
Phytotoxicity and Plant Defence Induction by Cinnamomum cassia Essential Oil Application on Malus domestica Tree: A Molecular Approach

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Emulsion Formulation and Application
2.3. Redox Status: Determination of Reduced (GSH) and Oxidized Glutathione (GSSG)
2.4. Oxidative Damage
2.4.1. Determination of Malondialdehyde (MDA)
2.4.2. Determination of Photosynthetic Pigments
2.5. Induction of Defences (RT-PCR)
2.6. Data Analysis
3. Results
3.1. Redox Status: Determination of Reduced (GSH) and Oxidized Glutathione (GSSG)
3.2. Oxidative Damage
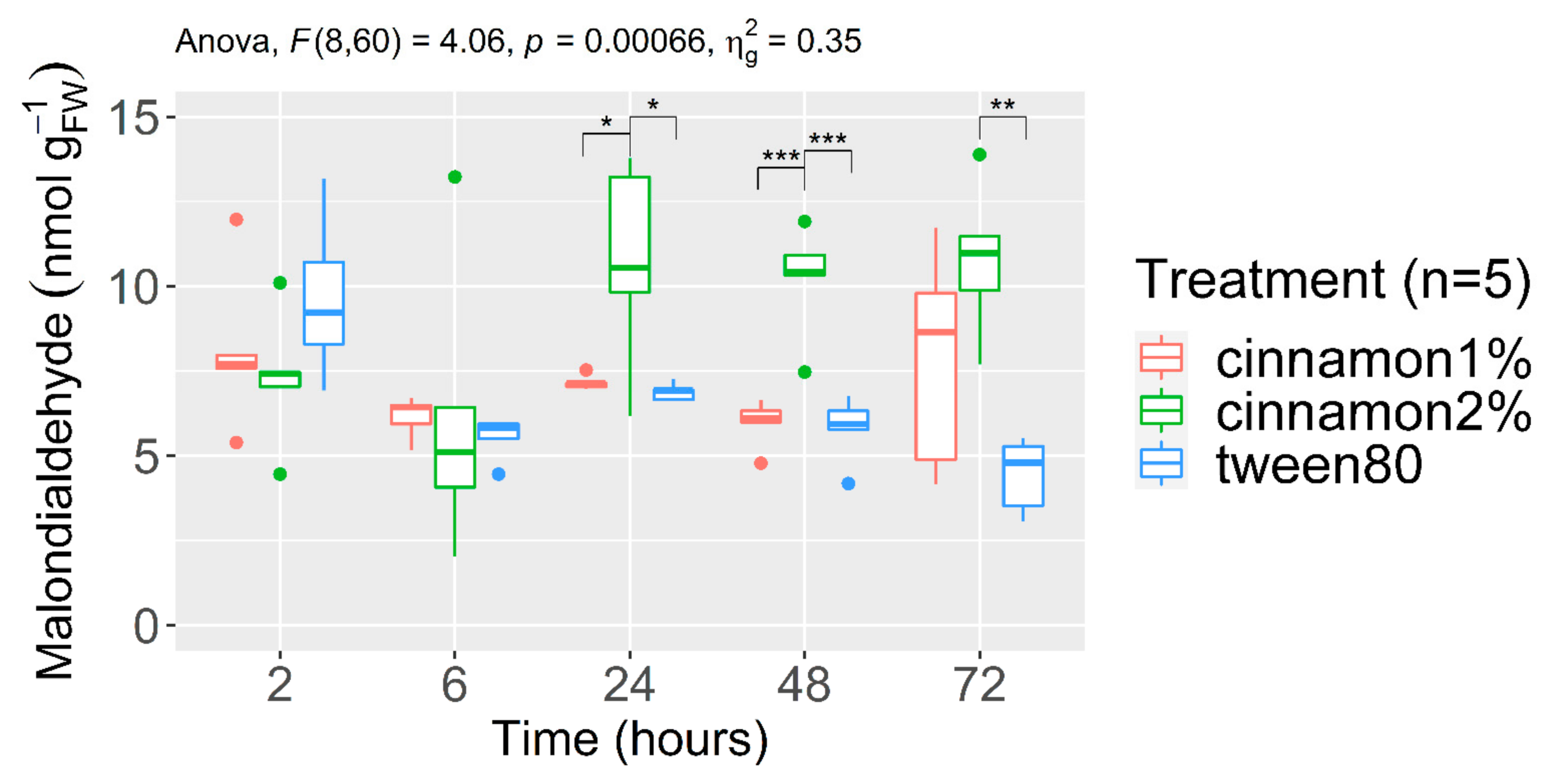
3.2.1. Malondialdehyde Content (MDA)
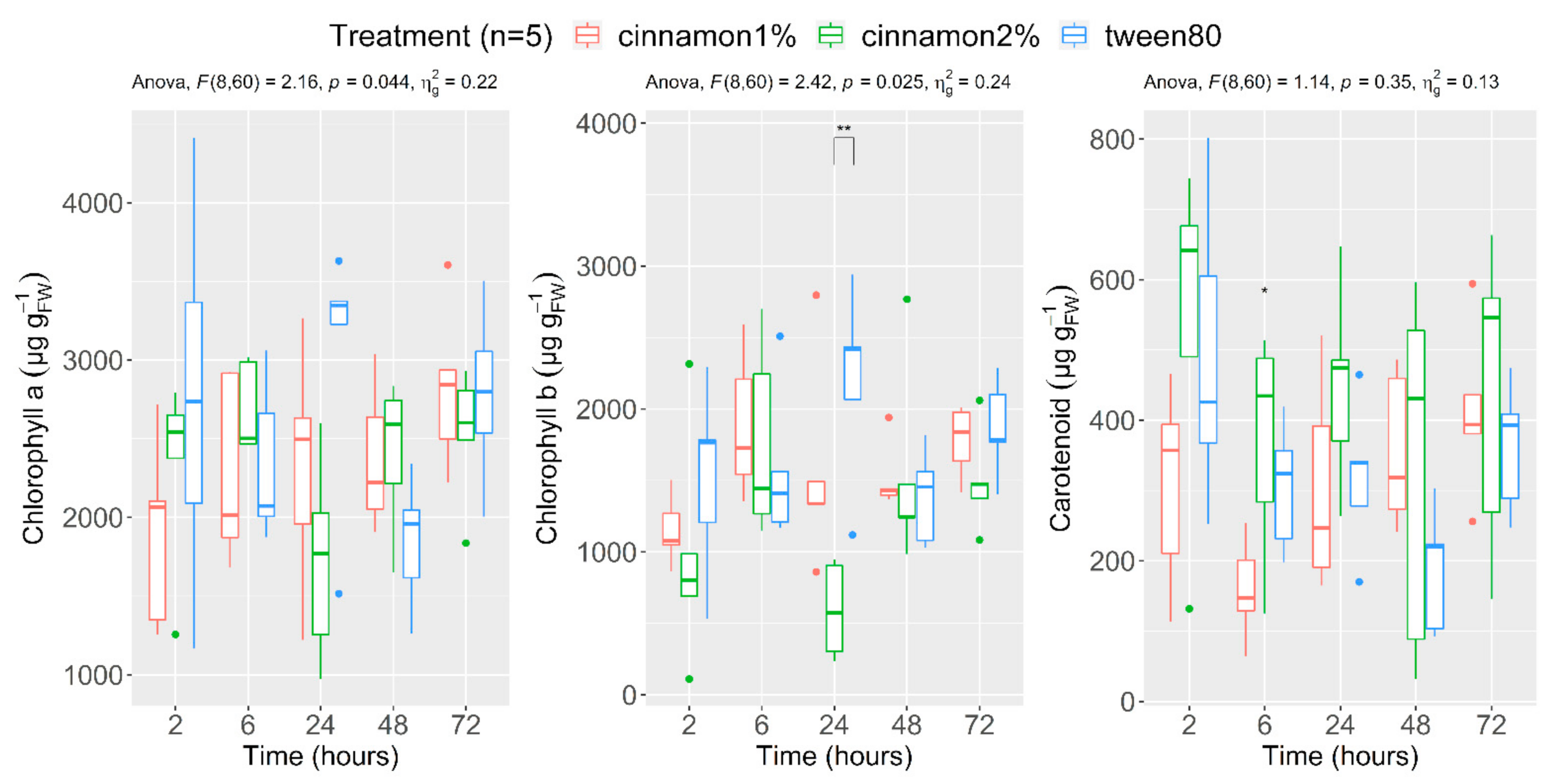
3.2.2. Photosynthetic Pigment Content (Chlorophyll a, Chlorophyll b and Carotenoids)
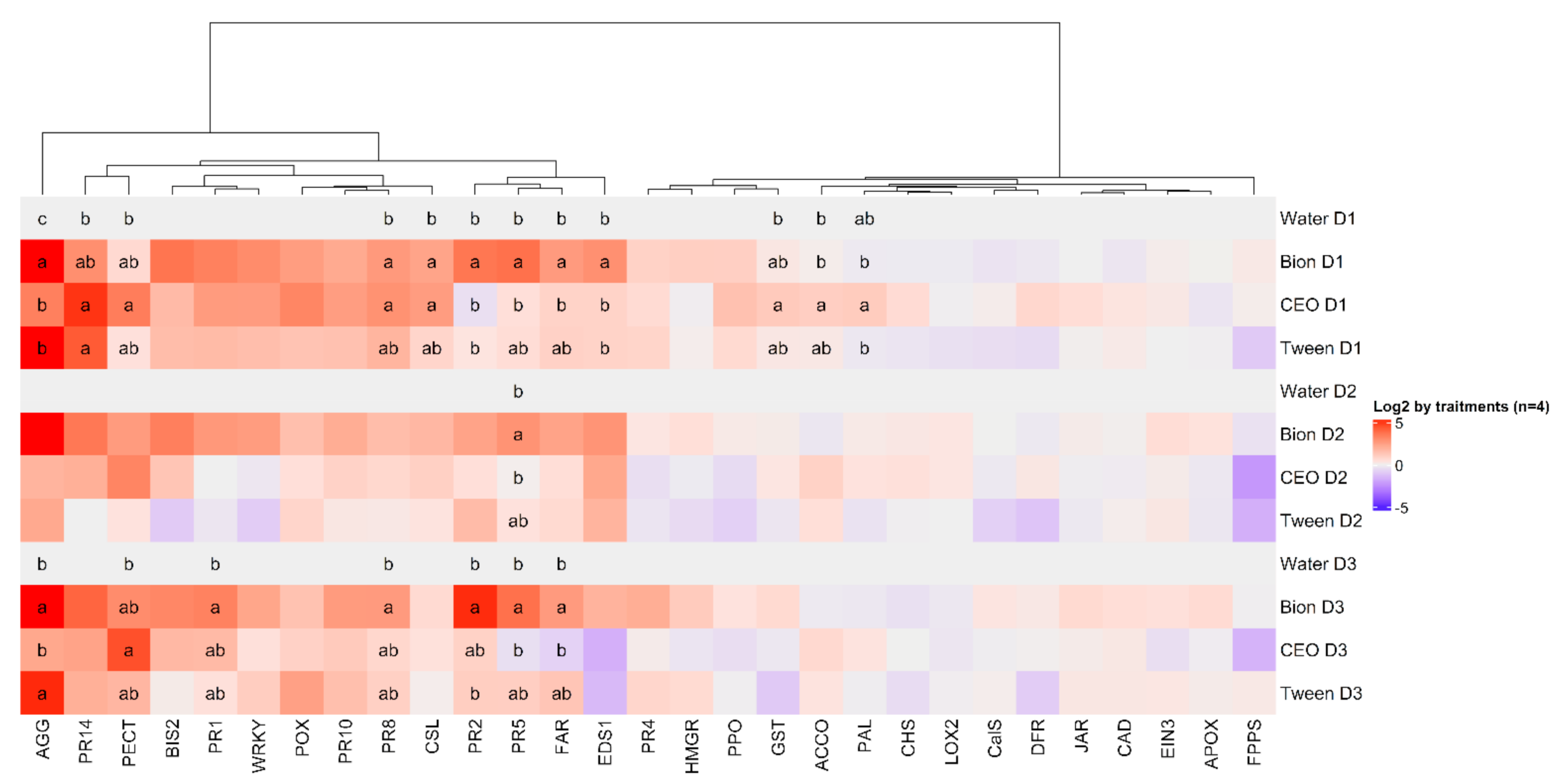
3.3. Induction of Defences
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Bryan, C.A.; Pendleton, S.J.; Crandall, P.G.; Ricke, S.C. Potential of plant essential oils and their components in animal agriculture-in vitro studies on antibacterial mode of action. Front. Vet. Sci. 2015, 2, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Clerck, C.; Maso, S.D.; Parisi, O.; Dresen, F.; Zhiri, A.; Haissam Jijakli, M. Screening of antifungal and antibacterial activity of 90 commercial essential oils against 10 pathogens of agronomical importance. Foods 2020, 9, 1418. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system—A review. Molecules 2018, 23, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, M.G.; Costa-Júnior, L.M.; Blank, A.F.; Lima, A.d.S.; Menezes, T.S.A.; Santos, D.d.A.; Alves, P.B.; Cavalcanti, S.C.d.H.; Bacci, L.; Arrigoni-Blank, M.d.F. Acaricidal activity of essential oils from Lippia alba genotypes and its major components carvone, limonene, and citral against Rhipicephalus microplus. Vet. Parasitol. 2015, 210, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef] [Green Version]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorg. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef]
- Coats, J.R.; Karr, L.L.; Drewes, C.D. Toxicity and Neurotoxic Effects of Monoterpenoids: In Insects and Earthworms. Nat. Occur. Pest Bioregulators 1991, 449, 305. [Google Scholar]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Mossa, A.T.H. Green Pesticides: Essential oils as biopesticides in insect-pest management. J. Environ. Sci. Technol. 2016, 9, 354–378. [Google Scholar] [CrossRef] [Green Version]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Renoz, F.; Demeter, S.; Degand, H.; Nicolis, S.C.; Lebbe, O.; Martin, H.; Deneubourg, J.-L.; Fauconnier, M.L.; Morsomme, P.; Hance, T. The modes of action of Mentha arvensis essential oil on the granary weevil Sitophilus granarius revealed by a label-free quantitative proteomic analysis. J. Pest Sci. 2022, 95, 381–395. [Google Scholar] [CrossRef]
- Werrie, P.-Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.-L. Phytotoxicity of Essential Oils: Opportunities and Constraints for the Development of Biopesticides. A Review. Foods 2020, 9, 1291. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Queval, G.; Mhamdi, A.; Chaouch, S.; Foyer, C.H. Glutathione. Arab. B. 2011, 9, e0142. [Google Scholar] [CrossRef]
- Queval, G.; Jaillard, D.; Zechmann, B.; Noctor, G. Increased intracellular H2O2 availability preferentially drives glutathione accumulation in vacuoles and chloroplasts. Wiley Online Libr. 2010, 34, 21–32. [Google Scholar] [CrossRef]
- Hajdinák, P.; Czobor, Á.; Lőrincz, T.; Szarka, A. The problem of glutathione determination: A comparative study on the measurement of glutathione from plant cells. Period. Polytech. Chem. Eng. 2019, 63, 1–10. [Google Scholar] [CrossRef]
- Davey, M.W.; Dekempeneer, E.; Keulemans, J. Rocket-powered high-performance liquid chromatographic analysis of plant ascorbate and glutathione. Anal. Biochem. 2003, 316, 74–81. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Krasuska, U.; Andrzejczak, O.; Soltys, D. Allelopathic Compounds as Oxidative Stress Agents: Yes or NO. In Signaling and Communication in Plants; Springer: Cham, Switzerland, 2015; Volume 23, pp. 155–176. [Google Scholar]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide- and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol. Plant. 1997, 100, 241–254. [Google Scholar] [CrossRef]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.W.; Stals, E.; Panis, B.; Keulemans, J.; Swennen, R.L. High-throughput determination of malondialdehyde in plant tissues. Anal. Biochem. 2005, 347, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araniti, F.; Landi, M.; Lupini, A.; Sunseri, F.; Guidi, L.; Abenavoli, M.R. Origanum vulgare essential oils inhibit glutamate and aspartate metabolism altering the photorespiratory pathway in Arabidopsis thaliana seedlings. J. Plant Physiol. 2018, 231, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Pouresmaeil, M.; Nojadeh, M.S.; Movafeghi, A.; Maggi, F. Exploring the bio-control efficacy of Artemisia fragrans essential oil on the perennial weed Convolvulus arvensis: Inhibitory effects on the photosynthetic machinery and induction of oxidative stress. Ind. Crops Prod. 2020, 155, 112785. [Google Scholar] [CrossRef]
- Poonpaiboonpipat, T.; Pangnakorn, U.; Suvunnamek, U.; Teerarak, M.; Charoenying, P.; Laosinwattana, C. Phytotoxic effects of essential oil from Cymbopogon citratus and its physiological mechanisms on barnyardgrass (Echinochloa crus-galli). Ind. Crops Prod. 2013, 41, 403–407. [Google Scholar] [CrossRef]
- Gao, Y.; Kan, C.; Chen, M.; Chen, C.; Chen, Y.; Fu, Y.; Wan, C.; Chen, J. Effects of Chitosan-Based Coatings Enriched with Cinnamaldehyde on Mandarin Fruit cv. Ponkan during Room-Temperature Storage. Coatings 2018, 8, 372. [Google Scholar] [CrossRef]
- Chowhan, N.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Phytotoxic effects of β-pinene on early growth and associated biochemical changes in rice. Acta Physiol. Plant. 2011, 33, 2369–2376. [Google Scholar] [CrossRef]
- Bresson, J.; Bieker, S.; Riester, L.; Doll, J.; Zentgraf, U. A guideline for leaf senescence analyses: From quantification to physiological and molecular investigations. J. Exp. Bot. 2018, 69, 769–786. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Alqarawi, A.A.; Abd Allah, E.F. Bioherbicides: Current knowledge on weed control mechanism. Ecotoxicol. Environ. Saf. 2018, 158, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.A.; Merzlyak, M.N. Non-destructive assessment of chlorophyll, carotenoid and anthocyanin in higher plant leaves: Principles and algorithms. Remote Sens. Agric. Environ. 2004, 78–94. Available online: https://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1265&context=natrespapers (accessed on 10 January 2022).
- Weber, H.; Chételat, A.; Reymond, P.; Farmer, E.E. Selective and powerful stress gene expression in Arabidopsis in response to malondialdehyde. Plant J. 2004, 37, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Mhamdi, A.; Chaouch, S.; Noctor, G. Regulation of basal and oxidative stress-triggered jasmonic acid-related gene expression by glutathione. Plant Cell Environ. 2013, 36, 1135–1146. [Google Scholar] [CrossRef]
- Gomez, L.D.; Noctor, G.; Knight, M.R.; Foyer, C.H. Regulation of calcium signalling and gene expression by glutathione. J. Exp. Bot. 2004, 55, 1851–1859. [Google Scholar] [CrossRef] [Green Version]
- Massimo, E. Maffei Monoterpenoid Plant-Plant Interactions Upon Herbivory. Curr. Bioact. Compd. 2012, 8, 65–70. [Google Scholar] [CrossRef]
- Ben-Jabeur, M.; Kthiri, Z.; Harbaoui, K.; Belguesmi, K.; Serret, M.D.; Araus, J.L.; Hamada, W. Seed Coating with Thyme Essential Oil or Paraburkholderia phytofirmans PsJN Strain: Conferring Septoria Leaf Blotch Resistance and Promotion of Yield and Grain Isotopic Composition in Wheat. Agronomy 2019, 9, 586. [Google Scholar] [CrossRef] [Green Version]
- Banani, H.; Olivieri, L.; Santoro, K.; Garibaldi, A.; Gullino, M.; Spadaro, D. Thyme and Savory Essential Oil Efficacy and Induction of Resistance against Botrytis cinerea through Priming of Defense Responses in Apple. Foods 2018, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Sukegawa, S.; Shiojiri, K.; Higami, T.; Suzuki, S.; Arimura, G.-i. Pest management using mint volatiles to elicit resistance in soy: Mechanism and application potential. Plant J. 2018, 96, 910–920. [Google Scholar] [CrossRef] [Green Version]
- Godard, K.A.; White, R.; Bohlmann, J. Monoterpene-induced molecular responses in Arabidopsis thaliana. Phytochemistry 2008, 69, 1838–1849. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, L.; Cai, X.; Li, X.; Bian, L.; Luo, Z.; Li, Z.; Chen, Z.; Xin, Z. (E)-Nerolidol is a volatile signal that induces defenses against insects and pathogens in tea plants. Hortic. Res. 2020, 7, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Jabeur, M.; Ghabri, E.; Myriam, M.; Hamada, W. Thyme essential oil as a defense inducer of tomato against gray mold and Fusarium wilt. Plant Physiol. Biochem. 2015, 94, 35–40. [Google Scholar] [CrossRef]
- Jhonata, L.d.S.; Paulo, E.d.S.; Eduardo, A.; Jose, E.B.P.P.; Suzan, K.V.B.; Marcelo, L.O.F.; Camila, C.L.d.A.; Mario, L.V.R. Essential oil of Cymbopogon flexuosus, Vernonia polyanthes and potassium phosphite in control of bean anthracnose. J. Med. Plants Res. 2015, 9, 243–253. [Google Scholar] [CrossRef]
- Perina, F.J.; de Andrade, C.C.L.; Moreira, S.I.; Nery, E.M.; Ogoshi, C.; Alves, E. Cinnamomun zeylanicum oil and trans-cinnamaldehyde against Alternaria brown spot in tangerine: Direct effects and induced resistance. Phytoparasitica 2019, 47, 575–589. [Google Scholar] [CrossRef]
- Pereira, R.B.; Lucas, G.C.; Perina, F.J.; Ribeiro, P.M., Jr.; Alves, E. Citronella essential oil in the control and activation of coffee plants defense response against rust and brown eye spot. Ciência Agrotecnol. 2012, 36, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Dugé De Bernonville, T.; Marolleau, B.; Staub, J.; Gaucher, M.; Brisset, M.N. Using molecular tools to decipher the complex world of plant resistance inducers: An apple case study. J. Agric. Food Chem. 2014, 62, 11403–11411. [Google Scholar] [CrossRef]
- Koul, O.; Walia, S.; Dhaliwal, G. Essential oils as green pesticides: Potential and Constraints. Biopestic. Int. 2008, 4, 63–84. [Google Scholar]
- Lins, L.; Dal Maso, S.; Foncoux, B.; Kamili, A.; Laurin, Y.; Genva, M.; Jijakli, M.H.; De Clerck, C.; Fauconnier, M.L.; Deleu, M. Insights into the Relationships Between Herbicide Activities, Molecular Structure and Membrane Interaction of Cinnamon and Citronella Essential Oils Components. Int. J. Mol. Sci. 2019, 20, 4007. [Google Scholar] [CrossRef] [Green Version]
- Mezőfi, L.; Sipos, P.; Vétek, G.; Elek, R.; Markó, V. Evaluation of kaolin and cinnamon essential oil to manage two pests and a fungal disease of sour cherry at different tree canopy levels. J. Plant Dis. Prot. 2018, 125, 483–490. [Google Scholar] [CrossRef]
- Washington, D.C.; Mavian, C.M.K. U.S. EPA, Pesticide Product Label, CINNACURE P1005, 01/21/1999. 1999. Available online: https://www3.epa.gov/pesticides/chem_search/ppls/058866-00011-19990121.pdf (accessed on 2 February 2022).
- Warneys, R.; Gaucher, M.; Robert, P.; Aligon, S.; Anton, S.; Aubourg, S.; Barthes, N.; Braud, F.; Cournol, R.; Gadenne, C.; et al. Acibenzolar-s-methyl reprograms apple transcriptome toward resistance to rosy apple aphid. Front. Plant Sci. 2018, 871, 1795. [Google Scholar] [CrossRef]
- Le Mire, G.; Siah, A.; Marolleau, B.; Gaucher, M.; Maumené, C.; Brostaux, Y.; Massart, S.; Brisset, M.N.; Haissam Jijakli, M. Evaluation of l-carrageenan, CpG-ODN, glycine betaine, spirulina platensis, and ergosterol as elicitors for control of zymoseptoria tritici in wheat. Phytopathology 2019, 109, 409–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Mire, G.; Siah, A.; Brisset, M.N.; Gaucher, M.; Deleu, M.; Jijakli, M.H. Surfactin protects wheat against Zymoseptoria tritici and activates both salicylic acid- and jasmonic acid-dependent defense responses. Agriculture 2018, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Queval, G.; Noctor, G. A plate reader method for the measurement of NAD, NADP, glutathione, and ascorbate in tissue extracts: Application to redox profiling during Arabidopsis rosette development. Anal. Biochem. 2007, 363, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Vergnes, S.; Ladouce, N.; Fournier, S.; Ferhout, H.; Attia, F.; Dumas, B. Foliar treatments with Gaultheria procumbens essential oil induce defense responses and resistance against a fungal pathogen in Arabidopsis. Front. Plant Sci. 2014, 5, 477. [Google Scholar] [CrossRef]
- Bernonville, T.D.d.; Gaucher, M.; Guyot, S.; Durel, C.E.; Dat, J.F.; Brisset, M.N. The constitutive phenolic composition of two Malus×domestica genotypes is not responsible for their contrasted susceptibilities to fire blight. Environ. Exp. Bot. 2011, 74, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Ördög, A.; Csiszár, J.; Poór, P. Plant glutathione transferases and light. Front. Plant Sci. 2019, 9, 1944. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.J. The integration of glutathione homeostasis and redox signaling. J. Plant Physiol. 2008, 165, 1390–1403. [Google Scholar] [CrossRef]
- Tamburini, E.; Ferrari, G.; Marchetti, M.; Pedrini, P.; Ferro, S. Development of FT-NIR models for the simultaneous estimation of chlorophyll and nitrogen content in fresh apple (Malus domestica) leaves. Sensors 2015, 15, 2662–2679. [Google Scholar] [CrossRef] [Green Version]
- Prieto, C.; Calvo, L. Performance of the Biocompatible Surfactant Tween 80, for the Formation of Microemulsions Suitable for New Pharmaceutical Processing. J. Appl. Chem. 2013, 2013, 930356. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox Homeostasis and Antioxidant Signaling: A Metabolic Interface between Stress Perception and Physiological Responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Zhuang, X.; Zhou, J.; Sun, L.; Wan, H.; Li, H.; Lyu, D. Exogenous melatonin alleviates cadmium uptake and toxicity in apple rootstocks. Tree Physiol. 2020, 40, 746–761. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.H.; Ma, F.W.; Wang, Y.H.; Zhang, J.K. The responses of the enzymes related with ascorbate-glutathione cycle during drought stress in apple leaves. Acta Physiol. Plant. 2011, 33, 173–180. [Google Scholar] [CrossRef]
- Ma, Y.H.; Ma, F.W.; Zhang, J.K.; Li, M.J.; Wang, Y.H.; Liang, D. Effects of high temperature on activities and gene expression of enzymes involved in ascorbate–glutathione cycle in apple leaves. Plant Sci. 2008, 175, 761–766. [Google Scholar] [CrossRef]
- Davoine, C.; Falletti, O.; Douki, T.; Iacazio, G.; Ennar, N.; Montillet, J.L.; Triantaphylidès, C. Adducts of Oxylipin Electrophiles to Glutathione Reflect a 13 Specificity of the Downstream Lipoxygenase Pathway in the Tobacco Hypersensitive Response. Plant Physiol. 2006, 140, 1484–1493. [Google Scholar] [CrossRef] [Green Version]
- Muramoto, S.; Matsubara, Y.; Mwenda, C.M.; Koeduka, T.; Sakami, T.; Tani, A.; Matsui, K. Glutathionylation and reduction of methacrolein in tomato plants account for its absorption from the vapor phase. Plant Physiol. 2015, 169, 1744–1754. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, K.; Matsui, K.; Takabayashi, J. Uptake and Conversion of Volatile Compounds in Plant–Plant Communication. In Deciphering Chemical Language of Plant Communication; Springer: Cham, Switzerland, 2016; pp. 305–316. [Google Scholar]
- Mano, J. Reactive carbonyl species: Their production from lipid peroxides, action in environmental stress, and the detoxification mechanism. Plant Physiol. Biochem. 2012, 59, 90–97. [Google Scholar] [CrossRef]
- Chaimovitsh, D.; Shachter, A.; Abu-Abied, M.; Rubin, B.; Sadot, E.; Dudai, N. Herbicidal Activity of Monoterpenes Is Associated with Disruption of Microtubule Functionality and Membrane Integrity. Weed Sci. 2017, 65, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Bozari, S.; Agar, G.; Aksakal, O.; Erturk, F.A.; Yanmis, D. Determination of chemical composition and genotoxic effects of essential oil obtained from Nepeta nuda on Zea mays seedlings. Toxicol. Ind. Health 2013, 29, 339–348. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Pierpoint, W.S.; Boller, T.; Conejero, V. Recommendations for naming plant pathogenesis-related proteins. Plant Mol. Biol. Report. 1994, 12, 245–264. [Google Scholar] [CrossRef]
- Bonasera, J.M.; Kim, J.F.; Beer, S.V. PR genes of apple: Identification and expression in response to elicitors and inoculation with Erwinia amylovora. BMC Plant Biol. 2006, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.D.M.; Varanda, C.M.R.; Félix, M.R.F. Induced resistance during the interaction pathogen x plant and the use of resistance inducers. Phytochem. Lett. 2016, 15, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Saboki, E.; Usha, K.; Singh, B. Pathogenesis Related (PR) Proteins in Plant Defense Mechanism Age-Related Pathogen Resistance. Curr. Res. Technol. Adv. 2011, 2, 1043–1054. [Google Scholar]
- Le Mire, G. Identification of Elicitors Inducing Resistance in Wheat against Zymoseptoria Tritici and Characterization of the Subsequent Triggered Defense-Signaling Pathways. 2018. Available online: https://orbi.uliege.be/handle/2268/222563 (accessed on 2 February 2022).
- Saini, S.S.; Teotia, D.; Gaid, M.; Thakur, A.; Beerhues, L.; Sircar, D. Benzaldehyde dehydrogenase-driven phytoalexin biosynthesis in elicitor-treated Pyrus pyrifolia cell cultures. J. Plant Physiol. 2017, 215, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Chizzali, C.; Gaid, M.M.; Belkheir, A.K.; Hänsch, R.; Richter, K.; Flachowsky, H.; Peil, A.; Hanke, M.V.; Liu, B.; Beerhues, L. Differential Expression of Biphenyl Synthase Gene Family Members in Fire-Blight-Infected Apple ‘Holsteiner Cox’. Plant Physiol. 2012, 158, 864–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkate, A.; Saini, S.S.; Teotia, D.; Gaid, M.; Mir, J.I.; Roy, P.; Agrawal, P.K.; Sircar, D. Comparative metabolomics of scab-resistant and susceptible apple cell cultures in response to scab fungus elicitor treatment. Sci. Rep. 2018, 8, 17844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werrie, P.-Y.; Burgeon, C.; Le Goff, G.J.; Hance, T.; Fauconnier, M.-L. Biopesticide Trunk Injection Into Apple Trees: A Proof of Concept for the Systemic Movement of Mint and Cinnamon Essential Oils. Front. Plant Sci. 2021, 12, 495. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, N.; Liu, R.; Bai, H.; Tao, W.; Chen, J.; Shi, Z. Cinnamaldehyde Facilitates Cadmium Tolerance by Modulating Ca2+ in Brassica rapa. Water. Air. Soil Pollut. 2021, 232, 19. [Google Scholar] [CrossRef]
- Xue, Y.F.; Zhang, M.; Qi, Z.Q.; Li, Y.Q.; Shi, Z.; Chen, J. Cinnamaldehyde promotes root branching by regulating endogenous hydrogen sulfide. J. Sci. Food Agric. 2016, 96, 909–914. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Werrie, P.-Y.; Juillard, A.; Heintz, C.; Brisset, M.-N.; Fauconnier, M.-L. Phytotoxicity and Plant Defence Induction by Cinnamomum cassia Essential Oil Application on Malus domestica Tree: A Molecular Approach. Agronomy 2022, 12, 512. https://doi.org/10.3390/agronomy12020512
Werrie P-Y, Juillard A, Heintz C, Brisset M-N, Fauconnier M-L. Phytotoxicity and Plant Defence Induction by Cinnamomum cassia Essential Oil Application on Malus domestica Tree: A Molecular Approach. Agronomy. 2022; 12(2):512. https://doi.org/10.3390/agronomy12020512
Chicago/Turabian StyleWerrie, Pierre-Yves, Anthony Juillard, Christelle Heintz, Marie-Noëlle Brisset, and Marie-Laure Fauconnier. 2022. "Phytotoxicity and Plant Defence Induction by Cinnamomum cassia Essential Oil Application on Malus domestica Tree: A Molecular Approach" Agronomy 12, no. 2: 512. https://doi.org/10.3390/agronomy12020512
APA StyleWerrie, P.-Y., Juillard, A., Heintz, C., Brisset, M.-N., & Fauconnier, M.-L. (2022). Phytotoxicity and Plant Defence Induction by Cinnamomum cassia Essential Oil Application on Malus domestica Tree: A Molecular Approach. Agronomy, 12(2), 512. https://doi.org/10.3390/agronomy12020512