Abstract
Canola is an important temperate oil crop that can be severely affected by drought. Understanding the physiological and molecular mechanisms involved in canola tolerance to water deficit is essential to obtain drought-tolerant productive cultivars. To investigate the role of antioxidant response and the possible involvement of calcium-dependent protein kinases (CDPKs) in canola tolerance to drought, we analyzed four genotypes with different sensitivity to water stress. Leaf relative water content, canopy temperature, PSII efficiency, electrolyte leakage index and lipid peroxidation were used as indicators to classify the cultivars as drought-tolerant or drought-sensitive. Antioxidant enzymes superoxide dismutase, guaiacol peroxidase and catalase displayed significantly higher activities in drought-tolerant than in drought-sensitive cultivars subjected to water deficit, suggesting that the efficiency of the antioxidant response is essential in canola drought tolerance. The increased expression of genes BnaCDPK6 and BnaCDPK14 under drought conditions, their differential expression in drought-tolerant and drought-sensitive genotypes, and the presence of multiple cis-acting stress-related elements in their promoter regions suggest that CDPKs are part of the signaling pathways that regulate drought response in canola. We propose the BnaCDPK genes and their regulator elements as potential molecular targets to obtain drought-tolerant productive canola cultivars through breeding or genetic transformation.
1. Introduction
Canola (Brassica napus L.) is one of the most widely grown oil crops worldwide. Canola oil has a high amount of polyunsaturated fatty acids, and it is extensively used in the food industry [1]. Although canola tolerant cultivars have been identified, as a temperate crop, canola is generally considered sensitive to drought, which has a substantial negative effect on this crop’s growth and development [2]. In general, abiotic stress adversely affects the major biological processes, resulting in alterations at the physiological, morphological, biochemical, molecular and cellular levels. Drought stress or scarcity of water is a critical environmental factor in agriculture worldwide. It negatively affects crop yield and, in the context of climate change, drought is expected to have a profound impact on sustainable crop production in arid and semi-arid soils. In canola, drought stress causes a decrease in plant biomass and induces changes in physiological and biochemical parameters, including decreases in relative water content, reduced chlorophyll content due to loss of pigments and damaged thylakoid membranes, reduced photosynthetic rate, stomatal conductance, cell expansion, growth, damage to cellular membranes and various genetic responses [3,4,5]. Photosynthesis is one of the key physiological processes that both drought and heat stress compromise by reducing photosynthetic efficiency and/or increasing the dissipation of excitation energy as heat, leading to alterations in fluorescence parameters. Under drought, plants reduce water loss by reducing the stomatal opening. However, this also restricts CO2 entry into the leaf, which may lead to the decrease of the photosynthesis rate [6,7,8]. Elevated CO2 has been reported to alleviate the negative effects of both heat stress and drought in wheat [9,10] Water stress induces the production of reactive oxygen species (ROS) in plant cells, which can act as signaling molecules, mediating many key physiological processes, but high ROS production is toxic to plants, and ROS can attack lipids, proteins, carbohydrates, nucleic acids and, ultimately, lead to cell death [11,12]. Plants have some mechanisms, such as physiological, biochemical and metabolic responses through the activation of many stress-responsive genes and the synthesis of proteins, to manage environmental stress [13]. Plant response mechanisms include enzymatic and non-enzymatic antioxidative systems, such as superoxide dismutase (SOD, EC 1.15.1.1), which is the first defense enzyme that converts superoxide to H2O2, which can be scavenged by catalase (CAT, EC 1.11.1.6) and different classes of peroxidases, including guaiacol peroxidase (GPOX, EC. 1.11.1.7) [14]. Several antioxidant and carbohydrate-metabolic enzymes have been reported to correlate with drought effects on wheat grain yield and have been proposed as bio-signatures for the breeding and selection of drought-tolerant genotypes [15]. The first step in abiotic stress response is sensing or perceiving stress, which is a complex process involving calcium signals and phosphorylation cascades that activate antioxidative systems [16]. One of the most common responses of stress perception is the increased accumulation of free calcium (Ca2+) in the cytosol leading to the altered expression of stress-responsive genes through Ca2+ sensors, such as calcium-dependent protein kinases (CDPKs/CPKs), calcineurin B-like proteins (CBLs), calmodulins (CaMs) and calmodulin-like proteins (CMLs) [17,18]. Among Ca2+ sensors, CDPKs play a key role in the response to abiotic and biotic stimuli [19]. They contain a single polypeptide chain, a calmodulin domain with four EF-hand Ca2+ binding motifs and a kinase domain; they perceive Ca2+ variations and quickly translate them into a phosphorylation signal [20]. Previous studies on CDPK genes in Arabidopsis have shown that they play a key role in response to abiotic and biotic stimuli and that they are specifically involved in drought resistance [21,22,23]. Canola and Arabidopsis belong to the same plant family (Brassicaceae) and, therefore, share some genetic similarities. At least 25 CDPK genes have been identified in the canola genome, some of which have been further analyzed under various abiotic stresses [24].
The aims of the present study were to investigate differential responses to water deficit in drought-sensitive and drought-tolerant canola genotypes. Leaf relative water content, leaf canopy temperature, PSII efficiency, leaf electrolyte leakage index and lipid peroxidation were measured to test whether they could constitute valid indicators of drought tolerance in canola. We hypothesized that the antioxidant response was likely to have an important role in canola drought tolerance, and we determined superoxide dismutase, guaiacol peroxidase and catalase enzymatic activities to check whether there were differences between sensitive and tolerant cultivars. Based on the reported role of CDPKs in Arabidopsis, we theorized they might be involved in canola response to drought. To test this hypothesis, we quantified the expression of three BnaCDPK genes in sensitive and tolerant canola cultivars subjected to water deficit, and we examined the BnaCDPK promoter regions to identify motifs that could indicate that their expression might be regulated in response to drought stress.
2. Materials and Methods
2.1. Plant Material, Experimental Design, and Drought Stress Treatments
Four cultivars of canola (Brassica napus L.), namely Zarfam and Slm046 (tolerant genotypes) and Okapi and Sarigol (sensitive genotypes), were used in this study [25,26]. A completely randomized factorial experiment with three replications was carried out in a growth chamber under controlled conditions (16 h/8 h day/night photoperiod, 22 °C). Seeds were surface-sterilized and grown in plastic pots filled with 5 kg clay loam soil for 3 weeks. Drought stress treatments were applied after 3 weeks. Irrigation regimes were based on pot weight and soil water content. Control plants were kept in good irrigation at 100% field capacity (FC), and two drought treatments, 70% FC and 40% FC, were applied for 3 more weeks. Leaves were then collected, frozen in liquid nitrogen and stored at −80 °C.
2.2. Relative Water Content, Leaf Temperature, Chlorophyll Fluorescence (Fv/Fm) and Electrolyte Leakage Index
Relative water content (RWC) is a physiological measure of cellular hydration. A completely developed young leaf from each plant was used. Fresh weight (FW) was measured; saturated weight (SW) was determined after keeping the leaves in water for 24 h at 4 °C in the dark, and dry weight (DW) was measured after 72 h in an oven at 45 °C. Then, RWC was estimated using the Weatherley formula [27]: RWC (%) = [(FW − DW)/(SW − DW)] × 100.
Leaf temperature (LT) is a determining factor in plant productivity. A portable infrared thermometer (Model OS1327D, OMEGA, Norwalk, CT, USA) was used to measure LT (°C). In order to avoid the effect of soil temperature, data were taken on high leaf coverage areas. Two to four measurements per pot were taken.
Chlorophyll fluorescence was measured with a portable fluorescence spectrometer (Handy PEA, Handsatech Instruments Ltd., King’s Lynn, UK) [28]. Leaf clips were applied for 30 min for dark adaptation, then minimal fluorescence (F0) and maximum fluorescence (Fm) were measured under a weak red light (<0.1 µmol m−2 s−1) and a 0.8 s saturating pulse (8000 µmol m−2 s−1). The variable fluorescence (Fv) and the maximum quantum yield of PSII (Fv/Fm) were calculated using the measured parameters F0 and Fm.
Leaf electrolyte leakage index (ELI) was used as a function of cellular injury from osmotic stress that was determined to be ELI (%) = (EC1/EC2) × 100 [29]. Plant material (0.8 g leaf) was immersed in sterile water in 20 mL tubes and incubated at 25 °C for 24 h; after incubation, the electrical conductivity of the solution (EC1) was measured; then, samples were autoclaved at 120 °C for 20 min, and the final electrical conductivity (EC2) was measured after cooling to 25 °C.
2.3. Lipid Peroxidation
The amount of malondialdehyde (MDA) is indicative of the level of damage to membrane lipid or lipid peroxidation. Leaf samples (0.05 g) were pulverized in liquid nitrogen, homogenized with 0.1% trichloroacetic acid (TCA) and centrifuged at 10,000× g for 15 min at 4 °C, then the supernatant (0.6 mL) was mixed with 0.6 mL of 20% (w/v) trichloroacetic acid (TCA) containing 0.5% (w/v) TBA; the mixture was heated at 95 °C for 30 min and then quickly cooled down in an ice bath. After centrifugation at 10,000× g for 10 min, the absorbance and nonspecific absorption of the supernatant was measured at 532 and 600 nm. The extinction coefficient (155 mM−1 cm−1) was used for calculating the amount of MDA as µmol MDA mg−1 fresh weight [30].
2.4. Enzyme Extraction, Protein Determination and Antioxidant Enzyme Activity
Leaf samples were ground by liquid nitrogen and extraction buffer containing 0.1 M Tris-HCl; 5% (w/v) sucrose and 0.1% 2-mercaptoethanol, at pH 7.5, was added (3:1 buffer volume:leaf FW). Then the homogenate was centrifuged (10,000× g) for 20 min at 4 °C, and the supernatant was used for enzymatic activities and protein determinations. The Bradford method [31] was used to determine the protein concentration.
Superoxide dismutase activity (SOD) was determined as the ability to inhibit the photochemical reduction of nitro blue tetrazolium [32]. The reaction mixture, 50 mM potassium phosphate buffer (pH 7.8), 13 mM methionine, 75 μM NBT, 0.1 μM EDTA, 4 μM riboflavin and the required amount of enzyme extract (total volume 1 mL) was illuminated with two 15 W fluorescent lamps for 15 min. A control sample contained the reaction mixture without the enzyme. One SOD unit represents enough enzyme to cause 50% inhibition in the reduction of NBT as observed at 560 nm. Guaiacol peroxidase (GPOX) activity is based on the ability to convert guaiacol to tetra guaiacol (ε = 26.6 mM−1 cm−1) as monitored at 436 nm [33]. The reaction mixture contained 100 mM potassium phosphate buffer (pH 7.0), 20 mM guaiacol, 10 mM H2O2 and enzyme extract (total volume 1 mL). Absorbance was measured at 436 nm for 5 min. One GPOX unit is defined as 1 μM oxidized guaiacol per min per mg protein (unit mg−1 protein). Catalase (CAT) activity assay is based on the disappearance of H2O2 at 240 nm (ε = 40 mM−1 cm−1) [34]. The reaction mixture contained 50 mM potassium phosphate buffer (pH 7.0), 33 mM H2O2 and enzyme extract (total volume 1 mL). Absorbance was measured at 240 nm for 180 s. CAT activity is expressed as units (µM H2O2 per min) per mg of protein.
2.5. RNA Isolation and Gene Expression
Total RNA was isolated and purified from canola leaves using the RNeasy plant mini kit (Qiagen, Valencia, CA, USA). The quality of the RNA samples was assessed in an agarose gel 2%, and its concentration was determined with a NanoDrop ND-1000 (Thermo Fisher Scientific, Waltham, MA, USA). cDNA was synthesized from RNA (1 µg) using the QuantiTect Reverse Transcription Synthesis kit (Qiagen. Valencia, CA, USA). Real-time PCR (RT-PCR) was performed using the fluorescent intercalating dye SYBRGreen with the Step One Plus Real-Time PCR System (Applied Biosystems, Waltham, MA, USA), according to the manufacturer’s instructions. Gene-specific primers were designed using the Primer3 Software (http://primer3.ut.ee/, accessed on 20 June 2019) (Table 1). The cycle threshold (Ct) was measured for three biological samples with three technical replicates per sample. For normalizing the level of target gene expression, BnaActin7 was used as a control constitutive gene. Analysis of the relative gene expression was carried out using the 2−ΔΔCt formula [35].

Table 1.
Primers used in this study.
2.6. Statistical Analyses
Data were analyzed using SAS (v. 9.4) by two-way ANOVA, considering drought and genotype as fixed factors, and the significance of the fixed factors and their interaction was evaluated for all data (Supplementary Table S1). Post-hoc comparisons of means were performed using the Duncan’s multiple range test (DMRT), p = 0.05.
3. Results
3.1. Effect of Drought Stress on Physiological Traits
The leaf relative leaf water content (RWC), leaf temperature (LT) and maximum quantum yield of PSII (Fv/Fm) were measured in the leaves of canola drought-tolerant cultivars (Zarfam and Slm046) and drought-sensitive cultivars (Okapi and Serigold) under control and drought-stress conditions. There were no significant differences among cultivars under well-watered conditions for any of the measured parameters.
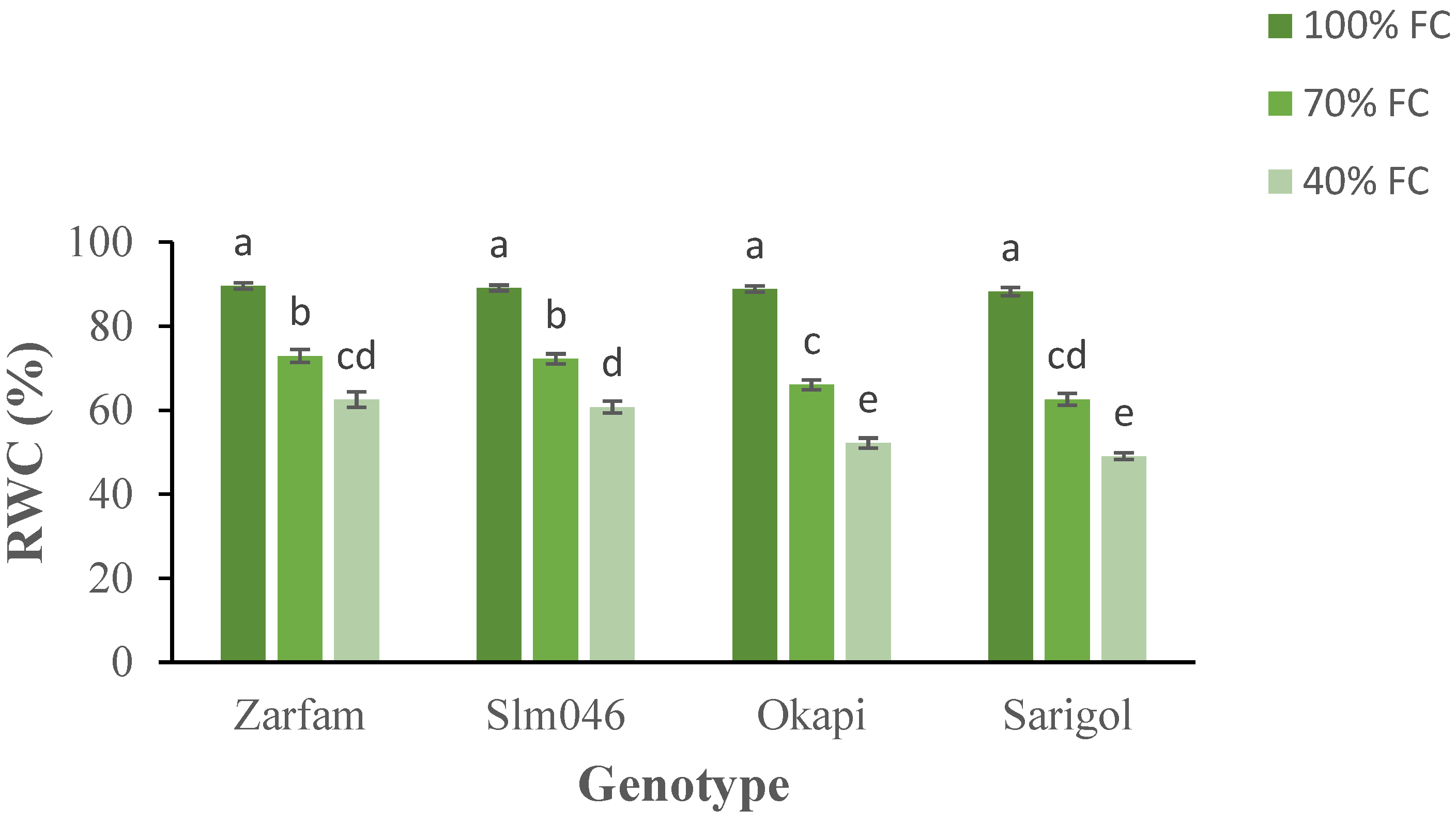
The relative leaf water content decreased significantly as a result of the drought treatments. Both moderate (70% FC) and severe drought (40% FC) significantly reduced RWC in comparison to well-watered (100% FC) conditions. Moreover, interaction between stress and genotypes was significant, and drought-sensitive genotypes were significantly more affected than tolerant genotypes under both drought regimes (Figure 1).
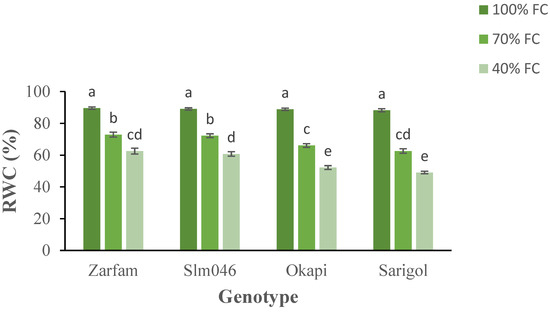
Figure 1.
Interaction effect of drought stress and genotype on the relative water content (RWC) of canola leaves. Data are means (n = 3) ± SE. Different letters indicate differences (p < 0.05) among treatments according to the Duncan test.
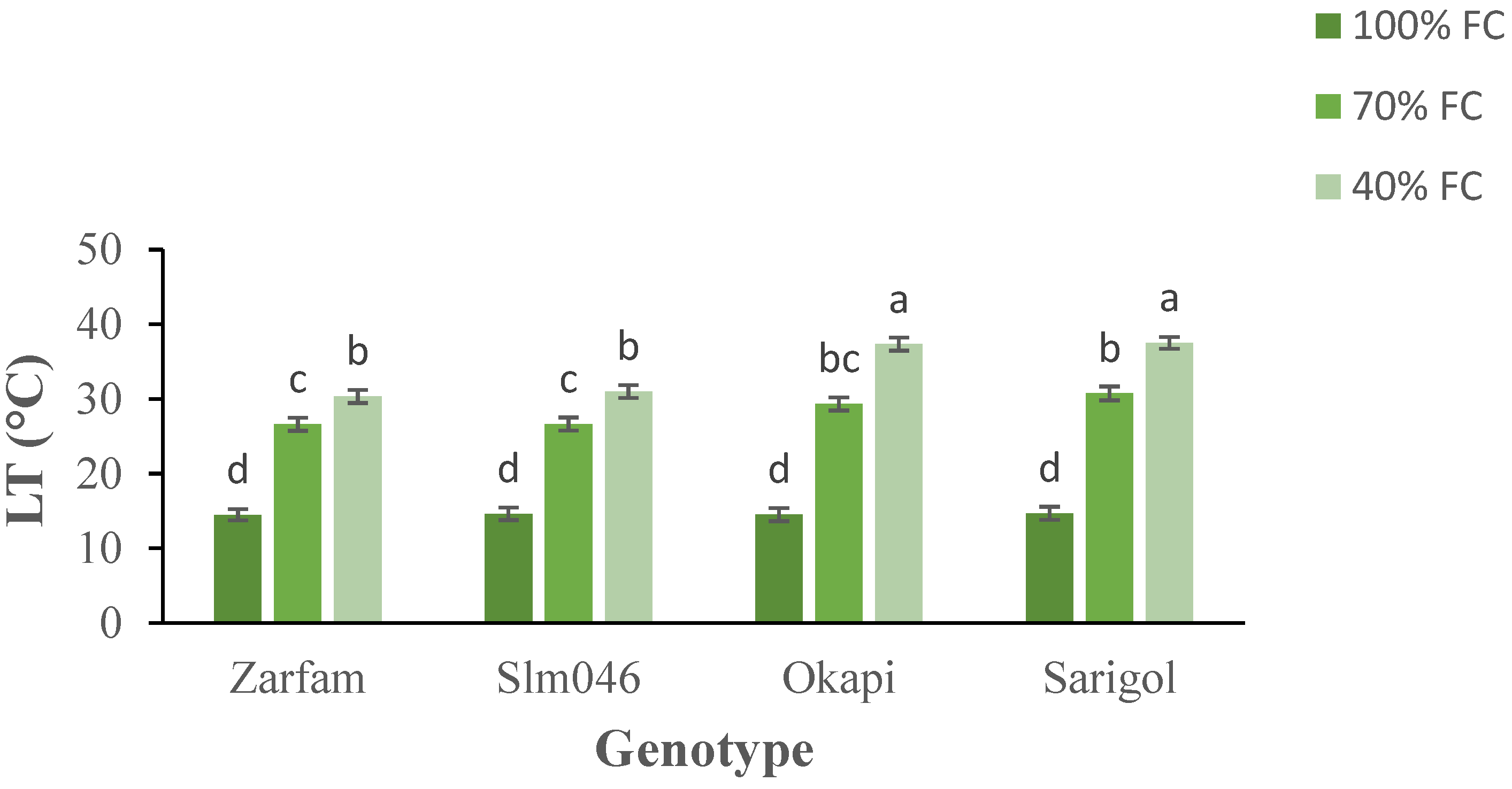
Leaf temperature increased with increasing drought. Interaction between stress and genotypes was significant, and the increase in LT was significantly lower in drought-tolerant genotypes than in drought-sensitive genotypes under the severe drought treatment (Figure 2).
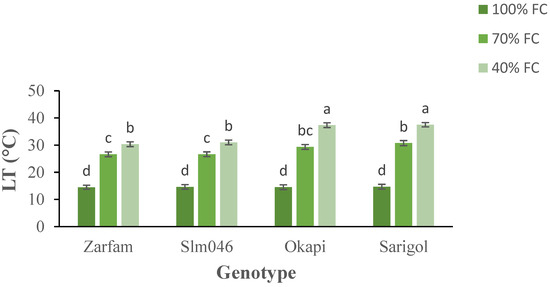
Figure 2.
Interaction effect of drought stress and genotype on the leaf temperature (LT) of canola plants. Data are means (n = 3) ± SE. Different letters indicate differences (p < 0.05) among treatments according to the Duncan test.
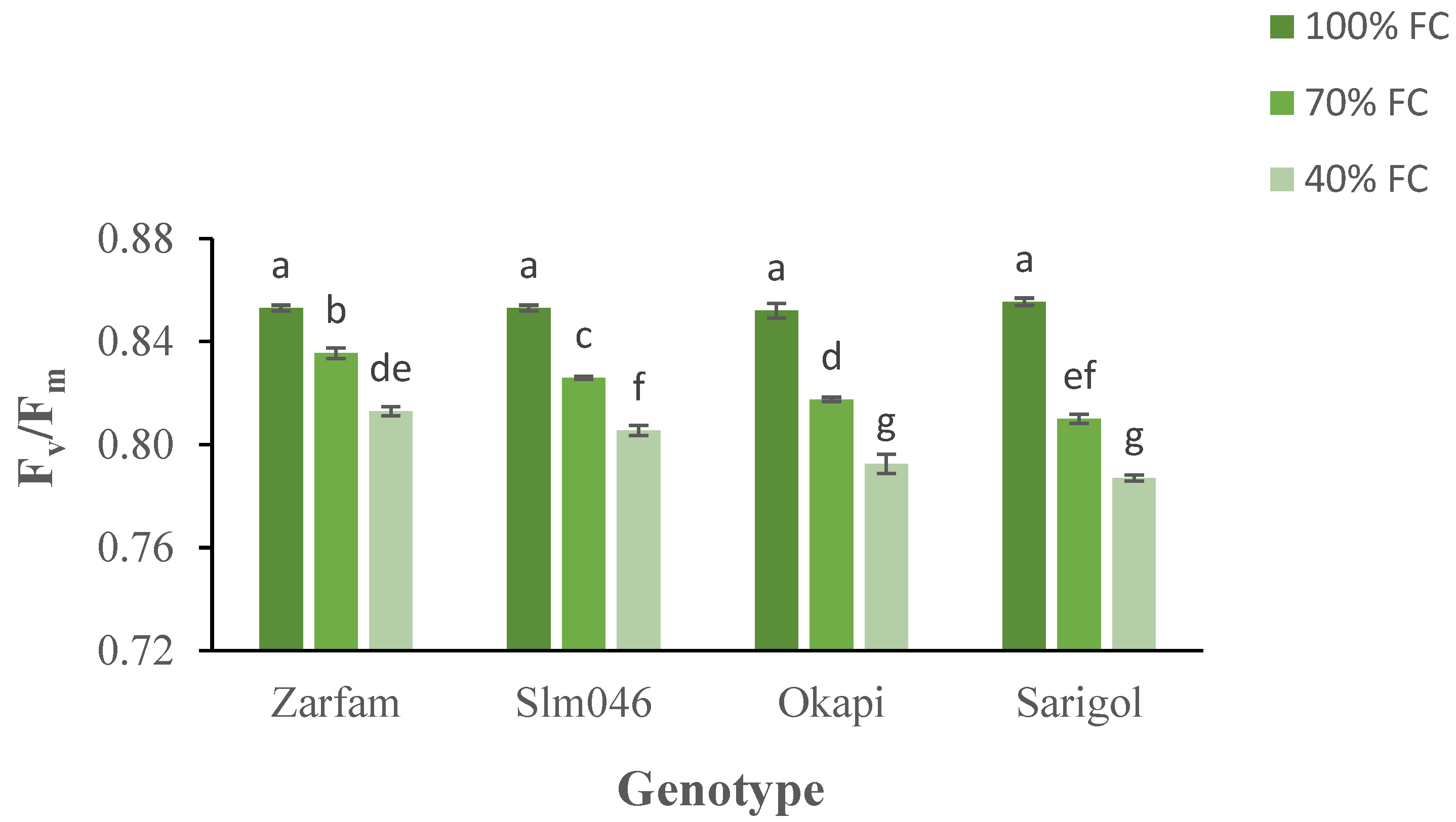
The maximum quantum yield of PSII (Fv/Fm) decreased under drought conditions, and differential responses between drought-tolerant and drought-sensitive genotypes were measured in both stress treatments. Interaction between stress and genotypes was significant. As compared to other genotypes, under severe drought, Zarfam maintained a significantly higher Fv/Fm value. (Figure 3).
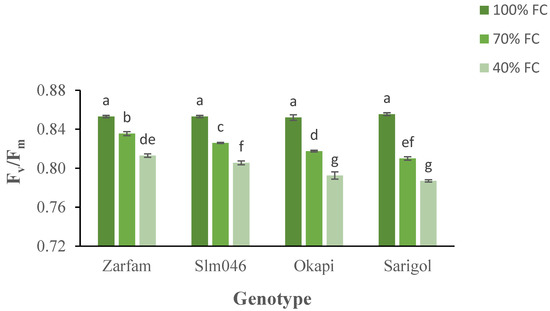
Figure 3.
Interaction effect of drought stress and genotype on the maximum quantum yield (Fv/Fm) of canola leaves. Data are means (n = 3) ± SE. Different letters indicate differences (p < 0.05) among treatments according to the Duncan test.
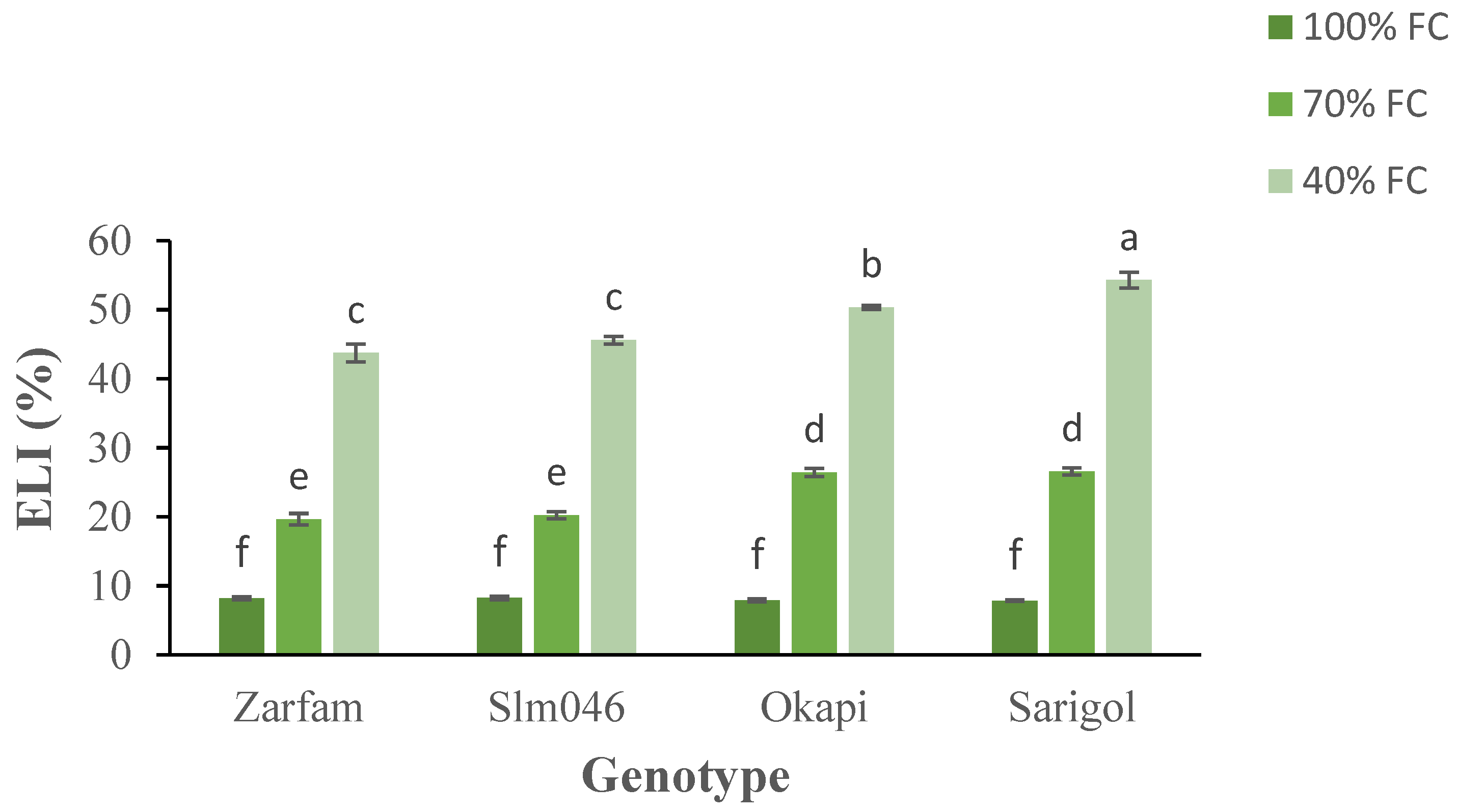
Analysis of electrolyte leakage index (ELI) showed increases for all genotypes under both drought treatments, and there were significant differences between tolerant and sensitive genotypes under both drought regimes. Interaction between factors was significant. Under severe drought, between the two sensitive cultivars, Sarigol was more affected than Okapi (Figure 4).
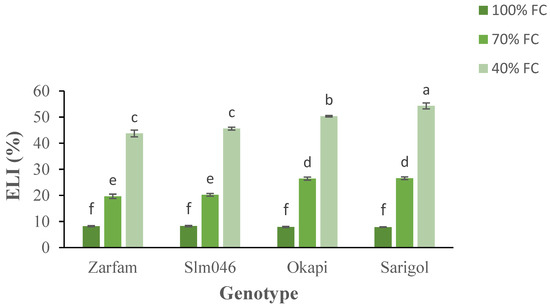
Figure 4.
Interaction effect of drought stress and genotype on the electrolyte leakage index (ELI) of canola leaves. Data are means (n = 3) ± SE. Different letters indicate differences (p < 0.05) among treatments according to the Duncan test.
3.2. Lipid Peroxidation
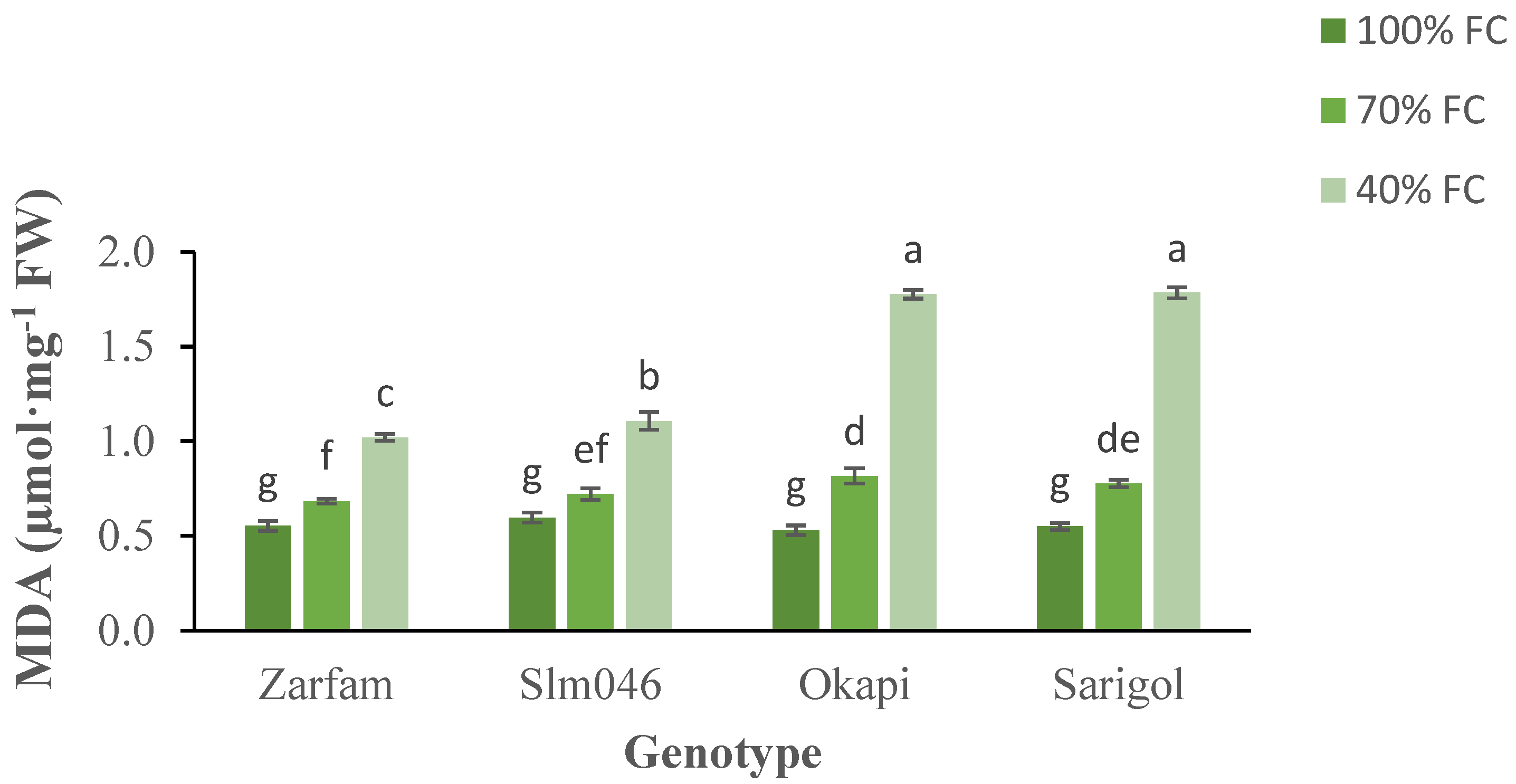
The malondialdehyde (MDA) content is indicative of lipid peroxidation. MDA content increased with drought for all genotypes. There were significant differences between tolerant and sensitive genotypes under severe drought. (Figure 5). Interaction between stress and genotypes was significant. As compared to other genotypes, under severe drought, Zarfam maintained a significantly lower MDA content, and this cultivar was less affected than Slm046, which is also drought-tolerant (Figure 5).
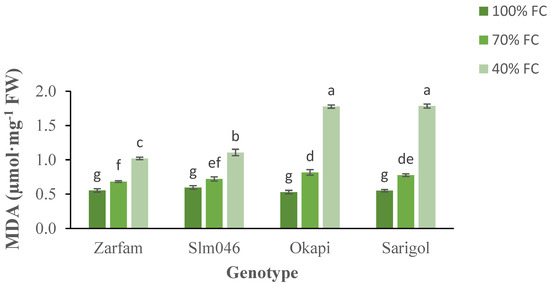
Figure 5.
Interaction effect of drought stress and genotype on the malondialdehyde (MDA) content of canola leaves. Data are means (n = 3) ± SE. Different letters indicate differences (p < 0.05) among treatments according to the Duncan test.
3.3. Antioxidant Enzyme Activities
There were no significant differences in superoxide dismutase (SOD) activity between drought-tolerant and drought-sensitive genotypes under control conditions. SOD activity significantly increased in all cultivars when moderate drought (70% FC) was applied, but still no significant differences among cultivars were detected (Figure 6A). Under more severe drought conditions (40% FC), SOD activity increased in the tolerant cultivars and decreased in the sensitive ones with significant differences between tolerant and sensitive genotypes. The same behavior was observed for guaiacol peroxidase (GPOX) (Figure 6B). There were no significant differences in catalase (CAT) activity among cultivars under control conditions. CAT activity increased significantly with drought severity in the tolerant cultivars, while it decreased in the sensitive ones (Figure 6C). Differences between tolerant and sensitive cultivars were significant under both drought regimes. Interaction between factors genotype and stress was significant for all three enzymatic activities.
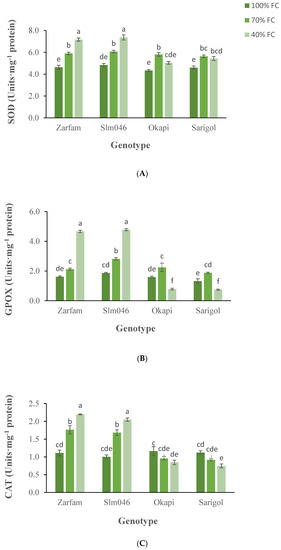
Figure 6.
Interaction effect of drought stress and genotype on SOD (A), GPOX (B) and CAT (C) enzymatic activities in canola leaves. Data are means (n = 3) ± SE. Different letters indicate differences (p < 0.05) among treatments according to the Duncan test.
3.4. Calcium-Dependent Protein Kinases (CDPK) Gene Expression in Canola Leaves and Promoter Analysis
There were significant differences in relative gene expression due to genotype and drought regime for the genes BnaCDPK6 and BnaCDPK14. Significant differences in BnaCDPK10 expression were a result of the drought treatment only (Supplementary Table S1). BnaCDPK6 and BnaCDPK14 expression increased with drought severity in all cultivars (Figure 7A,B). BnaCDPK10 expression increased when mild drought was applied and slightly decreased under the more severe conditions (Figure 7C).
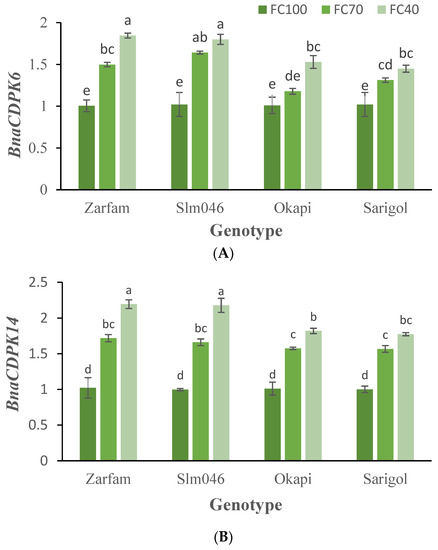
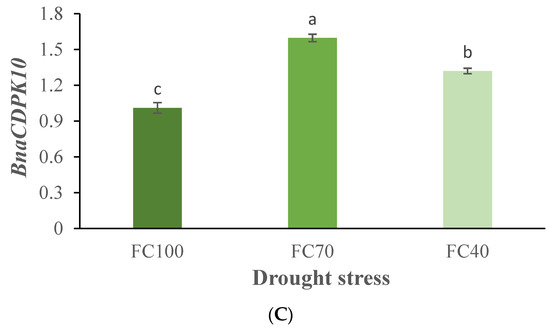
Figure 7.
Effects of drought stress and genotype on BnaCDPK6 (A), BnaCDPK14 (B) and BnaCDPK10 (C) transcript accumulation in canola leaves. Data are means (n = 3) ± SE. Different letters indicate differences (p < 0.05) among treatments according to the Duncan test.
A detailed analysis using the PLACE database [36] of BnaCDPK6, BnaCDPK10 and BnaCDPK14 promoter sequences, lying 2000 bp upstream of their coding sequences, identified several stress-responsive regulatory elements that are known to contribute to the expression of stress-related genes at a transcriptional level (Table 2).
Table 2.
Stress-responsive cis elements in the 2000 bp upstream region of BnaCDPK6, BnaCDPK10 and BnaCDPK14 coding sequences.
4. Discussion
This research was carried out to gain deeper insights into the physiological, biochemical and molecular response mechanisms induced by different levels of drought in canola plants. Several physiological and biochemical traits were scored under control and drought conditions to compare the drought response between tolerant and sensitive canola genotypes. The decrease in RWC is considered the main factor causing decreased growth in response to osmotic stress in plants [37] and particularly in canola [38]. RWC decreased and significant differences between the canola genotypes under different levels of drought stresses. Drought-sensitive genotypes were more affected by the decline in RWC than tolerant genotypes. Leaf temperature (LT) is an integrative trait and is affected by the following processes, such as the capacity of a vascular system to extract water from the soil in reaction to evaporative demand, photosynthetic potential via feedback on stomatal opening and the balance between sources and sinks. Thus, LT can be used to select for fitness in a range of environmental stresses and in favorable environments [39]. Temperature is also a determining factor in plant productivity, which acts as a double-edged sword, extreme fluctuations on either end resulting in stress conditions. High temperatures have been reported to impose constraints on plant reproduction and yield in canola [40], and low LT was reported a desirable trait under drought conditions [41]. In the current study, LT increased with drought, and significant differences between drought-tolerant and drought-sensitive canola genotypes were recorded. Chlorophyll fluorescence, as an efficient and non-destructive method, is a good way to measure the photochemical efficiency of PSII and detect the effect of stress in PSII [42]. Variation in the maximum quantum efficiency of PSII (Fv/Fm) is often confirmed by the concentration changes of specific photosynthetic pigments and cell structure modulations that are primarily affected by water stress and an estimate of this parameter provides information on photosynthetic electron transport [43,44]. Drought stress affects the reaction center and obstructs the electron transport system, which ultimately leads to a reduction in Fv/Fm and ROS generation [45,46]. Fv/Fm was significantly affected by the drought stress treatments in the four canola cultivars we analyzed, more so in the sensitive ones, suggesting that it might represent a valid indicator of the damage induced by water stress and be a tool for screening canola genotypes. Water stress increased the electrolyte leakage index (ELI) for all genotypes, and it was substantially higher in drought-sensitive genotypes, suggesting that ELI might also be considered a good indicator of drought tolerance in canola. Our results confirm Zarfam and Slm046 as tolerant and Okapi and Sarigol as sensitive genotypes [25,26]. ROS production leads to lipid peroxidation, the degradation of antioxidants and changes in gene expression [47,48]. Malondialdehyde (MDA) content is used as a sign of lipid peroxidation, which has often been used as a tool to evaluate oxidative damage in sensitive plants [49]. MDA contents significantly increased under drought conditions, and drought-sensitive canola genotypes showed a significantly higher increase in MDA in comparison with drought-tolerant genotypes. Together, high electrolyte leakage and high lipid peroxidation are indicative of the low stability of the cell membrane. It is assumed that the structural stability of cell membranes could be a main factor in protecting plant tissues against osmotic stress [50]. Redox homeostasis is the balance between the production and scavenging of ROS. When ROS production is higher than the cellular scavenging capacity, oxidative stress happens [51]. Plants possess antioxidant mechanisms to cope with oxidative damage and prevent damages to cells, and maximum antioxidant activity is displayed when plants need to overcome the oxidative stress [52]. The first enzyme in the antioxidant pathway is SOD, which converts highly reactive hydroxyl and superoxide radicals to less toxic H2O2, thus reducing damage to DNA, proteins and membranes [53]. H2O2 is still toxic to cells, and the scavenging of H2O2 is essential. It can be achieved through antioxidative defense systems, including GPOX and CAT [54]. SOD activity was higher in drought-tolerant canola genotypes under drought stress, indicating that they had better performance in reducing the stress damage caused by superoxide ions, involved in lipid peroxidation and membrane damage. Previous studies have reported that SOD activity increases in relation to drought stress [55,56,57]. GPOX activity was also enhanced under drought stress, and drought-tolerant genotypes displayed the highest activity, reinforcing the notion that they are more fit to tolerate drought stress. Previous studies also report increased GPOX activity in Brassica species under drought stress conditions [58]. The significant decrease in GPOX activity in drought-sensitive genotypes under severe drought stress suggests an increased damage that affects their ROS scavenging capacity. A similar conclusion can be drawn for the water-stress-induced decrease in CAT activity observed in the drought-sensitive genotypes. Previous works have shown that a reduction in CAT activity might happen under stressful conditions [54,59], which might be due to changes in the assembly of enzyme subunits or the inhibition of enzyme synthesis and degradation by peroxisomal proteases or photo-inactivation. Overall, the observed noticeable increased efficiency of the antioxidant enzyme complex of canola drought-tolerant genotypes might be correlated with tolerance mechanisms based on the fine-tuned regulation of its redox status in response to water deficit.
One of the most important responses to abiotic stress in plants is the change in cytosolic calcium concentration, as Ca2+ is an essential element in stress signaling. Calcium-dependent protein kinases (CDPKs) are activated by Ca2+ and mediate many signaling processes in plants, playing a synergistic role in the stress response of the plant [60,61]. In the present study, the transcription of CDPK genes BnaCDPK6, BnaCDPK10 and BnaCDPK14 was analyzed in response to water stress. The BnaCDPK6 and BnaCDPK14 relative expression increased with drought, with significant differences between tolerant and sensitive cultivars. BnaCDPK10 relative transcript accumulation increased under moderate drought and slightly decreased when more severe drought was applied. Canola and Arabidopsis belong to the Brassicaceae, and thus, both plants share genetic similarities. Our findings correspond to previous studies in Arabidopsis that have proven the involvement of several CDPKs in the plant’s response to drought [21,22,23]. Multiple canola CDPK genes have been checked for expression under various stresses [24], and our results are coincident with those previously reported for drought. The novelty of our results resides in the fact that BnaCDPK expression in response to water deficit is common to tolerant and sensitive genotypes, but importantly, expression is higher in tolerant than in sensitive cultivars, suggesting that BnaCDPKs are not only involved in drought-response mechanisms but also contribute to canola tolerance and open the possibility to canola engineering to overexpress the selected BnaCDPK genes and increase drought tolerance.
Gene ontology (GO) analyses provide an approach to associate transcriptome sequence data with biological understanding by linking differentially expressed genes to putative molecular functions [62]. The biological process, molecular function and cellular components related to the BnaCDPK genes were obtained from the Uniprot (https://www.uniprot.org/, accessed on 29 December 2021) summary output (Supplementary Table S2). Analysis of gene expression after stress application and GO might help improve our understanding of the mechanisms underlying the regulation of drought stress susceptibility and tolerance in this species. Furthermore, BnaCDPK genes were upregulated in the drought-tolerant genotypes, and this multifaceted stress response might be providing a considerable protection under water deficit conditions.
According to the results of the principal component analysis (PCA) (Supplementary Figure S1) and the simple correlations coefficients (Supplementary Table S3) between traits for canola genotypes under drought stress, significant positive and negative correlations were observed. The five indicators (leaf relative water content, leaf canopy temperature, PSII efficiency, leaf electrolyte leakage index and lipid peroxidation) used to classify the genotypes as drought-tolerant or drought-sensitive showed showed highly significant correlation values among them (Supplementary Table S3). The antioxidant enzymes also showed good correlation among them; however, their correlation with the drought indicators was not significant, probably due to the decreases in enzymatic activities observed for the sensitive cultivars when subjected to severe drought. BnaCDPK6 and BnaCDPK14 expression showed significant correlations with both the drought indicators and the antioxidant enzyme activities, which suggested that those CDPKs might be acting as regulatory proteins deciphering calcium signals triggered by water deficit stress.
Transcription factors (TFs) can regulate the expression of stress-responsive genes by binding to cis-acting elements located in the promoter sequence. The organization of promoter networks might provide insight into the prediction of how and when the expression of specific genes will be induced. A detailed analysis of the promoter region 2000 bp upstream of the coding sequence of BnaCDPK6, BnaCDPK10 and BnaCDPK14 revealed the presence of multiple cis elements. ABRE, GBox, WBox and ACGT motifs are preferred binding sites for the bZIP transcription factor, resulting in a specific response to tolerance to biotic and abiotic stresses [63]. ABRE (ABA-responsive-element) transcription factor regulates the gene response to osmotic stress and abscisic acid signaling [64,65]. WRKY and RAV transcription factors also play a key role in abscisic acid signaling [66,67]. MYB transcription factors constitute a large family and are involved in many plant processes, including drought stress responses [68]. The presence of these regulatory elements in the promoter region implies that BnaCDPKs might have critical functions in drought tolerance and represent positive regulators of the response to water stresses in canola.
Overall, our results suggest that ROS detoxification mechanisms, which contribute to drought tolerance in canola, might be triggered by BnaCDPKs gene expression, which appears to be regulated by cis elements in their promoter region. All analyzed physiological parameters (leaf relative water content, leaf canopy temperature, photosynthetic efficiency, electrolyte leakage, lipid peroxidation and antioxidant enzyme activities) appear to be valid indicators of drought tolerance in canola. Additionally, our results point to BnaCDPKs genes and their regulator elements as a potential molecular targets to obtain drought-tolerant productive canola cultivars through breeding or genetic transfor-mation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12010125/s1, Figure S1. Principal component analysis (PCA) of physiological and biochemical parameters and BnaCDPK genes expression in canola genotypes; Table S1. Analysis of variance of leaf relative water content (RWC), electrolyte leakage index (ELI), malondialdehyde (MDA), leaf temperature (LT), maximum quantum yield of PSII (Fv/Fm), antioxidant enzymes SOD, GPOX and CAT and Brassica napus calcium-dependent protein kinases BnaCDPK6, BnaCDPK10 and BnaCDPK14 relative gene expression in canola cultivars under drought stress; Table S2. Gene Ontology of BnaCDPK genes; Table S3. Simple correlation coefficients.
Author Contributions
For Conceptualization; H.A., A.A., J.J.P.; Methodology; H.A., A.A., J.J.P.; Software; H.A.; Validation; H.A., A.A., J.J.P.; Formal Analysis; H.A.; Investigation; H.A., A.A., J.J.P.; Resources; A.A., A.T., V.M., J.J.P.; Data Curation; H.A.; Writing–Original Draft Preparation; H.A.; Writing–Review & Editing; H.A., A.A., A.T., V.M., J.J.P.; Visualization; H.A., J.J.P.; Supervision; A.A., J.J.P.; Project Administration; A.A., A.T., V.M., J.J.P.; Funding Acquisition; J.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Agencia Estatal de Investigación (AEI), Spain (grant number AGL2017-88381-R).
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 22 December 2017).
- Elferjani, R.; Soolanayakanahally, R. Canola responses to drought, heat, and combined stress: Shared and specific effects on carbon assimilation, seed yield, and oil composition. Front. Plant Sci. 2018, 9, 1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhu, Z.; Guo, Q.; Ma, H.; Zhu, L. Alternate wetting and drying irrigation-mediated changes in the growth, photosynthesis and yield of the medicinal plant Tulipa edulis. Ind. Crops Prod. 2015, 66, 81–88. [Google Scholar] [CrossRef]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Efeoğlu, B.; Ekmekçi, Y.; Çiçek, N. Physiological responses of three maize cultivars to drought stress and recovery. S. Afr. J. Bot. 2009, 75, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Aberkane, H.; Belkadi, B.; Kehel, Z.; Filali-Maltouf, A.; Tahir, I.S.A.; Meheesi, S.; Amri, A. Assessment of drought and heat tolerance of durum wheat lines derived from interspecific crosses using physiological parameters and stress indices. Agronomy 2021, 11, 695. [Google Scholar] [CrossRef]
- Shokat, S.; Novák, O.; Široká, J.; Singh, S.; Gill, K.S.; Roitsch, T.; Großkinsky, D.K.; Liu, F. Elevated CO2 modulates the effect of heat stress responses in Triticum aestivum by differential expression of an isoflavone reductase-like gene. J. Exp. Bot. 2021, 72, 7594–7609. [Google Scholar] [CrossRef]
- Ulfat, A.; Shokat, S.; Li, X.; Fang, L.; Großkinsky, D.K.; Majid, S.A.; Roitsch, T.; Liu, F. Elevated carbon dioxide alleviates the negative impact of drought on wheat by modulating plant metabolism and physiology. Agric. Water Manag. 2021, 250, 106804. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The roles of reactive oxygen metabolism in drought: Not so cut and dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Shokat, S.; Großkinsky, D.K.; Roitsch, T.; Liu, F. Activities of leaf and spike carbohydrate-metabolic and antioxidant enzymes are linked with yield performance in three spring wheat genotypes grown under well-watered and drought conditions. BMC Plant Biol. 2020, 20, 400. [Google Scholar] [CrossRef]
- Whalley, H.J.; Knight, M.R. Calcium signatures are decoded by plants to give specific gene responses. New Phytol. 2013, 197, 690–693. [Google Scholar] [CrossRef]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and Calcineurin B–like Proteins: Calcium Sensors for Specific Signal Response Coupling in Plants. Plant Cell. 2002, 14, S389–S400. [Google Scholar] [CrossRef] [Green Version]
- Kudla, J.; Batistič, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef]
- Romeis, T.; Herde, M. From local to global: CDPKs in systemic defense signaling upon microbial and herbivore attack. Curr. Opin. Plant Biol. 2014, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.F.; Breton, G.; Harmon, A. Decoding Ca2+ signals through plant protein kinases. Annu. Rev. Plant Biol. 2004, 55, 263–288. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.-Y.; Wu, W.-H. AtCPK23 functions in Arabidopsis responses to drought and salt stresses. Plant Mol. Biol. 2007, 65, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tian, Y.-S.; Peng, R.-H.; Xiong, A.-S.; Zhu, B.; Jin, X.-F.; Gao, F.; Fu, X.-Y.; Hou, X.-L.; Yao, Q.-H. AtCPK6, a functionally redundant and positive regulator involved in salt/drought stress tolerance in Arabidopsis. Planta 2010, 231, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.-J.; Wei, F.-J.; Wang, C.; Wu, J.-J.; Ratnasekera, D.; Liu, W.-X.; Wu, W.-H. Arabidopsis calcium-dependent protein kinase CPK10 functions in abscisic acid- and Ca2+-mediated stomatal regulation in response to drought stress. Plant Physiol. 2010, 154, 1232–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Liu, W.-Z.; Zhang, Y.; Deng, M.; Niu, F.; Yang, B.; Wang, X.; Wang, B.; Liang, W.; Deyholos, M.K. Identification, expression and interaction analyses of calcium-dependent protein kinase (CPK) genes in canola (Brassica napus L.). BMC Genom. 2014, 15, 211. [Google Scholar] [CrossRef] [Green Version]
- Mirzai, M.; Moeini, A.; Ghanati, F. Effects of drought stress on the lipid peroxidation and antioxidant enzyme activities in two canola (Brassica napus L.) cultivars. J. Agric. Sci. Technol. (JAST) 2013, 15, 593–602. [Google Scholar]
- Zahedi, H.; Moghadam, H.T. Effect of drought stress on antioxidant enzymes activities with zeolite and selenium application in canola cultivars. Res. Crops 2011, 12, 388–392. [Google Scholar]
- Weatherley, P.E. Studies in the water relations of the cotton plant: I. The field measurements of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Strasserf, R.J.; Srivastava, A. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Polle, A.; Otter, T.; Seifert, F. Apoplastic peroxidases and lignification in needles of Norway spruce (Picea abies L.). Plant Physiol. 1994, 106, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H. Catalase In Vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Sabagh, A.E.; Hossain, A.; Barutçular, C.; Islam, M.S.; Ratnasekera, D.; Kumar, N.; Meena, R.S.; Gharib, H.S.; Saneoka, H.; da Silva, J.A.T. Drought and salinity stress management for higher and sustainable canola (Brassica napus L.) production: A critical review. Aust. J. Crop Sci. 2019, 13, 88–96. [Google Scholar] [CrossRef]
- Amani, I.; Fischer, R.A.; Reynolds, M.P. Canopy temperature depression association with yield of irrigated spring wheat cultivars in a hot climate. J. Agron. Crop Sci. 1996, 176, 119–129. [Google Scholar] [CrossRef]
- Morrison, M.J.; Stewart, D.W. Heat stress during flowering in summer Brassica. Crop Sci. 2002, 42, 797–803. [Google Scholar] [CrossRef]
- Balota, M.; Payne, W.A.; Evett, S.R.; Lazar, M.D. Canopy temperature depression sampling to assess grain yield and genotypic differentiation in winter wheat. Crop Sci. 2007, 47, 1518–1529. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Isoda, A. Effects of water stress on leaf temperature and chlorophyll fluorescence parameters in cotton and peanut. Plant Prod. Sci. 2010, 13, 269–278. [Google Scholar] [CrossRef]
- Miao, Y.; Bi, Q.; Qin, H.; Zhang, X.; Tan, N. Moderate drought followed by re-watering initiates beneficial changes in the photosynthesis, biomass production and Rubiaceae-type cyclopeptides (RAs) accumulation of Rubia yunnanensis. Ind. Crops Prod. 2020, 148, 112284. [Google Scholar] [CrossRef]
- Hussain, T.; Koyro, H.-W.; Zhang, W.; Liu, X.; Gul, B.; Liu, X. Low salinity improves photosynthetic performance in Panicum antidotale under drought stress. Front. Plant Sci. 2020, 11, 481. [Google Scholar] [CrossRef]
- Jia, Y.; Xiao, W.; Ye, Y.; Wang, X.; Liu, X.; Wang, G.; Li, G.; Wang, Y. Response of photosynthetic performance to drought duration and re-watering in maize. Agronomy 2020, 10, 533. [Google Scholar] [CrossRef] [Green Version]
- Biswas, K.; Adhikari, S.; Tarafdar, A.; Kumar, R.; Saha, S.; Ghosh, P. Reactive Oxygen Species and Antioxidant Defence Systems in Plants: Role and Crosstalk under Biotic stress. In Sustainable Agriculture in the Era of Climate Change; Springer: Berlin/Heidelberg, Germany, 2020; pp. 265–292. [Google Scholar]
- Choudhury, S.; Panda, P.; Sahoo, L.; Panda, S.K. Reactive oxygen species signaling in plants under abiotic stress. Plant Signal. Behav. 2013, 8, e23681. [Google Scholar] [CrossRef] [Green Version]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filek, M.; Walas, S.; Mrowiec, H.; Rudolphy-Skórska, E.; Sieprawska, A.; Biesaga-Kościelniak, J. Membrane permeability and micro-and macroelement accumulation in spring wheat cultivars during the short-term effect of salinity-and PEG-induced water stress. Acta Physiol. Plant. 2012, 34, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, P.M.; Baker, N.R. Oxidative stress: Antagonistic signaling for acclimation or cell death? Plant Physiol. 2010, 154, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Goswami, B.; Rankawat, R.; Gadi, B. Physiological and antioxidative responses associated with drought tolerance of Lasiurus sindicus Henr. endemic to Thar desert, India. Braz. J. Bot. 2020, 43, 761–773. [Google Scholar] [CrossRef]
- Guo, Z.; Ou, W.; Lu, S.; Zhong, Q. Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol. Biochem. 2006, 44, 828–836. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Zhang, S.; Liu, S.; Ma, H.; Chen, J.; Shen, Q.; Ge, C.; Zhang, X.; Pang, C.; Zhao, X. The compensation effects of physiology and yield in cotton after drought stress. J. Plant Physiol. 2018, 224, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shi, S.; Wang, B.; Zhao, J. Physiological and biochemical changes in different drought-tolerant alfalfa (Medicago sativa L.) varieties under PEG-induced drought stress. Acta Physiol. Plant. 2018, 40, 25. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Hasanloo, T.; Mohammadi, S. Physiological characteristics, antioxidant enzyme activities, and gene expression in 2 spring canola (Brassica napus L.) cultivars under drought stress conditions. Turk. J. Agric. For. 2015, 39, 413–420. [Google Scholar] [CrossRef]
- Jung, S. Variation in antioxidant metabolism of young and mature leaves of Arabidopsis thaliana subjected to drought. Plant Sci. 2004, 166, 459–466. [Google Scholar] [CrossRef]
- Ludwig, A.A.; Romeis, T.; Jones, J.D.G. CDPK-mediated signalling pathways: Specificity and cross-talk. J. Exp. Bot. 2004, 55, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Schulz, P.; Herde, M.; Romeis, T. Calcium-dependent protein kinases: Hubs in plant stress signaling and development. Plant Physiol. 2013, 163, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Falcon, S.; Gentleman, R. Using GOstats to test gene lists for GO term association. Bioinformatics 2007, 23, 257–258. [Google Scholar] [CrossRef] [Green Version]
- Llorca, C.M.; Potschin, M.; Zentgraf, U. bZIPs and WRKYs: Two large transcription factor families executing two different functional strategies. Front. Plant Sci. 2014, 5, 169. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.-i.; Hong, J.-h.; Ha, J.-o.; Kang, J.-y.; Kim, S.Y. ABFs, a family of ABA-responsive element binding factors. J. Biol. Chem. 2000, 275, 1723–1730. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biolology 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, C.Z.; Chen, Y.; Wang, C.; Kong, Y.H.; Wu, W.H.; Chen, Y.F. Arabidopsis RAV 1 transcription factor, phosphorylated by S n RK 2 kinases, regulates the expression of ABI 3, ABI 4, and ABI 5 during seed germination and early seedling development. Plant J. 2014, 80, 654–668. [Google Scholar] [CrossRef]
- Rushton, D.L.; Tripathi, P.; Rabara, R.C.; Lin, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J. WRKY transcription factors: Key components in abscisic acid signalling. Plant Biotechnol. J. 2012, 10, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Baldoni, E.; Genga, A.; Cominelli, E. Plant MYB Transcription factors: Their role in drought response mechanisms. Int. J. Mol. Sci. 2015, 16, 15811–15851. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).