
Biostimulant Application under Different Nitrogen Fertilization Levels: Assessment of Yield, Leaf Quality, and Nitrogen Metabolism of Tunnel-Grown Lettuce

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Growing Conditions, and Treatments
2.2. Yield and Morphological Parameter Measurements and Quality Analysis Samplings
2.3. Leaf Color, Pigment Content, and SPAD Index Measurement
2.4. Antioxidant Activities and Bioactive Compound Measurements
2.5. Leaf Nitrate Content and Nitrogen Use Efficiency
2.6. Statistical Analysis
3. Results
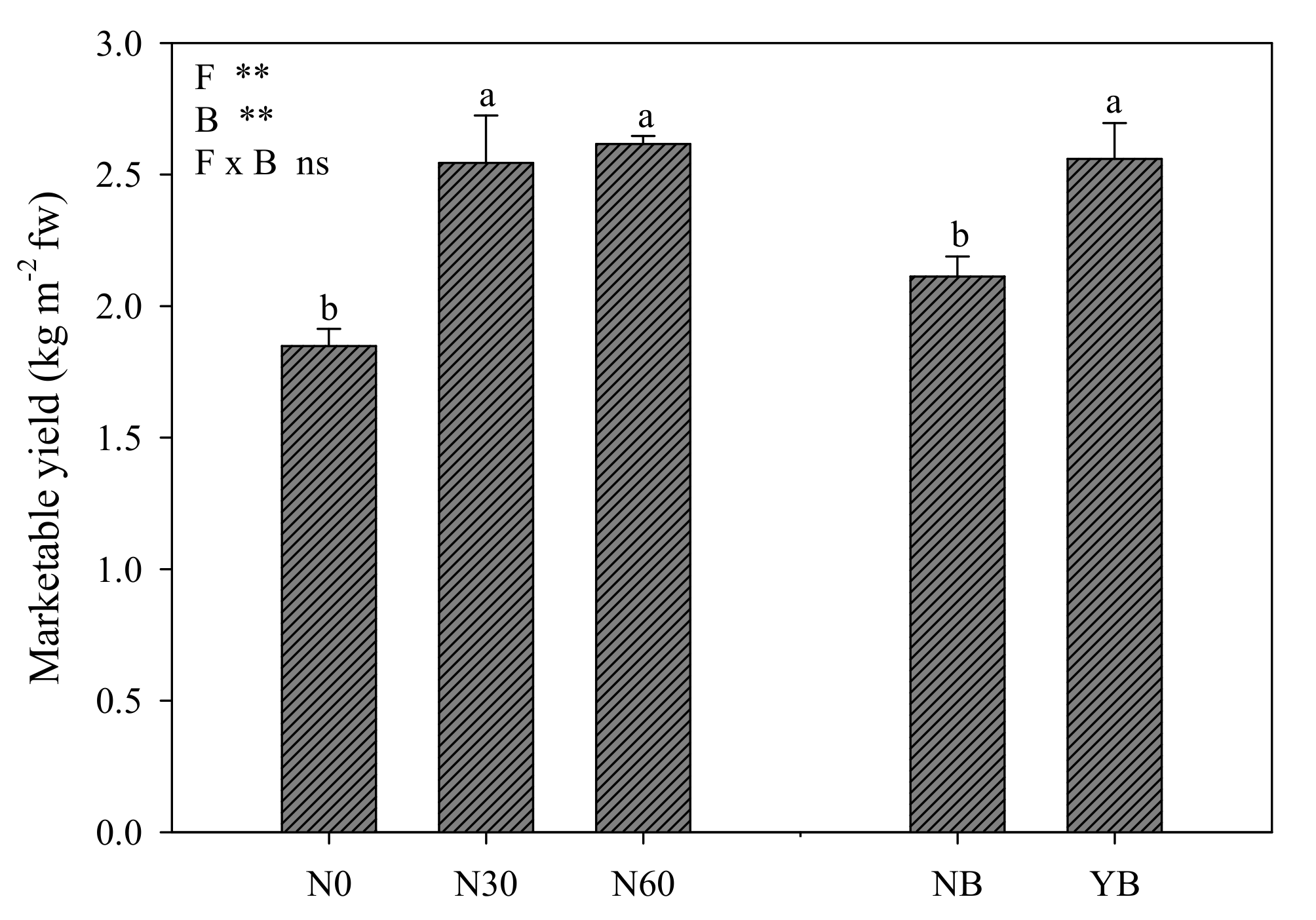
3.1. Marketable Yield and Morphological Parameters
3.2. Leaf Color, Pigment Content, and SPAD Index
3.3. Antioxidant Activities and Bioactive Compounds
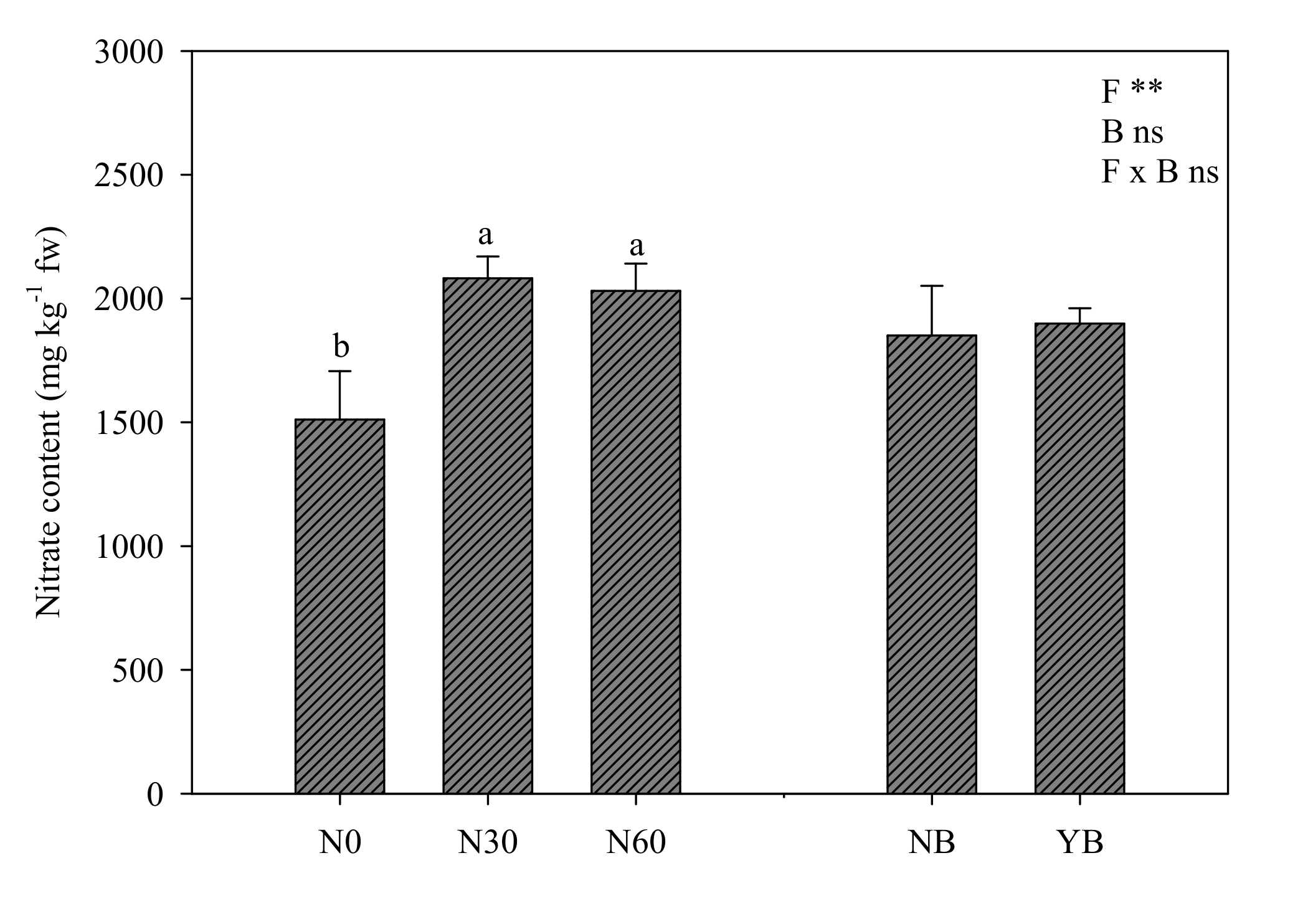
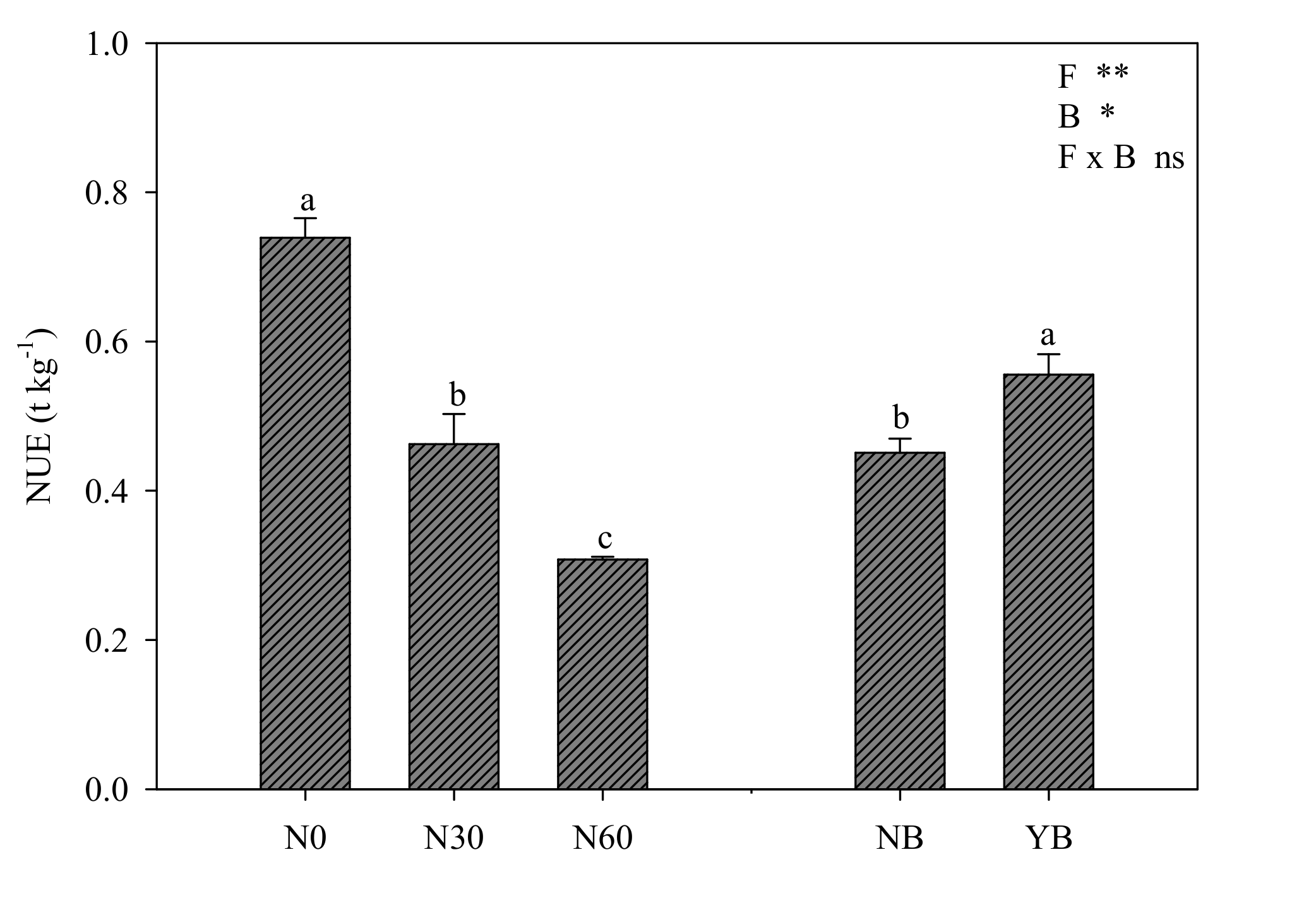
3.4. Nitrogen Metabolism: Leaf Nitrate Content and Nitrogen Use Efficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reimer, A.; Doll, J.E.; Basso, B.; Marquart-Pyatt, S.T.; Robertson, G.P.; Stuart, D.; Zhao, J. Moving toward sustainable farming systems: Insights from private and public sector dialogues on nitrogen management. J. Soil Water Conser. 2017, 72, 5A–9A. [Google Scholar] [CrossRef]
- Ronga, D.; Parisi, M.; Pentangelo, A.; Mori, M.; Di Mola, I. Effects of nitrogen management on biomass production and dry matter distribution of processing tomato cropped in southern Italy. Agronomy 2019, 9, 855. [Google Scholar] [CrossRef]
- Djidonou, D.; Zhao, X.; Simonne, E.H.; Koch, K.E.; Erickson, J.E. Yield, water-, and nitrogen-use efficiency in field-grown, grafted tomatoes. HortScience 2013, 48, 485–492. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Nocerino, S.; Rouphael, Y.; Colla, G.; Mori, M. Nitrogen use and uptake efficiency and crop performance of baby spinach (Spinacia oleracea L.) and Lamb’s Lettuce (Valerianellalocusta L.) grown under variable sub-optimal N regimes combined with plant-based biostimulant application. Agronomy 2020, 10, 278. [Google Scholar] [CrossRef]
- Lassaletta, L.; Billen, G.; Grizzetti, B.; Anglade, J.; Garnier, J. 50 year trends in nitrogen use efficiency of world cropping systems: The relationship between yield and nitrogen input to cropland. Environ. Res. Lett. 2014, 9, 105011. [Google Scholar] [CrossRef]
- Sutton, M.A.; Bleeker, A.; Howard, C.M.; Bekunda, M.; Grizzetti, B.; de Vries, W.; van Grinsven, H.J.M.; Abrol, Y.P.; Adhya, T.K.; Billen, G.; et al. Our Nutrient World: The challenge to produce more food and energy with less pollution. Cent. Ecol. Hydrol. 2013, 8, 95–108. [Google Scholar]
- Weitzberg, E.; Lundberg, J.O. Novel aspects of dietary nitrate and human health. Annu. Rev. Nutr. 2013, 33, 129–1598. [Google Scholar] [CrossRef] [PubMed]
- Tamme, T.; Reinik, M.; Roasto, M. Nitrates and nitrites in vegetables: Occurrence and health risks. Bioact. Foods Promot. Health 2010, 307–321. [Google Scholar] [CrossRef]
- Aires, A.; Carvalho, R.; Rosa, E.A.S.; Saavedra, M.J. Effects of agriculture production systems on nitrate and nitrite accumulation on baby-leaf salads. Food Sci. Nutr. 2013, 1, 3–7. [Google Scholar] [CrossRef]
- Rouphael, Y.; Spíchal, L.; Panzarová, K.; Casa, R.; Colla, G. High-throughput plant phenotyping for developing novel biostimulants: From lab to field or from field to lab? Front. Plant Sci. 2018, 9, 1197. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and physiological responses induced by protein hydrolysate based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Bonini, P.; Colla, G.; Rouphael, Y. Yield and nutritional quality of Vesuvian Piennolo tomato PDO as affected by farming system and biostimulant application. Agronomy 2019, 9, 505. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; Colla, G.; Mori, M. Effect of vegetal- and seaweed extract-based biostimulants on agronomical and leaf quality traits of plastic tunnel-grown baby lettuce under four regimes of nitrogen fertilization. Agronomy 2019, 9, 571. [Google Scholar] [CrossRef]
- Di Mola, I.; Ottaiano, L.; Cozzolino, E.; Senatore, M.; Giordano, M.; El-Nakhel, C.; Sacco, A.; Rouphael, Y.; Colla, G.; Mori, M. Plant-Based Biostimulants Influence the Agronomical, Physiological, and Qualitative Responses of Baby Rocket Leaves under Diverse Nitrogen Conditions. Plants 2019, 8, 522. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 64, 145–158. [Google Scholar] [CrossRef]
- Di Mola, I.; Conti, S.; Cozzolino, E.; Melchionna, G.; Ottaiano, L.; Testa, A.; Mori, M. Plant-Based Protein Hydrolysate Improves Salinity Tolerance in Hemp: Agronomical and Physiological Aspects. Agronomy 2021, 11, 342. [Google Scholar] [CrossRef]
- Rouphael, Y.; De Micco, V.; Arena, C.; Raimondi, G.; Colla, G.; De Pascale, S. Effect of Ecklonia maxima seaweed extract on yield, mineral composition, gas exchange and leaf anatomy of zucchini squash grown under saline conditions. J. Appl. Phycol. 2017, 29, 459–470. [Google Scholar] [CrossRef]
- Botta, A. Enhancing plant tolerance to temperature stress with amino acids: An approach to their mode of action. Acta Hortic. 2013, 1009, 29–35. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef]
- Cozzolino, E.; Giordano, M.; Fiorentino, N.; El-Nakhel, C.; Pannico, A.; Di Mola, I.; Rouphael, Y. Appraisal of biodegradable mulching films and vegetal-derived biostimulant application as eco-sustainable practices for enhancing lettuce crop performance and nutritive value. Agronomy 2020, 10, 427. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Stima Delle Superfici E Produzioni Delle Coltivazioni Agrarie, Floricole E Delle Piante Intere Da Vaso. Available online: http://dati.istat.it/Index.aspx?QueryId=33703# (accessed on 12 April 2021).
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health beneficts of lettuce (Lactuca sativa L.). J. Food Comp. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Baslam, M.; Idoia, G.; Nieves, G. The arbuscular mycorrhizal symbiosis can overcome reductions inyield and nutritional quality in greenhouse-lettuces cultivated at inappropriate growing seasons. Sci. Hortic. 2013, 164, 145–154. [Google Scholar] [CrossRef]
- Sofo, A.; Lundegårdh, B.; Mårtensson, A.; Manfra, M.; Pepe, G.; Sommella, E.; Scopa, A. Different agronomic and fertilization systems affect polyphenolic profile, antioxidant capacity and mineral composition of lettuce. Sci. Hort. 2016, 204, 106–115. [Google Scholar] [CrossRef]
- Perveen, S.; Iqbal, M.; Parveen, A.; Akram, M.S.; Shahbaz, M.; Akber, S.; Mehboob, A. Exogenous triacontanol-mediated increase in phenolics, proline, activity of nitrate reductase, and shoot k+ confers salt tolerance in maize (Zea mays L.). Braz. J. Bot. 2017, 40, 1–11. [Google Scholar] [CrossRef]
- Singh, M.; Khan, M.M.A.; Moinuddin, I.M.; Naeem, M. Augmentation of nutraceuticals, productivity and quality of ginger (Zingiber officinale Rosc.) through triacontanol application. Plant Biosyst. 2012, 146, 106–113. [Google Scholar] [CrossRef]
- Ries, S.K.; Richman, T.L.; Wert, V.F. Growth and yield of crops treated with triacontanol. J. Am. Soc. Hort. Sci. 1978, 103, 361–364. [Google Scholar]
- Naeem, M.; Khan, M.M.A.; Moinuddin, I.M.; Aftab, T. Triacontanol-mediated regulation of growth and other physiological attributes, active constituents and yield of Mentha arvensis L. Plant Growth Regul. 2011, 65, 195–206. [Google Scholar] [CrossRef]
- Grzegorczyk, I.; Bilichowski, I.; Mikicuik-Olasik, E.; Wysokinska, H. The effect of triacontanol on shoot multiplication and production of antioxidant compounds in shoot cultures of Salvia officinalis L. Acta Soc. Bot. Pol. 2006, 75, 11–15. [Google Scholar] [CrossRef][Green Version]
- Grzegorczyk, I.; Bilichowski, I.; Mikicuik-Olasik, E.; Wysokinska, H. In vitro cultures of Salvia officinalis L. as a source of antioxidant compounds. Acta Soc. Bot. Pol. 2005, 74, 17–21. [Google Scholar] [CrossRef]
- ILSA. Available online: https://www.ilsagroup.com/en/prodotti/prodotto/87/ilsastim.htm (accessed on 5 December 2019).
- Kumaravelu, G.; Livingstone, V.D.; Ramanujam, M.P. Triacontanol-induced changes in the growth, photosynthetic pigments, cell metabolites, flowering and yield of green gram. Biol. Plantarum 2000, 43, 287–290. [Google Scholar] [CrossRef]
- Chen, S.K.; Subler, S.; Edwards, C.A. Effects of agricultural biostimulants on soil microbial activity and nitrogen dynamics. Appl. Soil Ecol. 2002, 19, 249–259. [Google Scholar] [CrossRef]
- Islam, S.; Zaid, A.; Mohammad, F. Role of triacontanol in counteracting the ill effects of salinity in plants: A review. J. Plant Growth Regul. 2021, 40, 1–10. [Google Scholar] [CrossRef]
- Lichtenhaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leafextracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin–Ciocalteu Reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Kampfenkel, K.; Montagu, M.V.; Inzè, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Ann. Rev. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Kopta, T.; Pavlikova, M.; Sękara, A.; Pokluda, R.; Maršálek, B. Effect of bacterial-algal biostimulant on the yield and internal quality of lettuce (Lactuca sativa L.) produced for spring and summer crop. Not. Bot. Horti Agrobot. 2018, 46, 615–621. [Google Scholar] [CrossRef]
- Cristofano, F.; El-Nakhel, C.; Pannico, A.; Giordano, M.; Colla, G.; Rouphael, Y. Foliar and root applications of vegetal-derived protein hydrolysates differentially enhance the yield and qualitative attributes of two lettuce cultivars grown in floating system. Agronomy 2021, 11, 1194. [Google Scholar] [CrossRef]
- Khan, S.; Yu, H.; Li, Q.; Gao, Y.; Sallam, B.N.; Wang, H.; Jiang, W. Exogenous application of amino acids improves the growth and yield of lettuce by enhancing photosynthetic assimilation and nutrient availability. Agronomy 2019, 9, 266. [Google Scholar] [CrossRef]
- Kapitsimadi, C.; Gerasopoulos, D.; Olympios, C.; Passam, H. Effect of a long chain aliphatic alcohol (triacontanol) on growth and yield of different horticultural crops. Acta Hort. 1995, 379, 237–243. [Google Scholar] [CrossRef]
- Knowles, N.R.; Ries, S.K. Rapid growth and apparent total nitrogen increases in rice and corn plants following applications of triacontanol. Plant Physiol. 1981, 68, 1279–1284. [Google Scholar] [CrossRef]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of TCA cycle and N metabolism in Zea mays L. J. Agr. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef]
- European Community. Reg. n° 1258 of 2 December 2011. Off. J. Eur. Union. 2011, L 320, 15–17. [Google Scholar]
- Di Mola, I.; Rouphael, Y.; Colla, G.; Fagnano, M.; Paradiso, R.; Mori, M. Morphophysiological traits and nitrate content of greenhouse lettuce as affected by irrigation with saline water. HortScience 2017, 52, 1716–1721. [Google Scholar] [CrossRef]
- Di Mola, I.; Rouphael, Y.; Ottaiano, L.; Duri, L.G.; Mori, M.; De Pascale, S. Assessing the effects of salinity on yield, leaf gas exchange and nutritional quality of spring greenhouse lettuce. International Symposium on New Technologies for Environment Control, Energy-Saving and Crop Production in Greenhouse and Plant. Acta Hortic. 2018, 1227, 479–484. [Google Scholar] [CrossRef]
- Muthuchelian, K.; Murugan, C.; Harigovindan, R.; Nedunchezhian, N.; Kulandaivelu, G. Effect of triacontanol in flooded Erythrina variegata seedlings. 2. Changes in (CO2)-c-14 fixation, and ribulose-1, 5-b-isophosphate carboxylase, photosystem and nitrate reductase activities. Photosynthetica 1997, 30, 407–413. [Google Scholar]
- Tischner, R. Nitrate uptake and reduction in higher and lower plants. Plant Cell EnViron. 2000, 23, 1005–1024. [Google Scholar] [CrossRef]
- Hirel, B.; Bertin, P.; Quillere’, I.; Bourdoncle, W.; Attagnant, C.; Dellay, C.; Gouy, A.; Cadiou, S.; Retailliau, C.; Falque, M.; et al. Towards a better understanding of the genetic and physiological basis for nitrogen use efficiency in maize. Plant Physiol. 2001, 125, 1258–1270. [Google Scholar] [CrossRef]
- Coruzzi, G.; Bush, D.R. Nitrogen and C nutrient and metabolite signaling in plants. Plant Physiol. 2001, 125, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Mohammad, F. Triacontanol as a dynamic growth regulator for plants under diverse environmental conditions. Phy. Molecr. Biol. Plants 2020, 26, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Gruda, N. Impact of environmental factors on product quality of greenhouse vegetables for fresh consumption. Crc. Crit. Rev. Plant Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Dhalaria, R.; Verma, R.; Kumar, D.; Puri, S.; Tapwal, A.; Kumar, V.; Kuca, K. Bioactive Compounds of Edible Fruits with Their Anti-Aging Properties: A Comprehensive Review to Prolong Human Life. Antioxidants 2020, 9, 1123. [Google Scholar] [CrossRef]
- Li, J.; Zhu, Z.; Gerendás, J. Effects of nitrogen and sulfur on total phenolics and antioxidant activity in two genotypes of leaf mustard. J. Plant Nutr. 2008, 31, 1642–1655. [Google Scholar] [CrossRef]
- Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Woo, S.L. Appraisal of combined applications of Trichoderma virens and a biopolymer-based biostimulant on lettuce agronomical, physiological, and qualitative properties under variable N regimes. Agronomy 2020, 10, 196. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Fresh Weight | Head Diameter | Leaf Nb | Leaf DM |
|---|---|---|---|---|
| g head−1 | cm | n head−1 | % | |
| Fertilization | ||||
| N0 | 205.29 b | 24.25 b | 33.0 b | 4.83 |
| N30 | 282.69 a | 25.67 a | 44.5 a | 4.73 |
| N60 | 290.64 a | 25.50 a | 43.5 a | 4.90 |
| Biostimulants | ||||
| NB | 234.73 b | 24.56 b | 38.33 b | 4.61 b |
| YB | 284.36 a | 25.72 a | 42.33 a | 5.02 a |
| Significance | ||||
| Fertilization (F) | ** | ** | * | NS |
| Biostimulants (B) | ** | ** | ** | * |
| F × B | NS | NS | NS | NS |
| Treatments | SPAD Index | L* | a* | b* |
|---|---|---|---|---|
| Fertilization | ||||
| N0 | 33.77 c | 48.21 c | −10.30 | 29.03 |
| N30 | 38.54 b | 49.55 b | −10.02 | 28.60 |
| N60 | 39.54 a | 50.64 a | −10.02 | 27.92 |
| Biostimulants | ||||
| NB | 35.73 b | 47.85 b | −9.74 b | 27.52 b |
| YB | 38.83 a | 51.08 a | −10.48 a | 29.50 a |
| Significance | ||||
| Fertilization (F) | ** | ** | NS | NS |
| Biostimulants (B) | ** | ** | ** | * |
| F × B | NS | NS | NS | NS |
| Treatments | Chlorophyll a | Chlorophyll b | Total Chlorophylls | Carotenoids |
|---|---|---|---|---|
| mg g−1 fw | mg g−1 fw | mg g−1 fw | µg g−1 fw | |
| Fertilization | ||||
| N0 | 0.551 b | 0.213 | 0.764 b | 333 |
| N30 | 0.591 b | 0.222 | 0.813 b | 295 |
| N60 | 0.707 a | 0.254 | 0.960 a | 265 |
| Biostimulants | ||||
| NB | 0.635 | 0.237 | 0.871 | 299 |
| YB | 0.598 | 0.222 | 0.820 | 297 |
| Significance | ||||
| Fertilization (F) | * | NS | * | NS |
| Biostimulants (B) | NS | NS | NS | NS |
| F × B | NS | NS | NS | NS |
| Treatments | LAA | HAA | Total Phenols | TAA |
|---|---|---|---|---|
| mMTrolox eq. 100 g−1dw | mM AA eq. 100 g−1dw | mg Gallic Acid eq. g−1dw | mg g−1fw | |
| Fertilization | ||||
| N0 | 9.64 a | 5.96 a | 1.65 | 28.34 a |
| N30 | 8.60 ab | 5.65 a | 1.46 | 22.39 ab |
| N60 | 7.13 b | 5.12 b | 1.40 | 17.10 b |
| Biostimulants | ||||
| NB | 8.43 | 5.34 | 1.58 | 22.89 |
| YB | 8.49 | 5.81 | 1.42 | 22.34 |
| Significance | ||||
| Fertilization (F) | * | ** | NS | * |
| Biostimulants (B) | NS | NS | NS | NS |
| F × B | NS | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ottaiano, L.; Di Mola, I.; Cozzolino, E.; El-Nakhel, C.; Rouphael, Y.; Mori, M. Biostimulant Application under Different Nitrogen Fertilization Levels: Assessment of Yield, Leaf Quality, and Nitrogen Metabolism of Tunnel-Grown Lettuce. Agronomy 2021, 11, 1613. https://doi.org/10.3390/agronomy11081613
Ottaiano L, Di Mola I, Cozzolino E, El-Nakhel C, Rouphael Y, Mori M. Biostimulant Application under Different Nitrogen Fertilization Levels: Assessment of Yield, Leaf Quality, and Nitrogen Metabolism of Tunnel-Grown Lettuce. Agronomy. 2021; 11(8):1613. https://doi.org/10.3390/agronomy11081613
Chicago/Turabian StyleOttaiano, Lucia, Ida Di Mola, Eugenio Cozzolino, Christophe El-Nakhel, Youssef Rouphael, and Mauro Mori. 2021. "Biostimulant Application under Different Nitrogen Fertilization Levels: Assessment of Yield, Leaf Quality, and Nitrogen Metabolism of Tunnel-Grown Lettuce" Agronomy 11, no. 8: 1613. https://doi.org/10.3390/agronomy11081613
APA StyleOttaiano, L., Di Mola, I., Cozzolino, E., El-Nakhel, C., Rouphael, Y., & Mori, M. (2021). Biostimulant Application under Different Nitrogen Fertilization Levels: Assessment of Yield, Leaf Quality, and Nitrogen Metabolism of Tunnel-Grown Lettuce. Agronomy, 11(8), 1613. https://doi.org/10.3390/agronomy11081613