Abstract
Agricultural land abandonment is an emerging problem in European Union (EU), and about 11% of agricultural EU land is at high risk of abandonment in the coming 10 years. Land abandonment may have both positive and negative effects in ecosystems. Due to the potential for land abandonment to increase soil fertility, the study of vegetation succession effects on soil quality is of great importance. In this study, we investigated an abandoned vineyard where, after a period of 30 years, rows and alleys were characterized by two different forms of vegetation succession: natural recolonization by trees along the rows and by herbaceous vegetation in the alleys. No-tilled alleys covered by herbaceous vegetation of a neighboring conventionally cultivated vineyard were used as a comparison. Soil samples were chemically characterized (pH, extractable element, and available and total metals), and analyzed for the determination of carbon (C) and nitrogen (N) pools; hydrolytic and phenol oxidizing (PO) enzyme activities involved in C, N, and phosphorus (P) cycles; and the enzyme ratios. Results highlighted that natural recolonization by trees increased the organic C and N soil pools by 58% and 34%, respectively, compared to the natural recolonization by herbaceous vegetation. Moreover, natural recolonization by trees reduced β-glucosidase by 79%, urease by 100%, alkaline phosphastase by 98%, acid phosphatase specific hydrolytic activities by 50%, and catechol oxidase and laccase specific oxidative activities by 127% and 119%, respectively, compared to the renaturalization by herbaceous vegetation. In addition, the natural recolonization by trees reduced the C (βglu):C (PO) enzymes ratio by 16% compared to that of the conventional vineyard. Comparing the natural recolonization by herbaceous vegetation with that of the conventional vineyard revealed little significant difference (15% of the measured and calculated parameters); in particular, PO activities significantly decreased in the renaturalized vineyard with herbaceous vegetation by 49% (catechol oxidase) and 52% (laccase), and the C (βglu):C (PO) enzyme ratio showed a reduction (−11%) in the vineyard naturally recolonized by herbaceous vegetation compared to the conventional vineyard. This highlights that the type of vegetation succession that takes place after land abandonment may have a significant impact in terms of soil fertility and C accrual potential. These results help to focus attention on the practices used in agro-forestry that should be adopted in abandoned agro-ecosystems to increase their biodiversity, soil C stock, and soil quality, because these indicators are affected by the type of vegetative coverage.
1. Introduction
Land use changes often occur in agricultural ecosystems. In the Mediterranean basin, one of the most important of these is land abandonment [1]. In southern Europe, 24.5% of the lands under annual and permanent crops were abandoned between 1961 and 2011 [2], and about 11% of agricultural EU land is under high risk of abandonment in the coming 10 years [3]. Land abandonment by farmers can occur because of low productivity or unfavorable soil or climate conditions, rural depopulation, and declines in agriculture income [3]. During the course of land abandonment, many changes in vegetation, physical environmental conditions, and chemical soil properties take place [4]. Land abandonment can have positive effects in ecosystems, such as increased biodiversity, reduction in soil erosion, higher water retention capacity, and greater soil organic carbon (SOC) stock gain [2,5,6]. After land abandonment, colonization by natural vegetation begins, and soil properties gradually change [4,6,7]. Therefore, land abandonment also significantly impacts soil properties due to soil tillage suspension, vegetation colonization and succession, changes in soil physical environmental conditions, and soil quality parameters [4].
The main soil property studied following land abandonment is SOC content and its variations over time. Indeed, grassland colonization of abandoned lands induces a rapid SOC recovery [8,9] and the subsequent woody vegetation colonization of the grassland also triggers a net SOC gain [2]. Similarly, in a study across Europe, Baddeley et al. [10] found a SOC stock increase of 18 Mg ha−1 after land use change from cropland to grassland. Atallah et al. [11] found intermediate carbon (C) content values in abandoned lands between cultivated (olive orchards) and native woodlands in Lebanon, thus concluding that abandoned fields can also be an active C sink in sandy soils in the Mediterranean ecosystem. This general SOC increase that can be observed after land abandonment is mainly due to an increase in organic matter input and to a higher resistance in litter decomposition [6]. Other than SOC increase, in a study on vineyard abandonment and conversion to grassland over a period of 35 years, Novara et al. [12] found higher (+19%) microbial biomass C (MBC), and lower metabolic quotient (−50%) and basal respiration (−44%), in the grassland, thus indicating that vineyard abandonment induced a higher C substrate use efficiency. Other studies [4,13] focused on nitrogen (N) dynamics, showing a decrease in organic N mineralization after land abandonment. The resulting changes in N dynamics were associated with pH modification, soil organic matter variation, and microbial biomass and activity shift.
However, soil quality indicators, such as microbial and biochemical parameters (i.e., the enzymatic activities), have been less studied in the assessment of soil response to land abandonment [14,15], although their potential in describing soil changes was previously reported [16,17,18,19] and explained by their relatively rapid response to soil management variations. Indeed, soil microbial biomass has a key role in soil organic matter transformation and element cycling [20,21], and is involved in a wide range of metabolic processes that are mediated by several enzymatic activities [22]. Soil enzymatic activities are known to be sensitive and early indicators of changes in soil quality after land use change [16,22,23,24] because they play a crucial role in numerous important soil processes, such as organic matter decomposition, molecular N fixation, and C, N, and other major nutrients’ cycles [25,26,27]. Enzyme activities can also be used to calculate the soil enzyme ratios of C:N, C:phosphorous (P), and N:P acquisition activity, which are given by the natural logarithm of the enzyme activities involved in C, N, and P cycles, and are a measure of the enzymatic resources directed towards acquisition of organic P and N relative to C [23]. In the same manner, Sinsabaugh and Shah [28] proposed the utilization of the ratio between the natural logarithm of β-glucosidase activity and the natural logarithm of phenol oxidizing (PO) enzyme activity as an index of recalcitrant C abundance (C (βglu):C (PO) enzyme ratio), assuming an inverse relationship between SOC and the C (βglu):C (PO) enzyme ratio. Similarly, oxidative and hydrolytic enzyme activities can be combined (sum of the oxidative activities divided by the sum of the hydrolytic activities) in a C:N ratio [29,30] that expresses the relative enzyme investment in soil C and N mineralization.
In this study, we investigated an abandoned vineyard where, after a period of 30 years, in rows and alleys [31] a natural recolonization developed, with trees along the rows and herbaceous vegetation in the alleys, thus providing the opportunity to study the effects of abandonment on soil quality. Moreover, a neighboring conventionally managed vineyard with grassed alleys was taken as reference to assess if artificial grassing in a commercial vineyard may lead to comparable results to those of the abandoned vineyard. Specifically, we hypothesized that: (i) the different forms of natural recolonization, forest vs. herbaceous vegetation, of the abandoned vineyard induced differences in soil quality; and (ii) the natural recolonization of the abandoned vineyard increased the soil quality compared with that of a conventionally managed vineyard.
2. Materials and Methods
2.1. Site Description
The investigated site was the “Pantaleone Oasis” farm, located in Bagnacavallo (Ravenna, Northern Italy, 44°25′38.68″ N, 11°58′19.83″ E). This site was an agricultural farm traditionally cultivated until the 1980s, when it was abandoned and subsequently transformed into an area dedicated to ecological re-equilibrium (since 2006 it has been included in the “Sites of Community Importance”, in the EU-Natura 2000 network site and ecological rebalancing area, with Site Code: IT4070024). The farm has an area of about 7 ha and in the past was cultivated with vines and herbaceous crops (i.e., wheat, barley, alfalfa, and maize). The vineyard was cultivated in rows with a traditional technique known as “married vine” (see the scheme in Figure S1), according to which a fruit tree or woody plant (in this case maple) acted as support for one or two vines on the planted row [32]. A large alley (Figure 1), of approximately 20 m, was traditionally cultivated with herbaceous crops. As mentioned previously, in the past 30 years a process of abandonment took place during which no anthropic activity was undertaken, with the exception of interventions to avoid diffusion of non-indigenous plants (i.e., manual and/or mechanical eradication). At present, the farm is characterized by naturally recolonized rows of oaks (Quercus sp.) divided by alleys of herbaceous vegetation (Figure 1).

Figure 1.
The “Pantaleone” farm after re-equilibrium. Both pictures indicate the natural recolonization by trees in the rows and the natural recolonization by herbaceous vegetation in the alleys.
At the beginning of the abandonment (1988), the soil was classified as Calcaric Cambisol, mixed, superactive, and mesic, with a silty-loam texture, sub-alkaline pH (8.0), 16 g·kg−1 of SOC, 1.3 g·kg−1 of total nitrogen (TN), a C-to-N ratio of 12, 4.5 mg·kg−1 of available P, and 160 g·kg−1 of total carbonates.
Soil samples were collected from Pantaleone soil, both from the rows naturally recolonized by trees (natural recolonization by trees—NRT) and from the alleys naturally recolonized by herbaceous vegetation (natural recolonization by herbaceous vegetation —NRH), at a depth of 0–0.2 m. Composite soil samples (each composed of 6 core subsamples) were collected from 6 randomly chosen sites (each of about 0.4 ha), obtaining 3 composite samples from the rows and 3 from the alleys. Similarly, 3 composite soil samples (6 core subsamples for each sample) were taken from three randomly chosen grassed alleys (each of about 0.1 ha) of a neighboring farm, cultivated with vines in a conventional manner (vineyard grassed alleys—VGA). Soil samples for chemical analyses were dried, milled, and sieved at 2.0 mm, and those for microbiological and biochemical analysis were sieved at 4.0 mm and stored at +4 °C.
2.2. Soil Chemical Characterization
Soil pH was determined through International Standardized Methods (ISO 10390, 2005). Exchangeable cations were extracted with 1 M ammonium acetate at pH 7 and determined by inductively coupled plasma optical emission spectroscopy (ICP-OES, Spectro Arcos, Germany).
Available P (Olsen-P) was determined using the Olsen method [33] and expressed as mg·kg−1.
Total metal concentrations in soils were determined by ICP-OES after wet acid digestion. Briefly, an amount of 0.250 g of crushed soil was weighed into PTFE recipients, added to 6 mL of HCl 37% and 2 mL of HNO3 65%, and digested in a microwave oven (Milestone, Shelton, CT, USA). The digested suspension was filtered through Whatman no. 42 paper filters and brought to 20 mL with deionized water. Bioavailable metals were determined according to Lindsay et al. [34]. An amount of 25 g of soil was suspended in 50 mL of 0.005 M DTPA, 0.01 M CaCl2, and 0.1 M triethanolamine solution at pH 7.3. The suspension was then shaken in a horizontal shaker for 2 h at 60 rpm and filtered through Whatman no. 42 paper filters. The resulting solutions of both total and bioavailable trace metals were analyzed through OES-ICP for the determination of metals.
2.3. Soil Carbon and Nitrogen Pools
Total soil organic carbon (SOC) and TN were determined using a Flash 2000 elemental analyzer CHNS-O (Thermo-Fisher Scientific, Waltham, MA, USA). Soil dissolved organic C (DOC) and total dissolved N (TDN), and soil MBC and microbial N (MBN), were determined through the fumigation-extraction method [35].
The total extractable organic C (TEC), humic acid (HA), and fulvic acid (FA) were determined according to Ciavatta et al. [36], and were used for calculation of the humification rate, HR = (HA + FA)/SOC × 100; the degree of humification, DH = (HA + FA)/TEC × 100; and the humification index, HI = [TEC-(HA + FA)]/(HA + FA), or non-humified over humified C; as reported in Cavani et al. [37].
2.4. Soil Enzyme Activities
The main hydrolytic soil enzyme activities linked to C, N, and P cycles in soil were determined. β-Glucosidase (βglu) activity was determined by p-nitrophenol released after incubation of soil with p-nitrophenyl-β-D-glucoside for 1 h at 37 °C [38]. Urease activity (Ure) was determined according to Kandeler and Gerber [39] by ammonium released after incubation of soil with urea for 2 h at 37 °C. Alkaline and acid phosphomonoesterase (Alk PME, Ac PME) activities were estimated by determining PNP released after incubation of soil with p-nitrophenyl-phosphate at pH 11.0 and 6.0 respectively, for 1 h at 37 °C [40]. Protease activity (Prot) was determined by amino acids (tyrosine) released after incubation of soil with sodium caseinate for 2 h at 50 °C using Folin–Ciocalteu reagent [41].
Three soil oxidative enzyme activities linked to C and N cycles were determined. Dehydrogenase activity (Dehy) was determined by the 2-p-iodo-nitrophenyl formazan (INTF) produced from the reduction in 2-p-iodo-nitrophenyl-tetrazolium chloride, as described by von Mersi and Schinner [42]. Catechol oxidase activity (Cat) was determined using the catechol method as described by Perucci et al. [43], and laccase activity (Lac) was determined using the ABTS method, as described in Floch et al. [44]. Specific enzyme activities were also calculated by dividing each enzyme activity either by the SOC or the MBC content [16,45].
2.5. Soil Enzyme Ratios
The soil enzyme ratios (C:N, C:P, N:P, and C (βglu):C (PO)) proposed by Sinsabaugh et al. [23,28], and applied in other studies that address soil enzyme stoichiometry [46,47], were calculated (Equations (1)–(4)) using the natural logarithm of the measured hydrolytic and oxidative enzyme activities related to C, N, and P cycles:
where: ln = natural logarithm, βglu = β-Glucosidase activity, Prot = protease activity, Ure = urease activity, Alk PME = alkaline phosphomonoesterase activity, Ac PME = acid phosphomonoesterase activity, Cat = catechol oxidase activity, Lac = laccase activity, and PO = phenol oxidizing enzymes.
These ratios can be considered to be an expression of the enzyme nutrient acquisition activity, indicating, for example, if the enzymatic activity is mainly directed to the acquisition of organic N or organic C (i.e., the C:N enzyme ratio) [23,46,47]. The soil C (βglu):C (PO) enzyme ratio can indicate if the decomposition process developed by soil microorganisms, and thus soil enzymes, is mainly directed to the labile or the recalcitrant fraction of the soil organic matter [28]. Similarly, the relative C:N enzyme investment ratio was calculated according to Yin et al. [30] as the sum of two oxidative activities divided by the sum of two hydrolytic activities (Equation (5)); this indicates the enzyme investment in the mineralization of soil C and N.
2.6. Data Analysis
Means separation tests were performed using a pairwise t-test after assumption verification and applying the Bonferroni p-value adjustment. The significance of all statistical tests was assessed at α = 0.05. All statistics were performed using the R environment [48].
3. Results
3.1. Soil Chemical Characterization
Soil reaction (Table 1) remained sub-alkaline as at the beginning of the abandonment process and similar results were also recorded in the conventional vineyard.

Table 1.
Means ± standard error of soil reaction (pHH2O), exchangeable potassium (K.exc, mg·kg−1), exchangeable magnesium (Mg.exc, mg·kg−1), exchangeable sodium (Na.exc, mg·kg−1), and available phosphorus (Olsen-P, mg·kg−1). Asterisks indicate significant pairwise t-test comparisons (* p < 0.05, ** p < 0.01) between the theses considered: NRT = natural recolonization by trees, NRH = natural recolonization by herbaceous vegetation, VGA = vineyard grassed alleys.
Exchangeable metals were not affected by the location (row or alleys), excluding exchangeable Mg, which resulted higher in NRT than in NRH and VGA soils (Table 1), whereas VGA and NRHG showed no significant differences.
Available phosphorous (Olsen-P) was higher in NRT soil than in VGA, whereas no significant differences were observed among NRT and NRH, or among NRH and VGA (Table 1).
The concentrations of main total and bioavailable metals in soils are reported in Table 2. The results highlight that differences are particularly significant in the case of copper (Cu) and zinc (Zn). These two metals (both total and bioavailable) were found to be significantly higher in the NRT than in the NRH (+83% for Cu and +78% for Zn). The same trend was also observed for bioavailable manganese (Mn), with higher values in NRT than in NRH G, and significant differences were also found between NRT and VGA (Table 2).
Table 2.
Means ± standard error of total and bioavailable soil copper (Cu), manganese (Mn), and zinc (Zn) content expressed as mg·kgds−1. Asterisks indicate significant pairwise t-test comparisons (* p < 0.05, ** p < 0.01) between the theses considered: NRT = natural recolonization by trees, NRH = natural recolonization by herbaceous vegetation, VGA = vineyard grassed alleys.
3.2. Soil Carbon and Nitrogen Pools
All the C and N soil pools (Figure 2), and the derived ratios, showed the same trend, with higher values in the NRT soil than in the NRH and in the VGA. Significant results were not observed only in the case of the MBN and the MBC:MBN ratio (Figure 2).
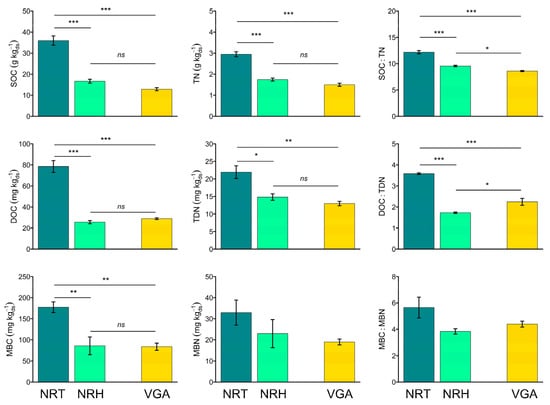
Figure 2.
Means of soil organic carbon (SOC), total nitrogen (TN), SOC:TN ratio (SOC:TN), dissolved organic carbon (DOC), total dissolved nitrogen (TDN), DOC:TDN ratio (DOC:TDN), microbial biomass carbon (MBC), microbial biomass nitrogen (MBN), and MBC:MBN ratio (MBC:MBN). Error bars represent the standard error. Asterisks indicate significant pairwise t-test comparisons (* p < 0.05, ** p < 0.01, *** p < 0.001) between the theses considered: NRT = natural recolonization by trees, NRH = natural recolonization by herbaceous vegetation, VGA = vineyard grassed alleys.
The C pools showed a mean increase of 58% and the N pools showed a mean increase of 34% in the NRT vs. NRH. Significant differences were also measured comparing the NRT with the VGA, with higher values of C (+60%) and N (+44%) pools in the NRT soil. Specifically, SOC was 54% and 64% higher in NRT compared to NRH and VGA, respectively; the DOC increased by 67% and 63% under NRT compared to NRH and VGA, respectively; MBC was 53% higher in NRT compared to both NRH and VGA. Regarding N pools, TN increased under NRT by 41% and 49% with respect to NRH and VGA, whereas TDN and MBN showed identical percentage increases in NRT compared to NRHG (31%) and VGA (42%). In general, C and N pools in the NRHG did not significantly differ from the VGA (Figure 2), and only in the case of the DOC:TDN ratio was NRH significantly lower than VGA, whereas the SOC:TN was found to be higher in NRG than in VGA. The same trend of C pools was found for the humified C content, which showed significantly higher values in NRT compared to NRH and VGA (Table 3).
Table 3.
Means ± standard error of humified C (g kgds−1), humification rate (HR, %), degree of humification (DH, %), and humification index (HI). Asterisks indicate significant pairwise t-test comparisons (* p < 0.05, ** p < 0.01) between the theses considered: NRT = natural recolonization by trees, NRH = natural recolonization by herbaceous vegetation, VGA = vineyard grassed alleys.
The humification ratio showed no significant differences between the three theses considered, whereas DH showed higher values in NRH compared to NRT with VGA, which did not show significant differences (Table 3). Conversely, an opposite trend was found for HI, which was found to be higher in the NRT than in the NRH, with both showing no significant differences compared to the VGA (Table 3).
3.3. Soil Enzyme Activities
Soil hydrolytic and oxidative enzyme activities showed slight or no differences between NRT and NRH locations (Table 4), whereas the majority (Ac PME, Prot, Dehy, Cat, and Lac) showed significant differences between NRT and VGA.
Table 4.
Means ± standard error of soil enzymatic activities: βglucosidase (βglu, mgNP kg−1 h−1), urease (Ure, mgN-NH4+ kg−1 h−1), alkaline phosphomonoesterase (Alk PME, mgNP kg−1 h−1), acid phosphomonoesterase (Ac PME, mgNP kg−1 h−1), protease (Prot, mgtyr kg−1 h−1), dehydrogenase (Dehy, mgINT kg−1 h−1), catechol oxidase (Cat, mmolcatechol kg−1 h−1), and laccase (Lac, mmolABTS+ kg−1 min−1). Asterisks indicate significant pairwise t-test comparisons (** p < 0.01, *** p < 0.001) between the theses considered: NRT = natural recolonization by trees, NRH = natural recolonization by herbaceous vegetation, VGA = vineyard grassed alleys.
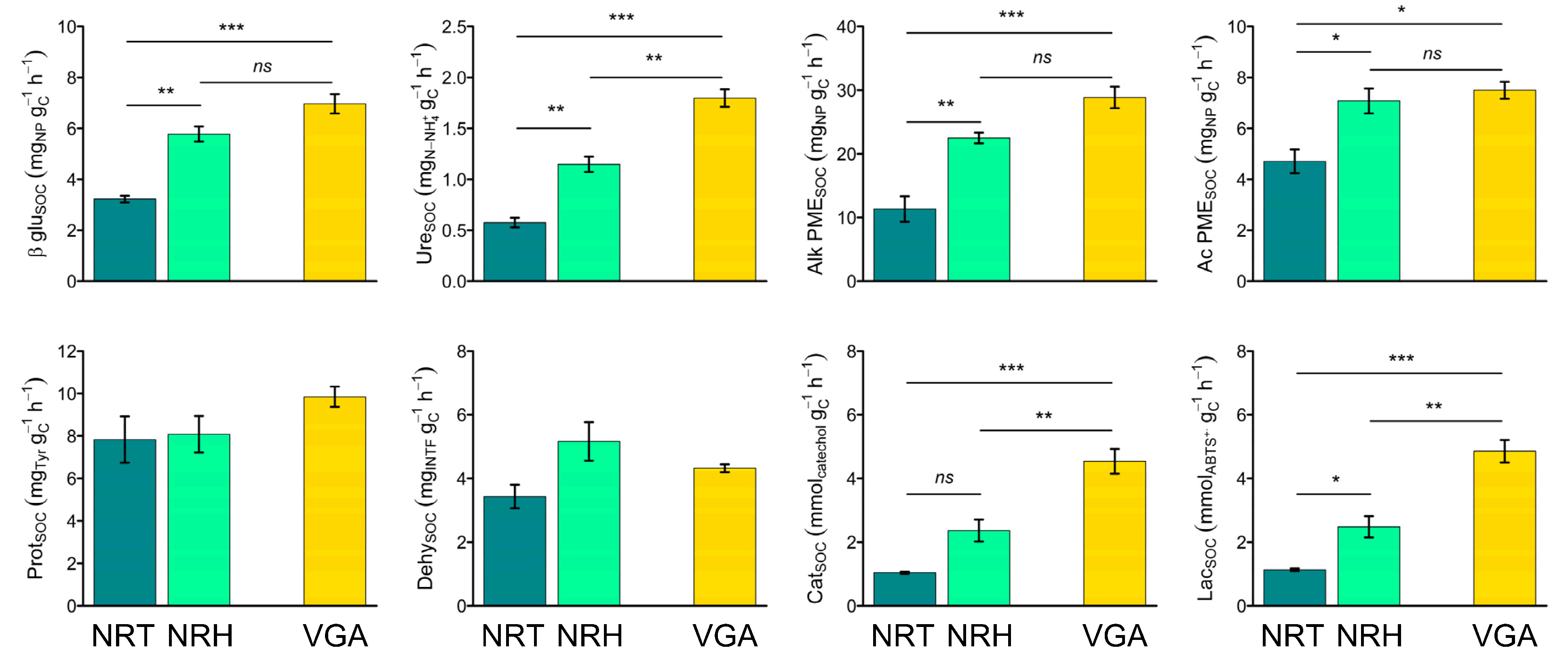
Specifically, Ac PME, Prot, and Dehy activities were found to be higher in NRT than in VGA, whereas Cat and Lac activities were found to be lower in NRT than in VGA and NRH than VGA (Table 4). The specific enzyme activities calculated on SOC content (Figure 3) showed the same trend with higher values in NRH than in NRT, with differences that were significant for βgluSOC, UreSOC, Alk PMESOC, Ac PMESOC, CatSOC, and LacSOC. Within these, only in the case of UreSOC, CatSOC, and LacSOC was the activity found to be higher in the VGA compared to both NRHG and NRT. By comparison, the other three activities (βgluSOC, Alk PMESOC, and Ac PMESOC) were higher in the VGA only compared to NRT. The specific enzyme activities determined on MBC content (Table S1) did not show significant differences, with the exception of CatMBC and LacMBC.
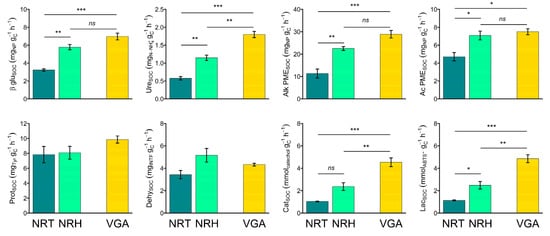
Figure 3.
Means of soil specific enzyme activities: βglucosidase (βgluSOC), urease (UreSOC), alkaline phosphomonoesterase (Alk PMESOC), acid phosphomonoesterase (Ac PMESOC), protease (ProtSOC), dehydrogenase (DehySOC), catechol oxidase (CatSOC), and laccase (LacSOC). Error bars represent the standard error. Asterisks indicate significant pairwise t-test comparisons (* p < 0.05, ** p < 0.01, *** p < 0.001) between the theses considered: NRT = natural recolonization by trees, NRH = natural recolonization by herbaceous vegetation, VGA = vineyard grassed alleys.
3.4. Soil Eco-Stoichiometric Ratios
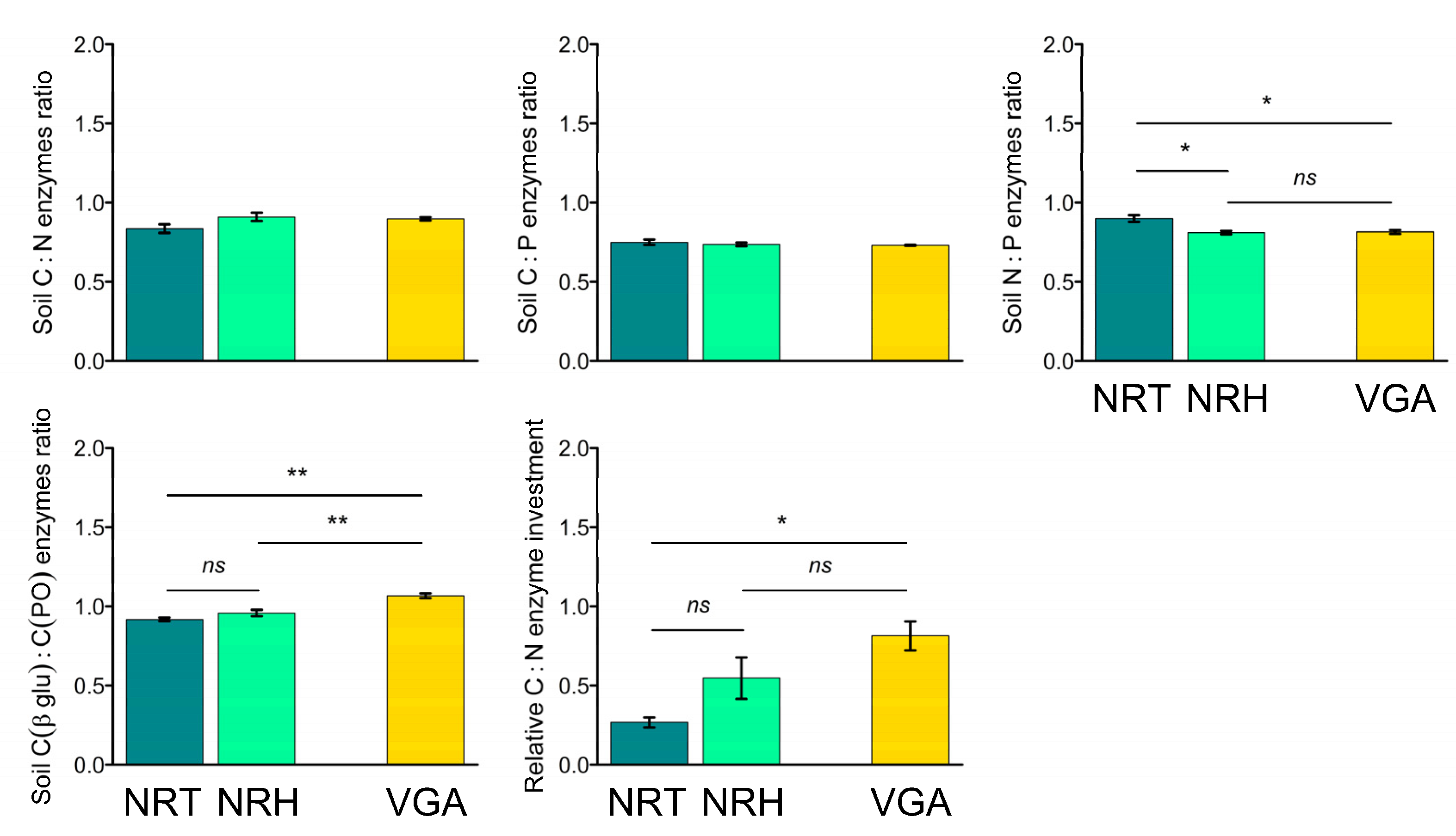
The C:N and the C:P soil enzyme ratios did not highlight significant differences between the three theses (Figure 4), whereas the C:N soil enzyme ratio showed higher values with the NRT in comparison to both NRH (+10%) and VGA (+9%). The opposite situation was observed for the C (βglu):C (PO) enzyme ratio (Figure 4), which showed higher values in the VGA (+14% and +10% than NRT and NRH, respectively) and no differences between NRT and NRH. Finally, the relative C:N enzyme investment highlighted significant differences only between NRT and VGA with VGA, which had values that were 67% higher.
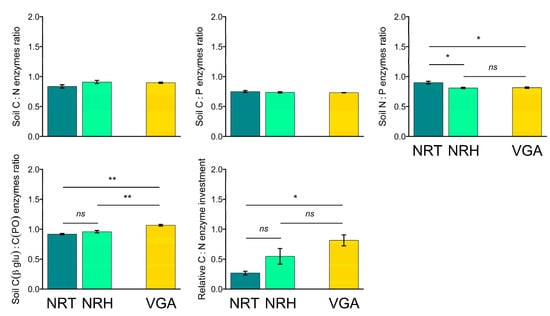
Figure 4.
Means of soil enzyme ratios (C:N, C:P, N:P, and C (βglu):C (PO)), and the C:N relative enzyme investment ratio. Error bars represent the standard error. Asterisks indicate significant pairwise t-test comparisons (* p < 0.05, ** p < 0.01) between the theses considered: NRT = natural recolonization by trees, NRH = natural recolonization by herbaceous vegetation, VGA = vineyard grassed alleys.
4. Discussion
4.1. Influences of Different Natural Recolonization on Soil Quality in the Abandoned Vineyard: Forest vs. Herbaceous Vegetation
The two different forms of vegetation succession developed in the rows and alleys of the Pantaleone abandoned farm induced significant differences for most of the measured parameters. The higher values of available P were registered in the NRT sites, in correlation with the highest SOC and the lowest pH. This evidence suggests that natural recolonization by trees more than by herbaceous vegetation can help in restoring soil fertility, with regard to P, which reached relatively more acceptable concentrations.
Total and bioavailable Cu were higher in NRT soils, perhaps as a consequence of previous cultivations; indeed, vines need Cu-based fungicides, which can accumulate in the soils and persist for many years after land abandonment [32]. The same explanation can be given for total Zn because this metal is present in several fungicides used in vineyards (i.e., mancozeb). Moreover, Mn and Zn availability was higher in NRT than in NRH, probably because of adsorption or retention phenomena caused by higher organic matter in NRT than in other soils [49].
All C and N pools showed higher values in NRT compared to NRH. Such differences were probably due to the increase in organic matter in these sites, similar to that described in Novara et al. [12]. The ecological re-equilibrium reached with trees (i.e., oak, maple, and hornbeam) clearly affected SOC content, evidencing values 2-fold higher in NRT soils than in NRH soils. The value of 36 g kg−1 reached as a consequence of re-equilibrium can be considered to be a high carbon content for a silty-loam soil in this region, as reported in Ungaro et al. [50]. Moreover, after 30 years of re-naturalization, it is likely close to the upper limit for SOC accumulation in this kind of “agro-ecosystem”. These findings (the increase in SOC by 54% in NRT) evidence the positive effect of natural recolonization by trees on SOC accumulation, confirming the results of previous studies on abandoned and re-naturalized olive orchards [11] or terraced vineyards [12]. The differences in the organic matter input to the soil between the two vegetation successions (trees vs. herbaceous vegetation) also impacted the humified C fraction, the degree of humification (DH), and the humification index (HI), which showed significant differences between NRT and NRH, with an opposite trend. The DH, that is the ratio between the humified C and the total extractable C, was higher in NRH, whereas humified C and HI, that is the ratio between the non-humified C and the humified C, were higher in NRT. In general, higher DH and lower HI values indicate higher soil ability to accumulate the SOC in the humic fraction [51]. The lower DH and higher HI values observed in NRT were due to the relatively higher non-humified C present in the soil extractable C fraction. These results appear to contradict other studies on afforestation or natural restoration, in which the accumulation of humified C was strictly correlated with the increase in SOC, and where the non-humified C fraction was lower [52,53,54]. It is likely that, in this “agro-ecosystem”, the input of organic C exceeds the humification capacity of the soil.
The increase in the MBC in NRT soils reflected the increase in SOC in the same system, confirming the positive effect of natural recolonization by trees on carbon content, as elsewhere described [6,12]. However, this did not impact the enzymatic activities, which were found to be similar in NRT and NRH. As already evidenced by Trasar-Cepeda et al. [45], in general, a strong decrease in organic C, as a result of soil use, influenced the enzyme activities involved in C, N, and P cycles (i.e., βglucosidase, urease, protease, invertase, and acid phosphomonoesterase), which showed lower values in correspondence of the lower organic C content. However, it is not possible to determine whether the observed modifications in the enzymatic activities were due to the lower content of organic C or to soil management (or, in this case, vegetation cover). In the aforementioned work [45], the determination of specific activity (calculated as the values of activity per unit of SOC) revealed that in soils affected by human activity, the soil specific enzyme activities were generally higher than those in abandoned soils. In this contest, where two forms of vegetation succession after land abandonment are compared, the higher specific enzyme activities were measured in the NRH soils. This leads to the supposition that, in the case of the NRT, the availability of substrates was higher, which induced the enlargement of the microbial biomass while maintaining, at the same time, an adequate microbial activity. As a consequence, natural recolonization by trees more than by herbaceous vegetation is able to favor good soil conditions for the microbial community, also leading to a higher C accrual potential capacity [51].
Considering C, N, and P availability in NRT and NRH soils, it was possible to observe that in NRT there was relatively higher available P than in NRH, and that in NRT the total organic C increased more than the total N. This leads to the supposition that, within these three nutrients, N may be the limiting one. This is confirmed by the N:P enzyme ratio, which was higher in NRT than in NRH, meaning that the microbial (enzymatic) activity was mainly focused on N rather than P [23].
4.2. Effects of Natural Recolonization and Conventional Management on Soil Quality in Two Neighboring Vineyards
The comparison of the conventional vineyard with the two forms of vegetation succession in the Pantaleone farm, focuses attention on the manner in which the results of sustainable management applied to a vineyard (similar to the agro-forestry perspective) may differ from those in an abandoned vineyard.
In general, the obtained results highlight a strong difference between NRT vs. VGA, which mainly derived from the different vegetation characterizing the two systems, as previously observed comparing NRT with NRH. Indeed, 30 years of land abandonment with natural recolonization by trees allowed the increase in soil C, N, and available elements, and Olsen-P concentration, which led to a decrease in the specific enzymatic activities because soil microorganisms did not need to recover a high quantity of nutrients from the soil. However, in NRT but not in VGA, it appears that total organic C content increased more than the total N, thus creating a stoichiometric imbalance that may have induced the expression of soil enzymes linked to N and P soil cycles (such as Prot and Ac PME), and the microbial activity (express as Dehy activity). Moreover, in NRT, a higher (+9%) N:P enzyme ratio and a lower (−205%) relative C:N enzyme investment ratio were measured compared to VGA, indicating that, in NRT, the enzyme activity was mainly dependent on N concentration [23,30]. It was observed [55,56] that lower availability of N than C would induce higher Lac activity, and that degrading the soil stabilized organic matter (“mining” activity) would recover the N needed. However, in the case of the current study, under NRT, we measured lower Lac (−53%) and Cat (−55%) activities compared to those of VGA, meaning that the lower N availability did not negatively affect the recalcitrant C fraction. This is in line with the calculated soil C (βglu):C (PO) enzyme ratio, which showed lower values (−16%) under NRT. Our results are consistent with those of Sinsabaugh and Shah [28], who reported that the soil C (βglu):C (PO) enzyme ratio is inversely related to recalcitrant C content and can be considered to be an index of recalcitrant C abundance.
Conversely to NRT, in NRH and VGA, all C and N pools were found to be similar. Previous studies, however, showed higher SOC and MBC content in the abandoned land compared to in a conventional vineyard [12,51]. In the case of this study, the absence of tillage in the alleys of both systems was likely the key factor explaining why no differences were observed [57,58].
The grassed alleys in the conventional vineyard and the herbaceous vegetated alleys in the abandoned vineyard were also differentiated in terms of the PO enzyme activity. Soil PO enzymes play an important role in the soil nutrient cycle as they can degrade lignin and humic substances, and can oxidize phenolic compounds releasing C and other nutrients [55,59]. Specifically, Cat and Lac (also when related to SOC) were found to be higher in the VGA soils (+35% vs. NRT and +34% vs. NRH for both Cat and Lac, and +77% vs. NRT and +48% vs. NRH for both Cat and Lac specific activities), thus indicating a higher activity in degrading the soil stabilized organic matter. Conversion of native ecosystems to agriculture typically leads to a loss of soil organic matter, in particular the labile fractions. In general, this increases soil PO activity (such as Cat and Lac), particularly on a specific basis [55], as occurred in the conventionally cultivated soils of our research. Moreover, in pineapple plantations in Tahiti, Waldrop et al. [60] reported that losses of about 50% in soil C and N, with respect to native tropical forest, resulted in a 10-fold increase in the PO activity per gram of organic matter. In addition, in our work, PO activities were lower when organic C concentration was higher, as in NRT, confirming the negative correlation of these activities with available organic C, N, and P content. This was confirmed by the soil C (βglu):C (PO) enzyme ratio, which was higher in VGA compared to NRH, evidencing an increased “mining” activity of the enzymes and a lower recalcitrant C content in VGA [28]. This highlights that, even if few differences occur between the grassed alleys in the conventional vineyard and the herbaceous vegetated alleys in the abandoned vineyard, in terms of nutrient concentration and microbial biomass, some microbial processes related to the nutrient acquisition activity (i.e., oxidative enzyme activity) are different and show a higher need in C and N recovery in the VGA compared to in the NRH.
Previous studies have highlighted the positive effect of cover crops or grassing in vineyards [57,61] compared to conventional systems characterized by tilled alleys. However, few studies were found comparing agriculture sustainable practices in vineyards with natural (or re-naturalized) systems [57].
5. Conclusions
The natural recolonization of an abandoned vineyard (by trees and by herbaceous vegetation) compared to a conventionally managed vineyard significantly impacted the soil quality, increasing the SOC (+64% with trees and +23% with herbaceous vegetation) and nutrient contents, soil microbial biomass, and activity (Dehy activity increased by 55% and 36% with trees and herbaceous vegetation, respectively), and reducing the soil C (βglu):C (PO) enzyme ratio (−16% with trees and −11% with herbaceous vegetation). In particular, the natural recolonization by trees induced a higher C accrual potential than recolonization by herbaceous vegetation, indicating that the secondary succession with trees may help to increase the SOC stock. The natural recolonization of the abandoned vineyard increased the soil quality compared with a conventionally managed vineyard, even if the alleys, naturally recolonized by herbaceous vegetation or conventionally covered by grasses, showed no statistically significant differences. Indeed, the two agricultural systems were found to be driven by different soil microbial acquisition activities: the abandoned vineyard showed higher soil hydrolytic enzymatic activities (i.e., protease, dehydrogenase, and acid phosphatase) and lower oxidative enzymatic activities. Conversely, the cultivated vineyard was characterized by higher oxidative enzymatic activities (i.e., catalase and laccase) and lower hydrolytic enzymatic activity. These differences in soil microbial acquisition activities reflect the differences in the soil organic C availability, which was higher in the abandoned vineyard and lower in the conventional vineyard, and perfectly match with the soil C:N and C (βglu):C (PO) enzyme ratio and the relative C:N enzyme investment. These findings suggest that natural recolonization may have a positive impact on soil quality, but may result in large differences according to vegetation type cover. Therefore, it is necessary to pay attention to the type of vegetation that is established, because natural recolonization by herbaceous vegetation can lead to a soil quality that is similar to that of a grassed alley that is not abandoned.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy11091841/s1, Figure S1: Scheme of vineyard traditional cultivation technique known as “married vine”, according to which a fruit or woody plant (maple) acted as a support for one or two vines on the planted row, Table S1: Means ± standard error of specific soil enzymatic activities: βglucosidase (βglu, mgNP mg MBC−1 h−1), urease (Ure, mgN-NH4+ mgMBC−1 h−1), alkaline phosphomonoesterase (Alk PME, mgNP mg MBC−1 h−1), acid phosphomonoesterase (Ac PME, mgNP mg MBC−1 h−1), protease (Prot, mgtyr mg MBC−1 h−1), dehydrogenase (Dehy, mgINT mg MBC−1 h−1), catechol oxidase (Cat, mmolcatechol mg MBC−1 h−1), and laccase (Lac, mmolABTS+ mg MBC−1 min−1). Asterisks indicate significant pairwise t-test comparisons (* p < 0.05, ** p < 0.01, *** p < 0.001) between the thesis considered: NRT = natural recolonization by trees, NRG = natural recolonization by grass, VGA = vineyard grassed alleys.
Author Contributions
Conceptualization, L.S. and C.M.; methodology, L.C.; formal analysis, L.S.; data curation, M.M.; writing—original draft preparation, L.S. and M.M.; writing—review and editing, M.M., L.C. and E.B.; supervision, C.C., M.T. and C.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available on request due to privacy.
Acknowledgments
The authors wish to thank Bruno Marangoni, Bagnacavallo municipality, the municipality union of “Bassa Romagna”, and Lestes (research and study association in natural sciences) for providing access to the Pantaleone protected area and soil sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nunes, A.N.; Coelho, C.O.A.; De Almeida, A.C.; Figueiredo, A. Soil erosion and hydrological response to land abandonment in a central inland area of Portugal. L. Degrad. Dev. 2010, 21, 260–273. [Google Scholar] [CrossRef]
- Gabarrón-Galeote, M.A.; Trigalet, S.; van Wesemael, B. Effect of land abandonment on soil organic carbon fractions along a Mediterranean precipitation gradient. Geoderma 2015, 249–250, 69–78. [Google Scholar] [CrossRef]
- Perpiña Castillo, C.; Kavalov, B.; Diogo, V.; Jacobs-Crisioni, C.; Batista e Silvia, F.; Lavalle, C. Agricultural land abandonment in the EU within 2015–2030. In JRC Working Papers (No. JRC113718); Joint Research Centre: Ispra, Italy, 2018; pp. 1–7. [Google Scholar]
- Raiesi, F. Land abandonment effect on N mineralization and microbial biomass N in a semi-arid calcareous soil from Iran. J. Arid. Environ. 2012, 76, 80–87. [Google Scholar] [CrossRef]
- Lasanta, T.; Nadal-Romero, E.; Arnáez, J. Managing abandoned farmland to control the impact of re-vegetation on the environment. The state of the art in Europe. Environ. Sci. Policy 2015, 52, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Gabarrón-Galeote, M.A.; Trigalet, S.; van Wesemael, B. Soil organic carbon evolution after land abandonment along a precipitation gradient in southern Spain. Agric. Ecosyst. Environ. 2015, 199, 114–123. [Google Scholar] [CrossRef]
- Bonet, A.; Pausas, J.G. Species richness and cover along a 60-year chronosequence in old-fields of southeastern Spain. Plant Ecol. 2004, 174, 257–270. [Google Scholar] [CrossRef]
- Deng, L.; Zhu, G.Y.; Tang, Z.S.; Shangguan, Z.P. Global patterns of the effects of land-use changes on soil carbon stocks. Glob. Ecol. Conserv. 2016, 5, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Szoboszlay, M.; Dohrmann, A.B.; Poeplau, C.; Don, A.; Tebbe, C.C. Impact of land-use change and soil organic carbon quality on microbial diversity in soils across Europe. FEMS Microbiol. Ecol. 2017, 93, 1–12. [Google Scholar] [CrossRef]
- Baddeley, J.A.; Edwards, A.C.; Watson, C.A. Changes in soil C and N stocks and C:N stoichiometry 21 years after land use change on an arable mineral topsoil. Geoderma 2017, 303, 19–26. [Google Scholar] [CrossRef]
- Atallah, T.; Sitt, K.; El Asmar, E.; Bitar, S.; Ibrahim, L.; Khatib, M.N.; Darwish, T. Effect of abandonment of olive orchards on soil organic carbon sequestration in Mediterranean Lebanon. Soil Res. 2015, 53, 745–752. [Google Scholar] [CrossRef]
- Novara, A.; La Mantia, T.; Rühl, J.; Badalucco, L.; Kuzyakov, Y.; Gristina, L.; Laudicina, V.A. Dynamics of soil organic carbon pools after agricultural abandonment. Geoderma 2014, 235–236, 191–198. [Google Scholar] [CrossRef]
- Zhang, K.; Cheng, X.; Dang, H.; Ye, C.; Zhang, Q. Soil nitrogen and denitrification potential as affected by land use and stand age following agricultural abandonment in a headwater catchment. Soil Use Manag. 2012, 28, 361–369. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Qiu, Y.; Chen, L. Analysis on soil nutrient characteristics for sustainable land use in Danangou catchment of the Loess Plateau, China. Catena 2003, 54, 17–29. [Google Scholar] [CrossRef]
- Gispert, M.; Emran, M.; Pardini, G.; Doni, S.; Ceccanti, B. The impact of land management and abandonment on soil enzymatic activity, glomalin content and aggregate stability. Geoderma 2013, 202–203, 51–61. [Google Scholar] [CrossRef]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Floch, C.; Capowiez, Y.; Criquet, S. Enzyme activities in apple orchard agroecosystems: How are they affected by management strategy and soil properties. Soil Biol. Biochem. 2009, 41, 61–68. [Google Scholar] [CrossRef]
- Mazzon, M.; Cavani, L.; Margon, A.; Sorrenti, G.; Ciavatta, C.; Marzadori, C. Changes in soil phenol oxidase activities due to long-term application of compost and mineral N in a walnut orchard. Geoderma 2018, 316, 70–77. [Google Scholar] [CrossRef]
- Giacometti, C.; Mazzon, M.; Cavani, L.; Triberti, L.; Baldoni, G.; Ciavatta, C.; Marzadori, C. Rotation and fertilization effects on soil quality and yields in a long term field experiment. Agronomy 2021, 11, 636. [Google Scholar] [CrossRef]
- Marinari, S.; Masciandaro, G.; Ceccanti, B.; Grego, S. Influence of organic and mineral fertilisers on soil biological and physical properties. Bioresour. Technol. 2000, 72, 9–17. [Google Scholar] [CrossRef]
- Masciandaro, G.; Ceccanti, B.; Gallardo-Lancho, J.F. Organic matter properties in cultivated versus set-aside arable soils. Agric. Ecosyst. Environ. 1998, 67, 267–274. [Google Scholar] [CrossRef]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. In Enzymes in the Environment: Activity, Ecology, and Applications; Burns, R.G., Dick, R.P., Eds.; CRC Press: New York, NY, USA, 2002; pp. 1–33. ISBN 0-8247-0614-5. [Google Scholar]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Nannipieri, P.; Trasar-cepeda, C.; Dick, R.P. Soil enzyme activity : A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Balota, E.L.; Kanashiro, M.; Filho, A.C.; Andrade, D.S.; Dick, R.P. Soil Enzyme Activities Under Long-Term Tillage and Crop Rotation Systems in Subtropical Agro-Ecosystems. Braz. J. Microbiol. 2004, 35, 300–306. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; Haddix, M.L.; Lee, D.D.; Conant, R.T.; Paul, E.A. A litter-slurry technique elucidates the key role of enzyme production and microbial dynamics in temperature sensitivity of organic matter decomposition. Soil Biol. Biochem. 2012, 47, 18–26. [Google Scholar] [CrossRef]
- Kwiatkowski, C.A.; Harasim, E.; Feledyn-Szewczyk, B.; Antonkiewicz, J. Enzymatic activity of loess soil in organic and conventional farming systems. Agriculture 2020, 10, 135. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Shah, J.J.F. Ecoenzymatic stoichiometry of recalcitrant organic matter decomposition: The growth rate hypothesis in reverse. Biogeochemistry 2011, 102, 31–43. [Google Scholar] [CrossRef]
- Cheeke, T.E.; Phillips, R.P.; Brzostek, E.R.; Rosling, A.; Bever, J.D.; Fransson, P. Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytol. 2017, 214, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Dijkstra, F.A.; Phillips, R.P.; Zhu, B.; Wang, P.; Cheng, W. Arbuscular mycorrhizal trees cause a higher carbon to nitrogen ratio of soil organic matter decomposition via rhizosphere priming than ectomycorrhizal trees. Soil Biol. Biochem. 2021, 157, 108246. [Google Scholar] [CrossRef]
- de Santiago-Martín, A.; Vaquero-Perea, C.; Valverde-Asenjo, I.; Quintana Nieto, J.R.; González-Huecas, C.; Lafuente, A.L.; Vázquez de la Cueva, A. Impact of vineyard abandonment and natural recolonization on metal content and availability in Mediterranean soils. Sci. Total Environ. 2016, 551–552, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Cavani, L.; Manici, L.M.; Caputo, F.; Peruzzi, E.; Ciavatta, C. Ecological restoration of a copper polluted vineyard: Long-term impact of farmland abandonment on soil bio-chemical properties and microbial communities. J. Environ. Manag. 2016, 182, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.R.; Cole, C.V.; Watandbe, F.; Dean, L. Estimation of Available Phosphorus in Soil by Extraction with sodium Bicarbonate. J. Chem. Inf. Model. 1954, 53, 1689–1699. [Google Scholar] [CrossRef]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA micronutrients soil test. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Eiol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Ciavatta, C.; Govi, M.; Antisari, L.V.; Sequi, P. Determination of organic carbon in aqueous extracts of soils and fertilizers. Commun. Soil Sci. Plant Anal. 1991, 22, 795–807. [Google Scholar] [CrossRef]
- Cavani, L.; Ciavatta, C.; Gessa, C. Identification of organic matter from peat, leonardite and lignite fertilisers using humification parameters and electrofocusing. Bioresour. Technol. 2003, 86, 45–52. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Phosphates in soils. Soil Biol. Biochem. 1977, 9, 167–172. [Google Scholar] [CrossRef]
- Ladd, J.N.; Butler, J.H.A. Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol. Biochem. 1972, 4, 19–30. [Google Scholar] [CrossRef]
- von Mersi, W.; Schinner, F. An improved and accurate method for determining the dehydrogenase activity of soils with iodonitrotetrazolium chloride. Biol. Fertil. Soils 1991, 11, 216–220. [Google Scholar] [CrossRef]
- Perucci, P.; Casucci, C.; Dumontet, S. An improved method to evaluate the o-diphenol oxidase activity of soil. Soil Biol. Biochem. 2000, 32, 1927–1933. [Google Scholar] [CrossRef]
- Floch, C.; Alarcon-Gutiérrez, E.; Criquet, S. ABTS assay of phenol oxidase activity in soil. J. Microbiol. Methods 2007, 71, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Liu, G.; Li, P.; Xue, S. Ecological stoichiometry of plant-soil-enzyme interactions drives secondary plant succession in the abandoned grasslands of Loess Plateau, China. Catena 2021, 202, 105302. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, X.; Green, S.M.; Dungait, J.A.J.; Wen, X.; Quine, T.A. Soil enzyme activity and stoichiometry along a gradient of vegetation restoration at the Karst Critical Zone Observatory in Southwest China. L. Degrad. Dev. 2019, 30, 1916–1927. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Lair, G.J.; Gerzabek, M.H.; Haberhauer, G. Sorption of heavy metals on organic and inorganic soil constituents. Environ. Chem. Lett. 2007, 5, 23–27. [Google Scholar] [CrossRef]
- Ungaro, F.; Staffilani, F.; Tarocco, P. Assessing and mapping topsoil organic carbon stock at regional scale: A scorpan kriging approach conditional on soil map delineations and land use. Land Degrad. Dev. 2010, 21, 565–581. [Google Scholar] [CrossRef]
- Francaviglia, R.; Renzi, G.; Ledda, L.; Benedetti, A. Organic carbon pools and soil biological fertility are affected by land use intensity in Mediterranean ecosystems of Sardinia, Italy. Sci. Total Environ. 2017, 599–600, 789–796. [Google Scholar] [CrossRef]
- Romeo, F.; Settineri, G.; Sidari, M.; Mallamaci, C.; Muscolo, A. Responses of soil quality indicators to innovative and traditional thinning in a beech (Fagus sylvatica L.) forest. For. Ecol. Manag. 2020, 465, 118106. [Google Scholar] [CrossRef]
- Saha, M.; Das, M.; Sarkar, A. Distinct nature of soil organic carbon pools and indices under nineteen years of rice based crop diversification switched over from uncultivated land in eastern plateau region of India. Soil Tillage Res. 2021, 207, 104856. [Google Scholar] [CrossRef]
- Ukalska-Jaruga, A.; Klimkowicz-Pawlas, A.; Smreczak, B. Characterization of organic matter fractions in the top layer of soils under different land uses in Central-Eastern Europe. Soil Use Manag. 2019, 35, 595–606. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L. Mosaic Patterns of thermal stress in the rocky intertidal zone: Implications for Climate Change. Ecol. Monogr. 2006, 76, 151–174. [Google Scholar] [CrossRef]
- Agnelli, A.; Bol, R.; Trumbore, S.E.; Dixon, L.; Cocco, S.; Corti, G. Carbon and nitrogen in soil and vine roots in harrowed and grass-covered vineyards. Agric. Ecosyst. Environ. 2014, 193, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Abad, J.; Hermoso De Mendoza, I.; Marín, D.; Orcaray, L.; Santesteban, L.G. Cover crops in viticulture. A systematic review (1): Implications on soil characteristics and biodiversity in vineyard. OENO One 2021, 55, 295–312. [Google Scholar] [CrossRef]
- Piotrowska-Długosz, A. Enzymes and soil fertility. In Enzymes in Agricultural Sciences; Gianfreda, L., Rao, M., Eds.; OMICS Group: Hyderabad, India, 2014. [Google Scholar]
- Waldrop, M.P.; Balser, T.C.; Firestone, M.K. Linking microbial community composition to function in a tropical soil. Soil Biol. Biochem. 2000, 32, 1837–1846. [Google Scholar] [CrossRef]
- Mania, E.; Alliani, N.; Giovannozzi, M.; Petrella, F.; Gangemi, L.; Guidoni, S. May vineyard floor management affects indicators of soil quality? Xe Congrès Int. des Terroirs Vitic. 2014, 2, 186–191. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).