
Exploring the Bush yam (Dioscorea praehensilis Benth) as a Source of Agronomic and Quality Trait Genes in White Guinea yam (Dioscorea rotundata Poir) Breeding

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Plant Materials
2.3. Experimental Design and Field Management
2.4. Data Collection
2.5. Statistical Analysis
3. Results
3.1. Variability in Key Agronomic and Tuber Quality Traits of D. praehensilis and D. rotundata
3.2. Genetic Variability and Broad-Sense Heritability of Agronomic and Tuber Quality Traits in D. praehensilis and D. rotundata
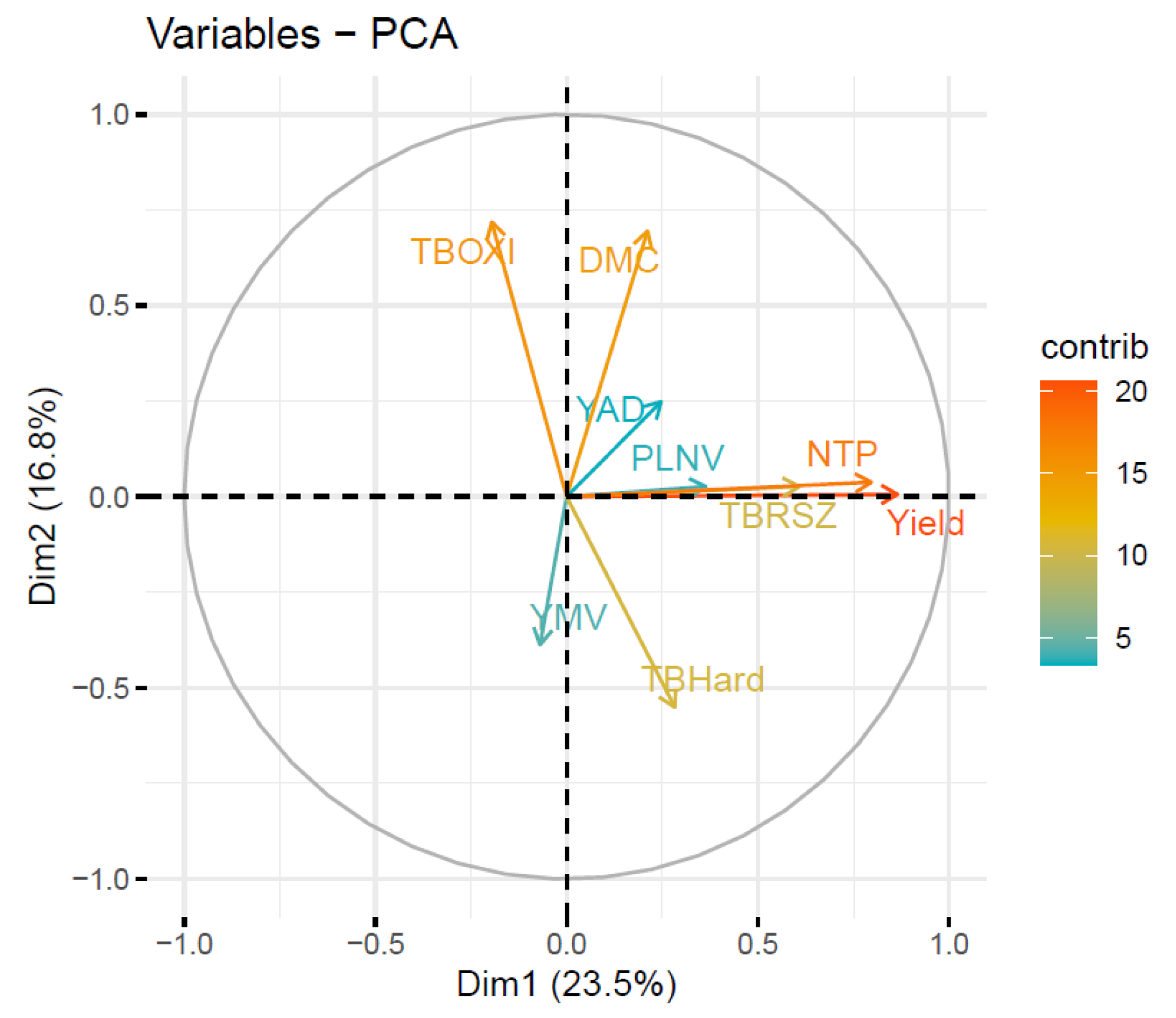
3.3. Principal Component Analysis of the Key Agronomic and Tuber Quality Traits
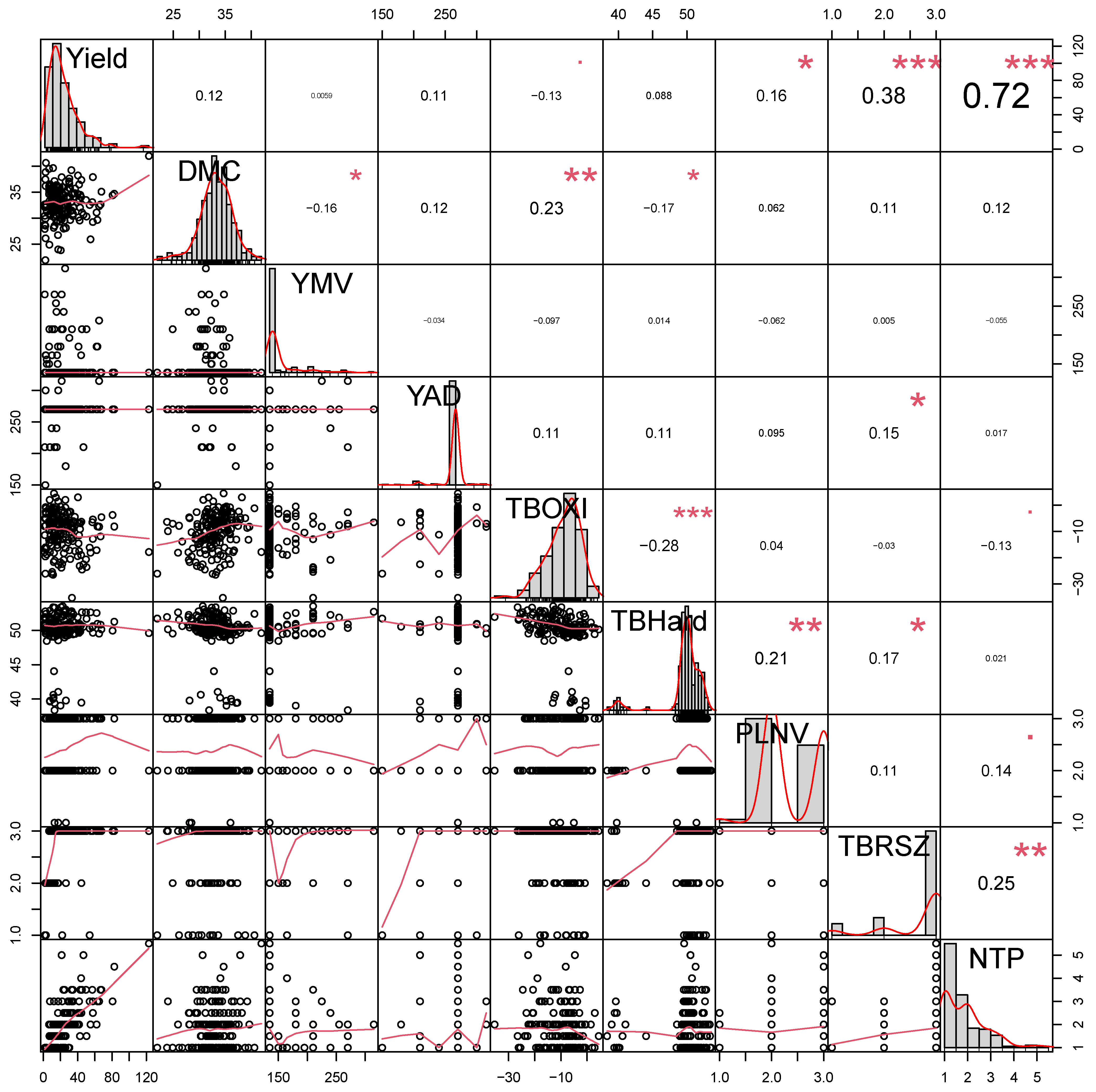
3.4. Relationships among Agronomic and Tuber Quality Traits
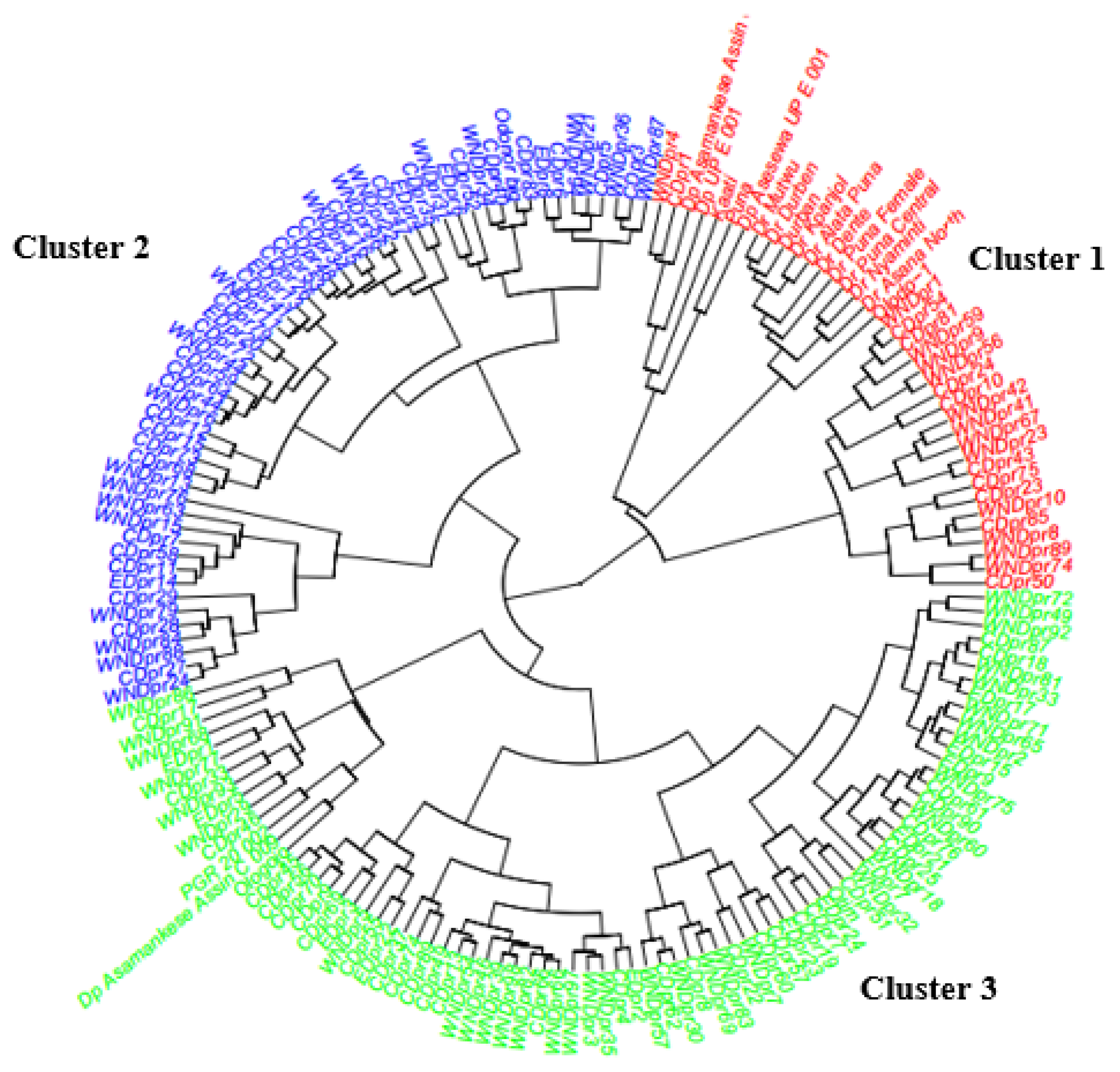
3.5. Hierarchical Clustering on Principal Components of D. praehensilis and D. rotundata Genotypes
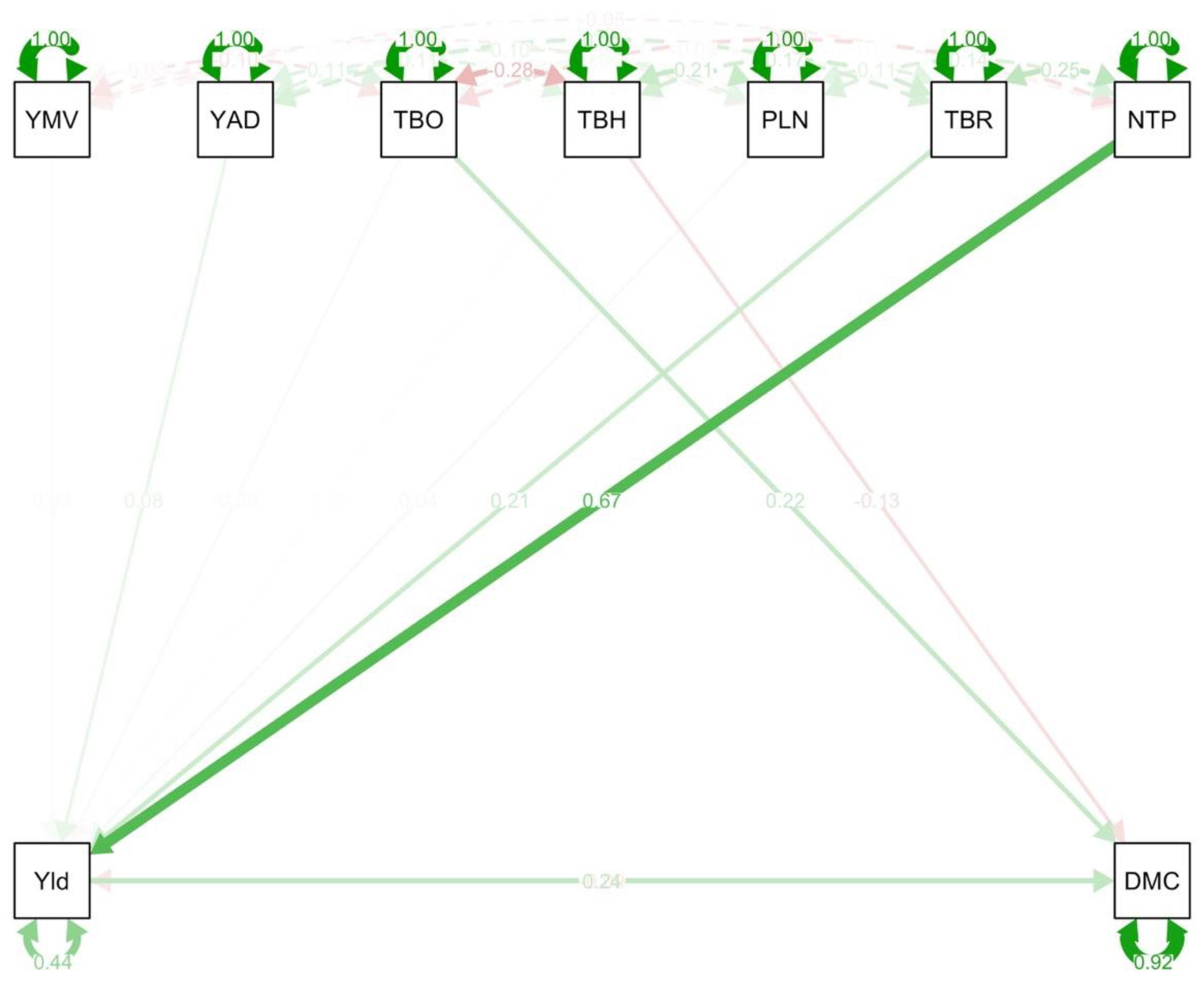
3.6. Path Coefficient Analysis among Assessed Traits of D. praehensilis and D. rotundata
4. Discussion
4.1. Variability in Key Agronomic and Tuber Quality Traits and Potential of D. praehensilis as Source of Genes for D. rotundata Breeding
4.2. Genetic Parameters and Broad-Sense Heritability of Assessed Traits
4.3. Correlation Coefficients, Principal Components, Path Coefficients, and Hierarchical Clusters among Assessed Traits of D. rotundata and D. praehensilis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asiedu, R.; Sartie, A. Crops that feed the World 1. Yams. Food Secur. 2010, 2, 305–315. [Google Scholar] [CrossRef]
- Sesay, L.; Norman, P.E.; Massaquoi, A.; Kobba, F.; Allieu, A.P.; Gboku, M.L.; Fomba, S.N. Assessment of farmers’ indigenous knowledge and selection criteria of yam in Sierra Leone. Sky J. Agric. Res. 2013, 2, 1–6. [Google Scholar]
- Alabi, T.R.; Adebola, P.O.; Asfaw, A.; De Koeyer, D.; Lopez-Montes, A.; Asiedu, R. Spatial multivariate cluster analysis for defining target population of environments in West Africa for yam breeding. Intern. J. Appl. Geospat. Res. 2019, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- FAO FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QL (accessed on 10 August 2021).
- Zannou, A.; Ahanchédé, A.; Struik, P.C.; Richards, P.; Zoundjihékpon, J.; Tossou, R.; Vodouhè, S. Yam and cowpea diversity management by farmers in the Guinea-Sudan transition zone of Benin. NJAS–Wagen. J. Life Sci. 2004, 52, 393–420. [Google Scholar] [CrossRef] [Green Version]
- Obidiegwu, J.E.; Akpabio, E.M. The geography of yam cultivation in southern Nigeria: Exploring its social meanings and cultural functions. J. Ethn. Foods 2017, 4, 28–35. [Google Scholar] [CrossRef]
- Thouvenel, J.C.; Dumont, R. Perte de rendement de l’igname infectee par le virus de la mosaique en Côte d’Ivoire. Agron. Trop. 1975, 45, 125–129. [Google Scholar]
- Adeniji, M.; Shoyinka, S.; Ikotun, T.; Asiedu, R.; Hughes, J.D.; Odu, B. Yield loss in guinea yam (Dioscorea rotundata poir.) due to infection by yam mosaic virus (YMV) genus potyvirus. Ife J. Sci. 2012, 14, 237–244. [Google Scholar]
- Frossard, E.; Aighewi, B.A.; Aké, S.; Barjolle, D.; Baumann, P.; Bernet, T.; Dao, D.; Diby, L.N.; Floquet, A.; Hgaza, V.K.; et al. The challenge of improving soil fertility in yam cropping systems of West Africa. Front. Plant Sci. 2017, 8, 1953. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, R.; Ishikawa, H.; Asfaw, A.; Asiedu, R. Low soil nutrient tolerance and mineral fertilizer response in White Guinea Yam (Dioscorea rotundata) genotypes. Front. Plant Sci. 2021, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Agre, P.A.; Norman, P.E.; Asiedu, R.; Asfaw, A. Identification of quantitative trait nucleotides and candidate genes for tuber yield and mosaic virus tolerance in an elite population of white Guinea yam (Dioscorea rotundata) using genome-wide association scan. BMC Plant Biol. 2021, 21, 552. [Google Scholar] [CrossRef]
- Agre, P.A.; Dassou, A.G.; Loko, L.E.; Idossou, R.; Dadonougbo, E.; Gbaguidi, A.; Mondo, J.M.; Muyideen, Y.; Adebola, P.O.; Asiedu, R.; et al. Diversity of white Guinea yam (Dioscorea rotundata Poir.) cultivars from Benin as revealed by agro-morphological traits and SNP markers. Plant Genet. Resourc. 2021, 19, 437–446. [Google Scholar] [CrossRef]
- Sugihara, Y.; Kudoh, A.; Oli, M.T.; Takagi, H.; Natsume, S.; Shimizu, M.; Abe, A.; Asiedu, R.; Asfaw, A.; Adebola, P.; et al. Population Genomics of Yams: Evolution and Domestication of Dioscorea Species. In Population Genomics; Springer: Cham, Switzerland, 2021; pp. 1–28. [Google Scholar] [CrossRef]
- Dempewolf, H.; Baute, G.; Anderson, J.; Kilian, B.; Smith, C.; Guarino, L. Past and future use of wild relatives in crop breeding. Crop Sci. 2017, 57, 1070–1082. [Google Scholar] [CrossRef]
- Lebot, V.; Abraham, K.; Kaoh, J.; Rogers, C.; Molisalé, T. Development of anthracnose resistant hybrids of the Greater Yam (Dioscorea alata L.) and interspecific hybrids with D. nummularia Lam. Genet. Resour. Crop Evol. 2019, 66, 871–883. [Google Scholar] [CrossRef]
- Couto, R.S.; Martins, A.C.; Bolson, M.; Lopes, R.C.; Smidt, E.C.; Braga, J.M.A. Time calibrated tree of Dioscorea (Dioscoreaceae) indicates four origins of yams in the Neotropics since the Eocene. Bot. J. Linn. 2018, 188, 144–160. [Google Scholar] [CrossRef]
- Darkwa, K.; Agre, P.; Olasanmi, B.; Iseki, K.; Matsumoto, R.; Powell, A.; Bauchet, G.; De Koeyer, D.; Muranaka, S.; Adebola, P.; et al. Comparative assessment of genetic diversity matrices and clustering methods in white Guinea yam (Dioscorea rotundata) based on morphological and molecular markers. Sci. Rep. 2020, 10, 13191. [Google Scholar] [CrossRef]
- Scarcelli, N.; Cubry, P.; Akakpo, R.; Thuillet, A.C.; Obidiegwu, J.; Baco, M.N.; Otoo, E.; Sonké, B.; Dansi, A.; Djedatin, G.; et al. Yam genomics supports West Africa as a major cradle of crop domestication. Sci. Adv. 2019, 5, eaaw1947. [Google Scholar] [CrossRef] [Green Version]
- Sugihara, Y.; Darkwa, K.; Yaegashi, H.; Natsume, S.; Shimizu, M.; Abe, A.; Hirabuchi, A.; Ito, K.; Oikawa, K.; Tamiru-Oli, M.; et al. Genome analyses reveal the hybrid origin of the staple crop white Guinea yam (Dioscorea rotundata). Proc. Natl. Acad. Sci. USA 2020, 117, 31987–31992. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, H.; Bhanu, N.A.; Srivastava, K.; Singh, M.; Shreya, H.A. Assessment of Genetic Diversity in Crop Plants—An Overview. Adv. Plants Agric. Res. 2017, 7, 279–286. [Google Scholar] [CrossRef]
- Pitalounani, W.E.N.; Dourma, M.; Wala, K.; Woegan, Y.; Gbogbo, A.; Batawila, K.; Dansi, A.; Tozo, K.; Akpagana, K. Agrodiversity, peasant management and importance of Dioscorea praehensilis (Benth.) in the Subhumid Zone of Togo. Afr. J. Food Agric. Nutr. Dev. 2017, 17, 12455–12475. [Google Scholar] [CrossRef]
- Adewumi, A.S.; Asare, P.A.; Adu, M.O.; Taah, K.J.; Akaba, S.; Mondo, J.M.; Agre, P.A. Farmers’ perceptions on varietal diversity, trait preferences and diversity management of bush yam (Dioscorea praehensilis Benth.) in Ghana. Sci. Afr. 2021, 12, e00808. [Google Scholar] [CrossRef]
- Mondo, J.M.; Agre, P.A.; Edemodu, A.; Adebola, P.; Asiedu, R.; Akoroda, M.O.; Asfaw, A. Floral Biology and Pollination Efficiency in Yam (Dioscorea spp.). Agriculture 2020, 10, 560. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae Tutorial (Version 1.3-5); Universidad Nacional Agraria: La Molina, Peru, 2021; 88p. [Google Scholar]
- Asfaw, A. Standard Operating Protocol for Yam Variety Performance Evaluation Trial; IITA: Ibadan, Nigeria, 2016; p. 27. [Google Scholar]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; John Wiley & Sons: Hoboken, NJ, USA, 1990. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Johnson, M.W.; Robinson, H.F.; Comstock, R.E. Genotypic and phenotypic correlations in soybeans and their implication in selection. Agron. J. 1955, 47, 477–483. [Google Scholar] [CrossRef]
- Deshmukh, S.N.; Basu, M.; Reddy, P.S. Genetic variability, character association, and path coefficient analysis of quantitative traits in Virginia bunch varieties of groundnut. Indian J. Agric. Sci. 1986, 56, 515–518. [Google Scholar]
- Wei, T.; Simko, V. R Package “corrplot”: Visualization of a Correlation Matrix (Version 0.84). 2017. Available online: https://github.com/taiyun/corrplot (accessed on 1 August 2021).
- Padhan, B.; Mukherjee, A.K.; Mohanty, S.K.; Lenka, S.K.; Panda, D. Genetic variability and inter species relationship between wild and cultivated yams (Dioscorea spp.) from Koraput, India based on molecular and morphological markers. Physiol. Mol. Biol. Plants 2019, 25, 1225–1233. [Google Scholar] [CrossRef]
- Anokye, M.; Tetteh, J.P.; Otoo, E. Morphological Characterization of Some Water Yam (Dioscorea alata L.) Germplasm in Ghana. J. Agric. Sci. Technol. 2014, 4, 518–532. [Google Scholar]
- Bassey, E.; Akpan, U. Evaluation of Guinea White Yam (Dioscorea rotundata Poir.) for Yield and Yield Components in Nigeria. J. Exp. Agric. Int. 2015, 8, 216–223. [Google Scholar] [CrossRef]
- Kouam, E.B.; Avana-Tientcheu, M.L.; Lekeumo, V.D.; Akitio, H.M.; Khasa, D.P.; Pasquet, R.S. Agro-ecological distribution of the phenotypic diversity of aerial yam (Dioscorea bulbifera L.) in Cameroon using multivariate analysis: Prospect for germplasm conservation and improvement. Open Agric. 2018, 3, 190–206. [Google Scholar] [CrossRef]
- Ayisah, K.D.; Mawussi, G.; Tchaniley, L.; Aziadekey, M.K. Potentials of Cultivated Varieties and Wild Yam Seeds as Efficient Alternative Plant Genetic Resources for Resistant Genotypes against Yam Mosaic Virus (YMV) in Togo. J. Exp. Agric. Int. 2020, 42, 82–93. [Google Scholar] [CrossRef]
- Padhan, B.; Panda, D. Variation of photosynthetic characteristics and yield in wild and cultivated species of yams (Dioscorea spp.) from Koraput, India. Photosynthetica 2018, 56, 1010–1018. [Google Scholar] [CrossRef]
- Norman, P.E.; Tongoona, P.B.; Danquah, A.; Danquah, E.Y.; Agre, P.A.; Agbona, A.; Asiedu, R.; Asfaw, A. Genetic parameter estimation and selection in advanced breeding population of white Guinea yam. J. Crop Improv. 2021, 35, 790–815. [Google Scholar] [CrossRef]
- Siadjeu, C.; Toukam, G.M.; Bell, J.M.; Nkwate, S. Genetic diversity of sweet yam Dioscorea dumetorum (Kunth) Pax revealed by morphological traits in two agro-ecological zones of Cameroon. Afr. J. Biotechnol. 2015, 14, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, R.; Nwadili, C.O.; Saski, C.A.; Agre, P.; Scheffler, B.E.; Augusto, J.; Lopez-Montes, A.; Onyeka, J.T.; Lava-Kumar, P.; Bandyopadhyay, R. An EST-SSR based genetic linkage map and identification of QTLs for anthracnose disease resistance in water yam (Dioscorea alata L.). PLoS ONE 2018, 13, e0197717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agre, P.; Asibe, F.; Darkwa, K.; Edemodu, A.; Bauchet, G.; Asiedu, R.; Adebola, P.; Asfaw, A. Phenotypic and molecular assessment of genetic structure and diversity in a panel of winged yam (Dioscorea alata) clones and cultivars. Sci. Rep. 2019, 9, 18221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asfaw, A.; Aderonmu, D.S.; Darkwa, K.; De Koeyer, D.; Agre, P.; Abe, A.; Olasanmi, B.; Adebola, P.; Asiedu, R. Genetic parameters, prediction, and selection in a white Guinea yam early-generation breeding population using pedigree information. Crop Sci. 2021, 61, 1038–1051. [Google Scholar] [CrossRef]
- Tewodros, M.; Firew, M.; Shimelis, H.; Endale, G. Interrelationship and Path analysis of Tuber yield and related traits in yam (Dioscorea spp.) from Ethiopia. Res. J. Pharm. Phytochem. 2020, 12, 207–218. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Squares | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sources of Variation | df | Tuber Yield (t ha−1) | DMC (%) | YMV | YAD | TBOXI | TBHard (N) | PLNV | TBRSZ | NTP |
| Replicate | 1 | 946.61 * | 323.41 * | 0.65 NS | 1.28 × 10−24 * | 288.63 NS | 0.15 NS | 0.003 NS | 0.003 NS | 7.47 * |
| Block | 14 | 194.37 NS | 9.97 NS | 1704.20 * | 271.42 * | 112.12 NS | 12.21 * | 0.44 * | 0.16 * | 1.28 NS |
| Species | 1 | 1824.46 * | 31.19 NS | 602.71 * | 1814.16 * | 238.16 NS | 2395.04 * | 2.06 * | 1.38 * | 0.69 NS |
| Genotype | 172 | 640.93 * | 19.58 * | 1861.65 * | 553.71 * | 92.37 NS | 2.59 * | 0.54 * | 0.68 * | 1.65 * |
| Residual | 159 | 209.96 | 12.62 | 0.64 | 1.91 × 10−24 | 81.98 | 0.37 | 0.01 | 0.01 | 0.74 |
| Species | Tuber Yield (t ha−1) ± SD | DMC (%) ± SD | YMV ± SD | YAD ± SD | TBOXI ± SD | TBHard (N) ± SD | PLNV ± SD | TBRSZ ± SD | NTP ± SD |
|---|---|---|---|---|---|---|---|---|---|
| D. praehensilis | 23.47 ± 18.53 a | 32.83 ± 3.16 a | 147.45 ± 31.00 b | 267.78 ± 16.48 a | −10.36 ± 7.13 a | 50.76 ± 1.15 a | 2.43 ± 0.52 a | 2.73 ± 0.61 a | 1.89 ± 0.93 a |
| Min. | 1.67 | 21.90 | 135.00 | 210.00 | −35.30 | 39.60 | 1.00 | 1.00 | 1.00 |
| Max. | 123.00 | 41.96 | 270.00 | 315.00 | 4.43 | 53.55 | 3.00 | 3.00 | 5.50 |
| D. rotundata | 16.39 ± 10.23 b | 34.00 ± 2.82 a | 157.50 ± 40.70 a | 260.00 ± 23.35 b | −6.47 ± 4.25 a | 40.10 ± 1.42 b | 1.92 ± 0.29 b | 2.25 ± 0.45 b | 1.71 ± 0.69 a |
| Min. | 7.67 | 28.20 | 135.00 | 210.00 | −13.08 | 39.00 | 1.00 | 2.00 | 1.00 |
| Max. | 44.34 | 37.14 | 270.00 | 270.00 | −0.55 | 41.03 | 2.00 | 3.00 | 2.50 |
| CV (%) | 63.40 | 10.79 | 0.53 | 0.27 | 89.67 | 1.22 | 4.94 | 4.35 | 45.90 |
| Genetic Parameters | |||||
|---|---|---|---|---|---|
| Traits | δ2g | δ2p | GCV (%) | PCV (%) | H2b (%) |
| Tuber yield (t ha−1) | 229.6 | 435.4 | 66.3 | 91.3 | 52.7 |
| DMC (%) | 4.0 | 16.6 | 6.1 | 12.4 | 24.1 |
| YMV | 994.8 | 995.4 | 21.4 | 21.4 | 99.9 |
| YAD | 162.0 | 164.0 | 4.8 | 4.8 | 98.8 |
| TBOXI | 4.4 | 89.2 | 20.8 | 93.5 | 4.9 |
| TBHard (N) | 8.5 | 8.9 | 5.8 | 6.0 | 95.5 |
| PLNV | 0.3 | 0.3 | 21.9 | 22.6 | 93.3 |
| TBRSZ | 0.3 | 0.5 | 21.2 | 24.7 | 73.9 |
| NTP | 0.5 | 1.2 | 37.6 | 58.5 | 41.3 |
| Traits | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 |
|---|---|---|---|---|---|---|---|---|---|
| Yield | 0.0041 | −0.8613 | −0.5015 | 0.0667 | −0.0207 | 0.0171 | −0.0355 | −0.0156 | 0.0015 |
| DMC | −0.0162 | −0.0257 | 0.0094 | 0.1242 | 0.8464 | −0.5164 | −0.0111 | −0.0188 | −0.0059 |
| YMV | 0.9992 | −0.0096 | 0.0268 | 0.023 | 0.0104 | −0.008 | 0.0018 | −0.0002 | 0.0006 |
| YAD | −0.0263 | −0.5051 | 0.8598 | −0.0626 | 0.0018 | 0.0295 | 0.0035 | −0.0026 | −0.0012 |
| TBOXI | −0.0232 | 0.0268 | 0.0886 | 0.9768 | −0.1813 | −0.0611 | 0.0062 | −0.0031 | −0.0084 |
| TBHard | 0.0014 | −0.0205 | 0.0073 | −0.1467 | −0.4997 | −0.8516 | 0.0122 | −0.0353 | −0.0414 |
| PLNV | −0.0011 | −0.0049 | −0.0001 | 0.0027 | −0.0154 | −0.0412 | 0.1198 | 0.0835 | 0.9883 |
| TBRSZ | 0.0001 | −0.0128 | −0.0029 | 0.0009 | −0.003 | −0.0371 | −0.1019 | 0.9913 | −0.073 |
| NTP | −0.0015 | −0.0301 | −0.0219 | −0.0006 | 0.0176 | 0.0068 | 0.9868 | 0.092 | −0.1269 |
| Eigenvalue | 2.115 | 1.516 | 1.207 | 0.994 | 0.876 | 0.765 | 0.712 | 0.557 | 0.258 |
| Variance (%) | 23.505 | 16.843 | 13.412 | 11.040 | 9.736 | 8.504 | 7.913 | 6.184 | 2.864 |
| Cumulative (%) | 23.505 | 40.348 | 53.760 | 64.800 | 74.535 | 83.040 | 90.952 | 97.136 | 100.000 |
| Traits | Cluster 1 ± SD (39) | Cluster 2 ± SD (56) | Cluster 3 ± SD (79) | F-value |
|---|---|---|---|---|
| Tuber yield (t ha−1) | 12.19 ± 12.70 c | 30.91 ± 22.14 a | 22.69 ± 14.20 b | 14.11 *** |
| Dry matter content (%) | 33.14 ± 3.67 ab | 33.81 ± 2.60 a | 32.17 ± 3.08 b | 4.76 ** |
| Yam mosaic virus | 145.77 ± 31.15 a | 137.68 ± 10.36 b | 155.60 ± 39.03 a | 5.66 * |
| Yam anthracnose disease | 256.15 ± 31.42 b | 270.54 ± 4.01 a | 270.38 ± 8.65 a | 11.94 *** |
| Tuber flesh oxidation | −8.17 ± 6.04 b | −9.43 ± 6.59 ab | −11.52 ± 7.56 a | 3.42 * |
| Tuber flesh hardness (N) | 47.44 ± 5.07 b | 50.47 ± 0.95 a | 50.98 ± 1.31 a | 25.30 *** |
| Plant vigour | 2.23 ± 0.48 b | 2.96 ± 0.19 a | 2.09 ± 0.36 c | 107.15 *** |
| Tuber size | 1.79 ± 0.73 b | 2.96 ± 0.19 a | 2.95 ± 0.22 a | 133.64 *** |
| Number of tubers per plot | 1.55 ± 0.86 b | 2.25 ± 1.03 a | 1.77 ± 0.78 b | 8.15 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adewumi, A.S.; Agre, P.A.; Asare, P.A.; Adu, M.O.; Taah, K.J.; Mondo, J.M.; Akaba, S. Exploring the Bush yam (Dioscorea praehensilis Benth) as a Source of Agronomic and Quality Trait Genes in White Guinea yam (Dioscorea rotundata Poir) Breeding. Agronomy 2022, 12, 55. https://doi.org/10.3390/agronomy12010055
Adewumi AS, Agre PA, Asare PA, Adu MO, Taah KJ, Mondo JM, Akaba S. Exploring the Bush yam (Dioscorea praehensilis Benth) as a Source of Agronomic and Quality Trait Genes in White Guinea yam (Dioscorea rotundata Poir) Breeding. Agronomy. 2022; 12(1):55. https://doi.org/10.3390/agronomy12010055
Chicago/Turabian StyleAdewumi, Adeyinka S., Paterne A. Agre, Paul A. Asare, Michael O. Adu, Kingsley J. Taah, Jean M. Mondo, and Selorm Akaba. 2022. "Exploring the Bush yam (Dioscorea praehensilis Benth) as a Source of Agronomic and Quality Trait Genes in White Guinea yam (Dioscorea rotundata Poir) Breeding" Agronomy 12, no. 1: 55. https://doi.org/10.3390/agronomy12010055
APA StyleAdewumi, A. S., Agre, P. A., Asare, P. A., Adu, M. O., Taah, K. J., Mondo, J. M., & Akaba, S. (2022). Exploring the Bush yam (Dioscorea praehensilis Benth) as a Source of Agronomic and Quality Trait Genes in White Guinea yam (Dioscorea rotundata Poir) Breeding. Agronomy, 12(1), 55. https://doi.org/10.3390/agronomy12010055