4.1. Standing Vegetative Biomass and Dry-Matter Production
The aboveground dry weight of tenera palms measured in this study were comparable to previous studies on either dura or tenera palms grown on different soils and under different management practices in Malaysia [
16,
18,
34] and in Indonesia [
35,
36] (
Figure S2). Our results suggest that accumulation of aboveground vegetative biomass in oil palm tends to remain relatively constant across different types of planting materials and environments. However, clear differences in aboveground vegetative biomass accumulation were observed on palms grown under suboptimal conditions such as peat in Malaysia [
37] and in West Africa [
15,
38]. The poor growing conditions of peat [
39,
40] and marked dry seasons prevailing in West Africa [
41] would have detrimentally affected the growth of the sampled palms there; hence, they accumulated less aboveground vegetative biomass.
The rapid increase in dry-matter production during the first 88 months of planting is typical of oil palm and is consistent with previous studies [
2,
16]. Production of aboveground vegetative dry matter in mature palms was in close agreement with studies conducted in Malaysia [
2,
16] but was much higher than in oil palms studied in West Africa [
15,
38] (
Table S3). Seasonal drought (6 consecutive months with monthly mean rainfall of less than 100 mm per month
−1) [
11] in West Africa would have affected the rate of leaf production and expansion [
36,
42], and in turn resulted in lower leaf dry-matter production. Ng et al. [
2], on the other hand, might have underestimated leaf dry-matter production by only considering 18 pruned leaves with average leaf dry weight of 3.95 kg while overestimating trunk dry-matter production by including leaf bases as part of the trunk.
If root dry-matter production is considered, the total vegetative dry-matter production estimated in this study agreed with Dufrene’s estimation [
38] of 171.3 kg palm
−1 year
−1, but this was about 25% higher than Corley et al. [
16] and 75% higher than Rees and Tinker [
15]. Dufrene [
38] and this study estimated annual root dry-matter production, but the other studies only considered net increase in root dry weight, and this perhaps explains the observed differences. If Equation (4), which was developed in Malaysia, was used to estimate the root dry-matter production in Corley et al. [
16], then total vegetative dry-matter production would be increased to 144.7 kg palm
−1 year
−1, quite close to the present estimate of 168.2 kg palm
−1 year
−1 (
Table S3). It is thus obvious that production of vegetative dry matter could be quite similar across different environments, but the partitioning of above- and belowground vegetative dry matter might vary depending on the prevailing growing conditions. Dufrene [
38] showed that oil palm tends to partition more dry matter to roots in places with pronounced dry seasons. In this study, the root dry-matter production was estimated using Equation (4) developed in Malaysia [
31]. The estimated root dry-matter production only represents about 10.4% of total aboveground vegetative dry-matter production. This is a considerably small fraction of dry matter partitioned to the roots as argued by Corley and Tinker [
11] (pp. 341–342). Thus, it seems that further studies are required to ascertain root dry-matter production of tenera palms at different ages of growth, since this has important implication on total dry-matter production, partitioning, and vegetative growth demand for nutrients.
Bunch dry matter produced in this study was comparable to the amounts produced elsewhere in Malaysia but was 1.6 to 2 times more than those in West Africa (
Table S3). This is not unexpected, as dry-matter requirements for vegetative growth appear to be met first before any surplus dry matter is diverted to bunch production. This surplus is smaller in less-conducive environments like West Africa, where a marked dry season prevails. Consequently, total dry-matter production across different environments might not vary as substantially as highlighted here. Total dry-matter production found in this study was only 15% more than Dufrene [
38], despite having produced 61% more bunch dry matter.
4.2. Nutrient Concentrations and Nutrient Demand
The variations in nutrient concentrations among vegetative tissues found in this study were consistent with previous studies [
2,
4,
5,
17,
18,
21,
43]. These results confirm that the uptake of nutrients cannot be reliably deduced from leaf nutrient analysis alone [
43], as high concentrations of nutrients are also found in other vegetative tissues such as rachis, petiole, leaf bases, and trunk. Classical fertilizer recommendation methodology based solely on leaf nutrient analysis data must consider the variations in nutrient concentrations among vegetative tissues. Considerable palm-to-palm variations in nutrient concentrations might be attributed to genetic differences, since tenera palms planted at the study site came from different genetic origins. It has been shown that nutrient uptake could vary by up to 30–40% among oil-palm planting materials [
43]. It is also possible that the variations observed were caused by genetic–environmental interactions. We therefore recommend establishing plantations with blocks of homogenous oil-palm planting materials in order to obtain a more reliable estimate of fertilizer requirements. In the case of existing plantations with mixed oil-palm planting materials, an appropriate sampling size should be established for every management block so as to minimize variation.
Much higher nutrient concentrations found in the cabbage were expected and have been documented [
2,
21]. This plant part is the growing point of the palm, so it is not surprising to observe a much higher concentration of nutrients here, as more nutrients are required to grow developing tissues, such as immature leaves and leaf bases. Higher K concentrations found in all vegetative tissues, particularly in the trunk, agreed with previous findings [
2,
18,
43,
44], and confirmed its prominent role in the nutrition of oil palm. The high concentrations of nutrients found in the leaf bases highlight the importance of this plant part as nutrient sink and stock, and it should be given due attention since it constitutes a significant portion of vegetative biomass (
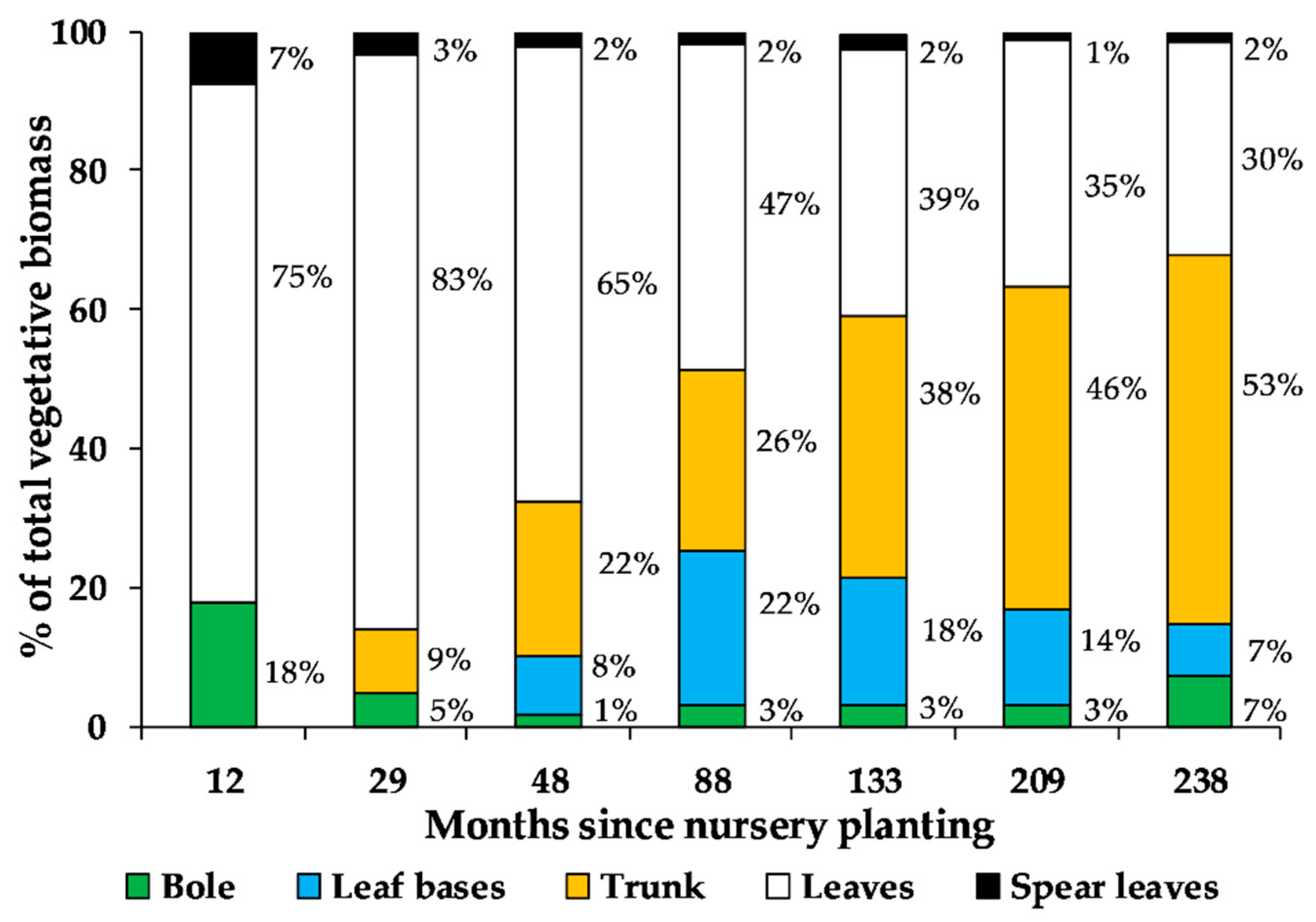
Table 2). The amounts of nutrients immobilized by leaf bases and recycled back to the soil through shedding of leaf bases could be significant [
45]. The low nutrient concentrations found in the roots have been reported [
2,
4,
5,
21].
The higher concentrations of N, P, K, Mg, and Ca found in the whole immature and mature tenera studied here (
Table 6), suggest that the tenera studied here might require more N, P, K, Mg, and Ca to sustain vegetative growth for a same unit of biomass produced. Recent advances in agronomic practices might have improved the uptake of nutrients, and led to the observed higher concentrations of N, P, K, Mg, and Ca. It is also possible that the presently studied tenera had higher nutrient concentrations in its vegetative tissues intrinsically as a result of advances in oil-palm breeding [
43].
The decline in concentrations of N and P with age, particularly in the trunk, has been reported [
2]. Henson and Chang [
46] analyzed published nutrient concentration data and found that only concentrations of N and P in the trunk declined with age, while N, P, and K concentrations in other palm tissues did not. The present study confirmed their findings but also found that concentrations of N and P in the leaves, concentration of Mg in the leaves and trunk, and concentrations of Ca and B in the trunk declined with age (
Table 7). The ability of oil palm to store K in vegetative tissues might explain the lack of correlation between K concentration and palm age [
18,
44]. This is supported by higher K concentrations found in the vegetative tissues such as rachis, petiole, leaf bases, trunk, and bole (
Table 5). If whole-palm nutrient concentration is considered, only K concentration did not decline with age (
Table 7). The physiological mechanism underlying the decline in concentration of nutrients in plant tissues with plant or organ age is complex and has not been fully understood [
47]. One plausible explanation is that as the plant grows and accumulates more biomass, the increase in plant or organ biomass is greater than the corresponding increase in nutrient content. This eventually decreases the concentration of nutrients due to the dilution effect of biomass [
47,
48]. Trunk K behaved quite differently in relation to palm age. It tended to increase with palm age initially before declining to a more stable level (
Figure S3). Ng et al. [
2] observed a similar trend of K concentration in the trunk (
Figure S3). They argued that oil palm tends to build up a K reserve in the trunk during the early years of planting, but this reserve will deplete gradually soon after fruit-bunch production peaks. The present observation seems to agree with Ng et al. [
2] as K concentration in the trunk tended to decrease with increasing fruit-bunch yield in both studies.
The variations in nutrient concentrations among bunch components found in this study were consistent with previous studies [
6,
19,
20,
21,
33]. The most striking variation is the nutrient concentrations in the mesocarp. The tenera bunches studied here had much higher N, P, K, Mg, and Ca concentrations in their mesocarps than previously studied dura and tenera bunches (
Table 8). Consequently, the tenera bunches studied here contain higher N, P, K, Mg, and Ca contents than previously studied dura and tenera bunches (
Table 9). It is thus obvious that for the same production level of fruit bunches, the tenera studied here would require more nutrients to support fruit-bunch production.
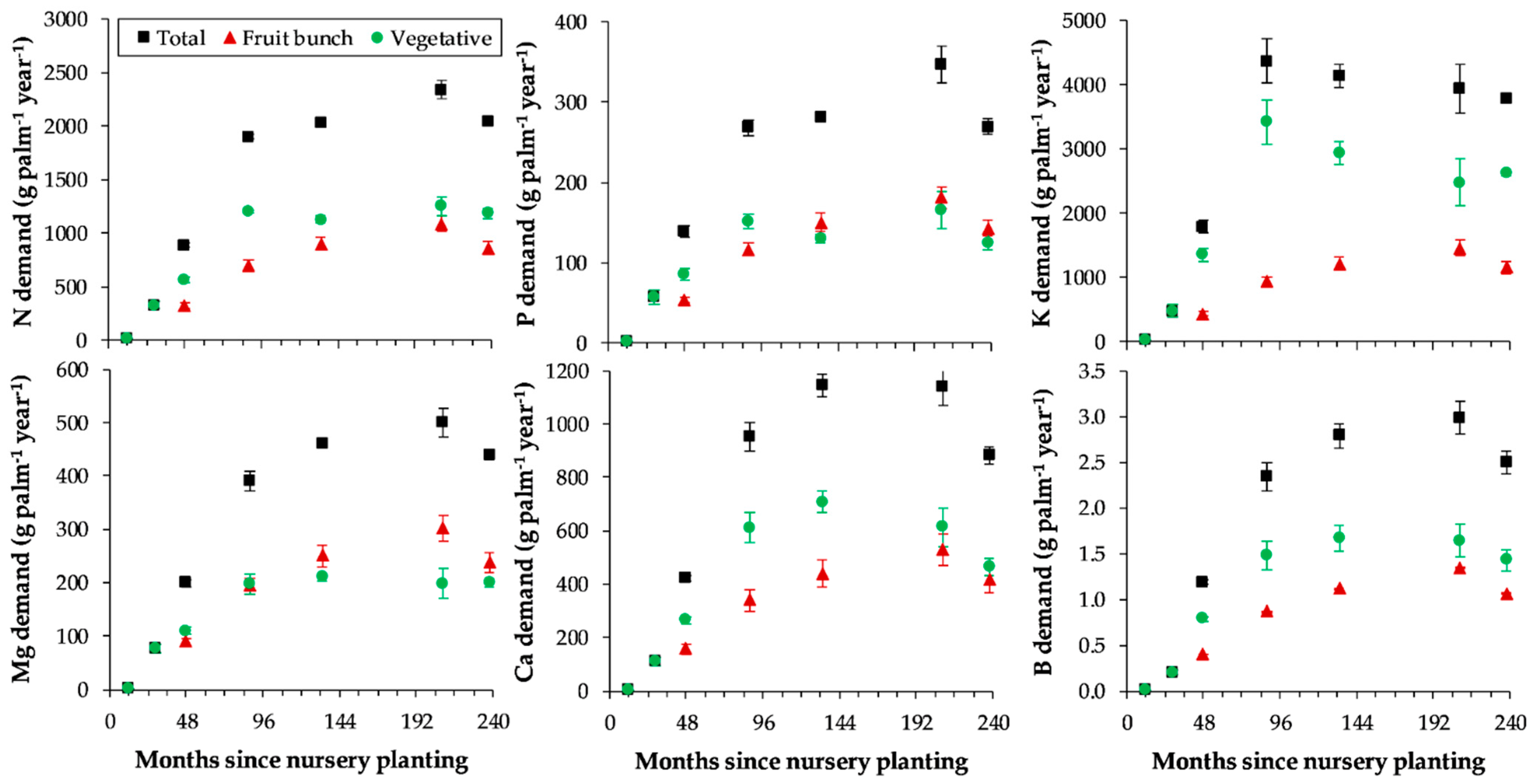
The rapid increase in nutrient demand in the first 88 months since nursery planting was mainly driven by growth rate (
Table 4), consistent with Ng et al. [
2] and Goh et al. [
21], and also suggests that derivation of nutrient requirements from palms of one age or from mature palms alone appears inadequate. Higher demand of N, P, Mg, Ca, and B at month 209 was attributed to high demand of these nutrients for fruit-bunch production (
Figure 3;
Table 4). The decline in K demand for vegetative growth after month 88 was consistent with the decline in trunk K concentration (
Figure 3 and
Figure S3). It appears that the studied tenera did not increase its K uptake when fruit-bunch demand for K increased due to increasing fruit-bunch production but instead used K reserved in the vegetative tissues, particularly trunk K, to meet fruit-bunch production. Thus, it is important to build up K reserves during the early years of planting to meet increased K demand in later years. Teoh and Chew [
18] estimated that trunk K reserve of fully fertilized tenera palms could support vegetative growth and fruit-bunch production for 2 to 6 years.
The substantial amounts of nutrients required annually by the tenera studied here re-emphasize the large fertilizer needs of oil palm and the importance of judicious manuring that meets agronomic, economic, and environmental objectives. The amounts of nutrients depicted in
Figure 3 were regarded as gross nutrient demand [
2,
11] since nutrients recycled from pruned leaves were not considered. If annual fruit-bunch production of 25 t ha
−1 is considered, the annual gross nutrient demand of mature tenera studied here would be 1.99 kg N, 0.28 kg P, 3.94 kg K, 0.42 kg Mg, and 0.99 kg Ca palm
−1 which is about 53, 57, 132, and 64% more N, P, K, and Ca than found in the dura studied by Ng et al. [
2], respectively, but Mg demand was only 2% higher than in the dura. No difference was found when annual B demand was compared with another mature tenera [
21], as both required 2.5 g palm
−1.
However, if nutrients contained in the pruned leaves were all recycled, the annual net demand of nutrients would be 1.04 kg N, 0.16 kg P, 1.85 kg K, 0.26 kg Mg, 0.47 kg Ca, and 1.3 g B palm
−1. These amounts were higher than net nutrient demand of fully fertilized mature dura [
2] and tenera [
18] studied previously. Teoh and Chew [
18] found that fully fertilized mature tenera grown on different soils in Malaysia required 1.34 to 1.59 kg K palm
−1 year
−1 if K recycled from pruned leaves was considered. The higher nutrient demand is not unexpected because of higher nutrient concentrations found in the vegetative tissues and fruit bunches of tenera studied here, and given that dry-matter production of either dura or tenera planting materials could be quite similar, especially when they are grown under favorable environments (
Table S3). Therefore, for the same level of dry-matter production, the tenera studied here would need more nutrients.
Fruit bunches accounted for the largest portion of required N, P, Mg, Ca, and B, except in the case of K (
Table 9), because a large portion of K is stored in vegetative tissues such as leaves, trunk, and leaf bases (
Table 6). Goh et al. [
44] found that fully fertilized mature tenera grown on different soils in Malaysia partitioned 70–73% of their K demand to vegetative tissues, whereas unfertilized tenera partitioned 55–70%. Thus, from the fertilizer management context, the production level of fruit bunches has great influence on the determination of fertilizer requirements of oil palm.







{kind=link}
{kind=link}
{kind=link}