
Physiological and Agronomical Responses of ‘Vairo’ Almond and ‘Big Top’ Nectarine Cultivars Grafted onto Different Prunus Rootstocks and Grown under Semiarid Mediterranean Conditions

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Site Description and Experimental Design
2.1.1. Almond
2.1.2. Peach
2.2. Agronomical Assessments before Harvest
2.2.1. Almond
2.2.2. Peach
2.3. Physiological Assessments before Harvest
2.3.1. Leaf Chlorophyll Concentration
2.3.2. Quantum Yield of Photosynthesis
2.3.3. Stem Water Potential
2.3.4. Stomatal Conductance
2.4. Agronomical Assessments at Harvest
2.4.1. Almond
2.4.2. Peach
2.5. Statistical Analysis
3. Results
3.1. Agronomical and Physiological Assessments before Harvest
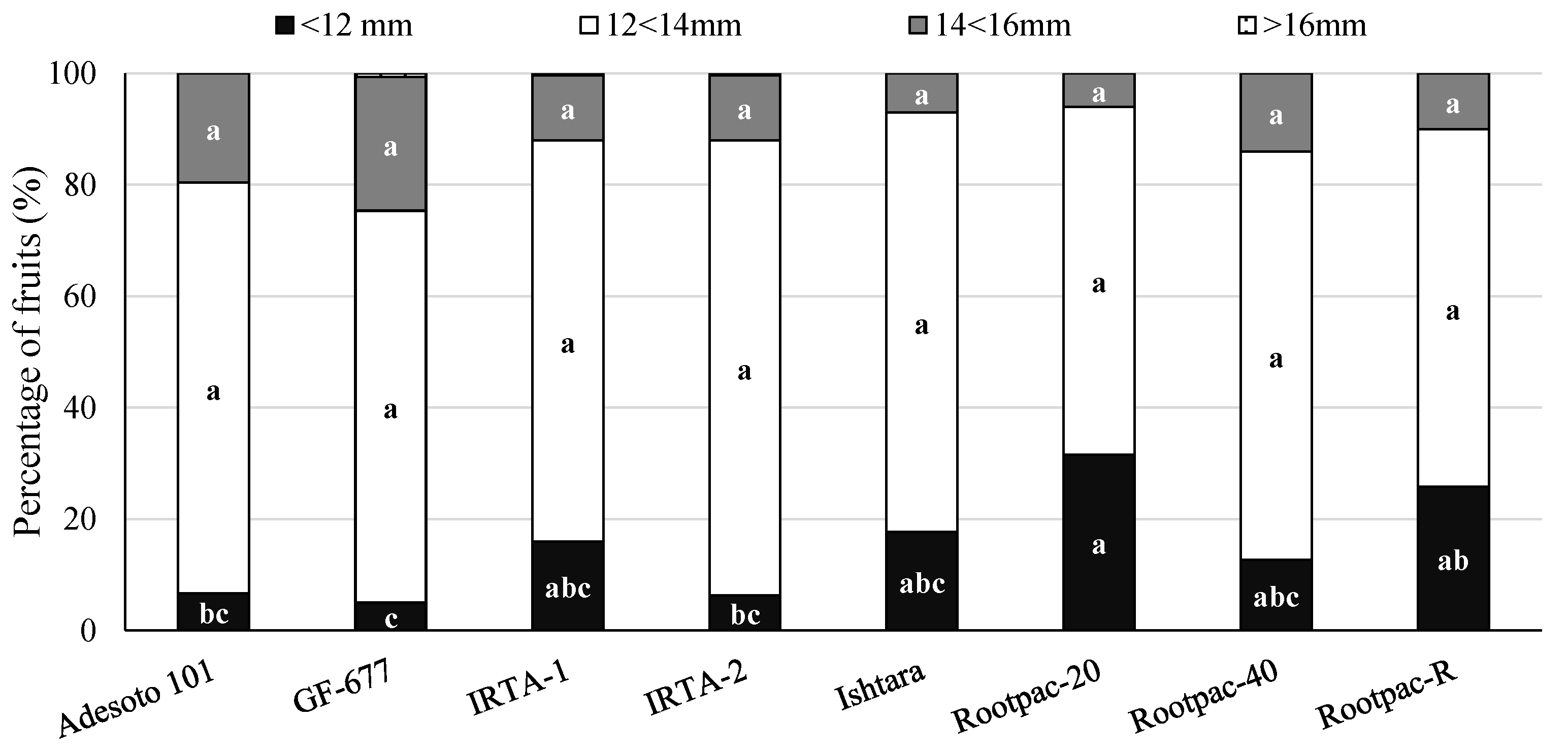
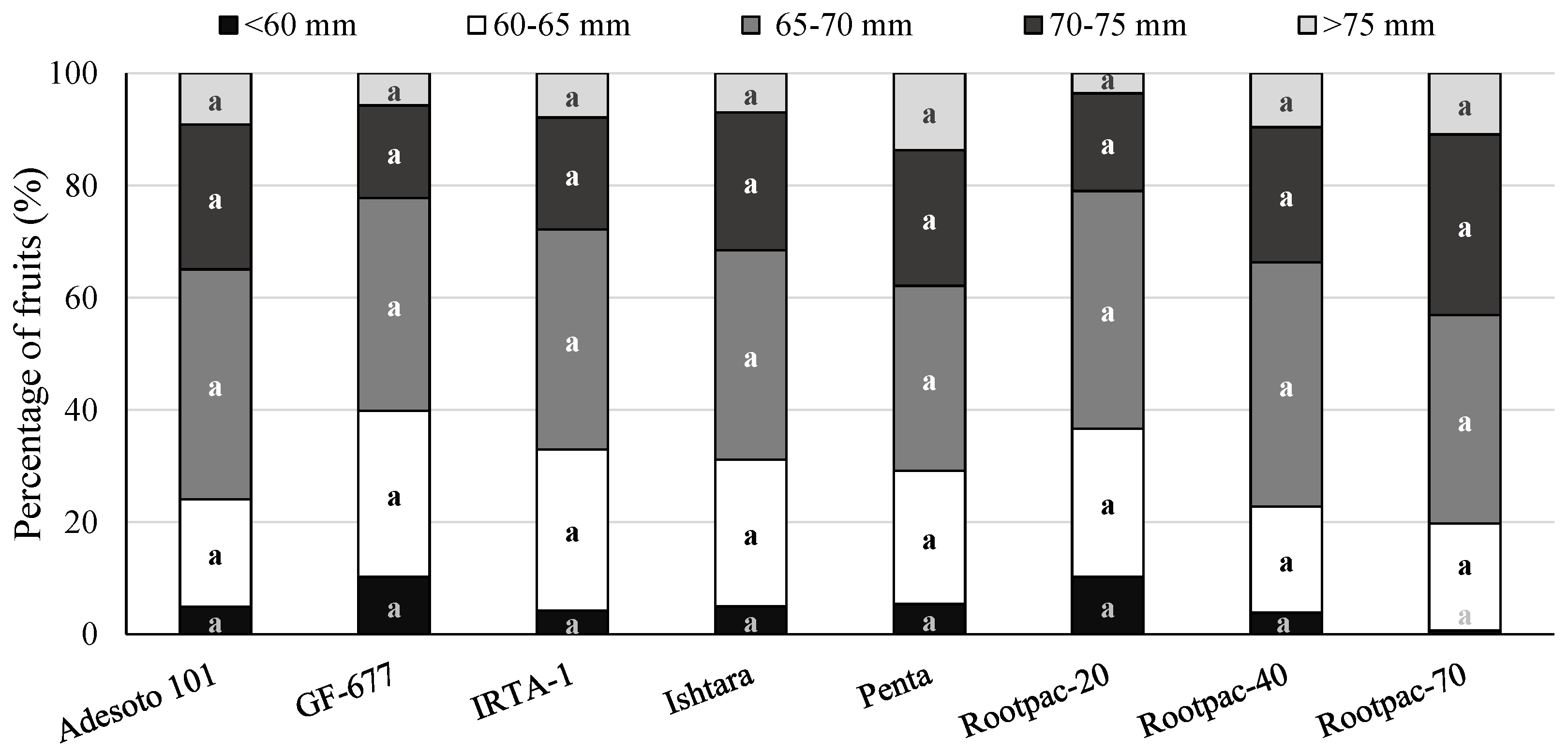
3.2. Agronomical Assessments at Harvest
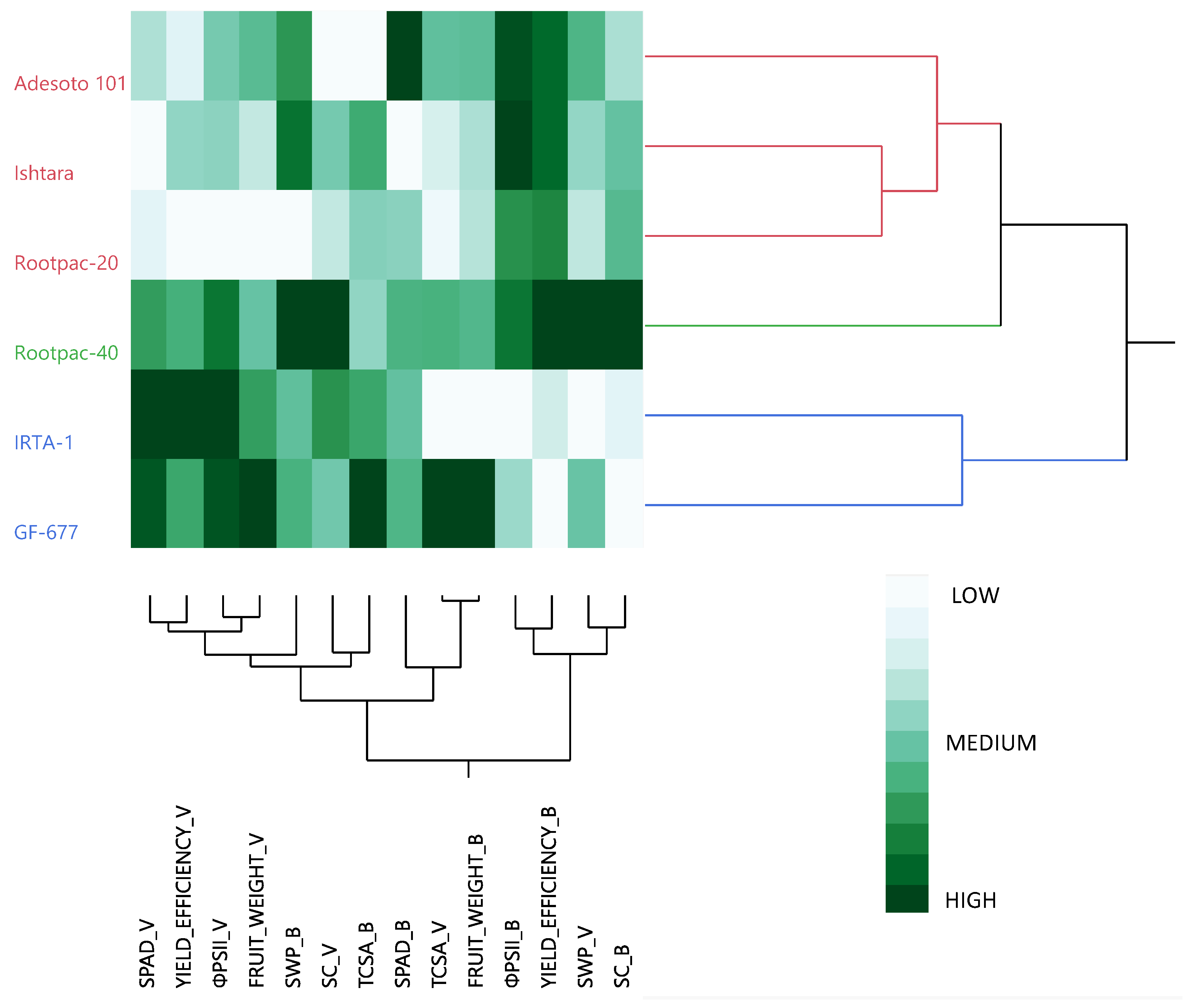
3.3. Overall Rootstock Performance and Interaction with Almond and Peach
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bassi, D.; Monet, R. Botany and Taxonomy. In The Peach: Botany, Production and Uses; Layne, D.R., Bassi, D., Eds.; CABI Publishing: Oxfordshire, UK, 2008; pp. 1–36. [Google Scholar]
- Socias i Company, R.; Ansón, J.M.; Espiau, M.T. Taxonomy, Botany and Physiology. In Almonds: Botany, Production and Uses; Socias i Company, R., Gradziel, T.M., Eds.; CABI Publishing: Oxfordshire, UK, 2017; pp. 1–41. [Google Scholar]
- Gradziel, T.M.; Martínez-Gómez, P. Almond breeding. In Plant Breeding Reviews; Jacnick, J., Ed.; Wiley & Blackwel: New York, NY, USA, 2013; Volume 37, pp. 207–258. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). The Importance of Ukraine and the Russian Federation for Global Agricultural Markets. Available online: https://www.fao.org (accessed on 15 December 2021).
- International Nut Council. Almond Working Group. Statistical Year Book. 2020–2021. Available online: https://www.nutfruit.org (accessed on 15 December 2021).
- MAPA. Short-Term Outlook for Eu Agricultural Markets in 2019 and 2020. Available online: https://www.mapa.gob.es (accessed on 15 December 2021).
- Iglesias, I.; Foles, P.; Oliveira, C. El cultivo del almendro en España y Portugal: Situación, innovación tecnológica, costes, rentabilidad y perspectivas. Rev. Frutic. 2021, 81, 6–49. [Google Scholar]
- Iglesias, I.; Echeverría, G. Current situation, trends and challenges for efficient and sustainable peach production. Sci. Hortic. 2022, 296, 110899. [Google Scholar] [CrossRef]
- Iglesias, I. Situación actual e innovación tecnológica en fruticultura: Una apuesta por la eficiencia y la sostenibilidad. Rev. De Frutic. 2022, 85, 6–45. [Google Scholar]
- Zhang, Y.; Barthe, G.; Grosser, J.W.; Wang, N. Transcriptome analysis of root response to Citrus blight bas on the newly as-sembled swingle citrumelo draft genome. BMC Genom. 2016, 17, 485. [Google Scholar] [CrossRef] [Green Version]
- Boini, A.; Manfrini, L.; Bortolotti, G.; Corelli-Grappadelli, L.; Morandi, B. Monitoring fruit daily growth indicates the onset of mild drought stress in apple. Sci. Hortic. 2019, 256, 108520. [Google Scholar] [CrossRef]
- Bellvert, J.; Nieto, H.; Pelechá, A.; Jofre-Cekalovic, C.; Zazurca, L.; Miarnau, X. Remote sensing energy balance model for the assessment of crop evapotranspiration and water status in an almond rootstock collection. Front. Plant Sci. 2021, 12, 608967. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, V.; Lopez-Lopez, M.; Espadafor, M.; Orgaz, F.; Testi, L.; Zarco-Tejada, P.J.; Lorite, I.J.; Fereres, E. Transpiration from canopy temperature: Implications for the assessment of crop yield in almond orchards. European. J. Agron. 2019, 105, 78–85. [Google Scholar] [CrossRef]
- Hernandez-Santana, V.; Fernández, J.E.; Rodriguez-Dominguez, C.M.; Romero, R.; Diaz-Espejo, A. The dynamics of radial sap flux density reflects changes in stomatal conductance in response to soil and air water deficit. Agric. Forest Meteorol. 2016, 218, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Yahmed, J.B.; Ghrab, M.; Mimoun, M.B. Eco-physiological evaluation of different scion-rootstock combinations of almond grown in Mediterranean conditions. Fruits 2016, 71, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, I. La intensificación sostenible como respuesta al Pacto Verde de la Unión Europea: Retos y ejemplos en la producción frutícola y en el consumo alimentario. Rev. Frutic. 2020, 79, 45–57. [Google Scholar]
- Maldonado, M.; Torguet, L.; Girabet, R.; Zazurca, L.; Martinez, G.; Miarnau, X. Nuevos modelos productivos para la intensi-ficación del almendro. Vida Rural 2019, 22, 50–55. [Google Scholar]
- Miarnau, X.; Torguet, L.; Batlle, I.; Alegre, S. El cultivo del almendro en alta densidad. Rev. Frutic. 2016, 49, 68–87. [Google Scholar]
- Díez-Palet, I.; Funes, I.; Savé, R.; Biel, C.; de Herralde, F.; Miarnau, X.; Vargas, F.; Àvila, G.; Carbó, J.; Aranda, X. Blooming under Mediterranean climate: Estimating cultivar-specific chill and heat requirements of almond and apple trees using a statistical approach. Agronomy 2019, 9, 760. [Google Scholar] [CrossRef] [Green Version]
- Gogorcena, Y.; Sánchez, G.; Moreno-Vázquez, S.; Pérez, S.; Ksouri, N. Genomic-Based Breeding for Climate-Smart Peach Varieties. In Genomic Designing of Climate-Smart Fruit Crops; Kole, C., Ed.; Springer Nature: Cham, Germany, 2020; pp. 271–332. [Google Scholar]
- Penso, G.A.; Citadin, I.; Scariotto, S.; Magalhae dos Santos, C.E.; Junior, A.W.; Bruckner, C.H.; Rodrigo, J. Development of peach flower buds under low Winter chilling conditions. Agronomy 2020, 10, 428. [Google Scholar] [CrossRef] [Green Version]
- Prudencio, A.S.; Sánchez-Pérez, R.; Martínez-García, P.J.; Dicenta, F.; Gradziel, T.M.; Martínez-Gómez, P. Genomic Designing for New Climate-Resilient Almond Varieties. In Genomic Designing of Climate-Smart Fruit Crops; Kole, C., Ed.; Springer Nature: Cham, Germany, 2020; pp. 1–22. [Google Scholar]
- Paniagua, L.L.; García-Martín, A.; Moral, F.J.; Rebollo, F.J. Aridity in the Iberian Peninsula (1960–2017): Distribution, tenden-cies, and changes. Theor. Appl. Climatol. 2019, 138, 811–830. [Google Scholar] [CrossRef]
- Ozturk, T.; Zeynep, P.C.; Turkes, M.; Levent Kurnaz, M. Projections of climate change in the Mediterranean Basin by using downscaled global climate model outputs. Int. J. Climatol. 2015, 35, 4276–4292. [Google Scholar] [CrossRef]
- Font i Forcada, C.; Reig, G.; Mestre, L.; Mignard, P.; Betrán, J.A.; Moreno, M.A. Scion × rootstock response on production, mineral composition, and fruit quality under heavy-calcareous soil and hot climate. Agronomy 2020, 10, 1159. [Google Scholar] [CrossRef]
- Jiménez, S.; Dridi, J.; Gutiérrez, D.; Moret, D.; Irigoyen, J.J.; Moreno, M.A.; Gogorcena, Y. Physiological, biochemical and molecular responses in four Prunus rootstocks submitted to drought stress. Tree Physiol. 2013, 33, 1061–1075. [Google Scholar] [CrossRef]
- Lordan, J.; Zazurca, L.; Maldonado, M.; Torguet, L.; Alegre, S.; Miarnau, X. Horticultural performance of ‘Marinada’ and ‘Vairo’ almond cultivars grown on a genetically diverse set of rootstocks. Sci. Hortic. 2019, 256, 108558. [Google Scholar] [CrossRef]
- Reig, G.; Garanto, X.; Mas, N.; Iglesias, I. Long-term agronomical performance and iron chlorosis susceptibility of several Prunus rootstocks grown under loamy and calcareous soil conditions. Sci. Hortic. 2020, 262, 10935. [Google Scholar] [CrossRef]
- Yahmed, J.B.; Ghrab, M.; Moreno, M.A.; Pinochet, J.; Mimoun, M.B. Performance of ‘Subirana’ flat peach cultivar budded on different Prunus rootstocks in a warm production area in North Africa. Sci. Hortic. 2016, 206, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Gascòn, J.; Figueras-Panillo, M.; Iglesias-Castellarnau, J.; Martin-Ramos, P. Comparison of SHD and ppen-center training systems in almond tree orchards cv. “Soleta”. Agronomy 2019, 9, 874. [Google Scholar] [CrossRef] [Green Version]
- Maldera, F.; Vivaldi, G.A.; Iglesias-Castellarnau, I.; Camposeo, S. Row orientation and canopy position affect bud differentia-tion, LAI and some agronomical traits of a super high-density almond orchard. Agronomy 2021, 11, 251. [Google Scholar] [CrossRef]
- Maldera, F.; Vivaldi, G.A.; Iglesias-Castellarnau, I.; Camposeo, S. Two almond cultivars trained in a super-high density orchard show different growth, yield efficiencies and damages by mechanical harvesting. Agronomy 2021, 11, 1406. [Google Scholar] [CrossRef]
- Vargas, F.J.; Romero, M.; Clave, J.; Verges, J.; Santos, J.; Batlle, I. Vairo’, ‘Marinada’, ‘Constanti’, and ‘Tarraco’ almonds. HortScience 2008, 43, 535–537. [Google Scholar] [CrossRef] [Green Version]
- Vargas, F.J.; Romero, M.A.; Clave, J.; Batlle, I.; Miarnau, X.; Alegre, S. Important traits in IRTA’s new almond cultivars. Acta Hortic. 2011, 912, 359–365. [Google Scholar] [CrossRef]
- Iglesias, I.; Echeverría, G. Differential effect of cultivar and harvest date on nectarine colour, quality and consumer acceptance. Sci. Hortic. 2009, 120, 41–50. [Google Scholar] [CrossRef]
- Iglesias, I.; Echeverría, G. Overview of peach industry in the European Union with special reference to Spain. Acta Hortic. 2021, 1304, 163–176. [Google Scholar] [CrossRef]
- Reig, G.; Alegre, S.; Cantín, C.M.; Gatius, F.; Puy, J.; Iglesias, I. Tree ripening and postharvest firmness loss of eleven commercial nectarine cultivars under Mediterranean conditions. Sci. Hortic. 2017, 219, 335–343. [Google Scholar] [CrossRef]
- Sakar, E.H.; Yamani, M.E.; Rharrabti, Y. Geometrical traits in almond fruit as affected by genotypic and environmental variations in Northern Morocco. Erwerbs-Obstbau 2019, 61, 103–112. [Google Scholar] [CrossRef]
- Toro, G.; Flexas, J.; Escalona, J.M. Contrasting leaf porometer and infra-red gas analyser methodologies: An oldl paradigm about the stomatal conductance measurement. Theor. Exp. Plant Physiol. 2019, 31, 483–492. [Google Scholar] [CrossRef]
- Kuiper, F.K.; Fisher, L. A Monte Carlo comparison of six clustering procedures. Biometrics 1975, 31, 777–783. [Google Scholar] [CrossRef]
- Ward, J.H., Jr. Hierarchical groupings to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Mestre, L.; Reig, G.; Betrán, J.A.; Pinochet, J.; Moreno, M.A. Influence of peach–almond hybrids and plum-based rootstocks on mineral nutrition and yield characteristics of ‘Big Top’ nectarine in replant and heavy-calcareous soil conditions. Sci. Hortic. 2015, 215, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Reig, G.; Font i Forcada, C.; Mestre, L.; Betrán, J.A.; Moreno, M.A. Potential of new Prunus cerasifera based rootstocks for adapting under heavy and calcareous soil conditions. Sci. Hortic. 2018, 234, 193–200. [Google Scholar] [CrossRef]
- Conejero, W.; Mellisho, C.D.; Ortuño, M.F.; Galindo, A.; Pérez-Sarmiento, F.; Torrecillas, A. Establishing maximum daily trunk shrinkage and midday stem water potential reference equations for irrigation scheduling of early maturing peach trees. Irrig. Sci. 2011, 29, 299–309. [Google Scholar] [CrossRef]
- Reig, G.; Lordan, J.; Fazio, G.; Grusak, M.A.; Hoying, S.; Cheng, L.; Francescatto, P.; Robinson, T. Horticultural performance and elemental nutrient concentrations on ‘Fuji’ grafted on apple rootstocks under New York climatic conditions. Sci. Hortic. 2018, 227, 22–37. [Google Scholar] [CrossRef]
- Reig, G.; Lordan, J.; Miranda, M.; Hoying, S.A.; Fargione, M.; Reginato, G.; Donahue, D.J.; Francescatto, P.; Fazio, G.; Robinson, T. Long-term performance of ‘Gala’, Fuji’ and ‘Honeycrisp’ apple trees grafted on Geneva® rootstocks and trained to four production systems under New York State climatic conditions. Sci. Hortic. 2019, 244, 277–293. [Google Scholar] [CrossRef]
- Musacchi, S.; Neri, D.; Iglesias, I. Training systems and sustainable orchard management for European pear (Pyrus communis L.) in the Mediterranean area: A Review. Agronomy 2021, 11, 1765. [Google Scholar] [CrossRef]
- Reig, G.; Mestre, L.; Betrán, J.A.; Pinochet, J.; Moreno, M.A. Agronomic and physicochemical fruit properties of ‘Big Top’ nectarine budded on peach and plum based rootstocks in Mediterranean conditions. Sci. Hortic. 2016, 210, 85–92. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Rootstock | Genetics | Origin a |
|---|---|---|---|
| ‘Vairo’ | Adesoto® 101 | P. insititia (open pollination of Pollizo de Murcia) | CSIC, Spain |
| INRA® GF-677 | P. amygdalus × P. persica | INRA, France | |
| IRTA-1 | P. amygdalus × P. persica | IRTA, Spain | |
| IRTA-2 | P. cerasifera × P. dulcis | IRTA, Spain | |
| Ishtara® (Ferciana) | (P. cerasifera × P. salicina) × (P. cerasifera × P. persica) | INRA, France | |
| Rootpac®20 (Densipac) | P. besseyi × P. cerasifera | AI, Spain | |
| Rootpac®40 (Nanopac) | (P. amydalus × P. persica) × (P. amydalus × P. persica) | AI, Spain | |
| Rootpac®R (Replantpac) | P. cerasifera × P. dulcis | AI, Spain | |
| ‘Big Top’ | Adesoto® 101 | P. insititia (open pollination of Pollizo de Murcia) | CSIC, Spain |
| INRA® GF-677 | P. amygdalus × P. persica | INRA, France | |
| IRTA-1 | P. amygdalus × P. persica | IRTA, Spain | |
| Ishtara® (Ferciana) | (P. cerasifera × P. salicina) × (P. cerasifera × P. persica) | INRA, France | |
| Penta | P. domestica | CREA Rome, Italy | |
| Rootpac®20 (Densipac) | P. besseyi × P. cerasifera | AI, Spain | |
| Rootpac®40 (Nanopac) | (P. amydalus × P. persica) × (P. amydalus × P. persica) | AI, Spain | |
| Rootpac®70 (Redpac) | (P. persica × P. davidiana) × (P. amygdalus × P. persica) | AI, Spain |
| Cultivar | Variable a | Experimental Factor | DF | F Value | Prob > F |
|---|---|---|---|---|---|
| ‘Vairo’ | FV | Rootstock (R) | 7 | 5.4126 | 0.0003 |
| Date (D) | 4 | 3.8528 | 0.0052 | ||
| R × D | 28 | 1.1000 | 0.3453 | ||
| KV | Rootstock (R) | 7 | 7.3616 | <0.0001 | |
| Date (D) | 4 | 8.8201 | <0.0001 | ||
| R × D | 28 | 0.6491 | 0.9099 | ||
| SPAD | Rootstock (R) | 7 | 7.6642 | <0.0001 | |
| Date (D) | 7 | 107.8379 | <0.0001 | ||
| R × D | 49 | 1.8097 | 0.0007 | ||
| ΦPSII | Rootstock (R) | 7 | 28.2668 | <0.0001 | |
| Date (D) | 7 | 79.1444 | <0.0001 | ||
| R × D | 49 | 1.1317 | 0.2508 | ||
| SWP | Rootstock (R) | 7 | 7.6969 | <0.0001 | |
| Date (D) | 7 | 155.8832 | <0.0001 | ||
| R × D | 49 | 1.7082 | 0.0025 | ||
| SC | Rootstock (R) | 7 | 10.1479 | <0.0001 | |
| Date (D) | 7 | 127.2204 | <0.0001 | ||
| R × D | 49 | 2.1929 | <0.0001 | ||
| ‘Big Top’ | FD | Rootstock (R) | 7 | 0.6391 | 0.7189 |
| Date (D) | 5 | 1605.9050 | <0.0001 | ||
| R × D | 35 | 0.3718 | 0.9994 | ||
| SPAD | Rootstock (R) | 7 | 1.3513 | 0.2764 | |
| Date (D) | 5 | 1.4189 | 0.2223 | ||
| R × D | 35 | 0.7191 | 0.8691 | ||
| ΦPSII | Rootstock (R) | 7 | 1.2210 | 0.3352 | |
| Date (D) | 3 | 4.5479 | 0.0039 | ||
| R × D | 21 | 1.0670 | 0.3827 | ||
| SWP | Rootstock (R) | 7 | 2.0320 | 0.0987 | |
| Date (D) | 3 | 90.6405 | <0.0001 | ||
| R × D | 21 | 2.5176 | 0.0005 | ||
| SC | Rootstock (R) | 7 | 2.7790 | 0.0326 | |
| Date (D) | 3 | 63.9466 | <0.0001 | ||
| R × D | 21 | 1.3391 | 0.1539 |
| ‘Vairo’ | ‘Big Top’ | ||||
|---|---|---|---|---|---|
| Rootstock | FV a,b (cm3) | KV a,b (cm3) | ΦPSII a,b | Rootstock | SC a,b |
| Adesoto 101 | 36.5 ab | 6.1 ab | 0.76 a | Adesoto 101 | 407.9 ab |
| GF-677 | 38.8 a | 6.8 a | 0.76 a | GF-677 | 432.8 a |
| IRTA-1 | 35.6 ab | 6.2 ab | 0.76 a | IRTA-1 | 439.9 a |
| IRTA-2 | 35.9 ab | 6.1 ab | 0.76 a | Ishtara | 467.6 a |
| Ishtara | 34.4 ab | 5.8 b | 0.75 ab | Penta | 365.3 b |
| Rootpac-20 | 32.5 b | 4.8 c | 0.74 b | Rootpac-20 | 462.9 a |
| Rootpac-40 | 39.1 a | 5.9 ab | 0.76 a | Rootpac-40 | 463.5 a |
| Rootpac-R | 32.7 b | 5.6 bc | 0.76 a | Rootpac-70 | 465.1 a |
| p ≤ 0.05 | 0.0003 | <0.0001 | <0.0001 | p ≤ 0.05 | 0.0326 |
| Rootstock | TCSA a,b (cm2) | Yield b (Kernel tree−1) | Yield b,c (Kernel ha−1) | Kernel Efficiency b (kg cm−2) | Percentage Vigor d | Theoretical Kernel Yield b,e (kg ha−1) | Shelling b (%) | Kernel Weight b (g) |
|---|---|---|---|---|---|---|---|---|
| Adesoto 101 | 242.3 bc | 7.9 cd | 3035.5 cde | 0.032 cd | 76.4 | 3739.9 cd | 28.0 d | 0.88 abc |
| GF-677 | 317.9 ab | 13.9 a | 6971.8 a | 0.043 ab | 100.0 | 6971.8 a | 31.9 ab | 0.98 a |
| IRTA-1 | 200.0 c | 9.1 bc | 4272.5 bc | 0.052 a | 52.1 | 6020.2 ab | 32.7 a | 0.91 ab |
| IRTA-2 | 345.9 a | 10.5 b | 4224.6 bcd | 0.031 cd | 110.9 | 3681.8 cd | 30.7 abc | 0.91 ab |
| Ishtara | 215.8 c | 7.9 cd | 3227.7 bcde | 0.037 bcd | 68.9 | 4212.9 bcd | 29.4 cd | 0.82 bcd |
| Rootpac-20 | 204.7 c | 5.5 d | 2122.5 e | 0.029 d | 63.6 | 2906.5 d | 29.1 cd | 0.76 d |
| Rootpac-40 | 255.5 bc | 10.7 b | 4674.7 b | 0.042 abc | 81.9 | 5442.3 abc | 29.6 bcd | 0.87 bc |
| Rootpac-R | 229.7 c | 6.6 cd | 2696.6 de | 0.029 d | 72.5 | 3435.1 d | 31.7 abc | 0.79 cd |
| p ≤ 0.05 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | - | <0.0001 | <0.0001 | <0.0001 |
| Rootstock | TCSA a,b (cm2) | Yield b (kg tree−1) | Yield b,c (kg ha−1) | Yield Efficiency b (kg cm−2) | Percentage Vigor d | Theoretical Yield e (kg ha−1) | Fruit Weight b (g) |
|---|---|---|---|---|---|---|---|
| Adesoto 101 | 120.5 b | 37.2 ab | 28,626.0 ab | 0.31 a | 74.2 | 35,772.1 a | 163.6 a |
| GF-677 | 169.1 b | 31.4 b | 24,146.6 ab | 0.19 ab | 100.0 | 24,146.6 ab | 174.5 a |
| IRTA-1 | 153.5 b | 36.1 ab | 27,780.1 ab | 0.23 ab | 93.5 | 29,441.0 ab | 154.8 a |
| Ishtara | 152.7 b | 47.2 a | 36,296.8 a | 0.31 a | 93.0 | 37,746.1 a | 160.1 a |
| Penta | 147.1 b | 29.1 b | 22,379.8 b | 0.20 ab | 88.6 | 25,018.1 ab | 164.7 a |
| Rootpac-20 | 142.7 b | 42.8 ab | 32,938.8 ab | 0.30 ab | 91.7 | 35,842.2 a | 159.7 a |
| Rootpac-40 | 141.1 b | 42.2 ab | 32,471.0 ab | 0.32 a | 80.6 | 38,554.9 a | 164.4 a |
| Rootpac-70 | 227.2 a | 37.1 ab | 27,981.3 ab | 0.15 b | 136.3 | 17,822.0 b | 171.4 a |
| p ≤ 0.5 | <0.0001 | 0.0295 | 0.0295 | 0.0023 | - | 0.0028 | 0.9192 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reig, G.; Iglesias, I.; Zazurca, L.; Torguet, L.; Martinez, G.; Miarnau, X. Physiological and Agronomical Responses of ‘Vairo’ Almond and ‘Big Top’ Nectarine Cultivars Grafted onto Different Prunus Rootstocks and Grown under Semiarid Mediterranean Conditions. Agronomy 2022, 12, 821. https://doi.org/10.3390/agronomy12040821
Reig G, Iglesias I, Zazurca L, Torguet L, Martinez G, Miarnau X. Physiological and Agronomical Responses of ‘Vairo’ Almond and ‘Big Top’ Nectarine Cultivars Grafted onto Different Prunus Rootstocks and Grown under Semiarid Mediterranean Conditions. Agronomy. 2022; 12(4):821. https://doi.org/10.3390/agronomy12040821
Chicago/Turabian StyleReig, Gemma, Ignasi Iglesias, Lourdes Zazurca, Laura Torguet, Guillem Martinez, and Xavier Miarnau. 2022. "Physiological and Agronomical Responses of ‘Vairo’ Almond and ‘Big Top’ Nectarine Cultivars Grafted onto Different Prunus Rootstocks and Grown under Semiarid Mediterranean Conditions" Agronomy 12, no. 4: 821. https://doi.org/10.3390/agronomy12040821
APA StyleReig, G., Iglesias, I., Zazurca, L., Torguet, L., Martinez, G., & Miarnau, X. (2022). Physiological and Agronomical Responses of ‘Vairo’ Almond and ‘Big Top’ Nectarine Cultivars Grafted onto Different Prunus Rootstocks and Grown under Semiarid Mediterranean Conditions. Agronomy, 12(4), 821. https://doi.org/10.3390/agronomy12040821