Abstract
To date, the effects of invasive plants on soil communities and the concomitant interactions thereof have been the subject of ever-growing concern. Solanum elaeagnifolium (S) is a noxious invasive weed originating from south-central America, which has been recognized as a serious threat around the Mediterranean basin and worldwide. Herein, we compared soil nematode communities in the rhizospheres of (S), against those of its native antagonist Cichorium intybus (C), in sites where these two ruderal plants coexist (“species” effect—hereafter “sp”). These sites differed regarding the time after the initial invasion of (S) and were regarded as recently and historically invaded (10 years and 70 years after invasion; “year” effect—hereafter “yr”). Neither “sp” nor “yr” affected total nematode abundance and nematode functional indices. Diversity was lower in (S) and was reduced even more with time after invasion in both plant species. Plant parasites decreased significantly from S–10yr to S–70yr, i.e., over time after invasion, while predators were fewer in (C). Distinct nematode communities in terms of genera structure were formed under the two plant species and these were significantly affected by the time after (S) invasion. Differences between these communities related to loss of genera but also to changes in the abundance of common ones, such as Acrobeloides and Pratylenchus. Our results showed that the responses of the examined soil communities to the invasion of (S) are not straightforward and cannot be easily explained on the basis of existing theories. However, the first-time results furnished herein may be useful to integrated management strategies in the future.
1. Introduction
Present day biological invasions are a result of anthropogenous activities during which species are transferred directly or indirectly beyond their native range, thus overcrossing the natural boundaries of their original dispersal [1]. Invasive alien species are recognized as a significant part of human-induced global environmental change, threatening biodiversity and ecosystem stability with major socioeconomic consequences [2,3,4,5,6,7,8]. Some of the most prominent environmental impacts of plant invasions include biodiversity and productivity decrease as well as alteration and degradation of habitats and ecosystem functions [4,9,10,11,12,13].
Research on the remarkable success of invasive plants has revealed that plant–soil feedback is probably an important factor in determining why some alien plants become invasive [14]. In general, positive feedback occurs when a plant species induces changes in the composition of the soil community that promotes its growth and dominance, while negative feedback occurs when the plant growth is reduced [15,16]. Although invasive plants are assumed to experience positive feedbacks more often, which are possibly associated with mutualistic symbionts and/or lower root pathogen pressure [17,18,19,20,21], the role of the plant–soil feedback in invasion success may not be straightforward [22,23]. Thus, ever-growing attention has been drawn to the role of the soil food web in the plant invasion process [24]. Since soil nematodes occur at multiple levels of the soil food web, the study of their community gives a better insight into the functionality of the food web when compared to other soil biota [25,26]. Previous studies in the framework of invasion ecology have revealed adverse effects of invasive plants on the abundance and/or diversity of soil nematodes, e.g., [27,28]. In some cases, lower numbers of plant-parasitic nematodes and/or the absence of plant-parasitic nematode specialists in the new environment of invasive plants have been considered to play a part in their invasive success [29,30]. Other studies have shown that nematode community patterns in cases of plant invasion depend on species and site characteristics [31], as well as on climatic conditions during the year of sampling [32].
Solanum elaeagnifolium Cav. (Solanaceae) is a notorious invasive alien plant species, which has been unintentionally introduced from the Americas into five continents as a contaminant of commodities, declared to date as a noxious invasive weed worldwide. In Morocco, it represents a severe threat to agriculture [33,34,35], and around the Mediterranean basin, a few initial accidental introductions in the past have resulted over the last 60 years in invasive populations occupying vast territories [36]. It is considered as a problematic weed both in its native range and in other regions of the world with similar climatic conditions [33,37]. Both in its native and alien ranges, it is wild-growing, mainly on disturbed land, as a ruderal plant occupying man-made habitats [35,38], with disturbance possibly facilitating its invasion [34,39]. It has been known to compete for moisture and nutrients with many crops, while some of them, such as cotton, lucerne, wheat, maize, etc. are more susceptible to infestation [40]. Other negative effects include the release of plant inhibitors, the hosting of agricultural pests and plant diseases, its toxicity to livestock and the reduction of land value [34,36,38,40]. A nationwide research generated in Greece regarding S. elaeagnifolium has shown that its range has increased by 1750% during the last decades and that it has the potential to successfully compete with native ruderal species of the Mediterranean such as Cichorium intybus L. (Asteraceae) [41]. Additionally, it interferes with ecosystem services such as pollination [42,43] and crop production, since it facilitates or could potentially facilitate plant pathogens and pests [44,45]. Moreover, the potential impact of S. elaeagnifolium on forest regeneration and newly planted forests has raised serious concerns [46].
Despite the significant environmental and socio-economic impacts of S. elaeagnifolium’s invasion and the important role that the soil community may play in its establishment and naturalization, nothing is known about how this species affects the soil communities of the invaded areas. In this framework, and given the ongoing invasion of S. elaeagnifolium across the world, the aim of this study was to examine the effects that invasive populations of S. elaeagnifolium might have on the abundance, diversity, trophic and functional structure and composition of the soil nematode community compared to locally antagonist populations of the native C. intybus. Given that the probability of an alien species to become invasive increases with the time since its introduction [47], the differences in the respective soil communities between recently- and well-established invasive S. elaeagnifolium populations in Greece were examined. We hypothesized that the invasion and long naturalization of S. elaeagnifolium would be reflected in several aspects of the soil nematode community. Our scope is to add knowledge regarding the potential underlying mechanisms of S. elaeagnigolium’s invasiveness.
2. Materials and Methods
2.1. Study Area
All sampling sites are located in the suburban area of the city of Thessaloniki in Northern Greece, within a radius of 25 km around the city. Thessaloniki’s climate is transient between Mediterranean and Continental, with a mean annual temperature of 15.9 °C and a mean annual precipitation of 445 mm. Thessaloniki was selected since the presence of S. elaeagnifolium in the outskirts of the city dates back to 1946 (or earlier), although its invasion in the area was only reported after the 1970s [48], and especially during the last decades [49,50,51,52]. To date, it is considered one the most abundant and widespread alien species in the urban and suburban areas of Thessaloniki, occupying dozens of km2, and one of the most important invasive plant species across Greece [41,53].
Specific sites were selected according to the documented time of S. elaeagnifolium’s invasion, based on its overall historical distribution in the area of Thessaloniki [50,51,52] and in Greece [41]. In the selected sites, S. elaeagnifolium (hereafter Solanum) commonly coexists with its native antagonist C. intybus (hereafter Cichorium), which is also a perennial and expansive ruderal species [51,52]. All selected sites were situated along rural roadsides at margins of cultivated fields with intense human activity, sharing similar abiotic conditions (climate, seasonality, main geological substrate) and hosting large populations of both Solanum and Cichorium.
2.2. Soil Sampling
The rhizospheres of Solanum and Cichorium were sampled once across 18 sites that differed in regard to the period of the initial documented invasion of Solanum. Half of them were invaded about 70 years before sampling, representing the historically invaded sites (n = 9) vs. the recently invaded ones (n = 9) that were invaded about 10 years before sampling [41,52].
In each selected site, one composite soil sample was collected for each plant species, consisting of three soil cores randomly collected from the rhizospheres of three individual plants. Soil cores were taken with an auger 5 cm wide and 20 cm long, directly underneath and next to the shoot of each individual plant. Although Solanum and Cichorium co-existed in all sampling sites, an effort was made to sample plants from the center of monospecific stands, so that Solanum and Cichorium rhizospheres were at least 1.5 m apart. A total of 36 samples (9 sites × 2 invasion periods × 2 plant species) were collected, transferred to the laboratory and stored at 4 °C until soil analyses and nematode extraction, which took place within 15 days after sampling.
2.3. Soil Physicochemical Properties
Soon after sampling, a part of the collected soil samples was sieved (mesh size 2 mm) and air-dried and was used for the determination of soil texture, water content (%), pH and soil organic C and N (Table 1). Soil texture was estimated by the Bouyoucos hydrometer method [54] and pH was measured using an electrode pH-meter in a 1:2 w/v soil:water suspension [55]. Soil organic C (Corg) was determined by a wet oxidation titration procedure using an acid dichromate system, while soil organic N (Norg) was measured by the Kjeldahl method [56].

Table 1.
Soil physicochemical properties (mean ± SE) in the rhizospheres of the alien Solanum elaeagnifolium (S) and the native Cichorium intybus (C) in recently and historically invaded sites. PERMANONA results for the effects of factors “year”, “species” and their interaction are shown (ns: non-significant. *: p < 0.05).
2.4. Nematode Extraction and Identification
From each composite soil sample, we took a subsample of 200 mL of soil to extract nematodes. Prior to extraction, each soil sample was carefully hand-mixed and soil clusters were broken up also by hand. Cobb’s modified sieving and decanting method was used for the extraction [57]. After extraction, living nematodes were counted under a stereo-microscope and then they were heat-killed and fixed with 4% formaldehyde solution. From each soil sample, 100 randomly selected nematodes were identified to genus level [58]. Each genus was further assigned to trophic groups [59] and was classified across the colonizer–persister scale (cp values) [60,61].
2.5. Nematode Functional Indices
The Maturity Index (MI) for free living nematodes and the Plant Parasitic Index (PPI) for plant feeding nematodes, which both indicate the successional status of the community, were calculated according to Bongers [62]. The Enrichment index (EI), the Channel Index (CI) and the Structure Index (SI), which indicate the functional structure of the food web, were also calculated according to the weighted faunal analysis proposed by Ferris et al. [63]. EI is based on the abundance of enrichment opportunistic nematodes and indicates rapid decomposition of low C/N organic matter mediated by bacteria, thus suggesting whether the soil environment is nutrient enriched (high EI) or depleted (low EI). SI weights the prevalence of omnivore and predatory nematodes in the soil food web as an indicator of long and complex soil food webs with high connectance and numerous trophic links, and indicates whether the soil ecosystem is highly structured with complex trophic links (high SI) or degraded with simpler trophic links (low SI). Finally, CI indicates the degree of fungal participation in the decomposition channel of soil food web [63].
2.6. Data Analysis
To reveal differences between historical and recently invaded sites as well as between the rhizospheres of the alien Solanum and the native Cichorium, we used Permutational Multivariate Analysis of Variance (PERMANOVA [64], with “year” (70-yr vs. 10-yr) and “species” (Solanum vs. Cichorium) as fixed factors in a two-factor crossed design (“year” × “species”). When the interaction effect (“year” × “species”) was significant, we proceeded to pair-wise comparisons of the factor “year” within each “species” and vice-versa. PERMANOVA was performed on (i) single variables, i.e., soil properties, abundances of individual trophic groups, total nematode abundance, number of genera and nematode indices, using the Euclidean distance measure, as well as (ii) multivariate data-sets, i.e., the entire ensemble of nematode genera and the genera ensemble of each trophic group, using the Bray–Curtis measure. The analysis performed 4999 permutations.
To detect which trophic groups and which genera were mainly responsible for the distinction of nematode communities due to plant identity and/or time after Solanum invasion, two classification trees were constructed; one based on trophic group abundances and one based on genera abundances. This analysis is a form of classification that involves repeatedly partitioning of a set of samples into increasingly homogeneous subsets with respect to the predictor variables [65], which in this case were the trophic groups and the nematode genera. Splits or rules defining how to partition the data are based on information statistics that measure how well the split decreases impurity (heterogeneity or variance) within the resulting subsets [66]. The process is recursive, growing from the root node (the whole set of samples) to the terminal nodes [67]. The analysis was conducted using Statistica 7 for Windows [68].
To assess the diversity of nematode communities, we used the method of diversity ordering described by Patil and Taillie [69], which is based on Renyi’s index [70]. Renyi’s parametric index of order α shows varying sensitivity to the rare and abundant species of a community, as the scale parameter α changes [71]. For each community it provides a profile of the most widely used diversity indices. For α = 0, the index equals the total number of species; for α = 1, it equals Shannon’s index; for α = 2, it equals Simpson’s index. For α tending to infinite, the index is most sensitive to the abundant species of a community. Thus, when two diversity profiles differ in the range of low α values, this is due to the number of species. In the range of high α values, differences between communities are due to the presence of abundant species. When two diversity profiles intersect, the two communities may be ranked differently depending on the diversity index used [72]. In our study, nematode genera instead of species were used, and calculations were performed with Past 3.17 [73].
3. Results
All sites had an alkaline pH and were characterized by low levels of organic C and organic N, and low water availability (Table 1). Soil texture was estimated as sandy loam at the historically invaded sites and loamy sand at the recently invaded sites. PERMANOVA revealed no significant differences of soil parameters in Solanum and Cihorium rhizospheres or between recently (10-yr) and historically (70-yr) invaded sites. The only exception was water availability, which was significantly lower in historically invaded sites.
A total of 52 nematode genera were recorded across all sites, which were appointed to six trophic groups: parasitic and non-parasitic plant feeders, bacterivores, fungivores, predators and omnivores. The mean values of trophic group abundances are given in Figure 1, while the mean values of total nematode abundance, number of genera and nematode functional indices are shown in Table 2. In all cases, the effects of the factors “year” (70-yr vs. 10-yr), “species” (Solanum vs. Cichorium) and their interaction are provided as generated by PERMANOVA. In cases of significant interaction effects, the significant differences revealed by pair-wise tests are given. In general, bacterivores were the most dominant trophic group (more than 30%), followed by the two plant feeding groups and fungivores. Predatory and omnivorous nematodes had very low contribution in all sites (around 1%), while omnivores were absent from S–70yr. Total nematode abundance was not affected by the presence of Solanum, while the number of genera was significantly reduced overtime (Table 2). The abundance of the bacterivorous and fungivorous trophic groups, as well as that of omnivores, did not differ significantly between Solanum and Cichorium rhizospheres or between recently and historically invaded sites (Figure 1). On the contrary, both the time after Solanum invasion (“year”), and the interaction of plant species identity and time after invasion (“species” × “year”), affected significantly the abundance of plant parasitic nematodes. More specifically, from recently to historically invaded sites, i.e., through time after the initial invasion of Solanum, parasitic plant feeders decreased significantly in the Solanum rhizosphere, which resulted in a significant difference between Solanum and Cichorium in the historically invaded sites. The same pattern of changes, although not significant, was followed also by the non-parasitic plant feeders. The abundance of predatory nematodes was significantly higher in Solanum rhizospheres.
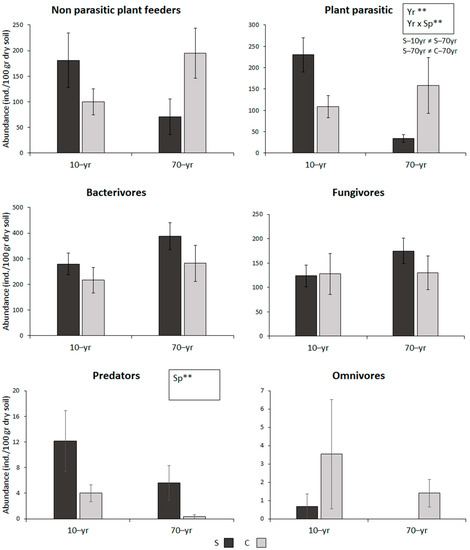
Figure 1.
Abundance of nematode trophic groups (mean ± SE) in the rhizospheres of the alien Solanum elaeagnifolium (S, dark grey bars) and the native Cichorium intybus (C, light grey bars) in recently (10yr) and historically (70yr) invaded sites. Results of PERMANONA, when significant, are indicated in boxes (**: p< 0.01). In case of significant interaction effect, the significant differences revealed by pair-wise tests are indicated.
Table 2.
Mean values (±SE) of total nematode abundance (individuals per 100 g dry soil), number of nematode genera and nematode functional indices in the rhizospheres of the alien Solanum elaeagnifolium (S) and the native Cichorium intybus (C) in recently (10yr) and historically (70yr) invaded sites. PERMANONA results for the effects of factors “year”, “species” and their interaction are shown (ns: non-significant; *: p < 0.05; **: p < 0.01).
Regarding nematode functional indices (Table 2), EI and CI were the only ones significantly affected by the interaction of plant species identity and time after the invasion of Solanum. EI values under Solanum decreased over time while the opposite was observed for Cichorium. The reverse trend was observed for CI. However, despite the significant “yr × sp” effect, these changes of EI and CI hardly failed to be significant according to the pair-wise tests (p values between 0.05 and 0.08), and therefore they are not presented in Table 2.
In Table 3, we provide the PERMANOVA results for the multivariate data sets, namely the whole ensemble of nematode genera and the genera ensemble of each trophic group. For the total nematode community, PERMANOVA revealed significant differences both due to time after Solanum invasion and due to plant species identity. The same holds for the structure of the plant parasitic, the bacterivore and the fungivore trophic groups. Regarding predators, the C–70yr sites differed from the rest, because only one genus was present, namely Aporcelaimellus, with trivial abundance (Table 3 and Table 4).
Table 3.
PERMANOVA results for the effects of factors “year”, “species” and their interaction on multiple variable data sets, i.e., abundances of nematode genera (ns: non-significant; *: p < 0.05, **: p < 0.01, ***: p < 0.001). In the case of significant interaction effect, the significant differences revealed by pair-wise tests are indicated.
Table 4.
Mean abundance (individuals per 100 g dry soil) of nematode genera and % participation to the total community in the rhizospheres of the alien Solanum elaeagnifolium (S) and the native Cichorium intybus (C) in recently (10 yr) and historically (70 yr) invaded sites.
The classification tree based on trophic group abundances is presented in Figure 2. The tree consisted of nine terminal and eight non-terminal nodes. Samples from historically invaded Solanum sites (S–70yr) were separated from the rest mainly due to the low abundance of plant parasitic nematodes (terminal node 2). For these samples the percentage of correct classification was 88.9%. The majority (66.7%) of samples from recently invaded Cichorium sites (C–10yr) was classified correctly in the terminal node 3, which was separated from the S–70yr node, mainly due to plant parasites once again. Both Cichorium samples from historically invaded sites (C–70yr) and Solanum samples from recently invaded sites (S–10yr) exhibited great variability and, therefore, only 55.6% of the S–10yr samples were classified correctly (terminal node 9), whereas correct classification for C–70yr samples was much lower and these samples were dispersed in several terminal nodes.
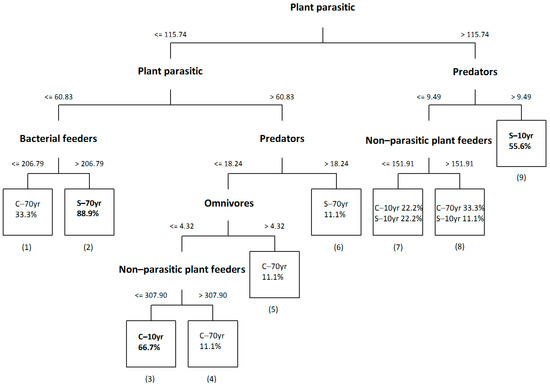
Figure 2.
Classification tree based on nematode trophic group abundance (individuals/100 g dry soil) in the rhizospheres of the alien Solanum elaeagnifolium (S) and the native Cichorium intybus (C) in recently (10yr) and historically (70yr) invaded sites. The percentage of correct classification of samples in each node is indicated. Numbers are assigned to all terminal nodes. Pure terminal nodes are indicated in bold.
In Figure 3, we present the classification tree applied on the abundances of all recorded genera. The tree consisted of eight terminal and seven non-terminal nodes. The C–70yr sites were separated from the rest due to the low abundance of the bacterivorous Acrobeloides, and are characterized by the high abundance of the plant parasitic Merlinius (77.8% correct classification of samples, terminal node 2). On the other hand, the S–10yr and S–70yr sites were characterized by higher abundance of Acrobeloides and support distinct communities (88.9% and 77.8% correct classification of samples in nodes 7 and 4 respectively). The main difference between them was related with the abundance of the plant parasitic Pratylenchus, which was high in S–10yr and decreased in S–70yr. Samples from C–10yr sites exhibited the greatest variability compared to the rest, since only 55.6% of the samples were classified in the same terminal node 5. Other nematode genera also important in the distinction of samples from different sites were the bacterivorous Eucephalobus, Panagrolaimus and the fungivorous Aphelenchus.
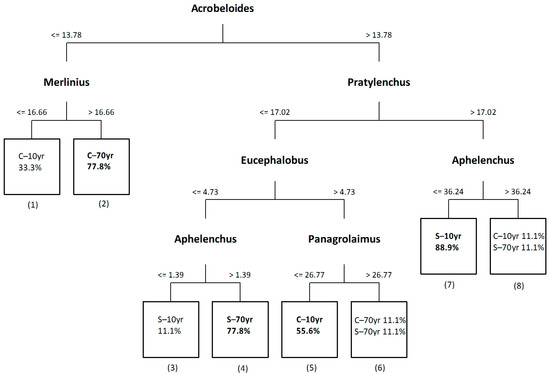
Figure 3.
Classification tree based on nematode genera abundance (individuals/100 g dry soil) in the rhizospheres of the alien Solanum elaeagnifolium (S) and the native Cichorium intybus (C) in recently (10yr) and historically (70yr) invaded sites. The percentage of correct classification of samples in each node is indicated. Numbers are assigned to all terminal nodes. Pure terminal nodes are indicated in bold.
The diversity profiles of nematode communities at all sites are presented in Figure 4. The most diverse community was the one at C–10yr, followed by S–10yr, C–70yr and finally S–70yr. Thus, diversity was higher in recently invaded sites under both focal plant species, whereas nematode communities in Cichorium rhizospheres were more diverse than those under Solanum, regardless of the time after Solanum invasion.
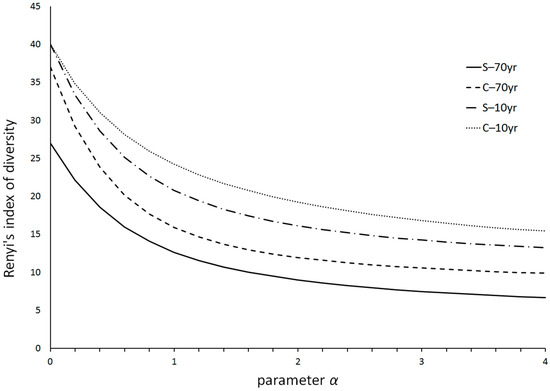
Figure 4.
Diversity profiles of nematode communities in the rhizospheres of the alien Solanum elaeagnifolium (S) and the native Cichorium intybus (C) in recently (10 yr) and historically (70 yr) invaded sites.
Table 4 shows the mean abundance and percent participation of the nematode genera to the total community for each “yr” × “sp” combination. Τhe bacterivore trophic group was the most rich in nematode genera, followed by that of plant parasites. There was a reduction in the number of genera from forty in the S–10yr sites to twenty-seven in S–70yr; five of them were plant parasitic, six were bacterivorous, one fungivorous and one omnivorous. Τhe communities at C–10yr, C–70yr and S–10yr were not over-dominated by a certain genus, but at S–70yr, the bacterivore Chiloplacus had 23% participation. Overall, most genera were cp-2 bacterivores or cp-3 plant parasites.
4. Discussion
In this study we tried to compare soil nematode communities in the rhizospheres of the alien invasive plant Solanum elaeagnifolium (shortly Solanum) to those of its native antagonist Cichorium intybus (shortly Cichorium), examining sites where these two ruderal plants coexist. These sites differ regarding the time after the initial invasion of Solanum and are regarded as historically invaded (70 yr after invasion) and recently invaded (10 yr after invasion). Therefore, in this study we tried to elucidate the effects of two factors on the soil communities; the first factor was the plant species per se (alien Solanum vs. native Cichorium) and the second factor was the years after Solanum invasion (10-yr vs. 70-yr).
The soil physicochemical characteristics of all our sites indicated a disturbed environment; sandy soil with low organic content, alkaline pH, and low water content, especially in the historically invaded sites. In these sites, Solanum covers much larger areas compared to 10 yr sites. Obviously its deep and extensive taproot system with long horizontal root clades [33] allows these plants to endure and thrive in dry environments [74].
As regards soil nematodes, their total abundance was not affected either by “species” or by “year”. Most often, nematode abundance is negatively correlated with the presence of invasive plants, e.g., [27,75,76]; however, contrasting responses of nematode abundance to Bromus tectorum L. invasion were found in two different grassland associations [24]. Most researchers focus on changes in nematode trophic structure after plant invasion, using the nematode community as an analogue of the soil food-web. In our study, the abundance of microbivorous trophic groups (bacterivores, fungivores, omnivores) did not differ significantly between the two focal plant species or between recently and historically invaded sites. The most striking effect of Solanum invasion was observed in the case of the plant parasites, which were fewer under the alien Solanum than under the native Cichorium, but only in the 70 yr sites. In the recently invaded sites (10 yr), the plant parasite assemblage supported by Solanum was even larger than that of Cichorium. It seems that plant parasitic nematodes and, to a lesser extent, their non-parasitic counterparts, decreased from 10 yr to 70 yr sites, i.e., through time after the initial invasion, but only in the Solanum rhizospheres. The lower pressure of root-herbivory on alien plants is considered one of the reasons for invasion success, and is related to the escape of invasive plants from their natural enemies when entering a new territory [77] or to the escape from feeding-specialist nematodes hosted by their native neighbors [30]. The release of chemical bioactive compounds into the soil is another potential mechanism regarding the invasion success of some plant species [78], and in the context of plant–soil interactions, these compounds may be also toxic for the plant parasites in the soil that feed on the roots of invaders [79]. Indeed, Solanum is known to contain several secondary metabolites in its leaves, fruits and roots, such as alkaloids [33,80], flavonoids [81,82,83] and terpenes [84], providing it with a strong allelopathic effect. Besides its phytotoxicity [82,85], Solanum has also been shown to have insecticidal, molluscicidal as well as nematicidal effects [85,86,87,88,89,90]. Although the “enemy-release” hypothesis and the chemical nature of Solanum may explain its lower root-herbivore load compared to the native Cichorium in our historically invaded sites, they cannot explain our findings in the recently invaded sites (10 yr). Similar results to those from our 10 yr sites, i.e., greater number of plant parasitic nematodes supported by the invasive rather than the native plant are reported in Porazinska et al. [32]. Neutel et al. [91] who studied changes in soil food webs in sand dunes during vegetation succession, observed an increase in phytophagous nematodes up to the first 15–25 years, while in later succession stages (50–100 years), phytophagous nematodes tend to disappear. Other studies [92] show that below-ground herbivory tends to gradually increase from early to late succession, suggesting that roots in early compared to later succession stage are often fast-growing, thinner, softer and, therefore, more susceptible to herbivores.
Nematode functional indices, apart from EI and CI, did nοt provide statistically significant results. Nevertheless, they offered an assessment of the prevailing conditions, which indicated disturbed and degraded soils that were able to support very simple food webs [61,62,63]. For example, the reduction of SI found in S–70yr, was due to the reduction of cp-4 and cp-5 nematodes, which are large persisters. A limited presence of larger size nematodes is associated with habitat degradation [93].
Both the species identity and the time after invasion affected the diversity of the nematode community. Lower numbers of nematode genera under the invasive rather than under native plants are reported in Porazinska et al. [27] and Fitoussi et al. [75]. However, in our study, the changes in diversity followed a quite different pattern. The inspection of all diversity profiles suggested that the alien Solanum supported indeed a less diverse community compared to the native Cichorium, but in the recently invaded sites this was not due to lower number of genera. Most importantly, the time after the initial invasion of Solanum had an even stronger negative effect on nematode diversity, i.e., loss of genera and increased participation of the widespread Chiloplacus in S-70yr sites (Figure 4, Table 4). Indeed, the number of genera was significantly affected only by “year” (Table 2). Belnap et al. [94] studied Bromus tectorum invasion comparing nematode communities in non-invaded, recently invaded (6–7 yr) and historically invaded (50+ yr) sites and found that alterations in soil communities, most particularly loss of genera, occurred mainly during the first years after invasion. In any case, the most possible explanation for reduced diversity in our sites is the depauperation of the above-ground plant communities. The introduction of Solanum in an area is often known to result in the formation of extended monospecific stands [36,41,95]. This dominance was especially evident in our historically invaded sites. Plants are the main drivers of changes in the soil ecosystem, since they provide food not only to root herbivores, such as plant-feeding soil nematodes, but also to microbivores, such as bacterivorous and fungivorous nematodes, by forming litter above and belowground and by releasing organic material from their roots as exudates [96,97]. Therefore, loss of nematode genera in our historically invaded sites did not occur only within the plant feeders, but within the free-living trophic groups, as well (Table 4).
Despite the fact that the differences in trophic structure of nematode communities were mainly driven by plant parasites (Figure 2), all genera contributed to the distinction of communities, however, with variable differentiating power (Table 3, Figure 3). Thus, nematode communities with distinct genera structure were formed under the studied plant species and these were significantly affected by the time after the Solanum invasion. Differences between these communities related to loss of genera but also to changes in abundance and/or percentage (%) participation of very common ones, such as Acrobeloides and Pratylenchus (Figure 3, Table 4).
5. Conclusions
The total nematode abundance was not affected significantly either by the species identity (alien Solanum vs native Cichorium) or by the time after Solanum invasion. The most striking effect of Solanum invasion on the trophic structure was observed in the case of root herbivores (mainly plant parasites). In the recently invaded sites (10 yr), the plant parasite load of Solanum was even larger than that of Cichorium, but this was reversed in the historically invaded sites (70 yr); the abundance of plant parasites was significantly reduced with time in the Solanum rhizospheres, so that root herbivory was more intense in Cichorium. Changes of the nematode functional indices indicated that long naturalization of Solanum increased degradation of the already disturbed invaded habitats. However, in most cases these changes were not statistically significant. The communities in Solanum rhizospheres were less diverse than those in Cichorium, due to changes in dominance patterns of abundant genera. Loss of nematode genera occurred only with time after invasion in both plant species, especially in Solanum. The whole ensemble of nematode genera contributed to the formation of distinct communities under the two plant species that were significantly affected by the time after Solanum invasion. Differences between these communities related to loss of certain genera but also to changes in abundance of others.
Our results showed that the responses of the soil communities to plant invasion, as studied herein, are not straightforward and cannot be easily explained on the basis of existing theories. To this end, more studies are needed to decipher the underlying mechanisms of invasive species success involving different invasive species, more ecosystems and more soil biota. However, the first-time results furnished herein may be useful to integrated management strategies in the future focused on this notorious alien invasive plant species.
Author Contributions
Conceptualization, N.K. and M.D.A.; data curation, M.K.; methodology, N.K., M.D.A. and M.K.; formal analysis, M.K.; investigation, M.K.; visualization, M.K.; validation, N.K. and M.D.A.; writing—original draft preparation, M.K.; writing—review and editing, M.D.A. and N.K.; supervision, M.D.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data available on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Soulé, M.E. The onslaught of alien species, and other challenges in the coming decades. Conserv. Biol. 1990, 4, 233–240. [Google Scholar] [CrossRef]
- Vitousek, P.M.; D’antonio, C.M.; Loope, L.L.; Rejmanek, M.; Westbrooks, R. Introduced species: A significant component of human-caused global change. N. Z. J. Ecol. 1997, 21, 1–16. Available online: https://www.researchgate.net/publication/246522062_Introduced_Species_A_Significant_Component_of_Human-Caused_Global_Change (accessed on 31 August 2022).
- Mack, R.N.; Simberloff, D.; Mark Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- McNeely, J.A. An introduction to human dimensions of invasive alien species. In The Great Reshuffling: Human Dimensions of Invasive Alien Species; IUCN: Gland, Switzerland; Cambridge, UK, 2001; pp. 5–20. [Google Scholar]
- Hulme, P.E. Biological invasions in Europe: Drivers, pressures, states, impacts and responses. In Biodiversity under Threat; Hester, R., Harrison, R., Eds.; Issues in Environmental Science and Technology; Cambridge University Press: Cambridge, UK, 2007; Volume 25, pp. 56–80. [Google Scholar]
- Bacher, S.; Blackburn, T.M.; Essl, F.; Genovesi, P.; Heikkilä, J.; Jeschke, J.M.; Jones, G.; Keller, R.; Kenis, M.; Kueffer, C. Socio-economic impact classification of alien taxa (SEICAT). Methods Ecol. Evol. 2018, 9, 159–168. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef]
- DiTomaso, J.M. Invasive weeds in rangelands: Species, impacts, and management. Weed Sci. 2000, 48, 255–265. [Google Scholar] [CrossRef]
- Simberloff, D. Non-native species do threaten the natural environment! J. Agric. Environ. Ethics 2005, 18, 595–607. [Google Scholar] [CrossRef]
- Charles, H.; Dukes, J.S. Impacts of invasive species on ecosystem services. In Biological Invasions; Nentwig, W., Ed.; Ecological Studies; Springer: Berlin, Germany, 2008; Volume 193, pp. 217–237. [Google Scholar]
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef]
- Vilà, M.; Hulme, P.E. Non-native species, ecosystem services, and human well-being. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P.E., Eds.; Volume Invading Nature—Springer Series in Invasion Ecology; Springer: Cham, Switzerland, 2017; Volume 12, pp. 1–14. [Google Scholar]
- Kuyper, T.W.; de Goede, R.G.M. Interactions Between Higher Plants and Soil-dwelling Organisms. In Vegetation Ecology, 2nd ed.; van der Maarel, E., Franklin, J., Eds.; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2013; pp. 260–284. [Google Scholar] [CrossRef]
- Bever, J.D.; Westover, K.M.; Antonovics, J. Incorporating the soil community into plant population dynamics: The utility of the feedback approach. J. Ecol. 1997, 85, 561–573. [Google Scholar] [CrossRef]
- Bever, J.D. Soil community feedback and the coexistence of competitors: Conceptual frameworks and empirical tests. New Phytol. 2003, 157, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Thelen, G.C.; Rodriguez, A.; Holben, W.E. Soil biota and exotic plant invasion. Nature 2004, 427, 731–733. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, B.E.; Klironomos, J.N. Breaking new ground: Soil communities and exotic plant invasion. BioScience 2005, 55, 477–487. [Google Scholar] [CrossRef]
- Reinhart, K.O.; Callaway, R.M. Soil biota and invasive plants. New Phytol. 2006, 170, 445–457. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A. Plant–soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Bradford, M.A.; Pernilla Brinkman, E.; van de Voorde, T.F.; Veen, G. Where, when and how plant–soil feedback matters in a changing world. Funct. Ecol. 2016, 30, 1109–1121. [Google Scholar] [CrossRef]
- Suding, K.N.; Stanley Harpole, W.; Fukami, T.; Kulmatiski, A.; MacDougall, A.S.; Stein, C.; van der Putten, W.H. Consequences of plant–soil feedbacks in invasion. J. Ecol. 2013, 101, 298–308. [Google Scholar] [CrossRef]
- Schittko, C.; Runge, C.; Strupp, M.; Wolff, S.; Wurst, S. No evidence that plant–soil feedback effects of native and invasive plant species under glasshouse conditions are reflected in the field. J. Ecol. 2016, 104, 1243–1249. [Google Scholar] [CrossRef]
- Belnap, J.; Phillips, S.L. Soil biota in an ungrazed grassland: Response to annual grass (Bromus tectorum) invasion. Ecol. Appl. 2001, 11, 1261–1275. [Google Scholar] [CrossRef]
- Ferris, H.; Venette, R.; Scow, K. Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralisation function. Appl. Soil Ecol. 2004, 25, 19–35. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T. Nematode indicators of organic enrichment. J. Nematol. 2006, 38, 3. Available online: https://pubmed.ncbi.nlm.nih.gov/19259424/ (accessed on 31 August 2022). [PubMed]
- Porazinska, D.L.; Pratt, P.D.; Glblin-Davis, R.M. Consequences of Melaleuca quinquenervia invasion on soil nematodes in the Florida Everglades. J. Nematol. 2007, 39, 305. Available online: https://pubmed.ncbi.nlm.nih.gov/19259503/ (accessed on 31 August 2022). [PubMed]
- Lazzaro, L.; Mazza, G.; d’Errico, G.; Fabiani, A.; Giuliani, C.; Inghilesi, A.F.; Lagomarsino, A.; Landi, S.; Lastrucci, L.; Pastorelli, R. How ecosystems change following invasion by Robinia pseudoacacia: Insights from soil chemical properties and soil microbial, nematode, microarthropod and plant communities. Sci. Total Environ. 2018, 622, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Knevel, I.C.; Lans, T.; Menting, F.B.; Hertling, U.M.; van der Putten, W.H. Release from native root herbivores and biotic resistance by soil pathogens in a new habitat both affect the alien Ammophila arenaria in South Africa. Oecologia 2004, 141, 502–510. [Google Scholar] [CrossRef]
- van der Putten, W.; Yeates, G.; Duyts, H.; Reis, C.S.; Karssen, G. Invasive plants and their escape from root herbivory: A worldwide comparison of the root-feeding nematode communities of the dune grass Ammophila arenaria in natural and introduced ranges. Biol. Invasions 2005, 7, 733–746. [Google Scholar] [CrossRef]
- Yeates, G.W.; Williams, P.A. Influence of three invasive weeds and site factors on soil microfauna in New Zealand. Pedobiologia 2001, 45, 367–383. [Google Scholar] [CrossRef]
- Porazinska, D.L.; Fujisaki, I.; Purcell, M.F.; Giblin-Davis, R.M. Plant invasions from a belowground nematocentric perspective. Soil Biol. Biochem. 2014, 77, 213–220. [Google Scholar] [CrossRef]
- Boyd, J.; Murray, D.; Tyrl, R. Silverleaf nightshade, Solanum elaeagnifolium, origin, distribution, and relation to man. Econ. Bot. 1984, 38, 210–217. [Google Scholar] [CrossRef]
- Mekki, M. Biology, distribution and impacts of silverleaf nightshade (Solanum elaeagnifolium Cav.). Bull. OEPP 2007, 37, 114–118. [Google Scholar] [CrossRef]
- Utah, W.; Rico, P. Datasheet on quarantine—Solanum elaeagnifolium. Bull. OEPP 2007, 37, 236–245. Available online: https://gd.eppo.int/download/doc/246_datasheet_SOLEL.pdf (accessed on 31 August 2022).
- Uludag, A.; Gbehounou, G.; Kashefi, J.; Bouhache, M.; Bon, M.C.; Bell, C.; Lagopodi, A. Review of the current situation for Solanum elaeagnifolium in the Mediterranean Basin. Bull. OEPP 2016, 46, 139–147. [Google Scholar] [CrossRef]
- Roche, C. Silverleaf Nightshade (Solanum elaeagnifolium Cav.). In PNW-Pacific Northwest Extension Publication; Cooperative Extension Service, Washington State Universitiy: Washington, DC, USA, 1991. [Google Scholar]
- Stanton, R.; Heap, J.; Carter, R.; Wu, H. Solanum elaeagnifolium Cav. In The Biology of Australian Weeds; Panetta, F., Ed.; RG and FJ Richardson: Melbourne, Australia, 2009; Volume 3, pp. 1–35. [Google Scholar]
- Chavana, J.; Singh, S.; Vazquez, A.; Christoffersen, B.; Racelis, A.; Kariyat, R.R. Local adaptation to continuous mowing makes the noxious weed Solanum elaeagnifolium a superweed candidate by improving fitness and defense traits. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Brunel, S. Pest risk analysis for Solanum elaeagnifolium and international management measures proposed. Bull. OEP 2011, 41, 232–242. [Google Scholar] [CrossRef]
- Krigas, N.; Tsiafouli, M.A.; Katsoulis, G.; Votsi, N.-E.; van Kleunen, M. Investigating the invasion pattern of the alien plant Solanum elaeagnifolium Cav. (Silverleaf Nightshade): Environmental and human-induced drivers. Plants 2021, 10, 805. [Google Scholar] [CrossRef]
- Tscheulin, T.; Petanidou, T.; Potts, S.G.; Settele, J. The impact of Solanum elaeagnifolium, an invasive plant in the Mediterranean, on the flower visitation and seed set of the native co-flowering species Glaucium flavum. Plant Ecol. 2009, 205, 77–85. [Google Scholar] [CrossRef]
- Tscheulin, T.; Petanidou, T. The presence of the invasive plant Solanum elaeagnifolium deters honeybees and increases pollen limitation in the native co-flowering species Glaucium flavum. Biol. Invasions 2013, 15, 385–393. [Google Scholar] [CrossRef]
- Tscheulin, T.; Petanidou, T.; Settele, J. Invasive weed facilitates incidence of Colorado potato beetle on potato crop. Int. J. Pest Manag. 2009, 55, 165–173. [Google Scholar] [CrossRef]
- Kashefi, J.; Lagopodi, A. New pathogens of Solanum elaeagnifolium investigated as possible biocontrol agents of the weed in Greece. In Proceedings of the Environmental Weeds and Invasive Plants, 3rd International Symposium of Environmental Weeds and Invasive Plants, Ascona, Switzerland, 2–7 October 2011. [Google Scholar]
- Ganatsas, P.; Tsitsoni, T.; Tsakaldimi, M.; Zagas, T. Reforestation of degraded Kermes oak shrublands with planted pines: Effects on vegetation cover, species diversity and community structure. New For. 2012, 43, 1–11. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog. Phys. Geogr. 2006, 30, 409–431. [Google Scholar] [CrossRef]
- Yannitsaros, A.; Economidou, E. Studies on the adventive flora of Greece. I. General remarks on some recently introduced taxa. Candollea 1974, 29, 111–119. [Google Scholar]
- Browicz, K. Nicotiana glauca and Solanum elaeagnifolium [Solanaceae]—Two xenophytes from South America and the history of their spreading in the eastern Mediterranean. Fragm. Flor. Geobot. Suppl. 1993, 2, 299–305. [Google Scholar]
- Lagoudakis, N.; Krigas, N.; Hanlidou, E.; Kokkini, S. Plant species with alkaloids in the city of Thessaloniki (N Greece): Distribution and alkaloid content. Bot. Chron. 2002, 15, 35–44. [Google Scholar]
- Krigas, N. Flora and Human Activities in the Area of Thessaloniki: Biological Approach and Historical Consideration. Ph.D. Thesis, Aristotle University of Greece, Thessaloniki, Greece, 2004. [Google Scholar]
- Krigas, N.; Kokkini, S. A survey of the alien vascular flora of the urban and suburban area of Thessaloniki, N Greece. Willdenowia 2004, 34, 81–99. Available online: http://www.jstor.org/stable/3997464 (accessed on 31 August 2022). [CrossRef]
- Arianoutsou, M.; Bazos, I.; Delipetrou, P.; Kokkoris, Y. The alien flora of Greece: Taxonomy, life traits and habitat preferences. Biol. Invasions 2010, 12, 3525–3549. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Schofield, R.; Taylor, A.W. The measurement of soil pH. Soil Sci. Soc. Am. J. 1955, 19, 164–167. [Google Scholar] [CrossRef]
- Allen, S. Chemical Analysis of Ecological Materials; Blackwell Scientific Publications: Oxford, UK, 1974. [Google Scholar]
- Jacob, J.J.; van Bezooijen, J. Manual for Practical Work in Nematology; Landbouwhogeschool Wageningen: Wageningen, The Netherlands, 1984. [Google Scholar]
- Bongers, T. Identification key: De Nematoden van Nederland. KNNV Bibliotheekuitgave 1994, 46, 408. [Google Scholar]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.; Freckman, D.W.; Georgieva, S. Feeding habits in soil nematode families and genera—an outline for soil ecologists. J. Nematol. 1993, 25, 315. Available online: https://pubmed.ncbi.nlm.nih.gov/19279775/ (accessed on 31 August 2022).
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Bongers, T. The Maturity Index, the evolution of nematode life history traits, adaptive radiation and cp-scaling. Plant Soil 1999, 212, 13–22. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Bongers, T.; de Goede, R.G. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Anderson, M.J. Permanova: A Fortran computer program for permutational multivariate analysis of variance. Ecol. Monogr. 2005, 83, 557–574. Available online: http://img2.timg.co.il/forums/1_124959686.pdf (accessed on 31 August 2022). [CrossRef]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Routledge: New York, NY, USA, 1984. [Google Scholar]
- Clark, L.A.; Pregibon, D. Tree-based models. In Statistical Models in S.; Hastie, T., Ed.; Routledge: New York, NY, USA, 1992. [Google Scholar]
- Friedl, M.A.; Brodley, C.E. Decision tree classification of land cover from remotely sensed data. Remote Sens. Environ. 1997, 61, 399–409. [Google Scholar] [CrossRef]
- Statistica. Data Analysis Software System; Version 7®; StatSoft, Inc.: Tulsa, OK, USA, 2004; Available online: www.statsoft.com (accessed on 30 August 2022).
- Patil, G.; Taillie, C. An overview of diversity. In Ecological Diversity in Theory and Practice; Grassle, J., Patil, G., Smith, W., Taillie, C., Eds.; International Cooperative Publishing House: Fairland, ML, USA, 1979. [Google Scholar]
- Rényi, A. On measures of entropy and information. In Proceedings of the Fourth Berkeley Symposium on Mathematical Statistics and Probability, Volume 1: Contributions to the Theory of Statistics, Berkley, CA, USA, 20 June–30 July 1961; pp. 547–562. [Google Scholar]
- Ricotta, C. From theoretical ecology to statistical physics and back: Self-similar landscape metrics as a synthesis of ecological diversity and geometrical complexity. Ecol. Modell. 2000, 125, 245–253. [Google Scholar] [CrossRef]
- Tóthmérész, B. Comparison of different methods for diversity ordering. J. Veg. Sci. 1995, 6, 283–290. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, A.; Ryan, P. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4. Available online: https://palaeo-electronica.org/2001_1/past/past.pdf (accessed on 31 August 2022).
- Travlos, I.S. Responses of invasive silverleaf nightshade (Solanum elaeagnifolium) populations to varying soil water availability. Phytoparasitica 2013, 41, 41–48. [Google Scholar] [CrossRef]
- Fitoussi, N.; Pen-Mouratov, S.; Steinberger, Y. Soil free-living nematodes as bio-indicators for assaying the invasive effect of the alien plant Heterotheca subaxillaris in a coastal dune ecosystem. Appl. Soil Ecol. 2016, 102, 1–9. [Google Scholar] [CrossRef]
- Renčo, M.; Čerevková, A.; Homolová, Z. Nematode communities indicate the negative impact of Reynoutria japonica invasion on soil fauna in ruderal habitats of Tatra National park in Slovakia. Glob. Ecol. Conserv. 2021, 26, e01470. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. Allelopathy and exotic plant invasion. Plant Soil 2003, 256, 29–39. [Google Scholar] [CrossRef]
- Renčo, M.; Jurová, J.; Gömöryová, E.; Čerevková, A. Long-term giant hogweed invasion contributes to the structural changes of soil nematofauna. Plants 2021, 10, 2103. [Google Scholar] [CrossRef] [PubMed]
- Sammani, A.; Shammaa, E.; Chehna, F. Qualitative and quantitative steroidal alkaloids of Solanum species distributed widely in Syria by TLC and HPLC. Int. J. Pharm. Sci. Rev. Res. 2013, 23, 23–27. Available online: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.1037.8833&rep=rep1&type=pdf (accessed on 31 August 2022).
- Chiale, C.A.; Cabrera, J.L.; Juliani, H.R. Kaempferol 3-(6 ″-cis-cinnamoylglucoside) from Solanum elaeagnifolium. Phytochemistry 1991, 30, 1042–1043. [Google Scholar] [CrossRef]
- Abdel aziz Balah, M. Herbicidal activity of constituents isolated from Solanum elaeagnifolium (Solanaceae). J. Crop Prot. 2015, 4, 487–496. Available online: https://jcp.modares.ac.ir/article-3-3802-en.html (accessed on 31 August 2022).
- Badawy, A.; Zayed, R.; Ahmed, S.; Hassanean, H. Phytochemical and pharmacological studies of Solanum elaeagnifolium growing in Egypt. J. Nat. Prod. 2013, 6, 156–167. Available online: http://www.journalofnaturalproducts.com/Volume6/22_Res_paper-21.pdf (accessed on 31 August 2022).
- Tsaballa, A.; Nikolaidis, A.; Trikka, F.; Ignea, C.; Kampranis, S.C.; Makris, A.M.; Argiriou, A. Use of the de novo transcriptome analysis of silver-leaf nightshade (Solanum elaeagnifolium) to identify gene expression changes associated with wounding and terpene biosynthesis. BMC Genom. 2015, 16, 1–15. [Google Scholar] [CrossRef][Green Version]
- Balah, M.A.; AbdelRazek, G.M. Pesticidal activity of Solanum elaeagnifolium Cav. leaves against nematodes and perennial weeds. Acta Ecol. Sin. 2020, 40, 373–379. [Google Scholar] [CrossRef]
- Wuyts, N.; Swennen, R.; De Waele, D. Effects of plant phenylpropanoid pathway products and selected terpenoids and alkaloids on the behaviour of the plant-parasitic nematodes Radopholus similis, Pratylenchus penetrans and Meloidogyne incognita. Nematology 2006, 8, 89–101. [Google Scholar] [CrossRef]
- Faizi, S.; Fayyaz, S.; Bano, S.; Yawar Iqbal, E.; Siddiqi, H.; Naz, A. Isolation of nematicidal compounds from Tagetes patula L. yellow flowers: Structure–activity relationship studies against cyst nematode Heterodera zeae infective stage larvae. J. Agric. Food Chem. 2011, 59, 9080–9093. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, A.B.; Chaieb, I.; Zarrad, K.; Laarif, A. Insecticidal activity of methanolic extract of silverleaf nightshade against Tribolium castaneum. Int. J. Entomol. Res. 2015, 3, 23–28. Available online: https://esciencepress.net/journals/index.php/IJER/article/view/1002 (accessed on 31 August 2022).
- Hamouda, A.B.; Zarrad, K.; Chaieb, I. Antifeedant and insecticidal properties of Solanum elaeagnifolium extracts on the African Cotton Leafworm. Azarian J. Agric. 2015, 2, 71–74. Available online: https://www.researchgate.net/publication/301676646_Antifeedant_and_insecticidal_properties_of_Solanum_elaeagnifolium_extracts_on_the_African_Cotton_Leafworm (accessed on 31 August 2022).
- Njeh, F.; Feki, H.; Koubaa, I.; Hamed, N.; Damak, M.; Ayadi, A.; Hammami, H.; Mezghani-Jarraya, R. Molluscicidal activity of Solanum elaeagnifolium seeds against Galba truncatula intermediate host of Fasciola hepatica: Identification of β-solamarine. Pharm. Biol. 2016, 54, 726–731. [Google Scholar] [CrossRef]
- Neutel, A.-M.; Heesterbeek, J.A.; Van de Koppel, J.; Hoenderboom, G.; Vos, A.; Kaldeway, C.; Berendse, F.; De Ruiter, P.C. Reconciling complexity with stability in naturally assembling food webs. Nature 2007, 449, 599–602. [Google Scholar] [CrossRef]
- Rasmann, S.; Bauerle, T.L.; Poveda, K.; Vannette, R. Predicting root defence against herbivores during succession. Funct. Ecol. 2011, 25, 368–379. [Google Scholar] [CrossRef]
- Archidona-Yuste, A.; Wiegand, T.; Eisenhauer, N.; Cantalapiedra-Navarrete, C.; Palomares-Rius, J.E.; Castillo, P. Agriculture causes homogenization of plant-feeding nematode communities at the regional scale. J. Appl. Ecol. 2021, 58, 2881–2891. [Google Scholar] [CrossRef]
- Belnap, J.; Phillips, S.L.; Sherrod, S.K.; Moldenke, A. Soil biota can change after exotic plant invasion: Does this affect ecosystem processes? Ecology 2005, 86, 3007–3017. [Google Scholar] [CrossRef]
- Petanidou, T.; Price, M.V.; Bronstein, J.L.; Kantsa, A.; Tscheulin, T.; Kariyat, R.; Krigas, N.; Mescher, M.C.; De Moraes, C.M.; Waser, N.M. Pollination and reproduction of an invasive plant inside and outside its ancestral range. Acta Oecol. 2018, 89, 11–20. [Google Scholar] [CrossRef]
- Zhi, D.; Li, H.; Nan, W. Nematode communities in the artificially vegetated belt with or without irrigation in the Tengger Desert, China. Eur. J. Soil Biol. 2008, 44, 238–246. [Google Scholar] [CrossRef]
- Sylvain, Z.A.; Wall, D.H. Linking soil biodiversity and vegetation: Implications for a changing planet. Am. J. Bot. 2011, 98, 517–527. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).