Abstract
6-pentyl-α-pyrone (6-PP) is a powerful Magnaporthiopsis maydis antifungal compound, recently discovered when the potent growth medium of Trichoderma asperellum was analyzed. Despite its high potential in plate assay, it was not inspected for plant treatment prevention. Late wilt disease, caused by the fungus M. maydis, threatens commercial maize production in high-risk areas. Thus, the search for control options against the pathogen is one of the top priorities in Israel, Egypt, and other countries. Disease-resistant maize genotypes can reduce the damages. Yet, aggressive variants of the fungus can overcome host resistance. The current study aimed at inspecting T. asperellum and its secreted metabolite, pure 6-PP, against the pathogen in plants over a full growth period. First, adding T. asperellum directly to seeds with sowing provides significant protection to sprouts (up to 42 days) in a growth room, with more than two-fold growth promotion and reduced pathogen root infection (detected by real-time PCR). The same procedure applied in a commercial field was less beneficial in rescuing the plants’ growth and yield. Still, it reduced the cobs’ symptoms by 11% and resulted in nine-fold lower levels of the pathogen’s DNA in the stem tissue. Second, the T. asperellum purified 6-PP compound (30 µg/seed) was used in seed coating and tested against the T. asperellum secretory metabolites’ crude (diluted to 50%). At the season’s end, these treatments improved plant biomass by 90–120% and cob weight by 60%. Moreover, the treatments significantly (p < 0.05) reduced the symptoms (up to 20%) and pathogen infection (94–98%). The current study’s results reveal the potential of 6-PP as a new fungicide against M. maydis. Such a treatment may protect maize plants from other soil diseases.
1. Introduction
The world’s corn (Zea mays L., maize) harvested area is the third largest after wheat and rice (FAOSTAT, 2020 Food and Agriculture Commodity Production data), reflecting its importance globally. Maize late-wilt disease (LWD) can cause acute damage to susceptible hybrids in infected areas under favorable conditions. Such a scenario prevails in Egypt [1,2], Israel [3], Portugal [4], Spain [5], India [6], and other less impacted regions [7,8]. LWD is typified by rapid dehydration of fodder and sweet maize, typically from the tasseling stage until just before maturity [9,10]. The disease’s causal agent is the fungus Magnaporthiopsis maydis, which is recognized by two additional synonyms, Cephalosporium maydis and Harpophora maydis [11,12,13]. The fungus is a hemibiotroph, soil-borne [14], and seed-borne [15], and can be spread as sclerotia, conidia, or hype on plant remains. The pathogen can survive for prolonged periods in the soil or by establishing inside alternative host plants, such as Lupinus termis L. (lupine) [16], Setaria viridis (green foxtail), Citrullus lanatus (watermelon), and Gossypium hirsutum L. (cotton) [17,18]. It may act as an endophyte in those secondary hosts [19] but also as an opportunist, causing severe disease in some environments and under host-associated conditions [20,21].
The M. maydis pathogenesis in LWD-sensitive corn hybrids is well-described in the academic literature [22]. Sabet et al. [23] reported that the roots’ penetration and inoculation occur throughout the first 21 days post-seeding. At the end of this period, the pathogen appears in the vascular system (the xylem) and spreads up in the shoot. At tasseling (ca. day 50–60, near the male flowering), the pathogen could be detected in the stem, its DNA levels peaked [24], and the first shoot symptoms were revealed shortly later [10]. On ca. day 70, the last week or two before harvest, the fungus hype and secreted substances impeded the water supply and, in severe cases, led to the host’s acute dehydration and death [25]. The symptoms were increased under drought stress [5,26,27]. Economic losses due to LWD were up to 40% in Egypt [28], 100% in Israel [24], and 51% in India [29]. A similar but delayed disease mode appears in resistant genotypes, and M. maydis can innoculate and be established in the seeds of these apparently healthy plants and increase its dispersal [3].
Nowadays, there is a necessity to improve our toolkit to investigate the pathogen, restrict its spread, and limit its impact [24]. Past efforts to control LWD had relied on using agricultural (flood following and balanced soil fertility) [30,31,32], biological [1,33,34,35], physical (solar heating) [36], allelochemical [37,38] and chemical solutions [39,40], tillage, cover crop or crop rotation [4,41,42], as well as other options [43,44,45], with various levels of success. Despite these efforts, the most efficient, environmentally friendly, and cost-effective way today to contain the disease is relying on resistant maize hybrids [6,46,47,48,49]. Yet, the presence of highly violent M. maydis isolates [50,51] that may overcome host resistance pushes scientists to seek other approaches.
In Israel, maize is an important crop, with a harvested area of 3,820 ha and volume production of 73,419 tons (FAOSTAT, 2020 Food and Agriculture Commodity Production data). The average production quantity of maize crops in metric ton yield per metric hectare exhibited an increasing trend, from 5–7 in 1980–1984 to 10–15 in 1990–2000 and 20–25 in 2010–2020 (Figure 1). Effective LWD risk management may be a factor in this success, mainly by avoiding sensitive maize cultivars’ growth [52].
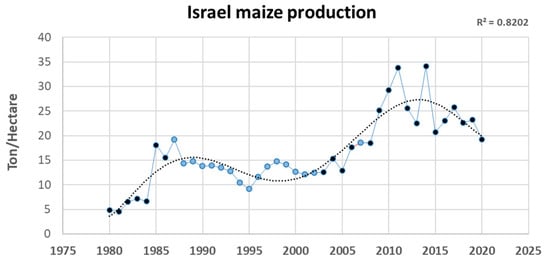
Figure 1.
Israel maize production per metric hectare between the years 1980 and 2020. Data from FAOSTAT, 2020 Food and Agriculture Commodity Production. Values in dark blue are official data. Values in light blue are unofficial data or estimations. The trend line is a sixth-order polynomial.
It was demonstrated that M. maydis is present in the host tissues of successfully chemically treated plants [52]. This may result in the development of the fungous’s fungicide immunity. The most effective fungicide against the late-wilt agent is Azoxystrobin [52,53]. Still, the rapid expansion of this fungicide tolerance has become increasingly problematic. Furthermore, the antifungals’ widespread use creates a major concern—their remaining toxicity and influence are endangering human health and the environment [54].
Several studies have already demonstrated microorganism-based LWD bio-control (most recently [1,34,55,56,57,58]). A particularly studied method is based on plant growth-promoting rhizobacteria, which could also enhance plant health (summarized by [59]). An alternative approach is using Trichoderma species against the LWD pathogen. Members in this genus can develop endophytic mutualistic interactions with various plant species [60], while other species have been used as a bio-protective barrier against phytoparasitic fungi [61]. Still, many knowledge gaps exist regarding the effectiveness of Trichoderma spp. against M. maydis. Therefore, their full potential as a biopesticide is yet to be explored. To demonstrate this potential, it was shown [33] that adding Chlorella vulgaris (microalgae) extracts to each of the Trichoderma species T. koningii and T. virens was an efficient treatment against LWD, resulting in a 72% disease frequency reduction in the greenhouse and 2.5-fold higher grain yield in the field.
Over the past few years, we tested nine Trichoderma isolates versus M. maydis. Three of these isolates revealed control ability against the maize pathogen: Trichoderma asperelloides (T.203) and Trichoderma longibrachiatum (T.7407 from a marine sponge); and Trichoderma asperellum (strain P1), an endophytic symbiont isolated from the seeds of an LWD-sensitive maize cultivar [57,58]. Growth-room seedlings pathogenicity trials show that all three Trichoderma isolates promoted the inspected plants’ growth indices, reduced LWD dehydration signs, and caused a significant reduction in M. maydis DNA inside the host tissues. Similar results were achieved in a full-season field trial (82 days) [55].
In follow-up work, we isolated a potent inhibitor chloroform extract (approx. 400 mg/L) from T. asperellum growth medium, capable of entirely preventing M. maydis growing [56]. Liquid chromatography-mass spectrometry (LC-MS) and gas chromatography-mass spectrometry (GC-MS) purification steps identified the secreted active ingredient as 6-pentyl-α-pyrone (also called pyrone 6-pentyl-2H-pyran-2-one or 6-PP). The pure compound exhibited high antifungal efficiency versus the LWD pathogen. Based on reports in the literature, this active ingredient is identified as a key bioactive compound of various Trichoderma species [62].
This metabolite plays a pivotal role in the biocontrol capability of the producing species. It displays multiple actions against phytoparasitic fungi, such as restricting their spore germination, mycelium pigmentation, and hyphal growth [63]. 6-PP also reduces fusaric acid production by Fusarium moniliform and mycotoxin deoxynivalenol by Fusarium graminearum [64]. For commercial production, a large amount of 6-PP can be generated by T. asperellum by utilizing forced aeration on a solid-state fermentation system [62]. Because 6-PP is a food-grade volatile compound, it may also be applied post-harvest to shield crops in storage.
The current study aimed at developing a biological environmentally friendly control strategy against M. maydis LWD based directly on T. asperellum, its secreted metabolites’ crude, or pure 6-PP. To this end, we conducted growth room trials in seedlings (up to 42 days) and full-season experiments in a net house or the field. Tracking M. maydis’s DNA within the host tissues using quantitative real-time PCR (q-PCR) enables direct evaluation of the treatments’ potential.
2. Materials and Methods
2.1. Rationale and Research Design
This three-year study examined the biocontrol potential of Trichoderma asperellum (P1 strain) and its secreted metabolites, specifically the 6-pentyl-α-pyrone (6-PP, Sigma-Aldrich, CAS number 27593-23-3, Buchs, Switzerland) purified active ingredient, against the maize late wilt pathogen, Magnaporthiopsis maydis. A test series was conducted to evaluate the potential of this new green biopesticide. These trials started by directly applying T. asperellum with the seeding in sprouts’ pathogenicity evaluation (growth room conditions up to 42 days) and under field conditions for the entire growing season. The following step examined the T. asperellum secreted metabolites (growth medium crude) against the purified active ingredient, 6-PP, in a semi-field environment (net house) full-season experiment. While the growing room provides a controlled environment, the net house offers a better field-like environment while maintaining more uniform environmental conditions and better regulation of soil composition, inoculation load, and water management. Both trials enable adding non-infected plants as a negative control. Therefore, it is an important step designed to achieve a complete image. Field experiments, while providing a more actual commercial growth environment, are unstable regarding physical and climatic conditions, pathogen existence, and other factors (e.g., soil composition and microbiome), as well as the cross-influence between treatments.
The preliminary steps—inspecting T. asperellum biocontrol potential using the sprout pathogenicity assay (in a growing room up to 42 days from sowing, DAS) and the field trial—were conducted using the late-wilt-susceptible maize genotype Prelude (a sweet maize cultivar produced by SRS Snowy River Seeds, Australia, marketed by Green 2000 Ltd., Bitan Aharon, Israel). The T. asperellum secreted metabolites’ examination in the net house was made using the sweet maize genotype Megaton cv. (from Limagrain, Saint-Beauzire, Puy-de-Dôme, France, supplied by Hazera Seeds Ltd.). These two cultivars were tested previously for their susceptibility to the late wilt pathogen [41,52] and identified as LWD-highly sensitive hybrids. A special effort was devoted to maintaining similar conditions in the growth room and mesh house experiments. Both trials were conducted based on the same soil conditions, inoculation method, and controls.
2.2. Fungal Species and Growth Conditions
All M. maydis isolates (Table 1) chosen for the current work were examined in our lab for their aggressiveness towards LWD-susceptible maize hybrids. They were found to have moderate pathogenicity levels (Ofir Degani, personal communication). The T. asperellum (P1 strain) endophyte was separated from the sweet maize Prelude cv. grains and identified as described before [58].

Table 1.
Fungi used in this research.
The fungi were grown on solid potato dextrose agar (PDA; Difco Laboratories, Detroit, MI, USA) under dark conditions at an optimal temperature of 28 ± 1 °C. They were transferred to a new growth dish by extracting a 6-mm-diameter agar disk from the margins of the culture and sowing it to a new PDA Petri dish. Plates were labeled and incubated in the dark at 28 ± 1 °C for 4–6 days. For submerged cultures (in a liquid substrate), five fungal disks were sown in an Erlenmeyer bottle containing 150-mL potato dextrose broth (PDB; Difco Laboratories, Detroit, MI, USA). The bottles were plugged with a breathable stopper and incubated for six days, shaken at 150 rpm in the dark, at 28 ± 1 °C. The growth medium metabolites were extracted by filtering the liquid PDB growth medium using a Buchner funnel and Whatman filter no. 3.
2.3. Trichoderma Asperellum Pathogenicity Evaluation in Growth Room Sprouts
2.3.1. Inoculation Method
Two-liter pots were filled with commercial field peat soil from a location with a long record of LWD infection (Amir field, Mehogi-1 plot, coordinates: 33°09′59″ N 35°36′5″ E) [52,53]. Control was a similar peat soil from a nearby farm (Gadash experimental, coordinates: 33°10′48.6″ N 35°35′11.6″ E) with no record of LWD. If such an infestation occurred, it was supposed to be minor. The soil was blended with 30% Perlite No. 4 (for aerating). Complementary inoculation was conducted in two steps to guarantee a high and uniform infection load: (1) pre-sowing soil infection with sterilized infected wheat grains; and (2) adding M. maydis colony agar disks with the seeding. The sterilized infected wheat grains were prepared as previously described [41]. Briefly, the wheat seeds were drenched in tap water overnight, filtered, and dried on paper towels in a fume hood. The seeds were sterilized in an autoclave for 30 min at 120 °C, and 150 g was incubated with 10 M. maydis mycelium agar disks of M. maydis (see Section 2.2) in sterilized plastic boxes (0.5 l volume) for 2–3 weeks at 28 ± 1 °C in the dark. The seeds were mixed every two days by shaking to ensure equal inoculation. The soil was infected by adding and mixing 12 g of sterilized and infected wheat grains to the top 10 cm of the ground in 2 L pots. The inoculated soil was kept moist until the maize was sown for a week. Additionally, three colony M. maydis colony agar disks were added to each seed with the sowing.
2.3.2. Trichoderma asperellum Biocontrol-Based Treatment
The biocontrol agent, T. asperellum, was grown in a liquid PDB (see Section 2.2) for six days, filtered, and chopped into small hyphae fragments, and 1 g (wet weight) of mycelia was added to each seed with the sowing.
2.3.3. Growth Room Trial Conditions
The experiment was conducted in 2019 in five biological repetitions and included two controls—infected unprotected plants and healthy plants (without any treatment). Two-liter pots were filled with field peat soil mixed with 30% Perlite, as in Section 2.3.1. Five Prelude cv. maize seeds were seeded in each pot to a depth of 4 cm. The plants were developed in a growing room for 42 days (fifth leaf appearance, V5 phenological stage) in an artificial light regime of 16 and 8 h of darkness, at 28 ± 3 °C with 45–50% humidity. Each pot was irrigated routinely with 100 mL of tap water every two days.
2.4. Trichoderma Asperellum Pathogenicity Evaluation in the Field
2.4.1. Overall Description of the Field Experiments
This study evaluated the T. asperellum biocontrol capability against M. maydis over a full growing season in a commercial field conditions simulation. The experiment was carried out on a trial farm (Gadash, Hula Valley, Upper Galilee, north Israel) throughout the summer and fall of 2020. The meteorological records through the growth period are listed in Table 2. The 2020 trial was conducted in autumn for pragmatic reasons (principally the field availability). Thus, due to the late season, the average meteorological parameters quantified during the experimental duration (relatively low temperature, radiation and evaporation, and high precipitation) were not optimal for the LWD burst.
Table 2.
Meteorological data for the 2020 field experiments 1.
The experiment field area has a history of mild LWD infestation. We infected the experimental plots (apart from the control plots) to attain a more acute disease. Therefore, the negative check plots were without pathogen enrichment. The Prelude cv. seeds were pre-treated (according to the regulations) with thiram, captan, carboxin, metalaxyl-M (Rogers/Syngenta Seeds, Boise, ID, the USA, provided by CTS, Tel Aviv, Israel). The experiment was performed with five random blocks per treatment/control. Each block comprised five garden beds (repeats), which were 9 m long, 6 m wide, and had two rows with row spacing of 96 cm.
2.4.2. Sowing Procedure and Irrigation
The plots were seeded to a 4 cm depth with 7 plants/meter and sprouted the next day with a frontal irrigation system. Irrigation of 0.6 l h−1 was conducted by drip irrigation line (20 mm) for each row (Dripnet PC1613 F, Netafim USA, Fresno, CA) and was controlled using a computerized system. The watering supply was 3000 L/0.1 ha per day. The watering was conducted every two days, and the overall water supplied throughout the season was ca. 400 mm. All the plants received insecticides and fertilization at the recommended dosages (Consultation Service, SAHAM, Israel Ministry of Agriculture).
2.4.3. Complementary Inoculation Method
The plants’ infection methodology and growth were according to [17,53]. The inoculation procedure includes further complementary phases to equalize and enhance the soil’s pathogen infection. To this end, the trial plots (excluding the negative check) were inoculated by adding 3 M. maydis-infected wheat grains to each corn seed with the seeding. These grains were prepared as detailed in Section 2.3.1 with a blend of M. maydis isolates (20 disks/150 g seeds), Hm2, Hm29, and Hm30 (see Table 1).
2.4.4. Trichoderma-Based Biocontrol Treatments
The bio-shield treatment was conducted by adding 3 sterilized wheat grains enriched with T. asperellum to each maize seed with the seeding. The wheat grain enrichment was conducted in a similar fashion to the pathogen enrichment described in Section 2.3.1.
2.4.5. Data Collection
Pollination occurred on 49 DAS (29 October 2020). During the season (up to the harvest), most plants had a healthy, vital appearance, typical growth, and crops (detailed in the Results) at the season’s ending. Yet, close examination of the lower stalk (first internode above the ground) and cobs’ spathes (the large bracts surrounding the cobs) revealed disease symptoms. Thus, the healthy (non-symptomatic) plant percentages were calculated and presented. Growth indexes (shoot height and wet weight, and the number of leaves) were determined in 5 plants (one repeat) selected randomly from the middle of every one of the experiment’s garden beds. Each experimental group (controls and biological intervention) included eight repetitions. Yield evaluation was conducted on all the plants in 4-7 rows (repeats) for the biological treatment and the controls. The molecular tracking (M. maydis DNA diagnosis) was performed on three arbitrarily selected representative plants in those rows.
2.5. Effect of Trichoderma Asperellum-Secreted Metabolites in Solid Media Cultures
The impact of T. asperellum-secreted metabolites on M. maydis, M. phaseolina, and F. verticillioides growth was assessed in solid growth media (PDA) as previously described [56]. To prepare the T. asperellum’s secreted metabolites, five mycelial discs (6 mm) were removed from the edges of 2–4-day-old colonies and incubated for six days in 150 mL PDB in the dark at 150 rpm at a temperature of 28 ± 1 °C. The growth medium was separated by Buchner funnel filtration through Whatman filter no. 3. The medium pH was adjusted to 5.1 ± 0.2 with NaOH and filtered again for sterilization using biofilter bottles (0.22-micron filter, BIOFIL 500 mL vacuum bottle filter, Indore, India). The Trichoderma cultures’ filtrate described above was used to prepare the PDA plates (instead of DDW). PDA powder was added to the filtrate according to the manufacturer’s instructions. After autoclave sterilization, the mixture was poured into Petri plates, which were seeded with a 6-mm-diameter M. maydis mycelial disc (from the margin of a fungus colony, see Section 2.2). The colonies were allowed to grow for six days and photographed.
2.6. 6-pentyl-α-pyrone Evaluation in a Net House
2.6.1. Application of the Purified T. asperellum Antifungal Compound(s) Using Seed Dressing
The T. asperellum isolate activity products were tested in a net house in pots during a full growth period. The experiment included six replicates (pots). The Megaton cv. corn (sweet, LWD super-susceptible) [41] was used in the experiments. Seed coating was performed according to a standard commercial procedure by blending the seeds with the compound (0.002 mL preparation/seed) until it was ultimately absorbed in the seeds. The inspected ingredient (30 or 60 μg 6-PP, and 100% or 50% T. asperellum crude, in 2 mL double-distilled water, DDW) was pipetted into a one-liter round-bottom glass flask containing 1000 seeds. The flask was rotated for ca. 10 min until the inspected ingredient was fully absorbed in the seeds. The seeds were taken out and dried in a fume hood on sterile paper towels for several hours. These seeds were kept in paper bags at 4 °C until use. The control treatment was conducted with similar seeds that underwent the addition of sterile water instead of the inspected ingredient.
2.6.2. Net House Full-Growth Season Pot Experiments Treatments and Sampling Program
The experiment was conducted in the R&D North Israel Experimental Farm located in the Hula Valley, Upper Galilee, northern Israel (33°09′08.2″ N 35°37′21.6″ E) during a full growing season. Each treatment consists of ten independent replicates (pots). The sprouts were thinned to one plant per pot towards the end of the sprouting growth stage (40 DAS). Inoculation with the M. maydis was carried out as described for the growth room experiment, with the following changes: in the first inoculation step, 40 g of infected sterilized wheat grains was added to the upper 20 cm of the soil of each pot. Control was peat soil from the experimental farm, having no record of LWD. If such an infestation existed, it was assumed to be very low. The evaluation of emergence (aboveground surface peek) percentages was performed 6 DAS. The male flowering was documented at 53 DAS. The growth parameters were evaluated at 40 DAS and the experiment end (96 DAS). Yield assessment and wilt determination were performed at harvest. Wilt valuation for each plant was based on 4 classifications—dead, diseased, symptoms, and healthy—as was demonstrated in [55], and the dry plants’ percentages were calculated and presented. On both sampling days, a qPCR detection of the fungal DNA in the plants’ roots or aboveground first internode was performed.
2.6.3. Net House Growth Conditions
The experiment’s local peat soil was similar to the growth room trial soil. It had no recognized record of LWD infestation (if it existed, it was assumed to be very low). To aerate the ground, the soil was blended with coarse perlite (No. 4) at a ratio of 2:1 in 10 L pots. The pots were positioned on concrete blocks to prevent the roots from contacting the ground and to better isolate the treatments. Each pot was sown with 5 seeds previously coated with thiram, captan, carboxin and metalaxyl-M. This seed treatment is standard general pesticide care. Seed samples were tested for vitality before seeding. Computerized irrigation was conducted with drip lines and was carried out from the seeding using 2 L per pot/day. According to the plants’ needs, adjustments were made in the water regime and pesticide treatments to prevent other diseases from influencing the plants during their growth (according to the Ministry of Agriculture’s recommended growing protocol). During the maize growing season, the average net house temperature was 24.7 °C, with a maximum of 43.4 °C and a minimum of 12.3 °C. The average humidity was 93.6%, with a maximum of 100% and a minimum of 42.2%.
2.7. Molecular Analysis
Quantifying M. maydis DNA in the host plant tissues was performed using the qPCR-based technique. The plants’ parts were washed vigorously with running tap water, then twice with sterile DDW and sliced into ca. 2 cm sections. The total weight of each repeat was adjusted to 0.7 g. The pathogen DNA was isolated and extracted according to a previously published protocol [66] with slight modifications [65]. Briefly, 0.7 g of plant tissue was ground with 4 mL of cetyltrimethyl ammonium bromide (CTAB). A total of 1.2 mL was transferred to Eppendorf test tubes and heated in a bath at 65 °C for 20 min. The test tubes were concentrated at 13,000 rpm at room temperature (24 °C) for 5 min, and the upper phase (700 µL) was transferred to new test tubes with the same volume of chloroform/isoamyl-alcohol (24: 1). The last cleaning stage was repeated twice. The upper phase (300 µL) was transferred to a new Eppendorf test tube with 200 µL of cold isopropanol, and the test tubes were maintained at 20 °C for 20–60 min. The test tubes were then concentrated at 13,000 rpm at 4 °C for 20 min, and the liquid was poured into a waste vessel. The DNA precipitate was resuspended and washed with 0.5 mL of ethanol. After a further concentration and ethanol wash cycle, the DNA precipitate in the test tubes was allowed to dry in a sterile hood overnight. Finally, the DNA was resuspended in 100 µL of ultra-pure water and kept at −20 °C until used for qPCR. DNA extraction from PDA colonies (positive control) was performed using a Plant/Fungi DNA Isolation Kit (Norgen, Biotek Corporation, Thorold, Ontario, Canada) according to the manufacturer’s instructions.
The molecular detection technique is based on a standard qPCR procedure to measure mRNA (converted to cDNA) levels [67]. It was adjusted here to detect M. maydis DNA using species-specific primers [68,69]. The A200a primers were applied for qPCR (Table 3). The housekeeping cytochrome C oxidase (COX) gene, encoding the cellular mitochondria’ last enzyme in the respiratory electron transport chain, served to normalize the relative M. maydis DNA amount in the samples examined [70]. For the COX gene COX F/R primer set, see Table 3. The relative DNA abundance was calculated using the ΔCt model [67,71]. All amplifications were carried out in four repetitions, and the same efficiency was assumed.
Table 3.
Primers for Magnaporthiopsis maydis detection 1.
All qPCR reactions were measured using the ABI PRISM 7900 HT Sequence Detection System (Applied Biosystems, Foster City, CA, USA) and 384-well plates. The reaction volume was 5 µL per well. All wells contained 0.25 μL of each primer (Forward/Backward at 10 μM concentration), 2.5 μL iTaq™ Universal SYBR Green Supermix solution (Bio-Rad Laboratories Ltd., Hercules, California, USA), and 2 µL of DNA sample. Reaction conditions were 95 °C for 60 s (pre-cycle activation phase), then 40 cycles of 95 °C for 15 s (denaturation), 60 °C for 30 s (annealing and extension), and, finally, the formation of a melting curve.
2.8. Statistical Analysis
The growth room and net house experiment pots and the field plots were scattered in a fully randomized design and analyzed using the same statistical method. Statistical processing and data evaluation were achieved using JMP software, version 15 (SAS Institute Inc., Cary, NC, USA). The results were evaluated using the one-way analysis of variance (ANOVA) followed by a Student’s t-test post hoc (without correction for multiple tests) at a significance level of p < 0.05. Due to the difficulty in creating uniform plant infection, high variability in M. maydis’s DNA levels and high standard error values are expected, making it challenging to obtain the alternative hypothesis.
3. Results
The current study aimed at evaluating the Trichoderma asperellum secreted metabolite, 6-pentyl-α-pyrone (6-PP), against the maize late wilt (LWD) pathogen, Magnaporthiopsis maydis. To achieve this goal, a series of trials were conducted, starting by inspecting the addition of T. asperellum directly to seeds with the sowing (sprouts in a growth room-controlled conditions and complete season field experiment). These trials were followed by a targeted evaluation of the T. asperellum recently purified and identified active secreted metabolite, 6-PP (net house experiment over an entire growing season). The powerful antifungal compound was applied in seed coating and tested against the T. asperellum growth medium crude or by adding the fungus itself with the seeding. The results indicate a potential new control treatment to protect susceptible maize plants from LWD, as detailed below.
3.1. Trichoderma Asperellum Pathogenicity Evaluation in Growth Room Sprouts
Young T. asperellum hyphae disks were cut from the margins of culture colonies, grown on rich medium plates, and added directly to the seeds with the sowing. Inspecting this procedure’s impact on seedling development (up to 42 days) and M. maydis establishment in their roots revealed a significant growth promotion and LWD recovery (Table 4). While the T. asperellum did not affect the aboveground emergence (at day 8), at the end of the sprouting phase (day 42, the fifth leaf stage), all growth indexes improved drastically (p < 0.05) compared to the infected, untreated plants. This improvement ranges from 1.8-fold in plant height to 2.7 and 3.3-fold in shoot and root fresh weight. These changes were accompanied by a two-fold reduction in the M. maydis DNA in the sprouts’ roots.
Table 4.
Trichoderma asperellum late wilt control in sprouts at 42 DAS 1.
3.2. Trichoderma Asperellum Pathogenicity Evaluation in the Field
The follow-up full-season trial was conducted in an experimental field known to have a mild infection with late wilt. Since the growing season’s climatic conditions were not optimal for this disease development (relatively rainy autumn season), we infected the field soil with the seeding. Still, during the growth period up to the harvest, the plants’ phenological development and growth indexes were normal, and the disease symptoms were minor. These parameters did not vary statistically between the treatments (Table 5).
Table 5.
Trichoderma asperellum late wilt control in the field at 41 (A) and 82 (B) DAS 1.
Only a close examination of the lower stem and cob spathes (the large bracts surrounding the cobs) revealed the disease dehydration symptoms and the impact of the T. asperellum treatment (Figure 2A). At the season’s end (day 82, 33 days after fertilization), the improvement in these two parameters (lower stem and cob symptoms) was 1.6- and 1.3-fold, respectively, in the biological treatment compared to the non-treated control (without statistical significance). In line with these results is the M. maydis’s DNA in the plants’ first aboveground internode (Figure 2B, qPCR results). In the T. asperellum treatment, the pathogen DNA dropped by five-fold compared to the control. This being said, the pathogen DNA levels in all treatments were relatively low and near the technique’s lower threshold (the qPCR method can detect variation in proportionate M. maydis specific DNA between 10 to 10−5 [7]).
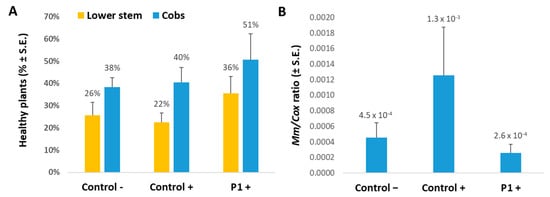
Figure 2.
Plants’ health and M. maydis infection severity in the field at day 82. Maize late-wilt disease symptoms (A) and qPCR diagnosis of M. maydis DNA in maize plants (B) at the end of the field experiment (33 days after fertilization) conducted in the summer and autumn of 2020. The experiment was conducted with the late-wilt-susceptible maize genotype Prelude. Control− are plots without complementary inoculation. Control+ are plots with M. maydis infection. The other treatments are infected plots treated with T. asperellum (P1) by adding it to the seeds with sowing. M. maydis relative DNA (Mm) normalized to the cytochrome C oxidase (Cox) DNA was evaluated in the plants’ first aboveground internode. Vertical upper bars signify the standard error of the mean of 4-10 replicates.
3.3. 6-pentyl-α-pyrone Evaluation in a Net House
So far, we have evaluated the addition of T. asperellum directly to the seeds with sowing. The following steps were taken to inspect its secreted metabolites’ bio-protective ability under late wilt stress. The potential of this Trichoderma species growth medium crude and its active ingredient 6-PP to restrict the LWD pathogen, M. maydis, in vitro was previously demonstrated [56]. It now appears that T. asperellum growth medium exhibits specificity activity against M. maydis and cannot repress two additional members of the maize post-flowering stalk rot complex [73], Fusarium verticillioides and Macrophomina phaseolina (Figure 3).
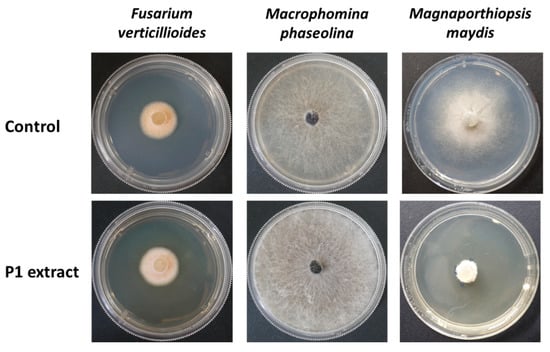
Figure 3.
Bioassay of the Trichoderma asperellum (P1) secreted metabolites. The fungus liquid growth medium (P extract) was used to prepare a rich growth medium (PDA). The plates were used to test the maize post-flowering stalk rot complex [73]. This fungal group includes M. maydis, Macrophomina phaseolina, and Fusarium verticillioides. The controls are regular water-based PDAs. Representative plates’ photos were taken after six days.
Until now, the T. asperellum secreted ingredients had never been tested as a shield against the LWD agent in plants. To accomplish this, we conducted a net house whole-season experiment. We tested the purified 6-PP metabolite against the growth medium crude and the direct addition of the Trichoderma to the seeds (Figure 4). The disease progression in this experiment was evident, and already at the end of the sprouting phase (day 40), significant growth reduction (p < 0.05) was observed in the infected, non-protected plants group (Table 6).

Figure 4.
Photos of the net house experiment, which was performed in late wilt-susceptible Megaton cv. The male flowering was documented at 53 DAS. Disease symptoms were already clearly visible from day 62 onwards.
Table 6.
6-pentyl-α-pyrone late wilt control in the net house at the sprouting phase 1.
At the V5 (five leaves) stage, the unprotected plants suffer a 37% aboveground peak decrease (6 DAS), a 64% lower survival rate, and 38–39% growth repression (40 DAS, Table 6). All the bio-treatments led to some recovery and improvement in these parameters, especially the 6-PP (30 µg), the crude (diluted to 50%), and the T. asperellum addition to the seeds. High dosages either of the clean 6-PP metabolite or the growth medium crude were apparently less effective, perhaps due to overdose, which can lead to phytotoxicity.
In a final evaluation made at the season’s end (day 96, 43 days post fertilization), the tendencies that started to be revealed on day 40 were enhanced and became significant (p < 0.05, Figure 5). Here, the bioprotective 30 µg 6-PP and 50% crude extract had similar (and even higher) results compared to the healthy plants’ control, while the non-treated group had up to 123% and 64% lower shoot and cob weights, respectively. Parallel plant health increases accompanied these dramatic growth changes (Figure 6).
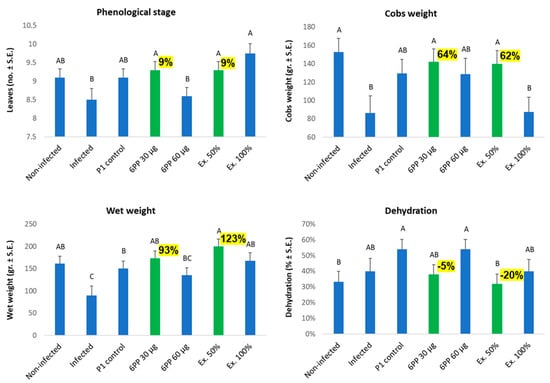
Figure 5.
The Trichoderma asperellum secreted metabolite, 6-pentyl-α-pyrone, impacts maize development under late wilt disease stress. Net house study, day 96 data. The purified active ingredient 6-PP was tested as seed coating at 30 and 60 µg/seed and compared to the T. asperellum (P1) direct addition to seeds or its crude secreted metabolites applied at 50% or 100% concentrations (Ex. 50% and Ex. 100%). The standard error of the mean of 7–10 replicates (plants) is shown. Highlighted values (in yellow) are important differences from the infected, untreated control. Statistically significant differences (p < 0.05) between treatments are specified by different letters (A–C).

Figure 6.
Net house representative plant photos on day 96. The experiment’s treatments and controls are depicted in Figure 5.
The dehydration levels of the plants were attributed to variations in the M. maydis DNA load tracked by the molecular qPCR method. As revealed by the highly sensitive qPCR method, some very low levels of M. maydis DNA (near the method’s lower sensitivity threshold) could be identified in the control treatment. However, such low levels were unable to evoke disease symptoms. High pathogen DNA was measured (at days 40 and 96) in the infected control plants group and the plants enriched with T. asperellum on the sowing day (Figure 7). This last result is unexpected since the T. asperellum addition to the seeds provides some protection against LWD growth reduction at the sprouting phase (40 DAS, Table 6). In contrast, the other treatments’ parameters fit well with the growth and health results. At harvest (day 96), the crude extract (especially the 50% dilution) had the best influence in repressing the pathogen (43-fold). Here, too, the 6-PP diluted (30 µg) seed coating was more efficient (17-fold) than the 6-PP 60 µg treatment (7-fold M. maydis decrease).
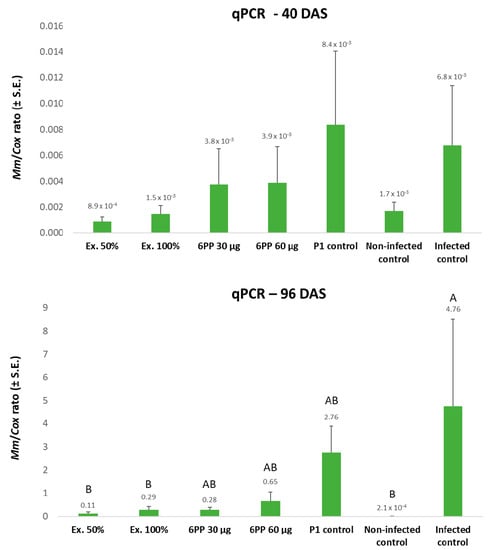
Figure 7.
qPCR estimate of M. maydis DNA on days 40 and 96 of the net house experiment. The relative amount of M. maydis DNA (Mm) normalized to the cytochrome C oxidase (Cox) DNA was assessed in the plants’ first aboveground internode. The experiment’s treatments and controls are depicted in Figure 5. Error lines represent the standard error of an average of 7–10 repetitions (plants). If they exist, statistically significant differences (p < 0.05) between treatments are specified by different letters (A–B).
4. Discussion
Natural microorganisms’ communities inhabiting plants include non-pathogenic members that can have shielding effects against pathogens. This work examined a new biological control agent candidate against the maize late wilt disease (LWD) pathogen, Magnaporthiopsis maydis. This pathogen control is challenging due to long-term soil survival, and resistant cultivars are constantly being threatened by aggressive pathogen variants [24].
In recent years, biopesticides have achieved significant consideration as an alternative to environmentally problematic chemical pesticides [54]. Understanding the roles of endophytic community assemblages and their impact on plant health would be a major contribution to sustainable integrated agriculture. Trichoderma species are used widely in agriculture to promote plant growth and control pathogens attacking crops, serving as valuable tools for sustainable food production [74]. Still, the action mechanism of this bio-protective species is poorly understood.
The maize-biofriendly species, T. asperellum, is an endophyte isolated in Israel from LWD-susceptible maize genotype seeds [58]. Here, this Trichoderma species showed protective ability against Magnaporthiopsis maydis in growth room sprouts and mature plants in the field. However, it is inferred from the results that this protection was affected most at the sprouting phase and that under some field conditions, it may fail. Obvious differences exist between the growth room and the field conditions that may explain the results presented here. As a start, the field’s unstable conditions may have a significant impact on the results. In our study, the field conditions were not optimal for LWD development, leading to weak dehydration symptoms appearing at the harvest. Indeed, this growing season was rainier and colder, and these circumstances were possibly the reason for lowered yields and a minor LWD outbreak. To support this, early (spring) sowing of corn in Egypt [75] and late summer planting in India [27] reduced LWD. It is possible that the full potential of the T. asperellum-based biological control was not manifested under these conditions.
Inspecting similar research results published recently by Estévez-Geffriaud (2021) [76] provides evidence that T. asperellum (strain T34 applied as seed treatment) positively affected maize productivity and health under late wilt disease stress. Regardless of the water regime, T34 improved kernel P and C content, kernel dry weight, and number. Under drought, this Trichoderma treatment improved leaf relative water content, water use efficiency, PSII maximum efficiency, and photosynthesis. Moreover, T34 reduced LWD incidence by up to 13% and enhanced the yield per plant in LWD moderate tolerant and resistant maize genotypes. Still, no yield improvement was measured in susceptible lines [76]. It may be inferred from Estévez-Geffriaud and other studies (for example, [33]) that the Trichoderma species, the specific strain, the plant’s LWD susceptibility degree, and environmental conditions could all affect the success of the biological treatment.
It should also be considered that T. asperellum protection is mainly affected during the sprouting stage, which is regarded as the pathogen entry and establishment phase [21,23]. Later in the season, this initial protection may lose effectiveness if the pathogen finds a way to establish itself in the plant’s vascular system and disrupt its development. If such a scenario is the case, integrated biological and chemical protection may assist in overcoming the pathogen’s establishment and provide us with the benefit of low fungicides usage with a live protective barrier [54].
Many other factors could affect this outcome (the differences between the growth room and field assays). For example, the time lapse between pathogen and antagonist inoculation can be crucial. The preventive and curative abilities of T. asperellum UDEAGIEM-H01 versus Fusarium oxysporum were evaluated in stevia plants (Stevia rebaudiana) under field conditions. The occurrence of Fusarium wilt was 10% in the protective treatment when the antagonist was inoculated six days prior to the pathogen, whereas 70% of disease occurrence was recorded in plants treated with the antagonist six days after the infection of seedlings with the pathogen [77]. Thus, controlled conditions experiments, as demonstrated here, are difficult to replicate on a field scale where pathogens are already present [74]. Further studies are needed to optimize the maize growing time at which T. asperellum could present the best LWD biocontrol abilities in the field.
One of the advances in using Trichoderma-based protection against LWD is that these species likely maintain a symbiotic lifestyle inside the host plant organs that may provide a season-long defense. Moreover, the Trichoderma species can trigger the maize disease resistance response systemically through the biosynthetic induction of brassinolide, jasmonic acid, salicylic acid, defense enzymes, and reactive oxygen species [78]. T. asperellum substantially impeded F. verticillioides growth in plate competition assay [79] and showed a biocontrol efficacy of 50% against Fusarium graminearum in the field [80]. These examples suggest that the field control application of T. asperellum may offer broad pathogen protection.
Yet, it should be considered that in severe cases, Trichoderma protection alone is insufficient to restrain the disease. When the pressure of infection is high, systemic fungicides must be applied. Trichoderma-based intervention should be part of an integrated management strategy, considering that some Trichoderma spp. are inefficient in combination with fungicides [54]. Compared to chemical pesticides, bio-based disease control does not generate resistance in pathogens, does not contaminate the environment, prevents the proliferation of secondary plagues, is compatible with organic production, and meets the requirements of profitable markets concerning maximum limits of chemical residues on fruits and vegetables. This work and others [55,56] suggest that T. asperellum bio-treatment against M. maydis is worth continuous future studies to maximize its potential. The findings presented in the current work imply that a major part of this antifungal activity is related to T. asperellum secreted metabolites in general and the 6-pentyl-α-pyrone (6-PP) active ingredient in particular.
Fungi are incredible producers of a huge range of xenobiotics, including some well-known compounds that inhibit the growth of other fungi. Secondary metabolites are pivotal defensive microorganism–pathogen interactions during host invasion and establishment, as well as the plant microbiome defense response. These include antifungals, mycotoxins, lactams/lactones, and other extracellular components. Such a compound is 6-PP [81]. This metabolite was recognized as one of the major bioactive compounds of various Trichoderma species [62], with a clear correlation between 6-PP biosynthesis and biocontrol capability. 6-PP inhibits spore germination, mycelial growth, and pigmentation of tested phytopathogenic fungi [63]. 6-PP also decreases the production of the mycotoxin deoxynivalenol (DON) by F. graminearum and fusaric acid by F. moniliforme [64]. Our study results support the importance of this molecule and are opening up a new research direction that could lead to a new LWD bio-pesticide product.
Significant knowledge gaps still exist regarding the action mechanism of 6-PP. The compound is oxygenated and thus was proposed to be part of the oxylipin family [82,83]. This suggests that regardless of their source, similar oxylipin-like structure compounds may provoke strong chemotropic reactions in Trichoderma—and possibly additional fungal species. Thus they may establish an interspecies signaling mechanism. In mycoparasitism, it was implied [84] that the plant oxylipin recruits the mycoparasite towards roots affected by fungal phytopathogens—or already as a preventive measure. At the same time, the fungal oxylipin 6-PP induces morphological adjustments in Trichoderma needed to launch a localized mycoparasitic attack.
The 6-PP mass release at this stage would not only have a repressive antifungal action. It might also act as a highly secreted signal in the regions where the colony contacts the prey fungus to promote hyphal redirection and hyphal growth as part of the mycoparasitism response. Other secondary metabolites produced by T. asperellum are likely involved in these events to refine and support the chemoattraction process and the mycoparasitism attack [84].
5. Conclusions
This research targeted the maize late wilt disease (LWD) agent, Magnaporthiopsis maydis, using the biocontrol Trichoderma asperellum and its secreted metabolites. Growth room sprouts assay and field trials that were less effective demonstrated that adding this Trichoderma species directly to seeds can provide some protection against the disease. The bio-friendly Trichoderma control is most likely attributed to this organism’s secreted metabolites since applying its growth medium extract in the seed coating provided the plants with significant protection. One of these ingredients is 6-pentyl-α-pyrone (6-PP), a potent antifungal compound tested here for the first time against M. maydis in infected plants. The success of the 6-PP seed coating was expressed not only in a drastic reduction in the pathogen’s lower stalk infection but also in the nearly full recovery of the plant growth indices. A follow-up study should implement the 6-PP seed coating in commercial field simulations. Moreover, it would be most interesting and important to identify other active ingredients in the T. asperellum and other beneficial Trichoderma spp. (such as T. longibrachiatum) growing medium extract.
Author Contributions
Conceptualization, O.D. and A.G.; data curation, O.D. and A.G.; formal analysis, O.D. and A.G.; funding acquisition, O.D.; investigation, O.D. and A.G.; methodology, O.D. and A.G.; project administration, O.D.; resources, O.D.; supervision, O.D.; validation, O.D. and A.G.; visualization, O.D. and A.G.; writing (original draft), O.D.; writing (review and editing), O.D. and A.G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a research grant, number 21-07-0004, from the Israel Ministry of Agriculture and Rural Development, a research grant from the Jewish Charitable Association (ICA)—Galilee Research Institute (Migal) accelerator, Israel (2020) and a research grant from the Galilee Research Institute (Migal) internal grant (2021).
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We would like to thank Soliman Khatib and Assaf Chen (Migal—Galilee Research Institute and Tel-Hai College, Israel) for their essential support and advice, and Shlomit Dor, Paz Becher and Galia Shofman (Migal—Galilee Research Institute and Tel-Hai College, Israel) for their technical assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Elshahawy, I.E.; Khattab, A.E.-N.A. Endophyte Chaetomium globosum improves the growth of maize plants and induces their resistance to late wilt disease. J. Plant Dis. Prot. 2022, 129, 1125–1144. [Google Scholar] [CrossRef]
- El-Shenawy, A.A.; Hassan, M.A.A.; El-Haress, S.M.A.; Abd-Elaziz, M.A.A. Assessment of Combining Ability in Some Newly Maize Inbred Lines for Grain Yield and Late Wilt Resistance. J. Plant Prod. 2022, 13, 45–48. [Google Scholar] [CrossRef]
- Degani, O.; Chen, A.; Dor, S.; Orlov-Levin, V.; Jacob, M.; Shoshani, G.; Rabinovitz, O. Remote evaluation of maize cultivars susceptibility to late wilt disease caused by Magnaporthiopsis maydis. J. Plant Pathol. 2022, 104, 509–525. [Google Scholar] [CrossRef]
- Patanita, M.; Campos, M.D.; Félix, M.D.R.; Carvalho, M.; Brito, I. Effect of Tillage System and Cover Crop on Maize Mycorrhization and Presence of Magnaporthiopsis maydis. Biology 2020, 9, 46. [Google Scholar] [CrossRef]
- Ortiz-Bustos, C.M.; López-Bernal, A.; Testi, L.; Molinero-Ruiz, L. Environmental and irrigation conditions can mask the effect of Magnaporthiopsis maydis on growth and productivity of maize. Plant Pathol. 2019, 68, 1555–1564. [Google Scholar] [CrossRef]
- Rakesh, B.; Gangappa, E.; Gandhi, S.; Gowda, R.P.V.; Swamy, S.D.; Ramesh, S.; Hemareddy, H.B. Mapping genomic regions controlling resistance to late wilt disease caused by Harpophora maydis in maize (Zea mays L.). Euphytica 2022, 218, 101. [Google Scholar] [CrossRef]
- Degani, O. A Review: Late Wilt of Maize—The Pathogen, the Disease, Current Status, and Future Perspective. J. Fungi 2021, 7, 989. [Google Scholar] [CrossRef] [PubMed]
- Johal, L.; Huber, D.M.; Martyn, R. Late wilt of corn (maize) pathway analysis: Intentional introduction of Cephalosporium maydis. In Pathways Analysis for the Introduction to the U.S. of Plant Pathogens of Economic Importance; Purdue University: West Lafayette, IN, USA, 2004. [Google Scholar]
- Samra, A.S.; Sabet, K.A.; Hingorani, M.K. Late wilt disease of maize caused by Cephalosporium maydis. Phytopathology 1963, 53, 402–406. [Google Scholar]
- Molinero-Ruiz, M.L.; Melero-Vara, J.M.; Mateos, A. Cephalosporium maydis, the cause of late wilt in maize, a pathogen new to Portugal and Spain. Plant Dis. 2010, 94, 379. [Google Scholar] [CrossRef]
- Klaubauf, S.; Tharreau, D.; Fournier, E.; Groenewald, J.; Crous, P.; de Vries, R.; Lebrun, M.-H. Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol. 2014, 79, 85–120. [Google Scholar] [CrossRef]
- Hernandez-Restrepo, M.; Groenewald, J.; Elliott, M.; Canning, G.; McMillan, V.; Crous, P.W. Take-all or nothing. Stud. Mycol. 2016, 83, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-B.; Hyde, K.D.; Yang, E.-F.; Kakumyan, P.; Bahkali, A.H.; Elgorban, A.M.; Karunarathna, S.C.; Phookamsak, R.; Lumyong, S. Morphological and phylogenetic appraisal of Ophioceras (Ophioceraceae, Magnaporthales). PLoS ONE 2021, 16, e0253853. [Google Scholar] [CrossRef] [PubMed]
- Sabet, K.A.; Samra, A.S.; Mansour, I.M. Saprophytic behaviour of Cephalosporium maydis and C. acremonium. Ann. Appl. Biol. 1970, 66, 265–271. [Google Scholar] [CrossRef]
- Michail, S.H.; Abou-Elseoud, M.S.; Nour Eldin, M.S. Seed health testing of corn for Cephalosporium maydis. Acta Phytopathol. Et Entomol. Hung. 1999, 34, 35–42. [Google Scholar]
- Sahab, A.F.; Osman, A.R.; Soleman, N.K.; Mikhail, M.S. Studies on root-rot of lupin in Egypt and its control. Egypt. J. Phytopathol. 1985, 17, 23–35. [Google Scholar]
- Degani, O.; Dor, S.; Abraham, D.; Cohen, R. Interactions between Magnaporthiopsis maydis and Macrophomina phaseolina, the causes of wilt diseases in maize and cotton. Microorganisms 2020, 8, 249. [Google Scholar] [CrossRef]
- Dor, S.; Degani, O. Uncovering the Host Range for Maize Pathogen Magnaporthiopsis maydis. Plants 2019, 8, 259. [Google Scholar] [CrossRef]
- Sabet, K.A.; Sanra, A.S.; Mansour, I.S. Interaction between Fusarium oxysporum F. vasinfectum and Cephalosporium maydis on cotton and maize. Ann. Appl. Biol. 1966, 58, 93–101. [Google Scholar] [CrossRef]
- Degani, O.; Becher, P.; Gordani, A. Pathogenic interactions between Macrophomina phaseolina and Magnaporthiopsis maydis in mutually infected cotton sprouts. Agriculture 2022, 12, 255. [Google Scholar] [CrossRef]
- El-Shafey, H.A.; Claflin, L.E. (Eds.) Late Wilt; APS Press: St. Paul, MN, USA, 1999; pp. 43–44. [Google Scholar]
- Agag, S.H.; Sabry, A.M.; El-Samman, M.G.; Mostafa, M.H. Pathological and Molecular Characterization of Magnaporthiopsis maydis Isolates Causing Late Wilt in Maize. Egypt. J. Phytopathol. 2021, 49, 1–9. [Google Scholar] [CrossRef]
- Sabet, K.A.; Samra, A.S.; Mansour, I.M.; Zaher, A.M. Pathogenic behaviour of Cephalosporium maydis and C. acremonium. Ann. Appl. Biol. 1970, 66, 257–263. [Google Scholar] [CrossRef]
- Degani, O. Control Strategies to Cope with Late Wilt of Maize. Pathogens 2022, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- El-Gremi, S.M.A.; Belal, E.B.A.; Ghazy, N.A. Cephalosporium maydis as affected by maize root exudates and role of the fungal metabolites in pathogenesis. J. Agric. Chem. Biotechnol. 2007, 32, 7605–7615. [Google Scholar] [CrossRef]
- Abd El-Rahim, M.F.; Fahmy, G.M.; Fahmy, Z.M. Alterations in transpiration and stem vascular tissues of two maize cultivars under conditions of water stress and late wilt disease. Plant Pathol. 1998, 47, 216–223. [Google Scholar] [CrossRef]
- Singh, S.D.; Siradhana, B.S. Date of sowing in relation to late wilt disease of maize. Indian Phytopathol. 1988, 41, 489–491. [Google Scholar]
- Samra, A.; Sabet, K.; Kamel, M.; Abd El-Rahim, M. Further studies on the effect of field conditions and cultural practices on infection with stalk-rot complex of maize. Arab Republic of Egypt. Plant Prot. Dept. Bull. 1971. Available online: http://library.wur.nl/webquery/clc/385177 (accessed on 25 August 2022).
- Payak, M.; Sharma, R. Research on Diseases of Maize; Indian Council of Agricultural Research: New Delhi, India, 1978; p. 228. [Google Scholar]
- Samra, A.S.; Sabet, K.A.; Abdel-Rahim, M.F. Effect of Soil Conditions and Cultural Practices on Infection with Stalk Rots; U.A.R. Ministry of Agric. Government Printing Offices: Cairo, Egypt, 1966; pp. 117–164. [Google Scholar]
- Singh, S.D.; Siradhana, B.S. Effect of macro and micronutrients on the development of late wilt of maize induced by Cephalosporium maydis. Summa Phytopath 1990, 16, 140–145. [Google Scholar]
- Mosa, H.; Motawei, A.; El-Aal, A.A. Nitrogen fertilization influence on combining ability for grain yield and resistance to late wilt disease in maize. J. Agric. Res. Kafrelsheikh Univ. 2010, 36, 278–291. [Google Scholar]
- Elshahawy, I.E.; El-Sayed, A.E.-K.B. Maximizing the efficacy of Trichoderma to control Cephalosporium maydis, causing maize late wilt disease, using freshwater microalgae extracts. Egypt. J. Biol. Pest Control 2018, 28, 48. [Google Scholar] [CrossRef]
- Doleib, N.M.; Farfour, S.A.; Al-shakankery, F.M.; Ammar, M.; Hamouda, R.A. Antifungal activates of cyanobacteria and some marine algae against Cephalosporium maydis, the cause of maize late wilt disease in vitro. Biosci. Res. 2021, 18, 536–543. [Google Scholar]
- El-Shabrawy, E.-S.; Shehata, H. Controlling maize late-wilt and enhancing plant salinity tolerance by some rhizobacterial strains. Egypt. J. Phytopathol. 2018, 46, 235–255. [Google Scholar] [CrossRef]
- Fayzalla, E.; Sadik, E.; Elwakil, M.; Gomah, A. Soil solarization for controlling Cephalosporium maydis, the cause of late wilt disease of maize in Egypt. Egypt J. Phytopathol. 1994, 22, 171–178. [Google Scholar]
- Tej, R.; Rodríguez-Mallol, C.; Rodríguez-Arcos, R.; Karray-Bouraoui, N.; Molinero-Ruiz, L. Inhibitory effect of Lycium europaeum extracts on phytopathogenic soil-borne fungi and the reduction of late wilt in maize. Eur. J. Plant Pathol. 2018, 152, 249–265. [Google Scholar] [CrossRef]
- Elshahawy, I.E.; El-Wahed, M.S.A. Suppression of Cephalosporium maydis by the resistance inducer beta-sitosterol. Eur. J. Plant Pathol. 2022, 163, 673–693. [Google Scholar] [CrossRef]
- Abd-el-Rahim, M.F.; Sabet, K.A.; El-Shafey, H.A.; El-Assiuty, E.M. Chemical control of the late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. 1982, 60, 31–49. [Google Scholar]
- Singh, S.D.; Siradhana, B.S. Chemical control of late wilt of maize induced by Cephalosporium maydis. Indian J. Mycol. Pl. Path. 1989, 19, 121–122. [Google Scholar]
- Degani, O.; Gordani, A.; Becher, P.; Chen, A.; Rabinovitz, O. Crop rotation and minimal tillage selectively affect maize growth promotion under late wilt disease stress. J. Fungi 2022, 8, 586. [Google Scholar] [CrossRef]
- Degani, O.; Gordani, A.; Becher, P.; Dor, S. Crop cycle and tillage role in the outbreak of late wilt disease of maize caused by Magnaporthiopsis maydis. J. Fungi 2021, 7, 706. [Google Scholar] [CrossRef]
- Hassan, E.O.; Shoala, T.; Attia, A.M.F.; Badr, O.A.M.; Mahmoud, S.Y.M.; Farrag, E.S.H.; EL-Fiki, I.A.I. Chitosan and nano-chitosan for management of Harpophora maydis: Approaches for investigating antifungal activity, pathogenicity, maize-resistant lines, and molecular diagnosis of plant infection. J. Fungi 2022, 8, 509. [Google Scholar] [CrossRef]
- El-Shabrawy, E. Use silica nanoparticles in controlling late wilt disease in maize caused by Harpophora maydis. Egypt. J. Appl. Sci. 2021, 36, 1–19. [Google Scholar] [CrossRef]
- El-Gazzar, N.; El-Bakery, A.M.; Ata, A.A. Influence of some bioagents and chitosan nanoparticles on controlling maize late wilt and improving plants characteristics. Egypt. J. Phytopathol. 2018, 46, 243–264. [Google Scholar] [CrossRef]
- Sunitha, N.C.; Gangappa, E.; Gowda, R.P.V.; Ramesh, S.; Biradar, S.; Swamy, D.; Hemareddy, H.B. Discovery of genomic regions associated with resistance to late wilt disease caused by Harpophora maydis (Samra, Sabet and Hing) in maize (Zea mays L.). J. Appl. Genet. 2021, 63, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.A.; Gangappa, E.; Swamy, S.R.A.D.S. Identification of high yielding inbred lines resistant to late wilt disease caused by Harpophora maydis in maize. Electron. J. Plant Breed. 2021, 12, 151–158. [Google Scholar] [CrossRef]
- Gazala, P.; Gangappa, E.; Ramesh, S.; Swamy, D. Comparative Breeding potential of two crosses for response to late wilt disease (LWD) in maize (Zea mays L.). Genet. Resour. Crop Evol. 2021, 68, 1751–1757. [Google Scholar] [CrossRef]
- Kamara, M.; Ghazy, N.; Mansour, E.; Elsharkawy, M.; Kheir, A.; Ibrahim, K. Molecular genetic diversity and line × tester analysis for resistance to late wilt disease and grain yield in maize. Agronomy 2021, 11, 898. [Google Scholar] [CrossRef]
- Ortiz-Bustos, C.M.; Testi, L.; García-Carneros, A.B.; Molinero-Ruiz, L. Geographic distribution and aggressiveness of Harpophora maydis in the Iberian peninsula, and thermal detection of maize late wilt. Eur. J. Plant Pathol. 2015, 144, 383–397. [Google Scholar] [CrossRef]
- Zeller, K.A.; Ismael, A.-S.M.; El-Assiuty, E.M.; Fahmy, Z.M.; Bekheet, F.M.; Leslie, J.F. Relative Competitiveness and Virulence of Four Clonal Lineages of Cephalosporium maydis from Egypt Toward Greenhouse-Grown Maize. Plant Dis. 2002, 86, 373–378. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S.; Movshowitz, D.; Fraidman, E.; Rabinovitz, O.; Graph, S. Effective chemical protection against the maize late wilt causal agent, Harpophora maydis, in the field. PLoS ONE 2018, 13, e0208353. [Google Scholar] [CrossRef]
- Degani, O.; Movshowitz, D.; Dor, S.; Meerson, A.; Goldblat, Y.; Rabinovitz, O. Evaluating Azoxystrobin seed coating against maize late wilt disease using a sensitive qPCR-based method. Plant Dis. 2019, 103, 238–248. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining biocontrol agents with chemical fungicides for integrated plant fungal disease control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef]
- Degani, O.; Rabinovitz, O.; Becher, P.; Gordani, A.; Chen, A. Trichoderma longibrachiatum and Trichoderma asperellum confer growth promotion and protection against late wilt disease in the field. J. Fungi 2021, 7, 444. [Google Scholar] [CrossRef]
- Degani, O.; Khatib, S.; Becher, P.; Gordani, A.; Harris, R. Trichoderma asperellum secreted 6-Pentyl-α-Pyrone to control magnaporthiopsis maydis, the maize late wilt disease agent. Biology 2021, 10, 897. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Dor, S. Trichoderma biological control to protect sensitive maize hybrids against late wilt disease in the field. J. Fungi 2021, 7, 315. [Google Scholar] [CrossRef] [PubMed]
- Degani, O.; Danielle, R.; Dor, S. The microflora of maize grains as a biological barrier against the late wilt causal agent, Magnaporthiopsis maydis. Agronomy 2021, 11, 965. [Google Scholar] [CrossRef]
- Ghazy, N.; El-Nahrawy, S. Siderophore production by Bacillus subtilis MF497446 and Pseudomonas koreensis MG209738 and their efficacy in controlling Cephalosporium maydis in maize plant. Arch. Microbiol. 2020, 203, 1195–1209. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—opportunistic, avirulent plant symbionts. Nat. Rev. Genet. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Harman, G.E. Overview of mechanisms and uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef]
- Hamrouni, R.; Molinet, J.; Dupuy, N.; Taieb, N.; Carboue, Q.; Masmoudi, A.; Roussos, S. The Effect of aeration for 6-Pentyl-alpha-pyrone, conidia and lytic enzymes production by Trichoderma asperellum strains grown in solid-state fermentation. Waste Biomass-Valorization 2019, 11, 5711–5720. [Google Scholar] [CrossRef]
- Salwan, R.; Rialch, N.; Sharma, V. Bioactive volatile metabolites of Trichoderma: An overview. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; 2019; pp. 87–111. Available online: https://link.springer.com/chapter/10.1007/978-981-13-5862-3_5 (accessed on 25 August 2022).
- Stracquadanio, C.; Quiles, J.M.; Meca, G.; Cacciola, S.O. Antifungal activity of bioactive metabolites produced by Trichoderma asperellum and Trichoderma atroviride in liquid medium. J. Fungi 2020, 6, 263. [Google Scholar] [CrossRef]
- Degani, O.; Dor, S.; Movshovitz, D.; Rabinovitz, O. Methods for studying Magnaporthiopsis maydis, the maize late wilt causal agent. Agronomy 2019, 9, 181. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.A.; Leslie, J.F. Cephalosporium maydis is a distinct species in the Gaeumannomyces-Harpophora species complex. Mycologia 2004, 96, 1294–1305. [Google Scholar] [CrossRef] [PubMed]
- Zeller, K.A.; Jurgenson, J.E.; El-Assiuty, E.M.; Leslie, J.F. Isozyme and amplified fragment length polymorphisms from Cephalosporium maydis in Egypt. Phytoparasitica 2000, 28, 121–130. [Google Scholar] [CrossRef]
- Weller, S.; Elphinstone, J.; Smith, N.; Boonham, N.; Stead, D. Detection of Ralstonia solanacearumstrains with a quantitative, multiplex, real-time, fluorogenic PCR (TaqMan) assay. Appl. Environ. Microbiol. 2000, 66, 2853–2858. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N., Jr. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef]
- Li, W.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identification of Candidatus liberibacter species associated with Citrus huanglongbing. J. Microbiol. Methods 2006, 66, 104–115. [Google Scholar] [CrossRef]
- Khokhar, M.K.; Hooda, K.S.; Sharma, S.S.; Singh, V. Post flowering stalk rot complex of maize-Present status and future prospects. Maydica 2014, 59, 226–242. [Google Scholar]
- Ferreira, F.V.; Musumeci, M.A. Trichoderma as biological control agent: Scope and prospects to improve efficacy. World J. Microbiol. Biotechnol. 2021, 37, 90. [Google Scholar] [CrossRef]
- El-Shafey, H.A.; El-Shorbagy, F.A.; Khalil, I.I.; El-Assiuty, E.M. Additional sources of resistance to the late-wilt disease of maize caused by Cephalosporium maydis. Agric. Res. Rev. Egypt. 1988, 66, 221–230. [Google Scholar]
- Estévez Geffriaud, V. Seed treatments for the protection of crops. Ph.D. Thesis, Universitat de Barcelona, 2021; 148p. Available online: https://www.tdx.cat/handle/10803/672956.
- Díaz-Gutiérrez, C.; Arroyave, C.; Llugany, M.; Poschenrieder, C.; Martos, S.; Peláez, C. Trichoderma asperellum as a preventive and curative agent to control Fusarium wilt in Stevia rebaudiana. Biol. Control 2021, 155, 104537. [Google Scholar] [CrossRef]
- Chen, J.; Vallikkannu, M.; Karuppiah, V. Systemically Induced resistance against Maize Diseases by Trichoderma spp. In Trichoderma; Springer: Gateway East, Singapore, 2020; pp. 111–123. [Google Scholar]
- Veenstra, A.; Rafudeen, M.S.; Murray, S.L. Trichoderma asperellum isolated from African maize seed directly inhibits Fusarium verticillioides growth in vitro. Eur. J. Plant Pathol. 2018, 153, 279–283. [Google Scholar] [CrossRef]
- He, A.-l.; Liu, J.; Wang, X.-H.; Zhang, Q.-G.; Song, W.; Chen, J. Soil application of Trichoderma asperellum GDFS1009 granules promotes growth and resistance to Fusarium graminearum in maize. J. Integr. Agric. 2019, 18, 599–606. [Google Scholar] [CrossRef]
- Collins, R.P.; Halim, A.F. Characterization of the major aroma constituent of the fungus Trichoderma viride. J. Agric. Food Chem. 1972, 20, 437–438. [Google Scholar] [CrossRef]
- Zeilinger, S.; Gruber, S.; Bansal, R.; Mukherjee, P.K. Secondary metabolism in Trichoderma-chemistry meets genomics. Fungal Biol. Rev. 2016, 30, 74–90. [Google Scholar] [CrossRef]
- Speckbacher, V.; Ruzsanyi, V.; Martinez-Medina, A.; Hinterdobler, W.; Doppler, M.; Schreiner, U.; Böhmdorfer, S.; Beccaccioli, M.; Schuhmacher, R.; Reverberi, M.; et al. The Lipoxygenase Lox1 Is Involved in Light- and Injury-Response, Conidiation, and Volatile Organic Compound Biosynthesis in the Mycoparasitic Fungus Trichoderma atroviride. Front. Microbiol. 2020, 11, 2004. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Ruiz, D.; Lichius, A.; Turrà, D.; Di Pietro, A.; Zeilinger, S. Chemotropism Assays for Plant Symbiosis and Mycoparasitism Related Compound Screening in Trichoderma atroviride. Front. Microbiol. 2020, 11, 601251. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).