
Competitiveness of Early Vigour Wheat (Triticum aestivum L.) Genotypes Is Established at Early Growth Stages

 , ,
, , 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm
2.2. Field Site and Climate
2.3. Experimental Design and Crop Management
2.4. Crop Assessments and Data Collection
2.5. Statistical Analyses and Modelling
3. Results
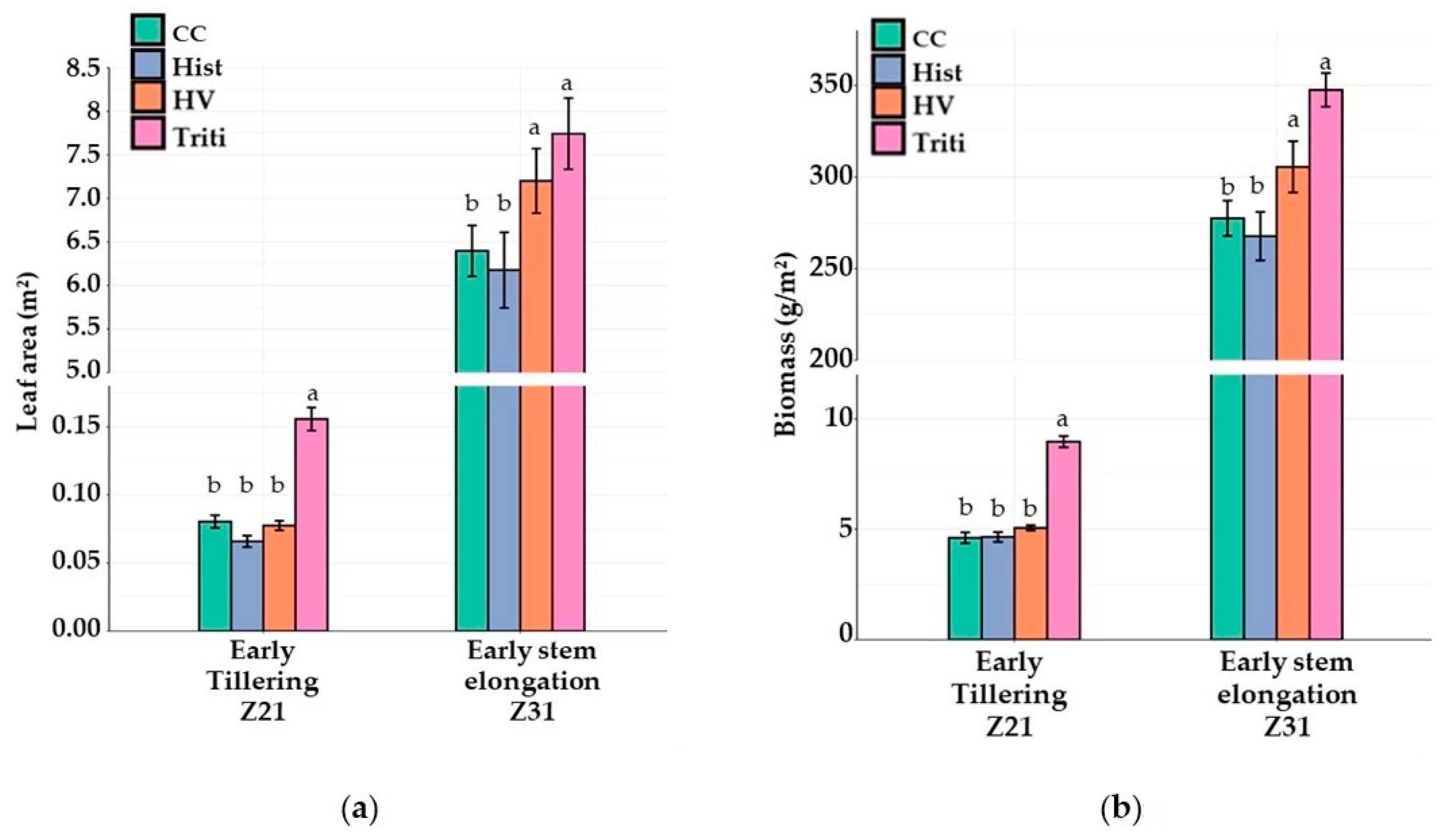
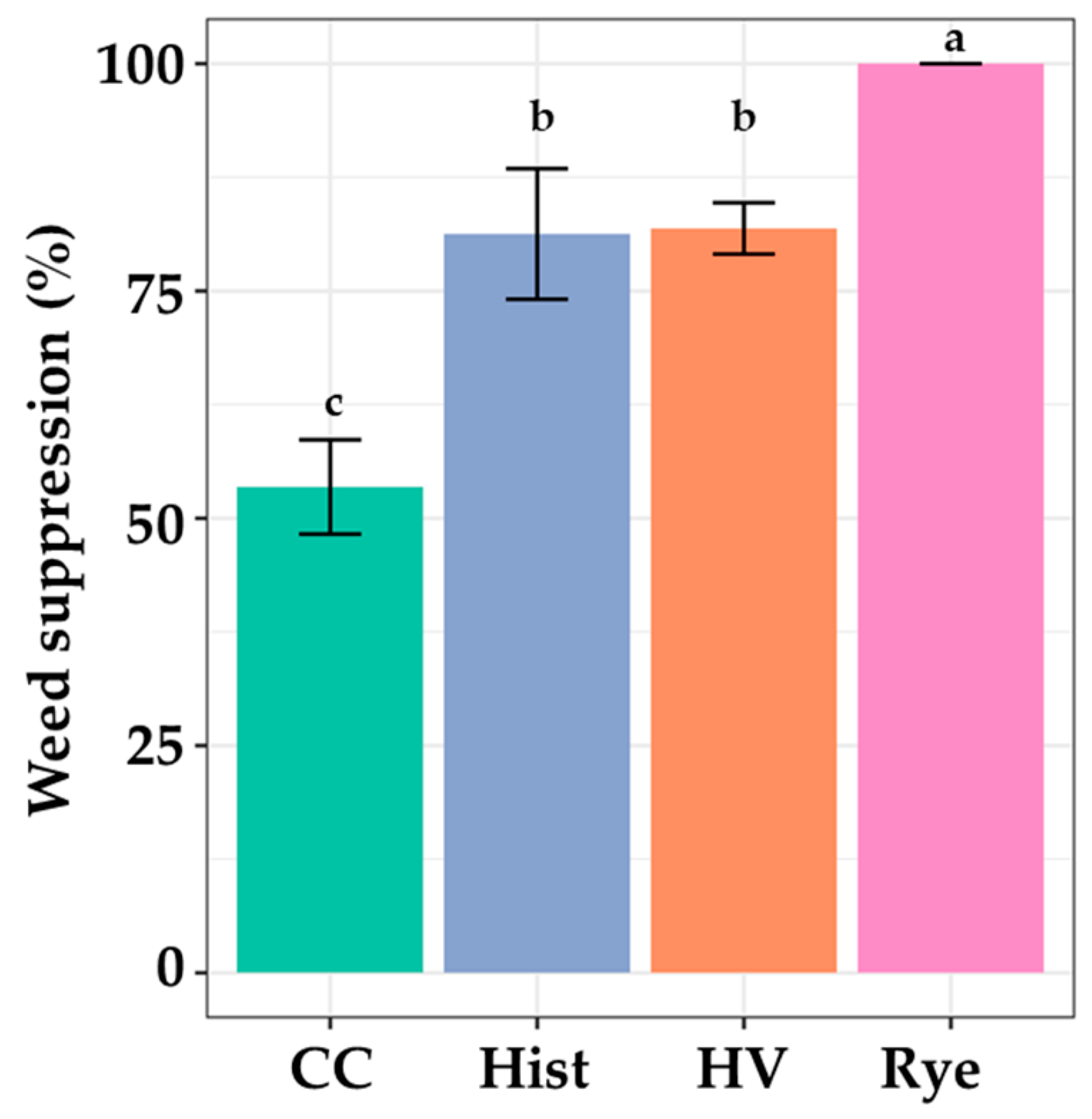
3.1. The 2018 Field Trial
3.2. The 2019 Field Trial
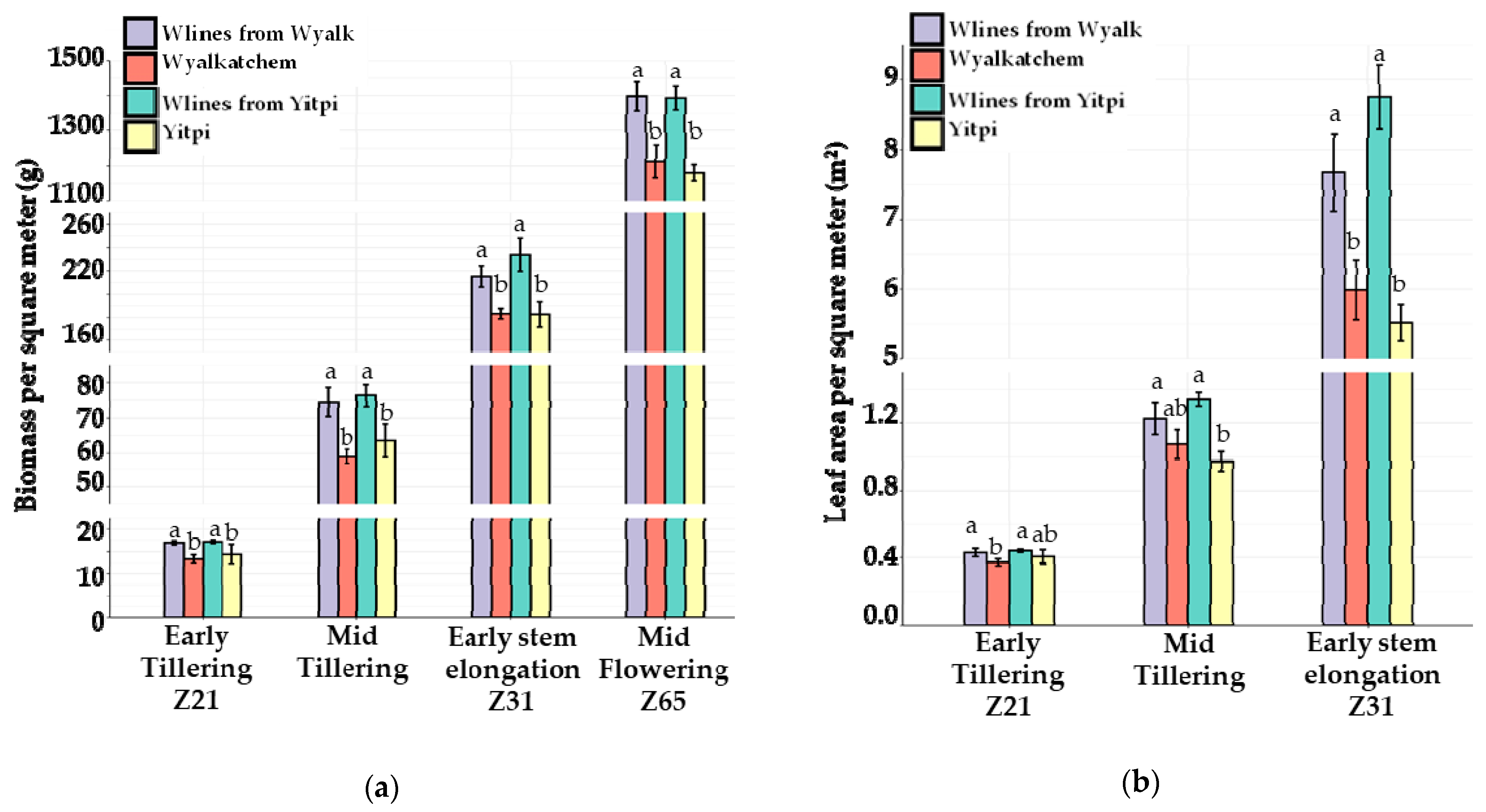
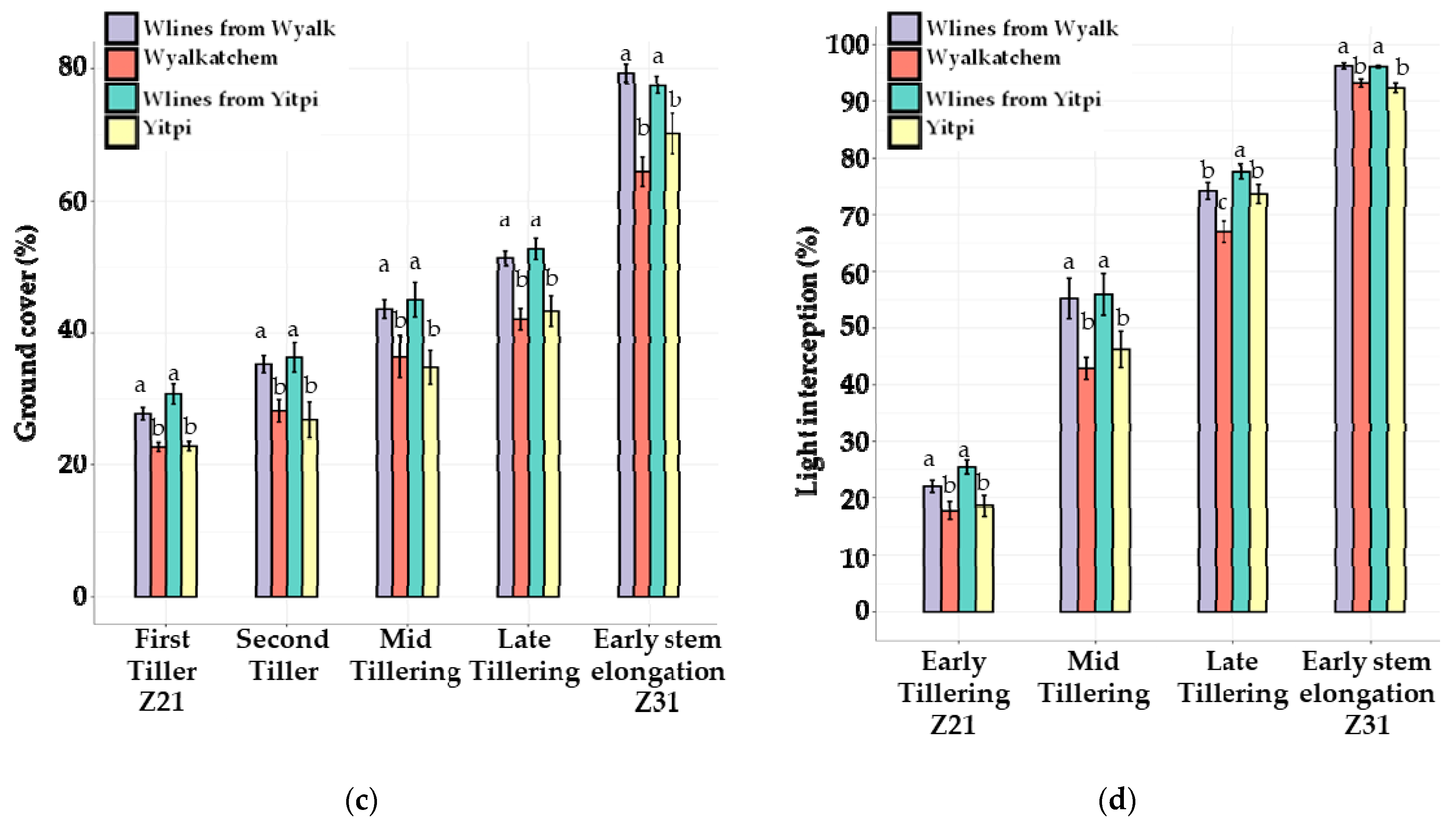
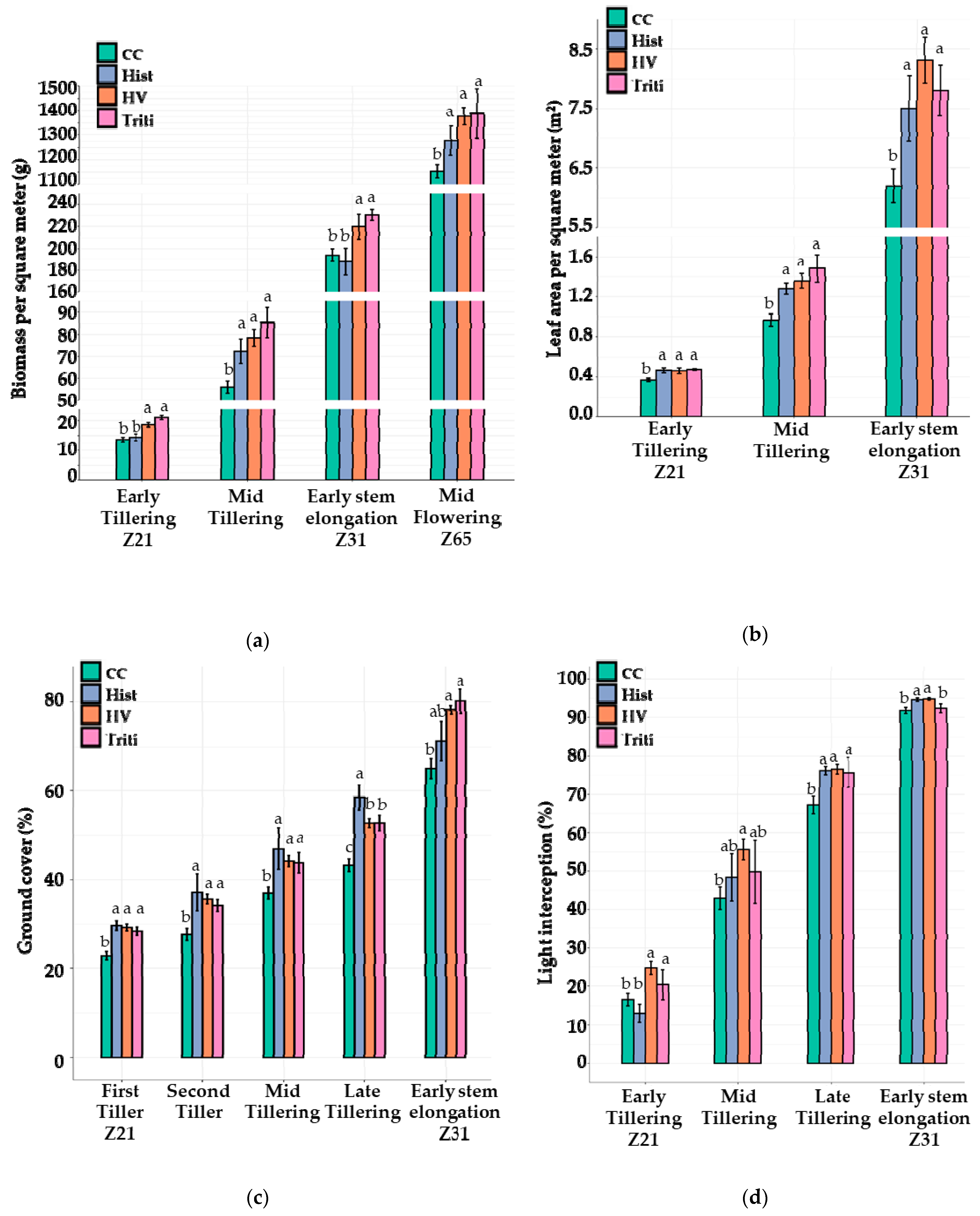
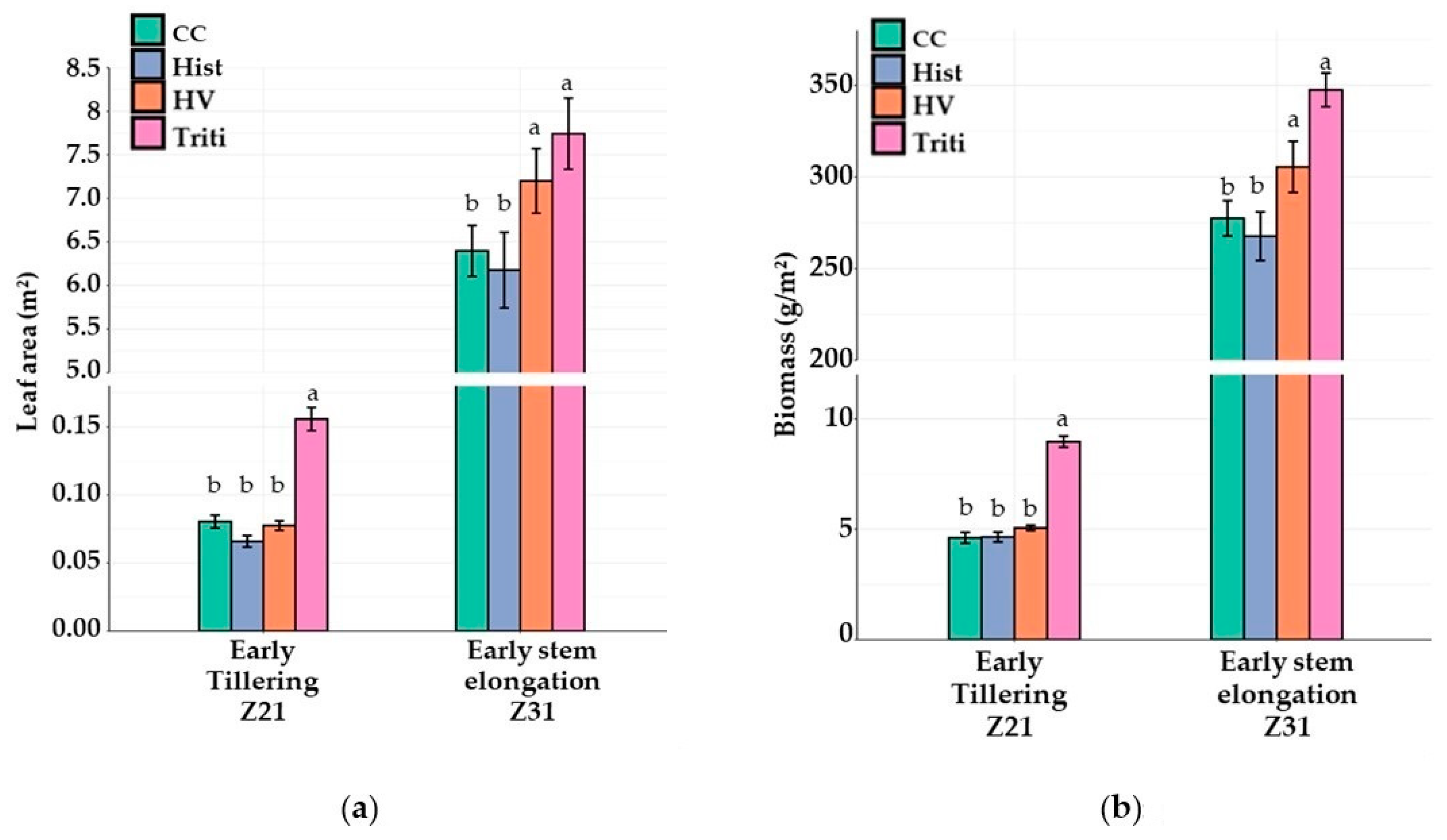
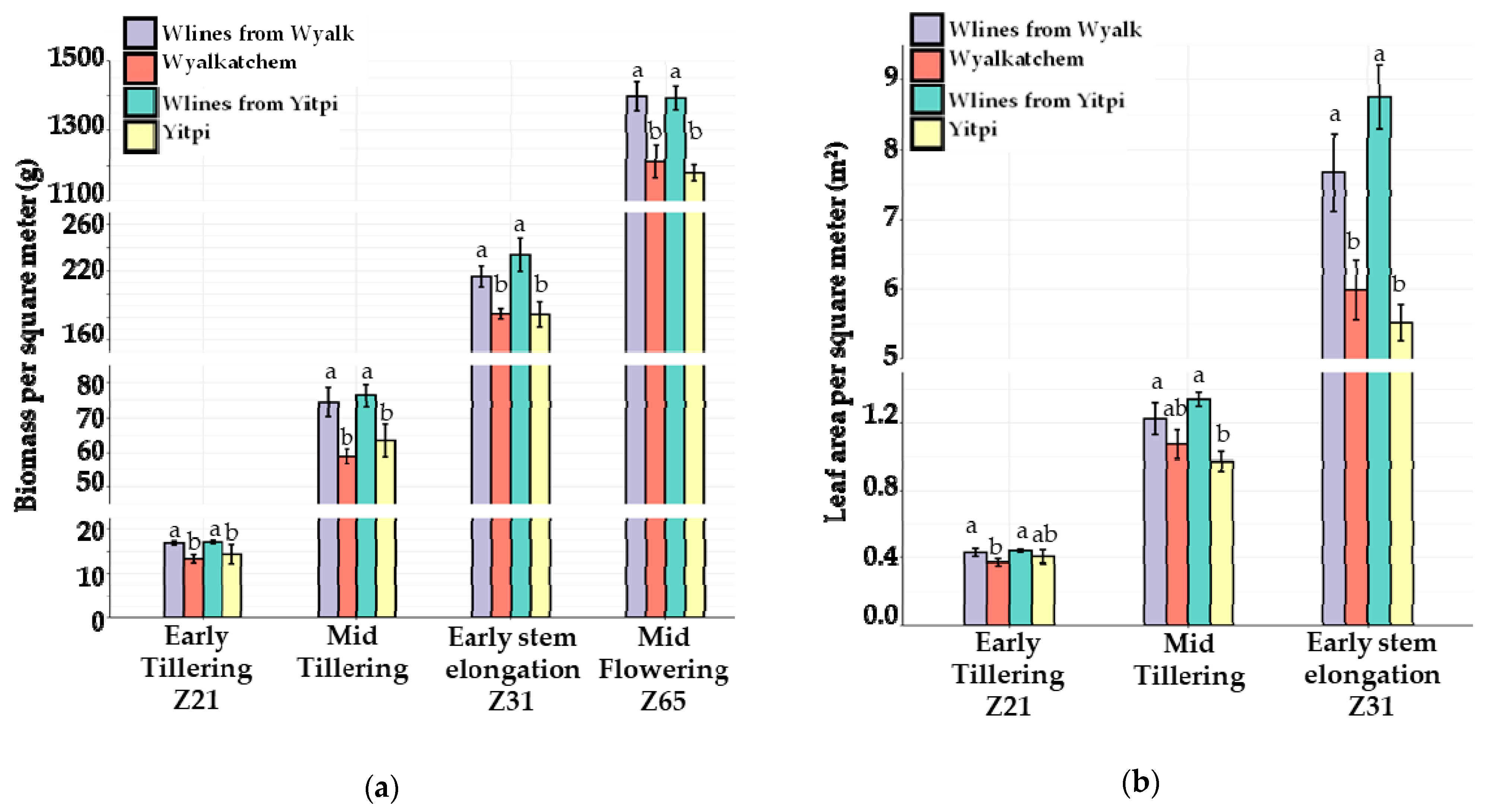
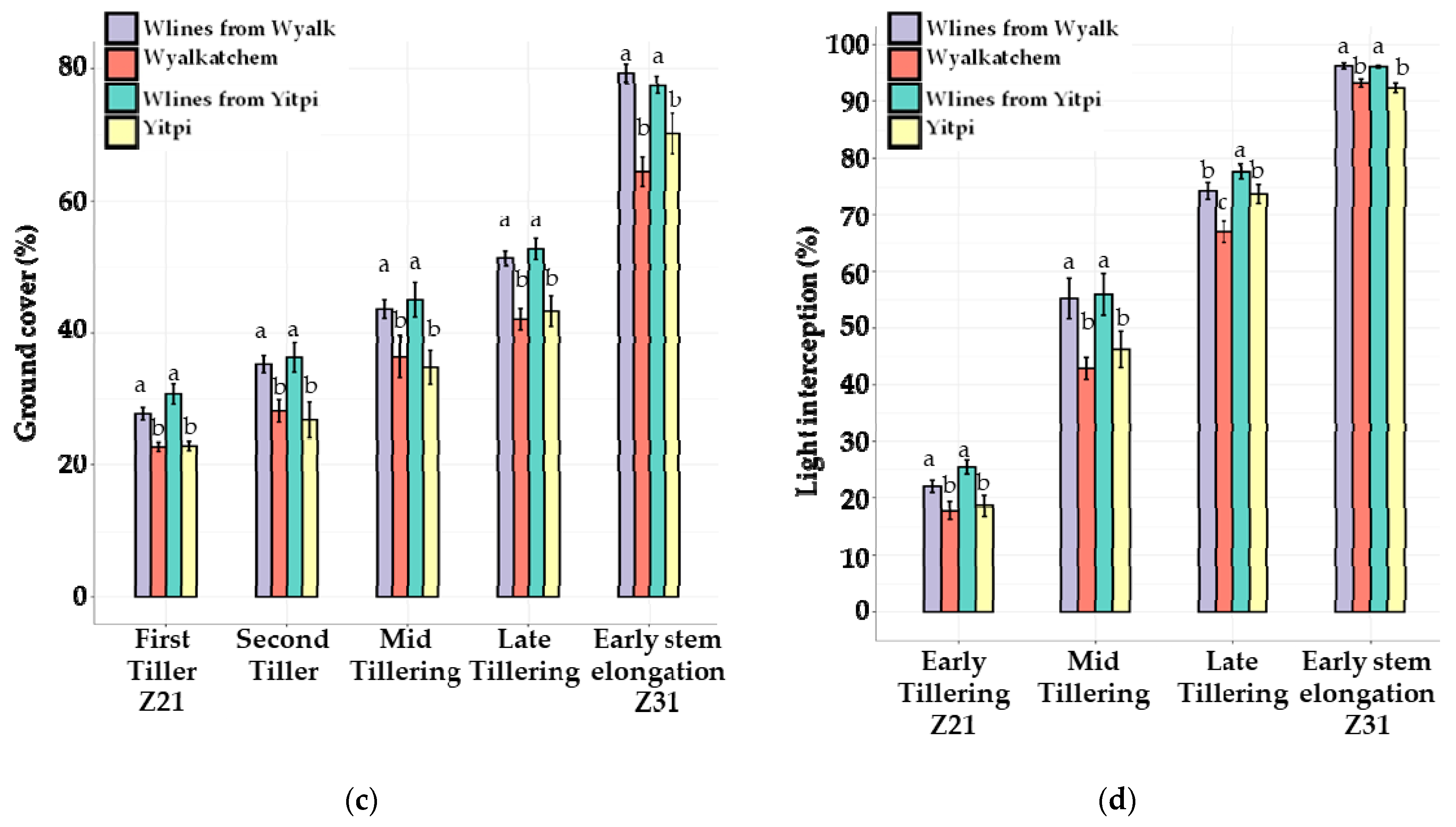
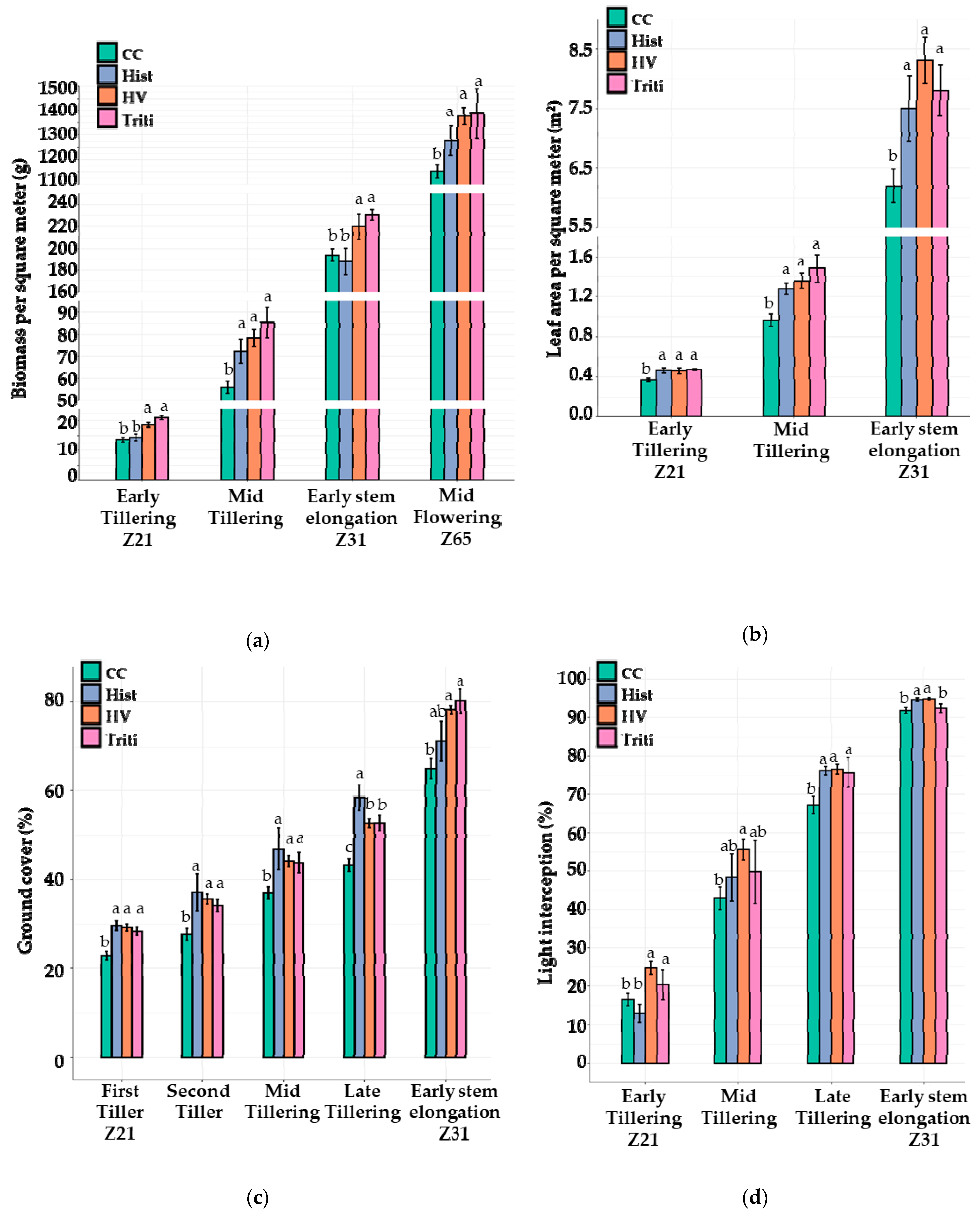
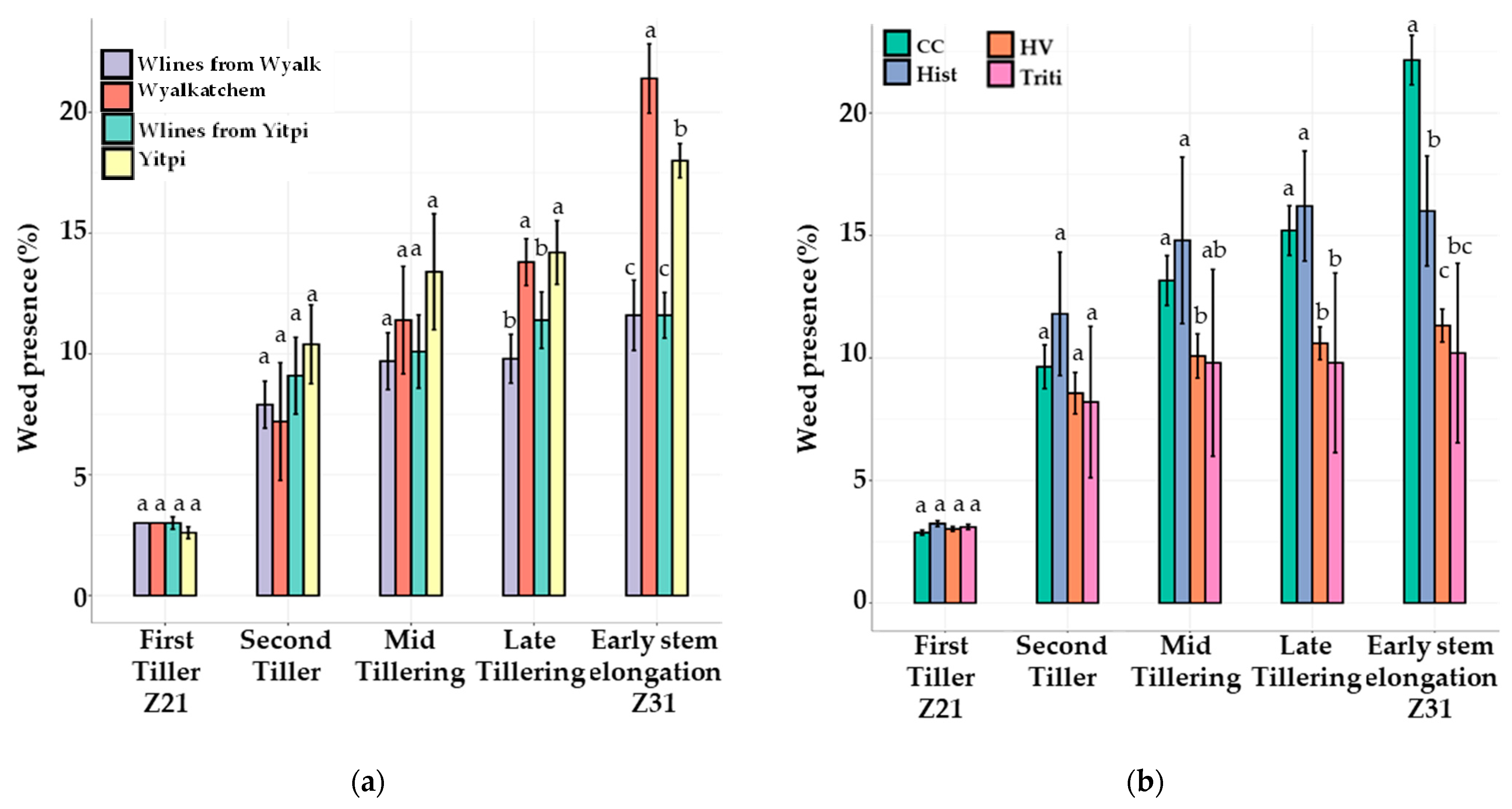
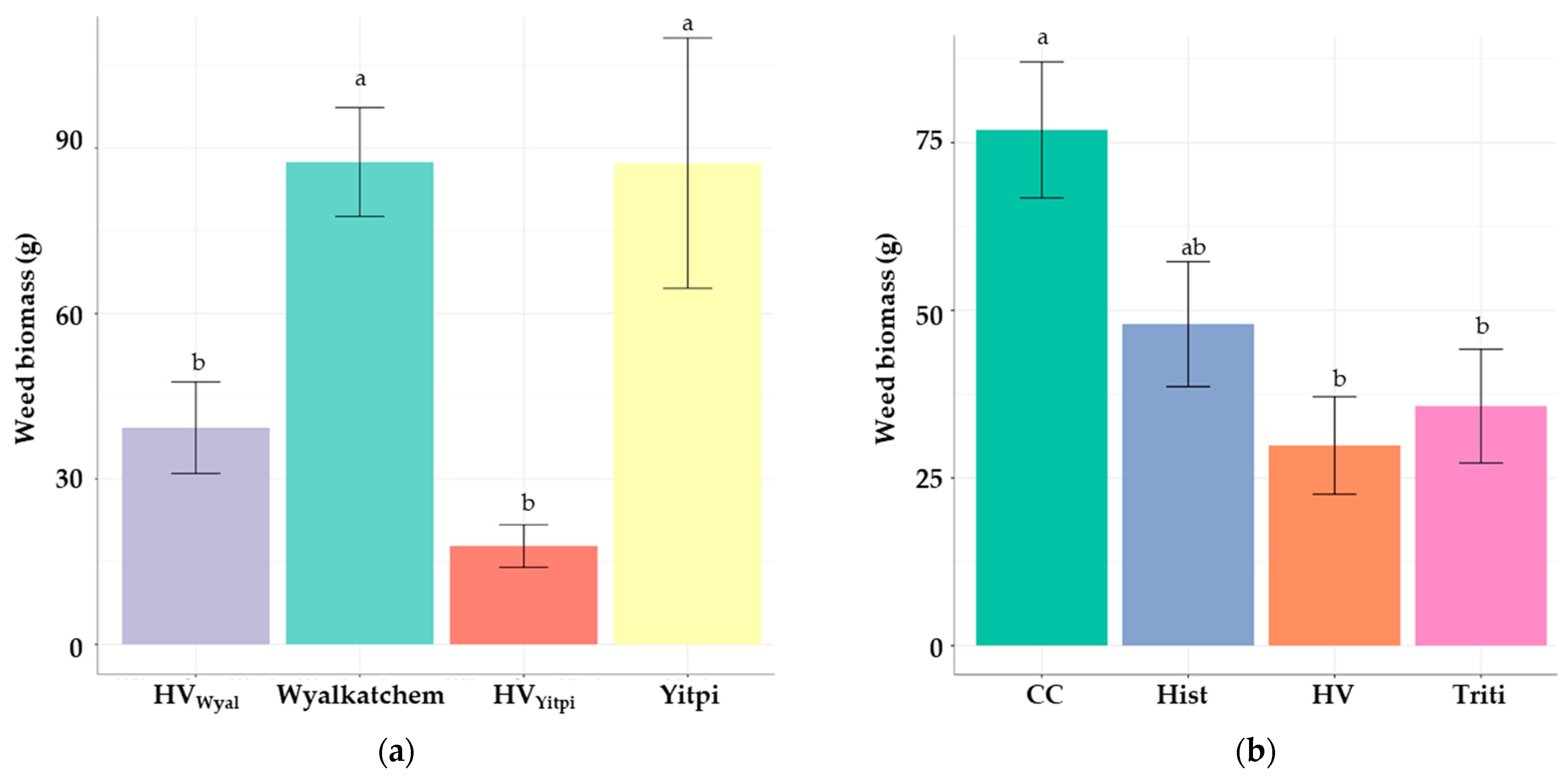
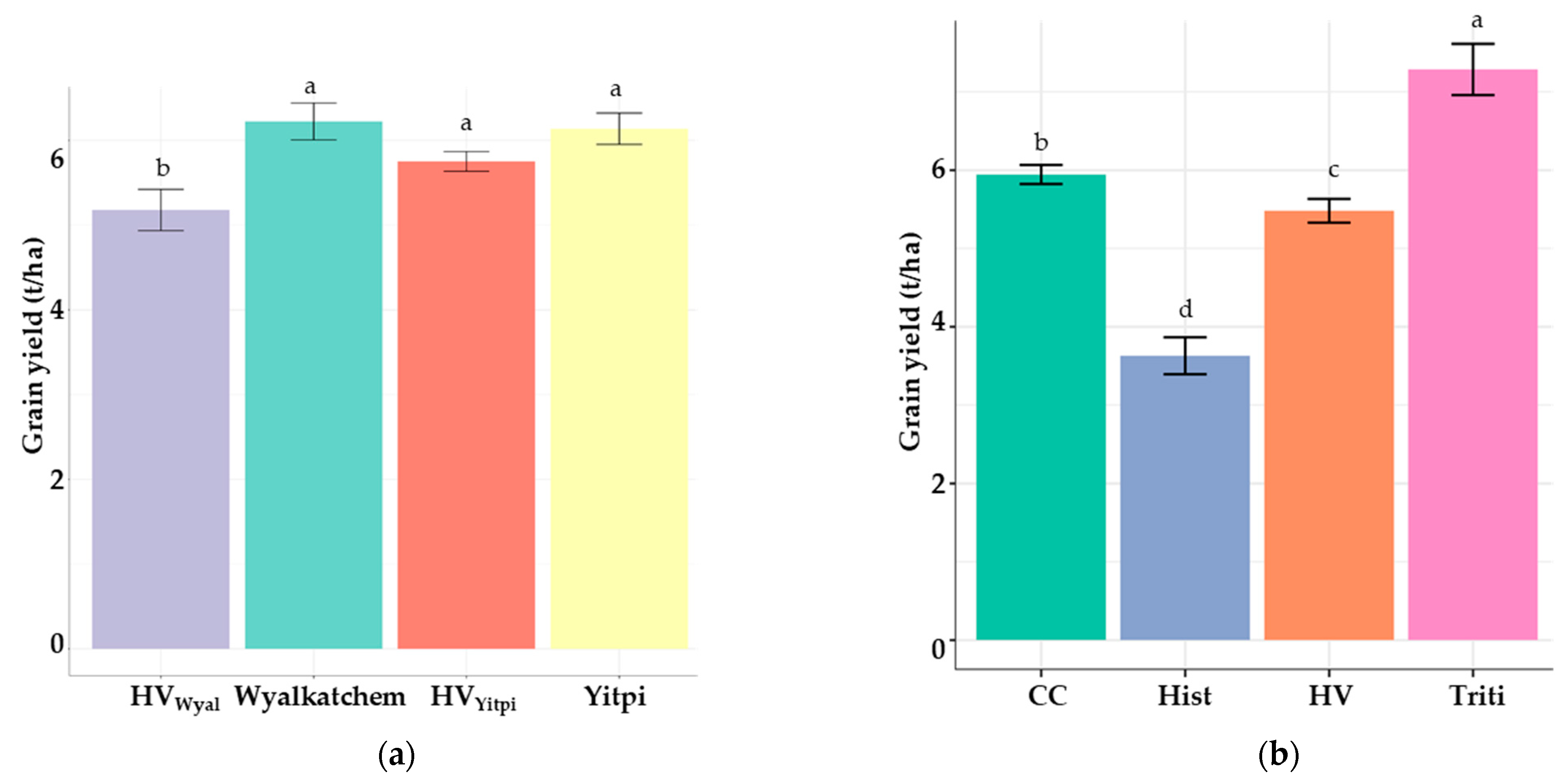
3.3. The 2020 Field Trial
3.3.1. Canopy Growth and Light Interception
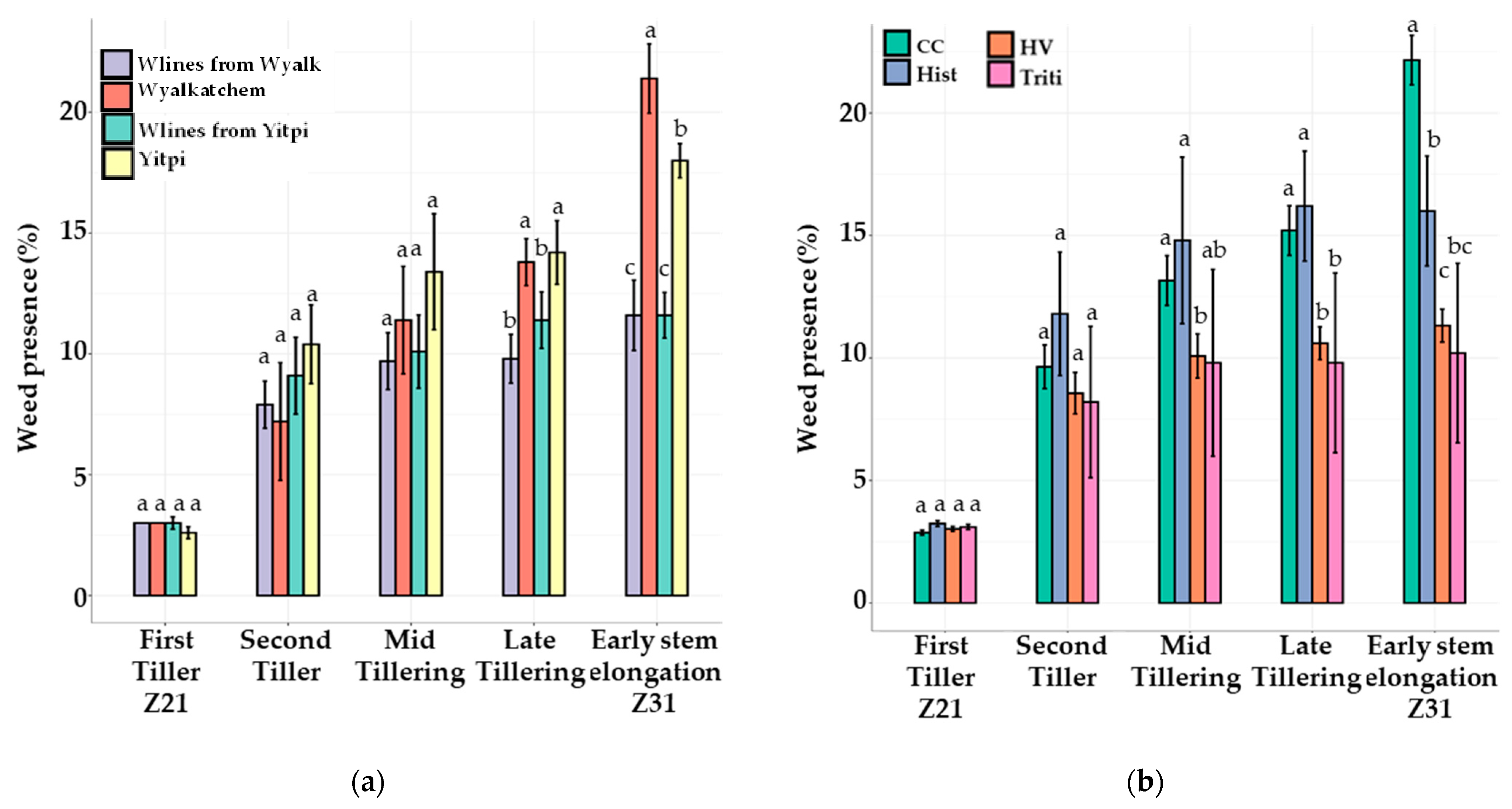
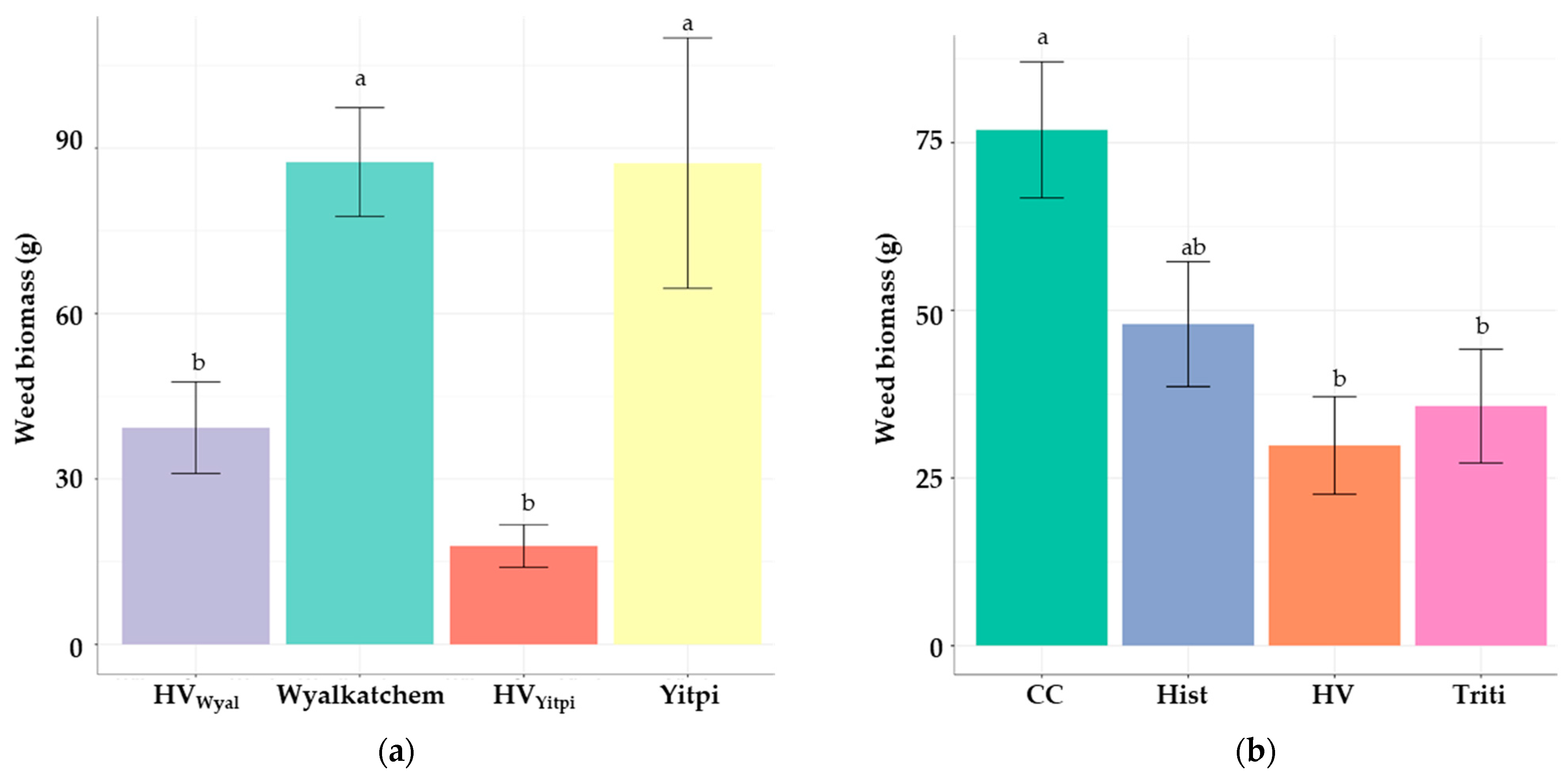
3.3.2. Weed Pressure
3.3.3. Yields
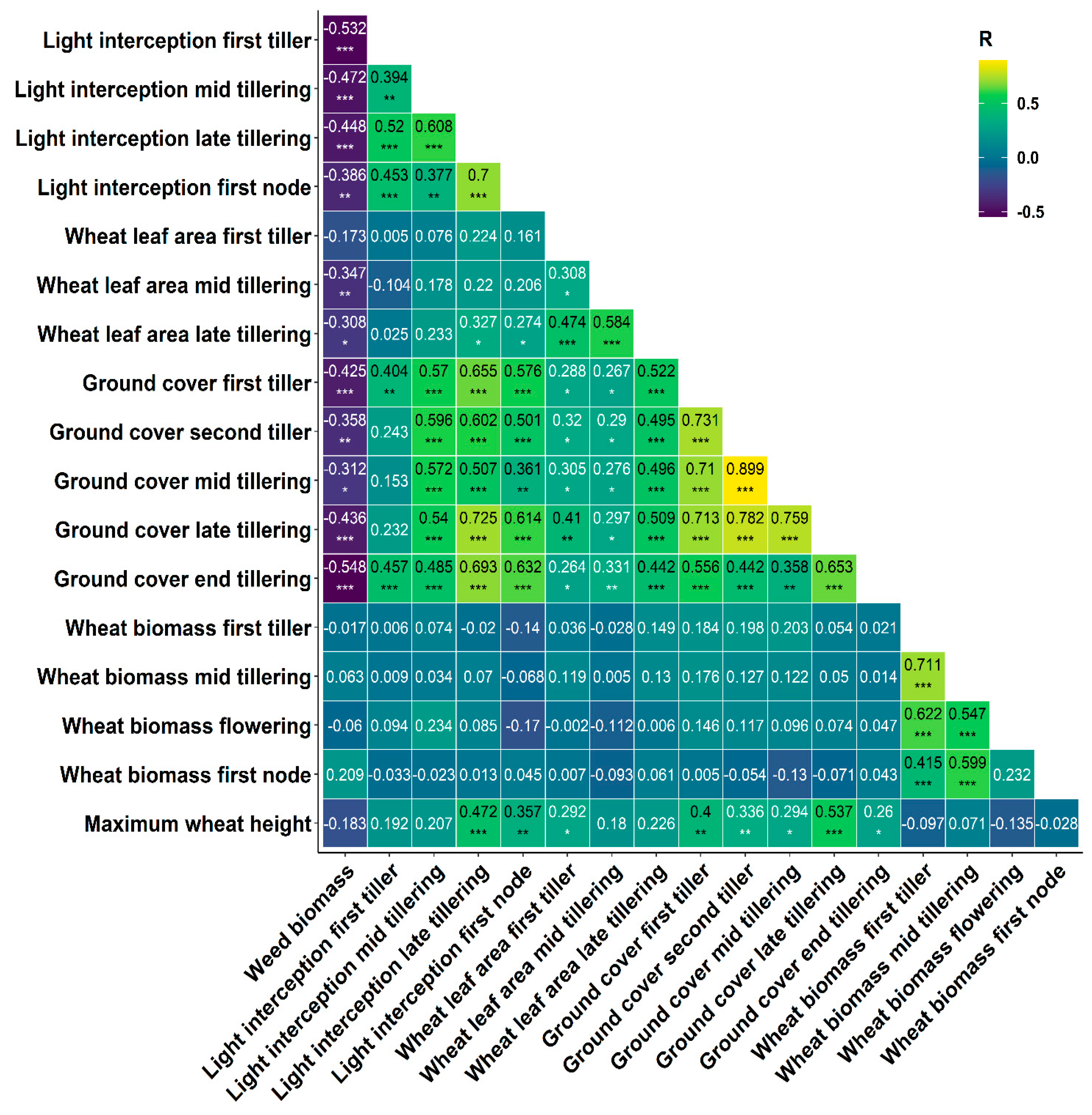
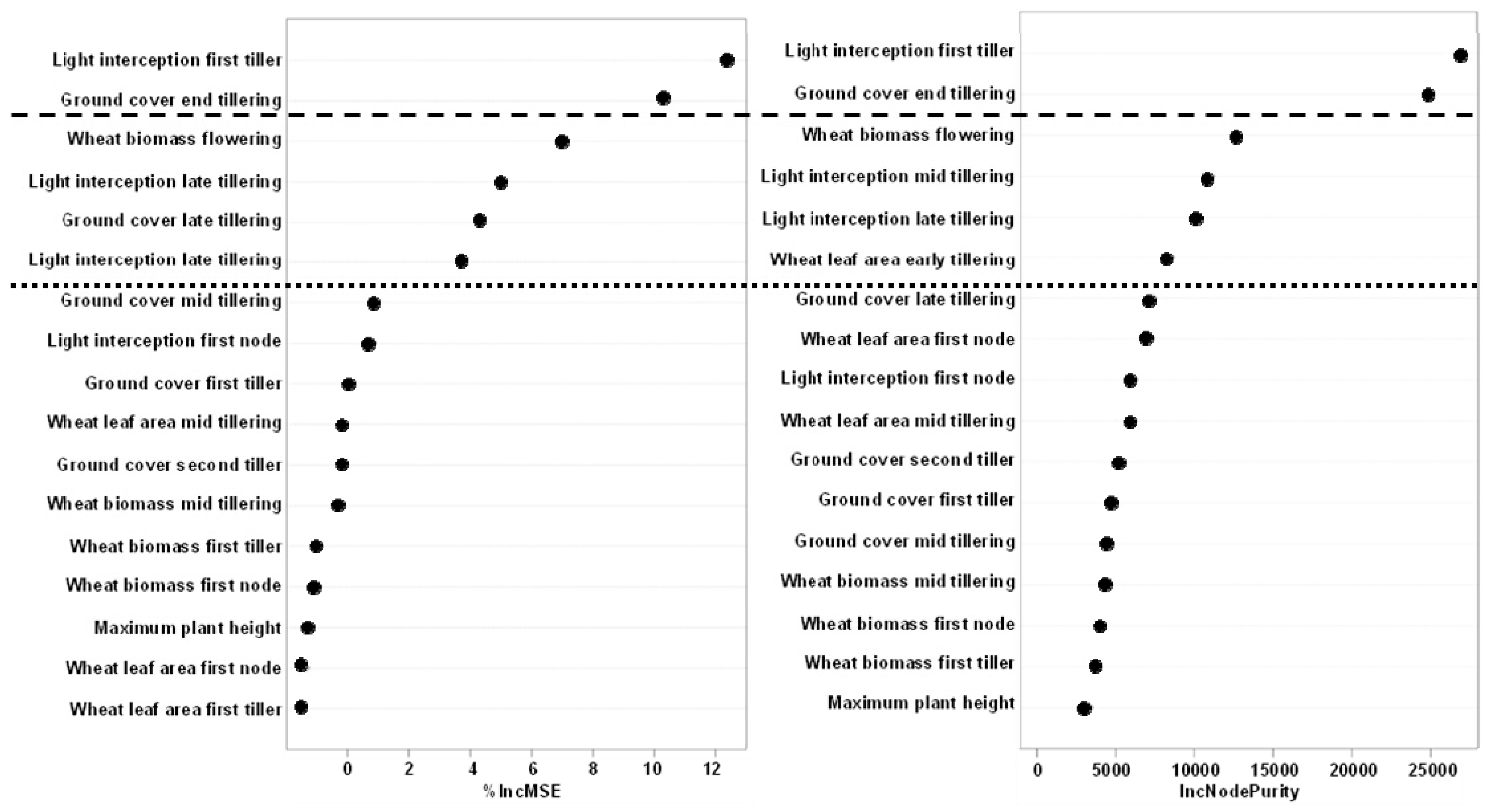
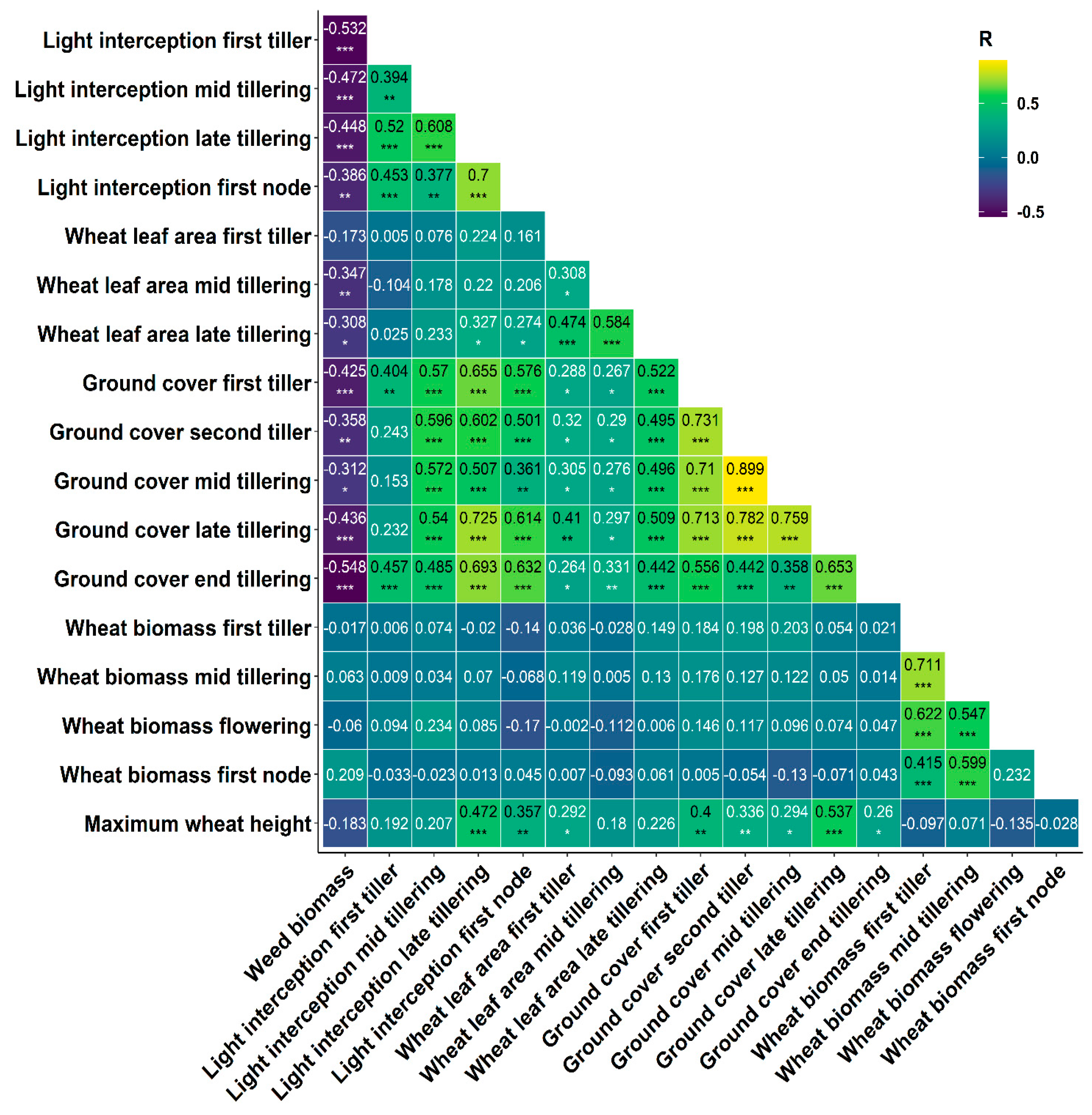
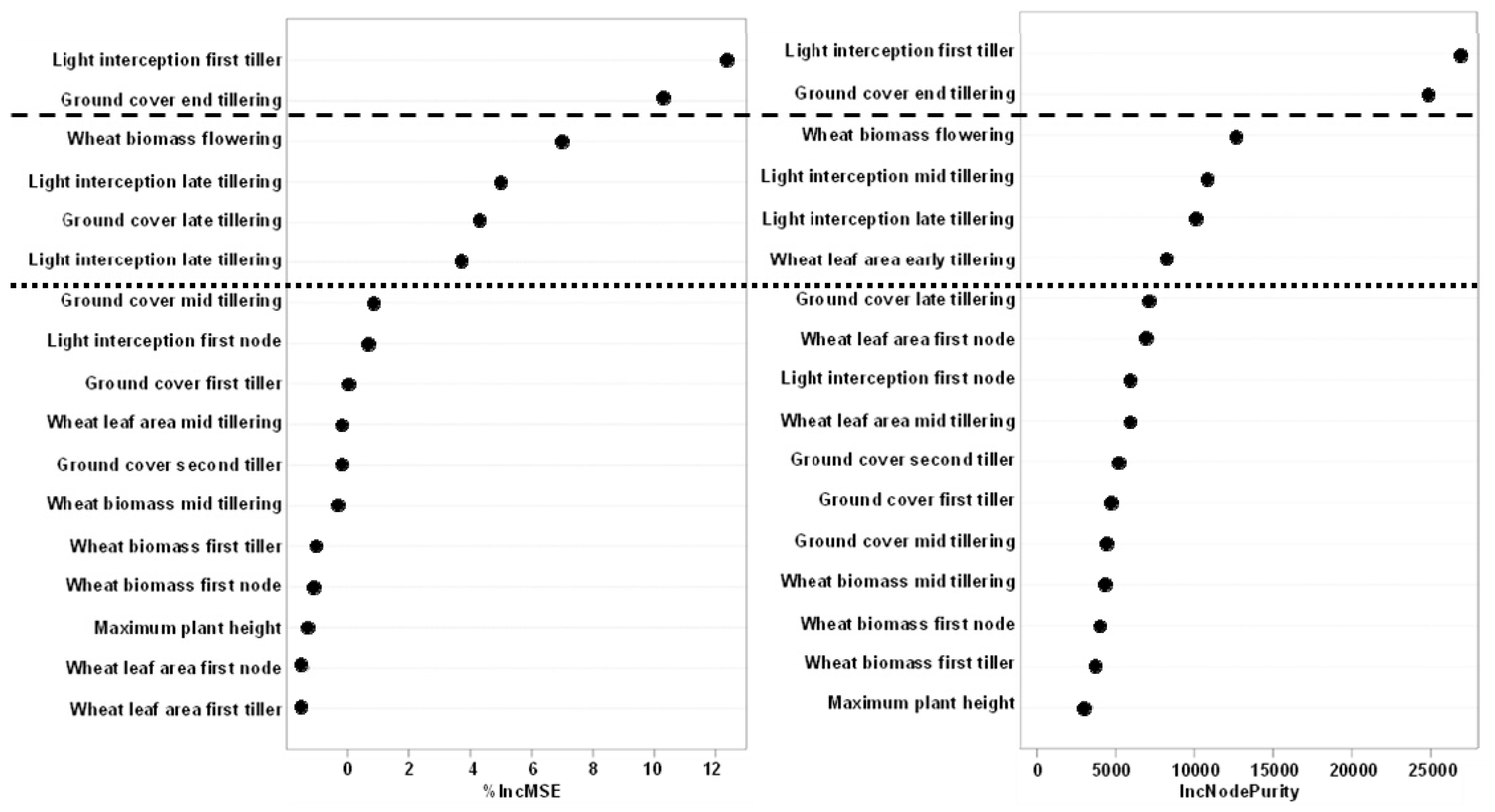
3.3.4. Modelling Weed Suppression Ability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Manschadi, A.M.; Manske, G.; Vlek, P. Root architecture and resource acquisition—Wheat as a model plant. In Plant Roots—The Hidden Half, 4th ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 1–22. [Google Scholar]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaumet, J.; Ghersi, G.; Rastoin, J. Food consumption in 2050. In Scenarios and Challenges for Feeding the World in 2050; Paillard, S., Dorin, T.S., Agrimonde, B., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 59–72. [Google Scholar]
- Fischer, R.A. Definitions and determination of crop yield, yield gaps, and of rates of change. Field Crops Res. 2015, 182, 9–18. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Gallandt, E.R.; Weiner, J. Crop-weed competition. In eLS; Wiley Online Library: Hoboken, NJ, USA, 2015; pp. 1–9. [Google Scholar]
- Evers, J.; Bastiaans, L. Quantifying the effect of crop spatial arrangement on weed suppression using functional-structural plant modelling. J. Plant Res. 2016, 129, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Lowry, C.J.; Smith, R.G. Weed control through crop plant manipulation. In Non Chemical Weed Control; Jabran, K., Chauhan, B.S., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2018; pp. 73–96. [Google Scholar]
- Lemerle, D.; Gill, G.; Murphy, C.E.; Walker, R.; Cousens, S.; Mokhtari, S.; Peltzer, D. Genetic improvement and agronomy for enhanced wheat competitiveness with weeds. Aust. J. Agric. Res. 2001, 52, 527. [Google Scholar] [CrossRef]
- Broster, J.C.; Koetz, E.A.; Wu, H. Herbicide resistance levels in annual ryegrass (Lolium rigidum Gaud.) and wild oat (Avena spp.) in southwestern New South Wales. Plant Prot. Q. 2013, 28, 126–132. [Google Scholar]
- Heap, I. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: https://weedscience.org/Home.aspx (accessed on 17 September 2021).
- Van Der Meulen, A.; Chauhan, B.S. A review of weed management in wheat using crop competition. Crop Prot. 2017, 95, 38–44. [Google Scholar] [CrossRef]
- Jordan, N. Prospects for weed control through crop interference. Ecol. Appl. 1993, 3, 84–91. [Google Scholar] [CrossRef]
- Lemerle, D.; Verbeek, B.; Orchard, B. Ranking the ability of wheat varieties to compete with Lolium rigidum. Weed Res. 2001, 41, 197–209. [Google Scholar] [CrossRef]
- Vandeleur, R.K.; Gill, G.S. The impact of plant breeding on the grain yield and competitive ability of wheat in Australia. Aust. J. Agric. Res. 2004, 55, 855–861. [Google Scholar] [CrossRef]
- Lemerle, D.; Smith, A.; Verbeek, B.; Koetz, E.; Lockley, P.; Martin, P. Incremental crop tolerance to weeds: A measure for selecting competitive ability in Australian wheats. Euphytica 2006, 149, 85–95. [Google Scholar] [CrossRef]
- Renton, M.; Chauhan, B.S. Modelling crop-weed competition: Why, what, how and what lies ahead? Crop Prot. 2017, 95, 101–108. [Google Scholar] [CrossRef]
- Kaur, S.; Kaur, R.; Chauhan, B.S. Understanding crop-weed-fertiliser-water interactions and their implications for weed management in agricultural systems. Crop Prot. 2018, 103, 65–72. [Google Scholar] [CrossRef]
- McPhee, C.; Aarssen, L. The separation of above- and below-ground competition in plants: A review and critique of methodology. Plant Ecol. 2001, 152, 119–136. [Google Scholar] [CrossRef]
- Barnes, P.; Beyschlag, W.; Ryel, R.; Flint, S.; Caldwell, M. Plant competition for light analysed with a multispecies canopy model. Oecologia 1990, 82, 560–566. [Google Scholar] [CrossRef]
- Reid, T.A. The Genetics of Competitive Ability in Spring Wheat. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2010. [Google Scholar]
- Lemerle, D.; Verbeek, B.; Cousens, R.D.; Coombes, N.E. The potential for selecting wheat varieties strongly competitive against weeds. Weed Res. 1996, 36, 505–513. [Google Scholar] [CrossRef]
- Richards, M.C. Crop competitiveness as an aid to weed control. In Proceedings of the Crop Protection Conf Weeds, Brighton, UK, 20–23 November 1989; pp. 573–578. [Google Scholar]
- Rebetzke, G.J.; Richards, R.A. Genetic improvement of early vigour in wheat. Aust. J. Agric. Res. 1999, 50, 291–301. [Google Scholar] [CrossRef]
- Maydup, M.L.; Graciano, C.; Guiamet, J.J.; Tambussi, E.A. Analysis of early vigour in twenty modern cultivars of bread wheat (Triticum aestivum L.). Crop Pasture Sci. 2012, 63, 987–996. [Google Scholar] [CrossRef]
- Black, C.C.; Chen, T.M.; Brown, R.H. Biochemical basis for plant competition. Weed Sci. 1969, 17, 338–344. [Google Scholar] [CrossRef]
- Gaudet, C.L.; Keddy, P.A. A comparative approach to predicting competitive ability from plant traits. Nature 1988, 334, 242–243. [Google Scholar] [CrossRef]
- Coleman, R.K.; Gill, G.S.; Rebetzke, G.J. Identification of quantitative trait loci for traits conferring weed competitiveness in wheat (Triticum aestivum L.). Aust. J. Agric. Res. 2001, 52, 1235–1246. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Botwright, T.L.; Moore, C.S.; Richards, R.A.; Condon, A.G. Genotypic variation in specific leaf area for genetic improvement of early vigour in wheat. Field Crops Res. 2004, 88, 179–189. [Google Scholar] [CrossRef]
- Mason, H.; Navabi, A.; Frick, B.; O’Donovan, J.; Spaner, D. The weed-competitive ability of Canada western red spring wheat cultivars grown under organic management. Crop Sci. 2007, 47, 1167–1176. [Google Scholar] [CrossRef]
- Zerner, M.C.; Rebetzke, G.J.; Gill, G.S. Genotypic stability of weed competitive ability for bread wheat genotypes in multiple environments. Crop Pasture Sci. 2016, 67, 695–702. [Google Scholar] [CrossRef]
- Rebetzke, G.; Condon, A.; Richards, R. Inheritance of coleoptile tiller appearance and size in wheat. Aust. J. Agric. Res. 2008, 59, 863–873. [Google Scholar] [CrossRef]
- Ogg, A.G., Jr.; Seefeldt, S.S. Characterising traits that enhance the competitiveness of winter wheat (Triticum aestivum) against jointed goatgrass (Aegilops cylindrica). Weed Sci. 1999, 47, 74–80. [Google Scholar] [CrossRef]
- Murphy, K.M.; Dawson, J.C.; Jones, S.S. Relationship among phenotypic growth traits, yield and weed suppression in spring wheat landraces and modern cultivars. Field Crops Res. 2008, 105, 107–115. [Google Scholar] [CrossRef]
- Huel, D.G.; Hucl, P. Genotypic variation for competitive ability in spring wheat. Plant Breed. 1996, 115, 325–329. [Google Scholar] [CrossRef]
- Richards, M.C.; Whytock, G.P. Varietal competitiveness with weeds. Asp. Appl. Biol. 1993, 34, 345–354. [Google Scholar]
- Thomas, J.; Schaalje, G.; Grant, M. Height, competition and yield potential in winter wheat. Euphytica 1993, 74, 9–17. [Google Scholar] [CrossRef]
- Cossani, C.M.; Sadras, V. Symmetric response to competition in binary mixtures of cultivars associates with genetic gain in wheat yield. Evol. Appl. 2021, 14, 2064–2078. [Google Scholar] [CrossRef]
- Zhang, L.; Richards, R.A.; Condon, A.G.; Liu, D.C.; Rebetzke, G.J. Recurrent selection for wider seedling leaves increases early biomass and leaf area in wheat (Triticum aestivum L.). J. Exp. Bot. 2015, 66, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Rebetzke, G.; Ingvordsen, C.; Newman, P.; Weston, L.A.; French, B.; Gill, G. Delivering weed-competitive wheat breeding lines to growers. In GRDC Grains Research Update; GRDC: Wagga Wagga, Australia, 2018; pp. 35–40. [Google Scholar]
- Mwendwa, J.M.; Brown, W.B.; Weidenhamer, J.D.; Weston, P.A.; Quinn, J.C.; Wu, H.; Weston, L.A. Evaluation of commercial wheat cultivars for canopy architecture, early vigour, weed suppression and yield. Agronomy 2020, 10, 983. [Google Scholar] [CrossRef]
- Beres, B.L.; Harker, K.N.; Clayton, G.W.; Bremer, E.; Blackshaw, R.E.; Graf, R.J. Weed-competitive ability of spring and winter cereals in the northern great plains. Weed Technol. 2010, 24, 108–116. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Li, Y.; Chen, D.; Walker, C.; Angus, J. Estimating the nitrogen status of crops using a digital camera. Field Crop Res. 2010, 3, 221–227. [Google Scholar] [CrossRef]
- Worthington, M.; Reberg-Horton, S.; Jordan, D.; Murphy, J. A comparison of methods for evaluating the suppressive ability of winter wheat cultivars against Italian ryegrass (Lolium perenne). Weed Sci. 2013, 61, 491–499. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Package “emmeans”: Estimated marginal means, aka least-squares means. In R Package Version 1.3.0; Available online: https://cran.r-project.org/bin/windows/base/ (accessed on 17 September 2021).
- Wang, L.A.; Zhou, X.; Zhu, X.; Dong, Z.; Guo, W. Estimation of biomass in wheat using random forest regression algorithm and remote sensing data. Crop J. 2016, 4, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Weiner, M. Classification and regression by Random Forest. R News 2002, 2, 18–22. [Google Scholar]
- Liland, K.; Mevil, B.; Wehrens, R. Multivariate regression methods. In Partial Least Squares Regression (PLSR), Principal Component Regression (PCR) and Canonical Powered Partial Least Squares (CPPLS); Available online: https://cran.r-project.org/web/packages/pls/pls.pdf (accessed on 17 September 2021).
- Gée, C.; Denimal, E.; Merienne, J.; Larmure, A. Evaluation of weed impact on wheat biomass by combining visible imagery with a plant growth model: Towards new non-destructive indicators for weed competition. Precis. Agric. 2021, 22, 550–568. [Google Scholar] [CrossRef]
- Korres, N.E.; Froud-Williams, R.J. Effects of winter wheat cultivars and seed rate on the biological characteristics of naturally occurring weed flora. Weed Res. 2002, 42, 417–428. [Google Scholar] [CrossRef]
- Seavers, G.P.; Wright, K.J. Crop canopy development and structure influence weed suppression. Weed Res. 1999, 39, 319–328. [Google Scholar] [CrossRef]
- Reid, T.A.; Navabi, A.; Cahill, J.C.; Salmon, D.; Spaner, D. A genetic analysis of weed competitive ability in spring wheat. Can. J. Plant Sci. 2009, 89, 591–599. [Google Scholar] [CrossRef]
- Pavlychenko, T.K.; Harrington, J.B. Competitive efficiency of weeds and cereal crops. Can. J. Res. 1934, 10, 77–94. [Google Scholar] [CrossRef]
- Pavlychenko, T.K.; Harrington, J.B. Root development of weeds and crops in competition under dry farming. Sci. Agric. 1935, 16, 151–160. [Google Scholar]
- Weiner, J.; Du, Y.L.; Zhang, C.; Qin, X.L.; Li, F.M. Evolutionary agroecology: Individual fitness and population yield in wheat (Triticum aestivum). Ecology 2017, 98, 2261–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerner, M.; Gill, G.; Vandeleur, R. Effect of height on the competitive ability of wheat with oats. Agron. J. 2008, 100, 1729–1734. [Google Scholar] [CrossRef]
- Zhao, Z.; Rebetzke, G.J.; Zheng, B.; Chapman, S.C.; Wang, E. Modelling impact of early vigour on wheat yield in dryland regions. J. Exp. Bot. 2019, 70, 2535–2548. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.; Storrie, A. Summer Fallow Weed Management in The Southern and Western Grains Regions of Australia—A Reference for Grain Growers and Adviser. 2014. Available online: https://grdc.com.au/__data/assets/pdf_file/0028/98632/summer-fallow-weed-management-manual.pdf.pdf?utm_source=website&utm_medium=download_link&utm_campaign=pdf_download&utm_term=South;%20West&utm_content=Summer%20fallow%20weed%20management (accessed on 17 September 2021).
- Schefe, C. Research for the Riverine plains 2016. 2016. Available online: https://riverineplains.org.au/wp-content/uploads/2016/10/Research-for-the-Riverine-Plains-2016.pdf (accessed on 17 September 2021).
- McLeod, R. Annual Costs of Weeds in Australia; eSYS Development Pty Limited; Centre for Invasive Species Solutions: Canberra, Australia, 2018. [Google Scholar]
- Kiær, L.P.; Weisbach, A.N.; Weiner, J. Root and shoot competition: A meta-analysis. J. Ecol. 2013, 101, 1298–1312. [Google Scholar] [CrossRef]
- Wilson, J. Shoot competition and root competition. J. Appl. Ecol. 1988, 25, 279–296. [Google Scholar] [CrossRef]
- Sardana, V.; Mahajan, G.; Jabran, K.; Chauhan, B.S. Role of competition in managing weeds: An introduction to the special issue. Crop Prot. 2017, 95, 1–7. [Google Scholar] [CrossRef]
- Evans, G.; Biernaskie, J.M.; Cockram, J.; Marr, E.C.; Oliver, A.G.; Ober, E.; Jones, H. Effects of breeding history and crop management on the root architecture of wheat. Plant Soil 2020, 452, 587–600. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wagga Wagga | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | Annual | In-Crop Season | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Long- term | Max temp (°C) | 31.2 | 30.5 | 27.2 | 22.2 | 17 | 13.7 | 12.5 | 14.4 | 17.4 | 21.1 | 25.2 | 29.2 | 21.8 | |
| Min temp (°C) | 16.4 | 16.9 | 14.1 | 10.1 | 6.8 | 4.3 | 3.3 | 4.3 | 6 | 8.5 | 11.1 | 14.2 | 9.7 | ||
| rain (mm) | 41.6 | 39.4 | 40.6 | 45.5 | 55.1 | 48.5 | 55.6 | 54 | 54.2 | 60.6 | 42.8 | 40.7 | 577.5 | 363.6 | |
| 2018 | Max temp (°C) | 34.3 | 32.4 | 29.9 | 27.7 | 18.4 | 14.8 | 13.8 | 14.7 | 19.1 | 26 | 26.3 | 32.2 | 24.1 | |
| Min temp (°C) | 8.3 | 16.2 | 13.6 | 11.2 | 5.8 | 4.1 | 1.7 | 2.9 | 3.8 | 9.5 | 13 | 17.5 | 9.8 | ||
| rain (mm) | 69.8 | 53.6 | 10.2 | 5 | 30 | 36.8 | 15.6 | 21 | 26.4 | 26.4 | 79 | 65.8 | 439.6 | 253.1 | |
| 2019 | Max temp (°C) | 37.9 | 31.9 | 28.5 | 24.7 | 17.4 | 14.4 | 13.8 | 14.1 | 19.6 | 26 | 27.1 | 33.6 | 24.1 | |
| Min temp (°C) | 22.3 | 16.7 | 15.3 | 11.8 | 6.5 | 2.8 | 3.4 | 1.1 | 3.7 | 8.2 | 11.4 | 16.1 | 9.9 | ||
| rain (mm) | 28.2 | 11.4 | 41 | 24 | 53.9 | 43 | 23 | 18.8 | 20 | 8 | 56.6 | 9.2 | 337.1 | 200.95 | |
| 2020 | Max temp (°C) | 34.2 | 31.2 | 26.7 | 20.6 | 16.7 | 14 | 13.5 | 13.8 | 19.2 | 22.5 | 29.2 | 28.9 | 22.5 | |
| Min temp (°C) | 17.7 | 18.1 | 14.2 | 9.5 | 4.8 | 3.9 | 2.9 | 4 | 6.7 | 10.4 | 13.3 | 13.1 | 9.9 | ||
| rain (mm) | 20 | 21.8 | 81 | 86.7 | 27 | 60.4 | 21 | 70.2 | 57.4 | 85 | 60.2 | 78.2 | 668.9 | 406.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hendriks, P.-W.; Gurusinghe, S.; Ryan, P.R.; Rebetzke, G.J.; Weston, L.A. Competitiveness of Early Vigour Wheat (Triticum aestivum L.) Genotypes Is Established at Early Growth Stages. Agronomy 2022, 12, 377. https://doi.org/10.3390/agronomy12020377
Hendriks P-W, Gurusinghe S, Ryan PR, Rebetzke GJ, Weston LA. Competitiveness of Early Vigour Wheat (Triticum aestivum L.) Genotypes Is Established at Early Growth Stages. Agronomy. 2022; 12(2):377. https://doi.org/10.3390/agronomy12020377
Chicago/Turabian StyleHendriks, Pieter-Willem, Saliya Gurusinghe, Peter R. Ryan, Gregory J. Rebetzke, and Leslie A. Weston. 2022. "Competitiveness of Early Vigour Wheat (Triticum aestivum L.) Genotypes Is Established at Early Growth Stages" Agronomy 12, no. 2: 377. https://doi.org/10.3390/agronomy12020377
APA StyleHendriks, P.-W., Gurusinghe, S., Ryan, P. R., Rebetzke, G. J., & Weston, L. A. (2022). Competitiveness of Early Vigour Wheat (Triticum aestivum L.) Genotypes Is Established at Early Growth Stages. Agronomy, 12(2), 377. https://doi.org/10.3390/agronomy12020377