Abstract
Sea daffodil is a perennial medicinal plant with several beneficial health properties that is enlisted as an endangered and protected species. For the exploitation and conservation of such valuable species, sustainable propagation is a key factor. For this purpose, the establishment of in vitro cultures using botanical seeds is an effective way to produce a large number of microplants which can be used as propagating material and/or bulblets with further uses for the isolation–production of biocactive substances. In this work, we performed two separate experiments where we studied (a) the effect of temperature (10 °C, 15 °C, 20 °C, 25 °C, and 30 °C) on seed germination in vitro and (b) the effect of temperature (10 °C, 20 °C, and 30 °C) and sucrose concentration (40 and 80 g/L) in growth medium on microplant development and the production of bulblets. Seed germination rates were high (over 70%) at a wide range of temperatures (15–30 °C), although the fastest and the most uniform germination was observed at 20–25 °C. Regarding the effect of temperature on microplant development and bulblet production, low temperatures (10 °C) had a negative effect on the number of leaves, bulb diameter, and fresh weight (F.W.) and dry weight (D.W.) of leaves, roots, and bulblets. The best growth of microplants (number of leaves, F.W. and D.W. of leaves, roots) and bulblet production (diameter, F.W., and D.W.) were observed in culture media with sucrose concentration 40 g/L and temperature of 30 °C. In culture medium with a sucrose concentration of 80 g/L the production of bulbs (diameter, F.W., and D.W.) was not inferior to culture media with a sucrose concentration of 40 g/L, although microplants had less root growth. It is concluded that the best treatment for the production of well-rooted microplants with large bulblets that could be used as propagating material or for the production of bulblets as a source for twin scales explants or pharmaceutical substances was the culture at 30 °C in culture medium with sucrose concentration 40 g/L. On the other hand, culture medium with a sucrose concentration of 80 g/L at 30 °C or 20 °C could be also used for the production of bulblets.
1. Introduction
The Mediterranean basin is abundant with wild medicinal plants that are highly appreciated for their bioactive properties and are an integral part of traditional medicine [1,2]. Among them, there are several halophytes that have received research attention due to their interesting agronomic, biochemical, and physiological properties [3,4,5]. Sea daffodil (Pancratium maritimum L.) is a perennial bulbous plant of the Amaryllidaceae family [6,7]. The species grows naturally on sandy beaches in many areas of the Mediterranean. Despite being a widespread plant species, human activities such as tourism, urbanization, coastline degradation, overcollection, and other ecological pressures have been reported to threaten plant populations in their natural habitats, causing it to be named an endangered and under protection plant species [8,9,10,11,12]. Sea daffodil has attracted the interest of numerous researchers due to the presence of valuable compounds such as alkaloids, flavonoids, phenolic compounds, and other substances contained in the bulbs, leaves, and flowers of the plant [13,14]. Bulbs are the most interesting plant part since they contain several alkaloids and other bioactive substances with important pharmaceutical properties, e.g., anti-virus, anti-cancer, etc. [15,16,17,18,19]. However, other plant parts such as leaves and roots are also good sources of bioactive compounds with anti-proliferative, antioxidant, antifungal, and other properties [14,20,21].
The valorization of Mediterranean flora under climate change conditions is necessary for the conservation of biodiversity and the sustainability of agroecosystems, focusing on species with the potential to produce high value-added products [1,2,3]. Halophytes are of particular interest due to their adaptation to harsh environmental conditions; therefore, special efforts for their commercial exploitation in saline farming systems have been made during the last years [22,23]. However, prior to suggesting the introduction of new species in Mediterranean farming systems, propagation is an important limiting factor that needs to be addressed for most of these species in order to ensure the genetic material conservation and the viability of this endeavor. For P. maritimum, special attention is needed considering its inclusion in the list of endangered and under-protection species in terms of bioethics and ecosystem sustainability [24]. The species is mostly asexually (vegetatively; with bulbs) and, less commonly, sexually (via seeds) propagated, while a great variation in success rates of sexual propagation among the various ecotypes has been suggested [8]. For commercial propagation of the species, the development of a standardized protocol for the production of mother bulbs is needed to produce seedlings on a large scale. For this purpose, in vitro propagation has been suggested, although a slow formation of new bulbs has been recorded [25]. The micropropagation of P. maritimum through direct and indirect regeneration has attracted the interest of researchers because it allows the fast production of large quantities of microplants or microbulbs [9,26,27,28,29,30]. For example, Dragassaki et al. [30] suggested the production of bulblets from twin-scale explants using culture medium supplemented with BA and/or NAA, while they also reported that the addition of growth regulators affected the formation of plantlets with roots and leaves. On the other hand, Georgiev et al. [27] suggested the use of sliced fruit for the in vitro production of callus and the further initiation of shoot cultures in MS growth medium containing sucrose (30 g/L), plant agar (5.5 g/L), a-naphtylacetic acid (α-ΝAA; 1.15 mg/L), and 6 benzylaminopurine (BAP; 2.0 mg/L). Georgiev et al. [26] also reported the use of auxins and cytokinins (e.g., kinetin, BAP, NAA, and 2,4-dichlorophe- noxyacetic acid (2,4-D)) in the culture medium for calli and shoot formation from P. maritimum explants derived from young stems and fruit. Similarly, Bogdanova et al. [28] reported the regeneration of bulblets obtained from explants of in vitro culture of P. maritimum mature seeds using MS culture medium supplemented with BAP (3 mg/L) and NAA (0.1 mg/L). The use of in vitro seed germination for explant production and plantlet formation was also reported by Yasemin et al. [29], who tested three different nutrient media containing agar, sucrose, and growth regulators (BAP and 2,4-D) in various amounts.
The generation of P. maritimum explants for the production of microplants or microbulbs (bulblets) through the in vitro germination of seeds has many advantages compared to the use of bulb parts (twin-scale explants derived from bulbs), such as avoiding the application of specific techniques to limit contamination [9,31,32]. Rooted seedlings produced via micropropagation are used successfully as planting material for ex vitrum plant cultivation [29]. Moreover, the formation of bulblets in microplants has been reported to favor the survival of microplants after transplantation to soil [33,34], while bulblets produced in vitro are a very good source of twin scales which can be used as explants for microplant production [9]. For the large-scale in vitro bulblet production of P. maritimum from seedlings produced in vitro, the culture conditions (e.g., temperature, culture medium, light) for seed germination and bulblet development is critical to be determined. The in vitro seed germination of P. maritimum, as in other plant species, is influenced by culture conditions, the substrate composition (e.g., plant growth regulators, sucrose), and special characteristics of the seeds (e.g., dormancy) [31]. The effect of plant growth regulators and sucrose concentration on in vitro germination of P. maritimum seeds has already been studied [9,25,29]. In contrast, limited literature reports concerning the effect of temperature on the in vitro germination of P. maritimum seeds are available since most of the studies regarding in vitro seed germination have been carried out at single temperatures, for example, at 12/16 °C (night/day) [9], 23 ± 1 °C [35], at 23 ± 2 °C [28], 25 ± 1 °C [36], or 23 °C [25].
To the best of our knowledge, there are no data available on the effect of temperature on the in vitro production of P. maritimum bulbs. Temperature is an important factor affecting plant growth, morphogenesis, and translocation of chemical substances (e.g., carbohydrates) to storage organs [37], and it has been shown to affect the in vitro bulblet production in bulbous plants [38]. Considering the above, the present study aimed to identify the effect of temperature on the in vitro germination of P. maritimum seeds and the effect of temperature in combination with the concentration of sucrose in the culture medium on in vitro bulblet production in seedlings produced in vitro. The presented results could contribute to the conservation of the species and also allow its further exploitation through commercial cultivation or bulblet production for the isolation of valuable bioactive compounds on a large scale without threatening the biodiversity of saline ecosystems. Moreover, our results provide useful information for the adoption of in vitro seed germination and bulblet production techniques that ensure the efficient use of the valuable genetic material of this endangered species.
2. Materials and Methods
2.1. Plant Material
The experiment was carried out at the University of the Peloponnese in September 2019. Mature seeds from closed pods collected from wild plants of Pancratium maritimum grown in the region of Finikounda (Southwest Peloponnese, Greece) were used. Seeds were collected in October 2005 and then stored in air-sealed glass jars at 5 ± 0.5 °C in the Laboratory of Agronomy at the University of the Peloponnese. Two experiments were conducted, as described below.
2.2. Experiment 1: The Effect of Temperature on In Vitro Seed Germination
After removing the auriferous parenchyma that covers them, seeds of Pancratium maritimum were thoroughly rinsed with tap water and then immersed in an 80% (v/v) ethyl alcohol solution for 50 sec. After that, seeds were disinfected through immersion in a sodium hypochlorite solution (1.2 % v/v) with the addition of 2 drops of Tween 20 (Sigma GmbH, Roedermark, Germany) per 100 mL of solution for 30 min under constant stirring and washed twice with sterilized deionized water. The sterilized seeds were transferred under aseptic conditions to ventilated disposable 9 cm Petri dishes containing the MS [39] basal mixture (M5519, Sigma-Aldrich, St. Louis, MO, USA) supplemented with 30 g/L sucrose and solidified with 4 g/L agar (Serva Electrophoresis GmbH, Heidelberg, Germany) for higher fluidity. The pH of the culture medium was adjusted to 5.75 [9].
Five seeds were placed in each Petri dish, and four replicates of twenty-five seeds each (i.e., five Petri dishes per replicate) were used for each temperature tested. The Petri dishes were placed in incubation chambers at 10 °C, 15 °C, 20 °C, 25 °C, and 30 °C under complete dark conditions. Measurements were taken daily for 28 days, and the results are presented at 4-day intervals. Germination was considered to occur when the length of the radicle was at least 2 mm.
2.3. Experiment 2: Effect of Temperature and Sucrose Concentration on Bulblet In Vitro Production
Pancratium maritimum seeds used in the previous experiment were also used in this experiment. The sterilization and transfer of the seeds to Petri dishes under aseptic conditions, as well as seed culture, were implemented according to the technique mentioned in Experiment 1. Then, Petri dishes were placed in an incubation chamber at 20 °C and under dark conditions for seed germination, as in Experiment 1.
Twenty days after the establishment of the seeds in vitro, only microplants with one leaf were selected (120 microplants at the one-leaf stage out of the 129 in total, for uniformity) and transferred under aseptic conditions to Petri dishes with two different culture media for bulb production (e.g., in vitro bulblet production in two stages) [9] (Figure 1A–D). Each microplant was transferred to a Petri dish containing MS (Murashige and Skoog M5519 basal mixture, Sigma-Aldrich, USA) modified culture medium with agar (Serva, Germany) at a concentration of 4 g/L and sucrose (Serva, Germany) at a concentration of 40 g/L or 80 g/L (e.g., bulblet production medium). In all culture media, the pH was adjusted to 5.75. For each sucrose concentration (e.g., 40 g/L or 80 g/L), 60 microplants were used (120 microplants in total). Moreover, twenty Petri dishes from each sucrose concentration were placed in three incubation chambers with different temperatures (10 °C, 20 °C, and 30 °C), while the photoperiod was adjusted at 16 h light/8 h dark and light intensity at 45 μmol/m2/sec.

Figure 1.
(A) In vitro seed germination of Pancratium maritimum at 20 °C after 8 days, (B) seedlings at the stage of one leaf—20 days after the establishment of seeds in vitro, (C) seedling-bulblet development in culture medium with sucrose 40 g/L at 10 °C after 72 days, (D) seedling-bulblet development in culture medium with sucrose 40 g/L at 30 °C after 72 days, (E) seedling-bulblet development in culture medium with sucrose 80 g/L at 20 °C after 72 days, (F) seedling-bulblet development in culture medium with sucrose 80 g/L at 30 °C after 72 days. Bars = 10 mm.
During the in vitro growth of the bulblet, the number of leaves, the length of the bulb, and the largest diameter of the bulblet were measured using an electronic caliper. Measurements started 18 days after the microplants establishment in the bulblet production culture media and were repeated at 18-day intervals until 72 days after the establishment (4 measurements in total). On the 72nd day, the fresh (F.W.) and dry weight (D.W.) of roots, leaves, and bulblets were measured. Dry matter content (D.M.C.) was recorded after drying the plant tissues at 72 °C to constant weight.
2.4. Experimental Design and Statistical Analysis
Experiment 1 was carried out in a completely randomized design with five treatments (n = 5). The data were subjected to a one-way ANOVA with the use of Statgraphics 5.1. plus (Statpoint Technologies, Inc., Warrenton, VA, USA), while when the F-test proved significant (p ≤ 0.05), means were compared by the method of Least Significant Difference (LSD) test.
Experiment 2 was carried out in a completely randomized experimental design with two factors (factor A: temperature level and factor B: sucrose concentration), with a total of six treatments (temperature level x sucrose concentration) and four replicates of five microplants each. As the interactions between these factors were statistically significant, the analysis of variance was carried out separately for the temperature levels at each sucrose concentration. For all parameters where the F-test proved significant (p ≤ 0.05), means were compared by the method of least significant difference (LSD). Comparisons of the two sucrose concentrations at each temperature level were made using t-tests.
3. Results
3.1. Experiment 1
Temperature significantly affected both the total percentage of germinated seeds and the rate of seed germination (Figure 2). The initiation of seed germination was favored at 20 °C, 25 °C, and 30 °C, with radicle appearance being recorded on day 4 after seed culture. In addition, on day 4, the percentage of germinated seeds was statistically significantly higher at 25 °C compared to 20 °C and 30 °C, whereas no seeds germinated at 10 °C and 15 °C. Moreover, for all the temperature treatments except for 10 °C, the germination percentage increase was very steep and reached approximately 70%, while at 15 °C, this steep increase continued up to day 8. Eight days after starting the experiment, seed germination percentage was also the highest at 25 °C, although no significant differences with 20 °C and 30 °C were recorded. On the other hand, 40% of seeds germinated at 15 °C, while no visible germination was observed at 10 °C. From day 8 onwards and until day 20, no significant differences were recorded between 20 °C, 25 °C, and 30 °C, while germination percentage at 30 °C did not differ significantly from that at 15 °C. After the 20th day and until the end of the experiment (28 days), the highest germination percentage was recorded at 20 °C, without being significantly different from the percentage at 25 °C. Moreover, no significant differences were detected in the germination rates between days 16 and 28 for 20 °C/25 °C. The temperature of 20 °C and 10 °C were the only ones where seed germination showed increasing trends until the end of the experiment, although the germination percentages at the latter conditions were the lowest overall. For the rest of the temperatures (15 °C, 25 °C, and 30 °C), germination percentages reached a plateau near the end of the experiment (20 to 24 days after the experiment started). After 28 days at 20 °C and 25 °C, the percentage of seeds with viable seedlings which could be used for in vitro propagation was 82% and 77%, respectively.
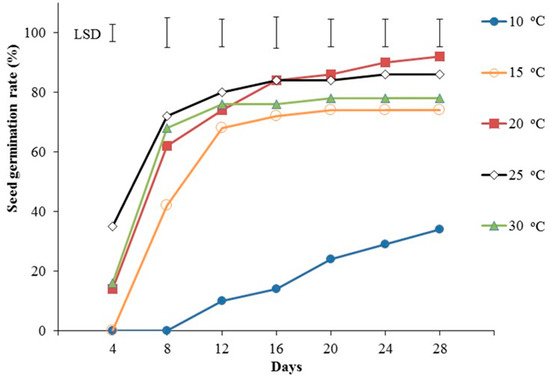
Figure 2.
Effect of temperature (10 °C, 15 °C, 20 °C, 25 °C, and 30 °C) on Pancratium maritimum seed germination rate (%). Vertical bars = LSD (p ≤ 0.05). Four replicates of 25 seeds per treatment.
3.2. Experiment 2
Temperature affected the number of leaves per microplant, which was statistically significantly lower at 10 °C (1 leaf on average) compared to 20 °C and 30 °C (2–3 leaves on average) throughout the experiment (72 days) when microplants were cultured in medium containing 40 g/L of sucrose (Figure 3). Moreover, no statistically significant differences in the average number of leaves per microplant between 20 and 30 °C were recorded, with the exception of day 54, where microplants cultured at 20 °C had a higher average number of leaves than those maintained at 30 °C. In the case of medium containing 80 g/L of sucrose, the average number of leaves did not differ between 20 °C and 30 °C or 30 °C and 10 °C on day 54 and 72, respectively, while the average number of leaves was significantly lower at 10 °C on day 18. Moreover, the lower sucrose content (40 g/L) had a beneficial effect on the average number of leaves at 20 °C and 30 °C, compared to the higher concentration (80 g/L), whereas no effects of sucrose content were recorded at the lowest tested temperature (10 °C). Finally, the highest overall number of leaves was recorded when microplants were cultured at 20 °C in a growing medium that contained 40 g/L (almost 3 leaves per microplant on average), although without significant differences with 30 °C.
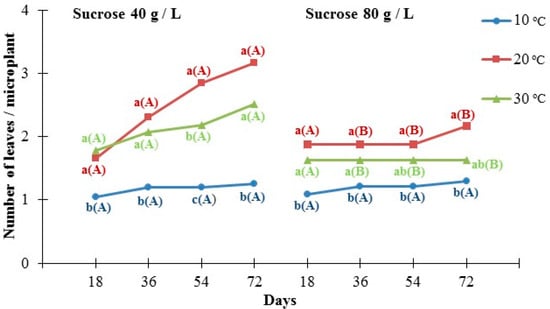
Figure 3.
The effect of two different concentrations of sucrose (40 g/L and 80 g/L) at three different temperatures (10 °C, 20 °C, and 30 °C) on the number of leaves per Pancratium maritimum microplant grown in culture media. In each sucrose concentration and each day of measurement, the same lowercase letters indicate non-statistically significant differences among the temperature treatments, according to LSD test (p ≤ 0.05). In each temperature and each day of measurement, the same uppercase letters in parenthesis indicate no statistically significant differences between sucrose concentrations, according to Student’s t-test (p ≤ 0.05). Four replicates of five explants for each treatment.
Regarding the effect of the temperature and the sucrose content of the culture medium on bulblet length, the bulblet length showed increasing trends with the progress of the experiment ranging between 10.4–11.5 mm and 10.8–12.1 mm on the 72nd day for 40 g/L and 80 g/L, respectively, without significant differences being recorded (data not shown). However, a varied response was recorded between the two sucrose concentrations, while the highest overall values of bulblet length for the medium containing 40 g/L or 80 g/L were recorded at 30 °C and 20 °C, respectively.
The diameter of the bulblet was significantly affected by both temperature and sucrose concentration in the culture medium, as shown in Figure 4. In the case of medium with a sucrose concentration of 40 g/L, microplants produced bulblets with a statistically significant higher diameter at 30 °C (6.4 mm on average) compared to 20 °C (4.1 mm on average) and 10 °C (3.6 mm on average). Similar results were recorded in the case of medium with the highest sucrose content (80 g/L), although no significant difference between 20 °C (5.4 mm on average) and 30 °C (6.2 mm on average) was detected. Moreover, when we compared the response of the microplants at 30 °C, no differences were observed in the diameter of the bulbs regardless of the sucrose concentration of the growing medium (40 or 80 g/L), whereas contrasting effects were observed at 20 °C and 10 °C, especially after 72 days of the experiment’s initiation.
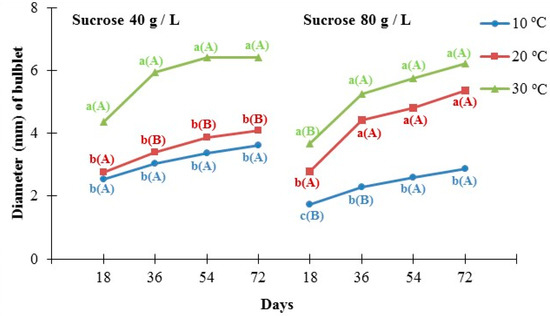
Figure 4.
Effect of two different concentrations of sucrose (40 g/L and 80 g/L) at three different temperatures (10 °C, 20 °C, and 30 °C) on the diameter of Pancratium maritimum bulblets produced in culture media In each sucrose concentration and each day of measurement, the same lowercase letters indicate non statistically significant differences among the temperature treatments, according to LSD test (p ≤ 0.05). In each temperature and each day of measurement, the same uppercase letters in parenthesis indicate no statistically significant differences between sucrose concentrations, according to Student’s t-test (p ≤ 0.05). Four replicates of five explants for each treatment.
The fresh weight (F.W.) of microplants was statistically significantly higher at 20 and 30 °C than that at 10 °C, regardless of the sucrose concentration in the culture medium, while the highest overall F.W. was recorded at 30 °C and sucrose concentration of 40 g/L (756.6 mg on average) (Figure 5A). Moreover, microplants grown in culture medium with a sucrose concentration of 40 g/L had statistically significantly higher F.W. compared to those cultured in medium with a sucrose concentration of 80 g/L, regardless of the temperature (Figure 5A). On the other hand, the F.W. of leaves was statistically significantly higher at 20 °C compared to 10 °C and 30 °C (154.8 mg on average in the case of medium with sucrose concentration 40 g/L) or compared to 30 °C (94.0 mg on average in the case of medium with sucrose concentration 80 g/L). Moreover, the fresh weight of leaves was higher for microplants cultured in medium with a sucrose concentration of 40 g/L than those cultured in medium with a sucrose concentration of 80 g/L, regardless of the temperature (Figure 5B). For the F.W. of roots, microplants formed very few roots when grown at 10 °C compared to 20 °C and 30 °C, regardless of the sucrose concentration, while the F.W. of roots was higher when the growing medium contained 40 g/L compared to 80 g/L. In particular, culture at 20 °C (230.2 mg at 40 g/L and 84.6 mg at 80 g/L) and 30 °C (305.1 mg at 40 g/L and 94.6 mg at 80 g/L) resulted in higher values than 10 °C (31.3 mg at 40 g/L and 26.8 mg at 80 g/L) for both sucrose concentrations. The highest overall F.W. was recorded at 20 °C and 30 °C for the medium with a sucrose content of 40 g/L (Figure 5C). Finally, the F.W. of bulblets was statistically significantly higher at 30 °C compared to 10 °C at 20 °C when the growth medium contained 40 g/L of sucrose (348.0 mg on average), while the lowest overall values were recorded at 10 °C for both sucrose concentrations. On the other hand, no significant differences between 20 °C and 30 °C were recorded in the case of sucrose content of 80 g/L (Figure 5D). Moreover, sucrose concentration did not significantly affect the F.W. of bulblets at 10 °C and 20 °C for both sucrose concentrations (40 g/L and 80 g/L), while growing at 30 °C and 40 g/L resulted in the highest overall F.W. values.
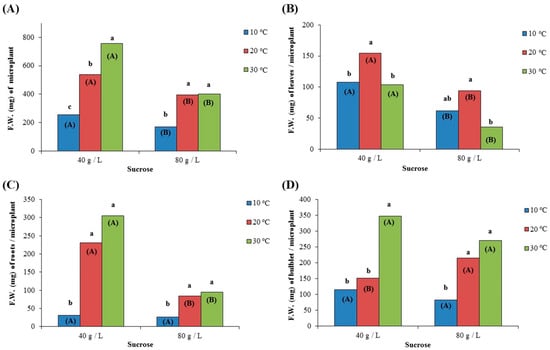
Figure 5.
Effect of two different concentrations of sucrose (40 g/L and 80 g/L) at three different temperatures (10 °C, 20 °C, and 30 °C) on the fresh weight (F.W. expressed mg) of (A) microplant; (B) leaves per microplant; (C) roots per microplant; and (D) bulblet per microplant, after growing for 72 days in culture medium. In each sucrose concentration, the same lowercase letters indicate non-statistically significant differences among the temperature treatments, according to the LSD test (p ≤ 0.05). In each temperature, the same uppercase letters in parenthesis indicate no statistically significant differences between sucrose concentrations, according to Student’s t-test (p ≤ 0.05).
The effect of temperature and sucrose concentration in the culture medium on the total D.W. of microplants, as well as the D.W. of leaves, roots per microplant, and bulblet per microplant, followed the same trends as in the case of F.W. (Figure 6A–D). The highest value of the D.W. for the whole microplant was recorded at 30 °C in the culture medium with a sucrose concentration of 40 g/L (118.7 mg on average), while the microplants with the highest leaf dry weight were those that were cultivated in a medium with 40 g/L of sucrose and incubated at 20 °C (30.2 mg on average). On the other hand, the microplants with the highest root dry weight were those that were cultivated in a medium with 40 g/L of sucrose and incubated at 20 °C (36.3 mg on average). In the case of the D.W. of bulblet per microplant, the highest values were recorded at 30 °C in the culture medium with 40 g/L (59.1 mg on average) and in the culture medium with 80 g/L (56.3 mg on average).
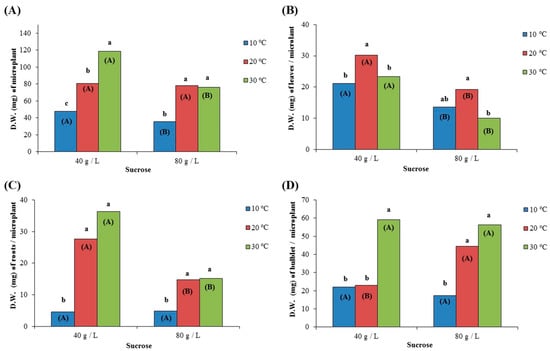
Figure 6.
Effect of two different concentrations of sucrose (40 g/L and 80 g/L) at three different temperatures (10 °C, 20 °C, and 30 °C) on the dry weight (D.W. expressed in mg) of (A) microplant; (B) leaf blades per microplant; (C) roots per microplant; and (D) bulblet per microplant, after growing for 72 days in culture media. In each sucrose concentration, the same lowercase letters indicate non-statistically significant differences among the temperature treatments, according to the LSD test (p ≤ 0.05). In each temperature, the same uppercase letters in parenthesis indicate no statistically significant differences between sucrose concentrations, according to Student’s t-test (p ≤ 0.05).
Dry matter content (D.M.C.) of leaves was not significantly affected in the case of microplants cultured in medium with 40 g/L (19.53–22.56%), while it was statistically significantly higher at 30 °C (28.04% on average) compared to 10 °C (22.10% on average) and 20 °C (20.46% on average) in the case of those growing in a medium with 80 g/L of sucrose (Figure 7A). Sucrose concentration had a significant effect on the D.M.C. of leaves only when microplants were cultured at 30 °C, where the incerased concentration of sucrose (80 g/L) resulted in a significant increase in the leaves’ D.M.C. (Figure 7A). Regarding the roots and bulblets, temperature did not significantly affect D.M.C. regardless of sucrose concentration (Figure 7B,C). On the other hand, the higher concentration of sucrose (80 g/L) in the culture medium resulted in a significant increase in the D.M.C. of roots (16.00–18.29%) for all the tested temperatures (Figure 7B), as well as in a significant increase in the D.M.C. of bulblets at 20 °C (Figure 7C).
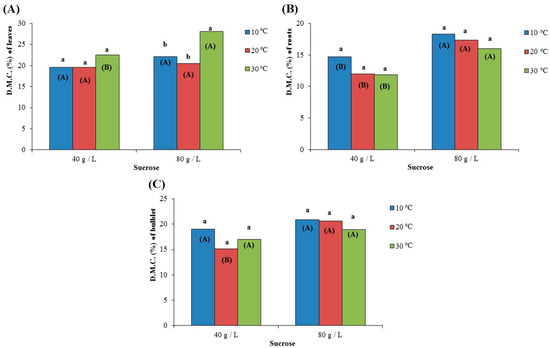
Figure 7.
Effect of two different concentrations of sucrose (40 g/L and 80 g/L) at three different temperatures (10 °C, 20 °C, and 30 °C) on the dry matter content (D.M.C. expressed as %) of (A) leaves, (B) roots and (C) bulblet, after growing for 72 days in culture media. In each sucrose concentration, the same lowercase letters indicate non-statistically significant differences among the temperature treatments, according to the LSD test (p ≤ 0.05). In each temperature, the same uppercase letters in parenthesis indicate no statistically significant differences between sucrose concentrations, according to Student’s t-test (p ≤ 0.05).
4. Discussion
High germination rates are essential for the sexual propagation of species, while further valorization of unexploited and endangered species, such as Pancratium maritimum, is associated with the establishment of germination protocols that ensure increased rates. The results from Experiment 1 indicate that the seed germination rate of P. maritimum was higher than 70% at temperatures between 15 °C and 30 °C. In particular, the percentage of seed germination was higher at 20 and 25 °C (92% and 86%, respectively). Similar seed germination rates have been reported by other researchers who used fresh mature seeds for germination either under in vitro [9,29,36] or in vivo conditions [40,41]. These findings are very important for the conservation of the genetic material of the plant as well as for the use of the seed as a starting material for in vitro cultures and the production of seedlings on a large scale. In our study, the high germination rates of P. maritimum at 20 °C and 25 °C (in the dark) were observed 28 days after the establishment of seeds in in vitro conditions. Similar germination rates of P. maritimum seeds were observed 30 days for in vitro germination by Yasemin et al. [29], also under dark conditions. Nikopoulos and Alexopoulos [9], who used freshly harvested seeds from the same genetic material tested in our study, observed similar seed germination rates but at 60 days after the establishment of seeds in vitro. This delay in germination could be due to the presence of light during their experiment (16 h light/8 h dark), as well as to the lower temperatures tested (12 °C in the dark/16 °C in the light). According to the literature, light has been reported to cause a delay in the germination of P. maritimum seeds in in vitro [36] and in vivo conditions [41], while Delipetrou [42] suggested the presence of photoinhibition and the inhibitory role of phytochrome in the germination of P. maritimum seeds. Although in vitro seed germination of P. maritimum may occur over a wide range of temperatures (10–30 °C), the fastest germination of seeds and the highest uniformity of seedlings and germination percentage were observed at temperatures between 20 °C and 25 °C. Moreover, the in vitro production of uniform seedlings is of great importance when applying the “two step” bulblet production method proposed by Nikopoulos and Alexopoulos [9]. Therefore, according to our results, temperatures between 20 °C and 25 °C and dark conditions proved to be the most suitable condition for faster and higher production of uniform seedlings.
Temperature is an important factor for plant growth in in vitro cultures [43]. In Experiment 2, following the suggestion for a “two-step” bulblet production method, microplants were grown in culture medium with varied sucrose content (40 g/L and 80 g/L) and three different temperatures (10 °C, 20 °C, and 30 °C). The obtained results indicated that the formation of leaves was inhibited at 10 °C, while it was favored at 20 °C. This trend was also reflected in leaf F.W. and D.W. In addition, microplant growth and bulblet production were inhibited at 10 °C, while the increase in bulblet diameter and the F.W. (and D.W. of bulblets were favored at 30 °C (in the case of sucrose concentration at 40 g/L) or at 20 and 30 °C (in the case of sucrose concentration at 80 g/L). Similarly, high temperatures (30 °C) favored root growth, especially in roots cultured in medium with a sucrose concentration of 40 g/L, as well as the D.M.C. of leaves cultured in medium with a sucrose concentration of 40 g/L. Moreover, the effect of temperature on the F.W. and D.W. of the different parts (roots, leaves, bulbs) of P. maritimum microplants showed a varied response. These observations indicate that temperature may have a regulatory role in plant growth and in morphogenesis during the in vitro propagation of the species [37,44].
Regarding the effect of sucrose concentration in the culture medium, it was observed that the higher concentration tested (80 g/L) did not favor the formation of leaves in microplants both at 20 °C and 30 °C. In addition, the F.W. and D.W. of leaves, roots, and whole microplants were lower in culture media with the highest sucrose concentration (80 g/L). Ptak et al. [45] have associated high sucrose concentrations in the culture medium with the increased biosynthesis of metabolites in in vitro conditions, which usually occurs under stress conditions, suggesting that high sucrose content may play an inhibitory role in microplant growth, thus the lower F.W and D.W. values recorded in our study. In contrast, high sucrose concentration (80 g/L) favored bulblet growth at 20 °C but did not affect bulblet growth at 30 °C, as shown by the F.W. and the D.W. of bulblets (Figure 6D and Figure 7D). Therefore, it seems that the effect of sucrose concentration on in vitro bulblet production of P. maritimum is temperature dependent. Many researchers have already reported that high concentrations of sucrose in the culture medium may favor the in vitro production of bulblets in P. maritimum [28] and other plant species, such as Lilium Asiatic hybrids [46] or Narcissus confusus and N. asturiensis [34,47]. However, in many cases, the positive effect of high sucrose concentrations on in vitro bulblet production was also associated with the use of plant growth regulators, such as BAP and NAA in P. maritimum [9], BAP and CCC in Hippeastrum hybridum [48], and paclobutrazol in Leucojum aestivum [45]. On the other hand, Azandi and Khosh–Khui et al. [49] reported that the in vitro bulblet production in Lilium ledebourii was favored when the sucrose concentration in the culture medium was low (e.g., 30 g/L).
Wu et al. [50] reported that bulb formation involves a shoot-to-bulblet transition, while Gao et al. [51] attributed a specific signaling role for bulblet development to sucrose. Additionally, Wu et al. [50] suggested that during the in vitro bulblet formation in Lily “Sorbonne”, sucrose synthase and not invertase had a dominant role in sucrose metabolism. In the case of P. maritimum, bulbs have a high starch content [52], and changes in sucrose metabolism may play an important role in bulblet development in vitro. Thus, the effect of sucrose concentration on the activity of sucrose-metabolizing enzymes may also justify the interaction of sucrose concentration in culture medium and temperature observed in this work since enzyme activity is also affected by temperature [37].
5. Conclusions
It is concluded that P. maritimum seeds may germinate in high percentages (over 70%) in a wide range of temperatures (15–30 °C), but when aiming for massive bulblet production in vitro, the temperature of 20 °C (or at most up to 25 °C) is recommended since it allowed faster and more uniform seed germination. Moreover, our results suggest transferring microplants to a bulbing culture medium (“two-step bulblet formation”) containing sucrose at 40 g/L and at 30 °C. These particular conditions may allow the development of bulblets of great weight (over 300 mg) and with very well-developed roots from the cultured microplants in a short period of time (about 70–80 days after transfer). Both characteristics are an important advantage when plants are intended to be transplanted in soil since they result in higher survival rates. Additionally, bulblets can be successfully used to obtain twin scales and produce new plants in vitro. However, the production of bulblets in higher sucrose concentration (80 g/L) at 20 °C or 30 °C could also be an alternative approach, mainly for the in vitro production of bulblets from which substances with medicinal properties could be isolated. Therefore, in vitro propagation could be a useful technique not only for seedling production and further commercial cultivation of the species but also for the isolation of bioactive compounds through the mass production of bulblets. Moreover, eliciting during bulblet production may also induce the biosynthesis and isolation of target compounds and further facilitate the valorization of this valuable and underexplored species.
Author Contributions
Conceptualization, A.A.A.; methodology, A.A.A., E.M. and E.K.; formal analysis, A.A.A.; investigation, E.M.; data curation, A.A.A.; writing—original draft preparation, A.A.A., E.K. and S.A.P.; writing—review and editing, A.A.A., E.K. and S.A.P.; visualization, A.A.A.; supervision, A.A.A.; project administration, A.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Halophytic herbs of the Mediterranean basin: An alternative approach to health. Food Chem. Toxicol. 2018, 114, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Karkanis, A.; Polyzos, N.; Kompocholi, M.; Petropoulos, S.A. Rock Samphire, a Candidate Crop for Saline Agriculture: Cropping Practices, Chemical Composition and Health Effects. Appl. Sci. 2022, 12, 737. [Google Scholar] [CrossRef]
- Petropoulos, S.; Ntatsi, G.; Levizou, E.; Barros, L.; Ferreira, I. Nutritional profile and chemical composition of Cichorium spinosum ecotypes. LWT Food Sci. Technol. 2016, 73, 95–101. [Google Scholar] [CrossRef]
- Abogadallah, G.M. Differential regulation of photorespiratory gene expression by moderate and severe salt and drought stress in relation to oxidative stress. Plant Sci. 2011, 180, 540–547. [Google Scholar] [CrossRef]
- Blamey, M.; Grey-Wilson, C. Mediterranean Wild Flowers; Blamey, M., Grey-Wilson, C., Eds.; HarperCollins: New York, NY, USA, 1993. [Google Scholar]
- Giovino, A.; Domina, G.; Bazan, G.; Campisi, P.; Scibetta, S. Taxonomy and conservation of Pancratium maritimum (Amaryllidaceae) and relatives in the Central Mediterranean. Acta Bot. Gall. 2015, 162, 289–299. [Google Scholar] [CrossRef]
- Zahreddine, H.; Clubbe, C.; Baalbaki, R.; Ghalayini, A.; Talhouk, S.N. Status of native species in threatened Mediterranean habitats: The case of Pancratium maritimum L. (sea daffodil) in Lebanon. Biol. Conserv. 2004, 120, 11–18. [Google Scholar] [CrossRef]
- Nikopoulos, D.; Alexopoulos, A.A. In vitro propagation of an endangered medicinal plant: Pancratium maritimum L. J. Food Agric. Environ. 2008, 6, 393–398. [Google Scholar]
- Grassi, F.; Cazzaniga, E.; Minuto, L.; Peccenini, S.; Barberis, G.; Basso, B. Evaluation of biodiversity and conservation strategies in Pancratium maritimum L. for the Northern Tyrrhenian Sea. Biodivers. Conserv. 2005, 14, 2159–2169. [Google Scholar] [CrossRef]
- Sanaa, A.; Fadhel, N. Ben Genetic diversity in mainland and island populations of the endangered Pancratium maritimum L. (Amaryllidaceae) in Tunisia. Sci. Hortic. 2010, 125, 740–747. [Google Scholar] [CrossRef]
- De Castro, O.; Avino, M.; Di Maio, A.; Menale, B.; Guida, M. Sanger and next generation sequencing in the characterisation of arbuscular mycorrhizal fungi (AMF) in Pancratium maritimum L. (Amaryllidaceae), a representative plant species of Mediterranean sand dunes. Planta 2018, 248, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Berkov, S.; Pavlov, A.; Georgiev, V.; Weber, J.; Bley, T.; Viladomat, F.; Bastida, J.; Codina, C. Changes in apolar metabolites during in vitro organogenesis of Pancratium maritimum. Plant Physiol. Biochem. 2010, 48, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Leporini, M.; Catinella, G.; Bruno, M.; Falco, T.; Tundis, R.; Loizzo, M.R. Investigating the Antiproliferative and Antioxidant Properties of Pancratium maritimum L. (Amaryllidaceae) Stems, Flowers, Bulbs, and Fruits Extracts. Evid. Based Complement. Altern. Med. 2018, 2018, 9301247. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Shaala, L.A.; Youssef, D.T.A. Non-alkaloidal compounds from the Bulbs of the Egyptian plant Pancratium maritimum. Z. Für Nat. C 2014, 69, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Kekre, N.; Naderi, J.; McNulty, J. Induction of apoptotic cell death specifically in rat and human cancer cells by pancratistatin. Artif. Cells Blood Substit. Immobil. Biotechnol. 2005, 33, 279–295. [Google Scholar] [CrossRef] [PubMed]
- Cedrón, J.C.; Ravelo, Á.G.; León, L.G.; Padrón, J.M.; Estévez-Braun, A. Antiproliferative and structure activity relationships of Amaryllidaceae alkaloids. Molecules 2015, 20, 13854–13863. [Google Scholar] [CrossRef]
- Rokbeni, N.; M’rabet, Y.; Cluzet, S.; Richard, T.; Krisa, S.; Boussaid, M.; Boulila, A. Determination of phenolic composition and antioxidant activities of Pancratium maritimum L. from Tunisia. Ind. Crops Prod. 2016, 94, 505–513. [Google Scholar] [CrossRef]
- Sanaa, A.; Boulila, A.; Boussaid, M.; Fadhel, N. Ben Alginic acid and derivatives, new polymers from the endangered Pancratium maritimum L. Ind. Crops Prod. 2013, 44, 290–293. [Google Scholar] [CrossRef]
- Tayoub, G.; Al-Odat, M.; Amer, A.; Aljapawe, A.; Ekhtiar, A. Antiproliferative effects of Pancratium maritimum extracts on normal and cancerous cells. Iran. J. Med. Sci. 2018, 43, 52–64. [Google Scholar]
- Nair, J.J.; Van Staden, J. Antifungal activity based studies of Amaryllidaceae plant extracts. Nat. Prod. Commun. 2017, 12, 1953–1956. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Tzortzakis, N.; Sokovic, M.; Ciric, A.; Barros, L.; Ferreira, I.C.F.R. Bioactive compounds content and antimicrobial activities of wild edible Asteraceae species of the Mediterranean flora under commercial cultivation conditions. Food Res. Int. 2019, 119, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Correa, R.C.G.; Di Gioia, F.; Ferreira, I.; Petropoulos, S.A. Halophytes for future horticulture: The case of small-scale farming in the mediterranean basin. In Halophytes for Future Horticulture: From Molecules to Ecosystems towards Biosaline Agriculture; Grigore, M.-N., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 1–28. ISBN 9783030178543. [Google Scholar]
- Juan Vicedo, J. Pancratium Maritimum. The IUCN Red List of Threatened Species 2018. Available online: https://www.iucnredlist.org/species/18990540/57467022 (accessed on 20 October 2022).
- Sirin, U.; Kanmaz, E. In vitro propagation of sea daffodil (Pancratium maritimum L.) using seedling explants. Fresenius Environ. Bull. 2017, 25, 7710–7716. [Google Scholar]
- Georgiev, V.; Ivanov, I.; Pavlov, A. Obtaining and selection of Pancratium maritimum L. in vitro cultures with acetylcholinesterase inhibitory action. Biotechnol. Biotechnol. Equip. 2010, 24, 149–154. [Google Scholar] [CrossRef][Green Version]
- Georgiev, V.; Ivanov, I.; Berkov, S.; Pavlov, A. Alkaloids biosynthesis by Pancratium maritimum L. shoots in liquid culture. Acta Physiol. Plant. 2011, 33, 927–933. [Google Scholar] [CrossRef]
- Bogdanova, Y.; Pandova, B.; Yanev, S.; Stanilova, M. Biosynthesis of lycorine by in vitro cultures of Pancratium maritimum L. (Amaryllidaceae). Biotechnol. Biotechnol. Equip. 2009, 23, 919–922. [Google Scholar] [CrossRef]
- Yasemin, S.; Köksal, N.; Büyükalaca, S. Effects of Disinfection Conditions and Culture Media on in vitro Germination of Sea Daffodil (Pancratium maritimum). J. Biol. Environ. Sci. 2018, 12, 13–22. [Google Scholar]
- Dragassaki, M.; Economou, A.S.; Vlahos, J.C. Bulblet formation in vitro and plantlet survival extra vitrum in Pancratium maritimum L. Acta Hortic. 2003, 616, 347–352. [Google Scholar] [CrossRef]
- George, E.F. Plant tissue culture procedure—Background. In Plant Propagation by Tissue Culture, 3rd ed.; George, E.F., Hall, M.A., De Klerk, G.-J., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 1, pp. 1–28. ISBN 9781402050046. [Google Scholar]
- Slabbert, M.M.; de Bruyn, M.H.; Ferreira, D.I.; Pretorius, J. Regeneration of bulblets from twin scales of Crinum macowanii in vitro. Plant Cell Tissue Organ Cult. 1993, 33, 133–141. [Google Scholar] [CrossRef]
- Nikopoulos, D.; Nikopoulou, D.; Alexopoulos, A.A. Methods for the preservation of genetic material of Pancratium maritimum (Amaryllidaceae). J. Food Agric. Environ. 2008, 6, 538–546. [Google Scholar]
- Santos, A.; Fidalgo, F.; Santos, I.; Salema, R. In vitro bulb formation of Narcissus asturiensis, a threatened species of the Amaryllidaceae. J. Hortic. Sci. Biotechnol. 2002, 77, 149–152. [Google Scholar] [CrossRef]
- Panayotova, L.G.; Ivanova, T.A.; Bogdanova, Y.Y.; Gussev, C.V.; Stanilova, M.I.; Bosseva, Y.Z.; Stoeva, T.D. In vitro cultivation of plant species from sandy dunes along the Bulgarian Black Sea Coast. Phytol. Balc. 2008, 14, 119–123. [Google Scholar]
- Paradiso, R.; Buonomo, R.; De Pascale, S.; Cardarelli, M. Evaluation of spontaneous species for the innovation in floriculture: Pancratium maritimum L. as ornamental plant. Acta Hortic. 2010, 881, 563–566. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.; Murphy, A. Plant Physiology and Development; Sinauer Associates Publishers: Sunderland, MA, USA, 2015; p. 855. [Google Scholar]
- Cheesman, L.; Finnie, J.F.; Van Staden, J. Eucomis zambesiaca baker: Factors affecting in vitro bulblet induction. South Afr. J. Bot. 2010, 76, 543–549. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Benvenuti, S. Seed ecology of Mediterranean hind dune wildflowers. Ecol. Eng. 2016, 91, 282–293. [Google Scholar] [CrossRef]
- Balestri, E.; Cinelli, F. Germination and early-seedling establishment capacity of Pancratium maritimum L. (Amaryllidaceae) on coastal dunes in the north-western Mediterranean. J. Coast. Res. 2004, 20, 761–770. [Google Scholar] [CrossRef]
- Delipetrou, P. Ecophysiological Study of Seed Germination in Maritime Plants with Special Emphasis on the Effect of Light. Ph.D. Thesis, National and Kapodistrian University of Athens, Athens, Greece, 1996. [Google Scholar]
- Saadon, S.; Zaccai, M. Lilium candidum bulblet and meristem development. Vitr. Cell. Dev. Biol. Plant 2013, 49, 313–319. [Google Scholar] [CrossRef]
- Ascough, G.D.; Rice, L.J.; Van Staden, J. Considerations for evaluating flower abscission in potted plants with multiple inflorescences—Plectranthus as a case study. South Afr. J. Bot. 2008, 74, 753–756. [Google Scholar] [CrossRef][Green Version]
- Ptak, A.; Simlat, M.; Kwiecień, M.; Laurain-Mattar, D. Leucojum aestivum plants propagated in in vitro bioreactor culture and on solid media containing cytokinins. Eng. Life Sci. 2013, 13, 261–270. [Google Scholar] [CrossRef]
- Varshney, A.; Sharma, M.P.; Adholeya, A.; Dhawan, V.; Srivastava, P.S. Enhanced growth of micropropagated bulblets of Lilium sp. inoculated with arbuscular mycorrhizal fungi at different P fertility levels in an alfisol. J. Hortic. Sci. Biotechnol. 2002, 77, 258–263. [Google Scholar] [CrossRef]
- Sellés, M.; Viladomat, F.; Bastida, J.; Codina, C. Callus induction, somatic embryogenesis and organogenesis in Narcissus confusus: Correlation between the state of differentiation and the content of galanthamine and related alkaloids. Plant Cell Rep. 1999, 18, 646–651. [Google Scholar] [CrossRef]
- Sultana, J.; Sultana, N.; Siddique, M.N.A.; Islam, A.K.M.A.; Hossain, M.M.; Hossain, T. In vitro bulb production in Hippeastrum (Hippeastrum hybridum). J. Cent. Eur. Agric. 2010, 11, 469–474. [Google Scholar] [CrossRef]
- Azadi, P.; Khosh-Khui, M. Micropropagation of Lilium ledebourii (Baker) Boiss as affected by plant growth regulator, sucrose concentration, harvesting season and cold treatments. Electron. J. Biotechnol. 2007, 10, 582–591. [Google Scholar] [CrossRef]
- Wu, Y.; Ren, Z.; Gao, C.; Sun, M.; Li, S.; Min, R.; Wu, J.; Li, D.; Wang, X.; Wei, Y.; et al. Change in Sucrose Cleavage Pattern and Rapid Starch Accumulation Govern Lily Shoot-to-Bulblet Transition in vitro. Front. Plant Sci. 2021, 11, 564713. [Google Scholar] [CrossRef]
- Gao, S.; Zhu, Y.; Zhou, L.; Fu, X.; Lei, T.; Chen, Q.; Yu, X.; Zhou, Y.; Li, W.; Hu, J.; et al. Sucrose signaling function on the formation and swelling of bulblets of Lilium sargentiae E.H. Wilson. Plant Cell Tissue Organ Cult. 2018, 135, 143–153. [Google Scholar] [CrossRef]
- Pouris, J.; Meletiou-Christou, M.S.; Chimona, C.; Rhizopoulou, S. Seasonal functional partitioning of carbohydrates and proline among plant parts of the sand daffodil. Agronomy 2020, 10, 539. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).