1. Introduction
Medicinal Cannabis (Cannabis sativa) is a new crop to Australia with licenced cultivation approved as recently as 2016 [
1]. In 2021, the Australian Government’s Department of Health, Office of Drug Control (ODC), listed 48 companies and organisations that had been granted licenses to cultivate and produce medicinal Cannabis [
2]. There is a demand for new and improved agronomic cultivation practices to protect and maximise yields and consequently profit margins for this growing industry.
Medicinal Cannabis is cultivated for its terpenophenolic phytocannabinoids, which interact with a mammalian neuronal signalling pathway, the endocannabinoid system [
3,
4]. Over 140 cannabinoids have been identified from Cannabis, but the two main targets for medicinal cultivation are Δ9-tetrahydrocannabinolic acid (THCA) and cannabidiolic acid (CBDA) [
5,
6,
7]. These secondary metabolites when decarboxylated to their neutral forms of Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD) are used for the treatment of non-communicable diseases including multiple sclerosis, childhood epilepsy, appetite stimulation, and sleep disorders [
8,
9,
10,
11,
12]. Recently, cannabigerolic acid (CBGA) and cannabigerovarinic acid (CBGVA) have been shown to have anticonvulsant effects in epilepsy mouse models [
7]. These cannabinoids were bioactive in their acidic forms that are usually decarboxylated during the heating processes used to manufacture Cannabis therapeutics [
7]. They are also considered minor cannabinoids as they are found in significantly lower abundance than THCA and CBDA. Based on this recent finding, it is likely that minor cannabinoids may gain importance as target compounds as clinical and pharmacological research further advances.
Commercial yields of medicinal Cannabis are usually cited as being measured in g dry flower m
2 (or equivalent imperial units) [
13,
14,
15]. Therefore, as with any commercial crop, biomass yield is critical to productivity. However, as medicinal Cannabis is harvested for its cannabinoids, a high concentration of target metabolite g dry flower (eg % CBDA or THCA) is also essential. The unit of m
2 relates to plant density within the growing environment and is therefore a product of infrastructure (e.g., fertigation design, bench ergonomics) and the morphology of the plants (e.g., height, lateral branch number and angle, floral structure). A unit of time is unaccounted for in the cited calculation, but most commercial growers will aim for >5 cycles of production in a 12-month period, so the speed of maturation is also important. The combination of biomass yield, cannabinoid concentration, plant density, and cycle time offers four different ways in which yield can be targeted for improvement.
Grafting is an ancient technique routinely applied for the improvement of horticultural crops including perennial fruit trees and vines, such as citrus and grape, and annuals including tomatoes, watermelon, and capsicum [
16]. It is used globally and extensively, for example, in Korea, an area of more than 23,000 ha was used to cultivate watermelon in 2005 and of these >90% were grafted; in the Netherlands, 75% of commercial tomato plants are grafted [
17]. The benefits of grafting include yield increase, abiotic stress tolerance, biotic stress resistance, quality changes, modified stem architecture, and improved water use efficiency [
17,
18].
Grafting involves the selection of at least two plants, one to provide the rootstock and the other the harvested above-ground growth, the scion. The grafting process connects the sections, resulting in a conjoined vascular system [
19]. Most annual glasshouse crops are grown from seed. The seeds for both plant organs (root and shoot) are germinated, often with staggered timing to ensure that both sections are the optimal size when the graft is made. For example, watermelon seeds that will form a scion are sown 7–8 days after the sowing of the rootstock seeds, and then, grafting takes place a further 7–8 days after the scion germination [
17]. The process of grafting can be summarised in four steps: (1) rootstock and scion selection and production, (2) joining of the two sections through wounding to form a union, (3) fusing of the union, and (4) acclimation of the grafted plant [
17].
Medicinal Cannabis differs from most annual crops cultivated under protected cropping because plants are usually clonally propagated rather than grown from seed. This is because Cannabis is a highly heterozygous, dioecious (and therefore an obligate out-crossing) species that does not produce true-to-type progeny and only the unfertilised female plants produce the prime cannabinoid-laden flowers. Clonal propagation thus removes the risk of producing male plants and ensures a genetically uniform crop. Clones are generated from mother plants that are maintained in a vegetative (non-flowering) state [
20]. New clones are excised from the mother plants, placed into a propagation medium, and maintained under controlled humidity until roots form, and the clone can transition to vegetative growth [
20]. In order to apply grafting to medicinal Cannabis without adapting commercial propagation procedures, both the rootstock and the scion would need to be clonally propagated from vegetative tissue rather than heterozygous seeds. The greatest cost to production for Australian medicinal Cannabis is labour, which accounts for ~46% of production costs [
21]. Therefore, any proposed new cultivation methodology must balance the yield benefit against any additional labour requirements.
We aimed to develop a grafting methodology specifically for medicinal Cannabis that utilized clonal propagation for both rootstock and scion. The method had to require minimal manual handling to keep labour costs down and confer a yield and or/cultivation advantage.
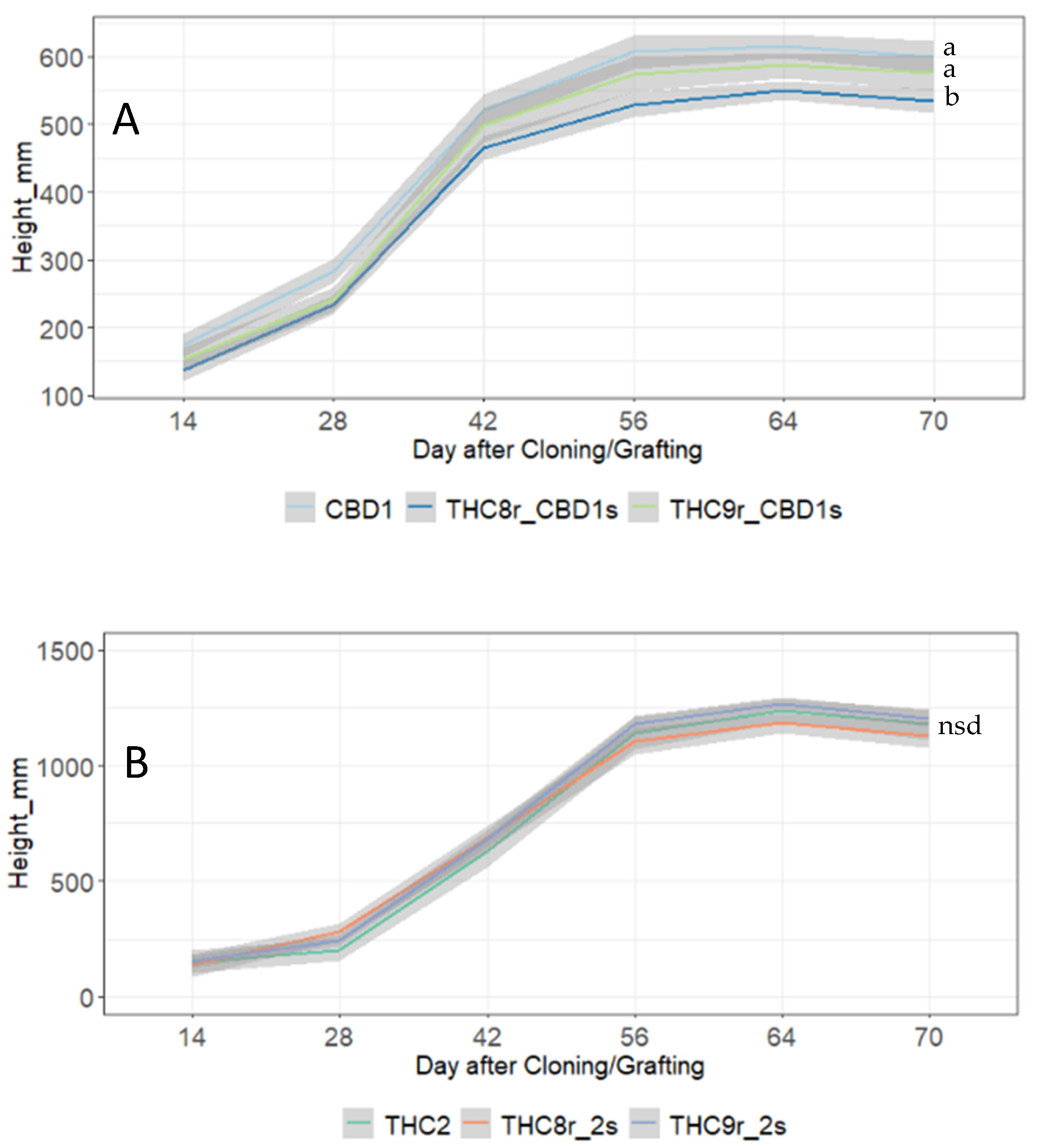
We aimed to improve performance of two commercial lines through grafting. The first, “CBD1”, is a high CBDA accumulating cultivar but exhibits a low flower biomass yield. The second, “THC2”, is a tall, high yielding, high THCA accumulating line that exhibits highly variable root development and therefore results in a high clonal failure rate and inconsistent plant yield performance.
A number of environmental, abiotic, and biotic factors have been cited as affecting cannabinoid content and/or profile [
22]. They include nitrogen fertilization rates [
23], light spectrum [
24,
25,
26], pruning [
27], and drought [
28]. Conversely, however, a number of studies have found little or no impact on cannabinoids of treatments such as flooding, wounding, botrytis infection as a growth regulator [
29], or blue light [
30]. It is currently unknown what role, if any, the root plays in determining the cannabinoid profile. This study aimed to identify root effects on the cannabinoid profile by grafting plants with diverse chemotypes and/or morphology.
2. Materials and Methods
2.1. Plant Material
Eight medicinal Cannabis genotypes were supplied from Cann Group Ltd. The eight varieties are presented in
Table 1. They comprised of one high CBDA producing line and seven high THCA lines.
All plants were cultivated in an ODC approved secure glasshouse facility. All experiments were conducted under a Commonwealth license and associated permits. The temperature was maintained at 25 °C and humidity at 50%. Plants were grown under natural light with supplementary lighting (Heliospectra, Elixia LX6xx, C-Plate, heliospectra.com, 20 March 2022) to extend the photoperiod of vegetative plants. Blackout curtains were used to shorten the photoperiod for flowering plants. Water and nutrients were supplied by a fertigation system. The curtain, climate, and fertigation systems were automated (Priva, priva.com, 20 March 2022).
Experimental plants were cloned from donor mothers. New growth stems of approximately 15 cm were excised from the mother. All leaves up the sides of the stem were removed, leaving the top leaf bunch. The bottom of the stem was then cut diagonally across a node using a scalpel, to form a clone approximately 12 cm in height. The top leaf bunch was trimmed to the height of the smallest emerging leaf to reduce water loss and prevent the clones from overlapping in the propagation dome. The bottom 1 cm of the stem, from which the roots would form, were lightly scraped with a scalpel and then dipped in hormone gel (Clonex Purple, Yates, DuluxGroup, Canterbury-Bankstown, VA, 3169, Australia) and placed in an organic propagation cube (Eazyplug CT12, The Netherlands,
https://www.eazyplug.nl/, accessed on 20 March 2022). Once the propagation tray was full of new clones, it was placed in a propagation dome (Smart Garden heavy duty 3-piece propagation kit, Epping Hydroponics) for 2 weeks under an 18 h light: 6 h dark (18L:6D) photoperiod in a growth cabinet (Conviron A2000,
https://www.conviron.com, accessed on 20 March 2022) at a light intensity of 100 µMol m
2 s
1 and a temperature of 25 °C. Humidity monitors were placed in a dome and the humidity was progressively reduced over the fourteen-day propagation period. Plants with established roots were then potted into 1 l pots containing a 30:70% blend of perlite and coco-coir (Professors Nutrients,
https://www.professorsnutrients.com.au, accessed on 20 March 2022); vegetative growth was fourteen days (except for experiment 3, detailed below) under glasshouse conditions. At the end of the fourteen-day period of vegetative growth, plants that were used in flowering experiments were repotted into 9 L flowering pots containing a 30:70% perlite: coco-coir blend and transferred to the flowering zone in which daylength was reduced to a 12L:12D photoperiod.
All trials were conducted on spaced plants to maximize light penetration and eliminate the risk of shading. Vegetative growth trials followed a fully randomized design on a single bench in the glasshouse, and the flowering trial was designed as a randomized block design across four benches within the flowering zone. Replicate numbers for each experiment are described below.
Experiment 1: The effect of root stock age on grafting success was tested using the highest-yielding, reliably performing genotype, THC7, as a rootstock and the low-yielding, high-CBDA genotype, CBD1, as a scion. The rootstock clones were cut from the donor mother plants successively over four timepoints so that on the day of grafting, they would be 10, 7, 4, and 0 days old. Six rootstocks were cultivated for each treatment. Rootstocks from days 10, 7, and 4 were cloned into coco-peat plugs and placed into a growth cabinet as previously described.
For grafting rootstocks age 4–10 days old, all equipment was cleaned with 70% PA before the procedure. A clone with developing roots was removed from the growth cabinet and laid horizontally on a cutting board. The leaf bunch at the top of the stem was excised with a scalpel to leave a 10 cm length of stem (still implanted in the propagation plug). The stem and propagation plug were returned to an upright position, and the scion was prepared. The scion was cut freshly from a CBD1 mother plant as a 15 cm section. All leaves up the sides of the stem were removed with secateurs, retaining the leaf bunch at the top. The stem was then cut with a scalpel to leave a length of approximately 10 cm (
Figure 1C). The leaf bunch was trimmed to the height of the smallest developing leaf to minimize water loss, and the bottom 1 cm of the stem was slightly shaved on two opposite sides. A 1 cm incision was made vertically into the rootstock stem, and the scion was slid into place (cleft graft) and secured with a 3.3 mm recycled-plastic grafting clip (PT8 3.3, Royal Brinkman).
The day 0 treatment involved cutting fresh clonal stem sections from THC7 mother plants and directly grafting a fresh CBD1 scion (see method below).
2.2. Single Step Grafting Method
All equipment was sterilized using 70% iso-propyl alcohol (PA).
Rootstock preparation. A rootstock stem (THC7, 8 or 9) was freshly cut from the mother, laid on a cutting board, and all leaves up the sides of the stem were removed with secateurs (
Figure 1A,B). The leaf bunch at the top of the stem was excised approximately 1 cm below the lowest leaf with a scalpel, to leave a 10 cm stem section (
Figure 1C). The bottom 1 cm of the stem, from which the roots would form, was lightly scraped with a scalpel. A vertical incision was made centrally down from the top of the stem, where the scion would connect, to a length of approximately 1 cm with a scalpel (
Figure 1E).
A scion approximately 15 cm in length was freshly cut from a CBD1 or THC2 mother plant (
Figure 1A,B). All leaves except the top-most bunch were removed, and the remaining bunch was trimmed to the length of the youngest emerging leaf to limit water loss (
Figure 1D). The area of the scion and rootstock that was to form the union was lightly wiped with a tissue soaked with 70% PA. The base of the scion stem was slightly shaved on two opposite sides using a scalpel (
Figure 1D). The scion was then slid into the incision in the rootstock stem, and a recycled-plastic grafting clip was used to hold the union. A 3.3 mm grafting clip was used (PT8 3.3 mm, Royal Brinkman,
https://www.royalbrinkman.com accessed on accessed on 20 March 2022) (
Figure 1F).
The bottom 1 cm of the rootstock stem was dipped in hormone gel (Clonex Purple, Yates, DuluxGroup, Australia) and then placed in propagation plug (Eazyplug, The Netherlands). Grafted clones were placed in the propagation domes which were sprayed with water and maintained at a humidity of ~100% for 7 days (
Figure 1G). Significant wilting occurs immediately after single-step grafting (
Figure 1H), and an experienced medicinal Cannabis grower may assume the method has failed. The grafts recover from the wilting but may take up to four days post-grafting to resume a healthy appearance (
Figure 1I).
Over the remaining 7 days, the humidity was allowed to progressively decrease until clones with developed roots could be potted out and enter the main glasshouse area (
Table 2).
Experiment 3: Seven high THCA accumulating lines (
Table 1), were used as rootstocks for CBD1 scions utilizing the fresh-on-fresh (day 0) grafting approach (as previously described) and grown for an extended vegetative period of 29 days as a fully randomized trial on a single bench (N = 5–6). The height and fresh weight of the grafted plants was recorded at the end of the trial.
Experiment 4: Two genotypes, THC8 and THC9, were selected as the rootstocks for CBD1 and THC2 scions based on results from Experiment 3. Plants were cultivated as described above and followed the standard production schedule for flowering plants (described above). Eight replicates of all controls (CBD1, THC9, 2, and 8) and grafts THC9r_2s and THC9r_CBD1s were included, but survival of THC8 rootstock grafts was lower, so 7 replicates of THC8r_CBD1s and 5 replicates of THC8r_2s were included. During vegetative growth, the trial followed a fully randomized design on a single bench, but when plants transferred to flowering, a randomized block design was employed to evenly distribute (as much as possible) the replicate plants across the four benches in the flowering area. Bamboo canes and ties were used to support the flowering plants. Plant height was measured every two weeks, and the morphological measures were made on the main stem and six longest side branches per plant when the plants had reached maturity at 76 day after cloning (DAC) 76. Harvest was carried out on 89–91 DAC.
2.3. Harvest Method
Plants were excised at the base, and then, the whole plant was weighed. The large fan leaves were removed, and the flowers were manually stripped from the stem and trimmed using a mechanical trimmer (TrimPro ROTOR,
https://www.trimpro.com, accessed on 20 March 2022). The trimmed flowers were re-weighed (flower fresh weight), and a subsample (approximately 50 g) was also weighed into a foil tray. The subsamples were dried in a dedicated drying room at 21 °C and 50% humidity until no further reduction in weight was observed (9 days). The subsamples were then re-weighed; the percent biomass and the total flower dry weight (g plant
−1) were calculated.
Root Area: After the plants were harvested, four replicates (one from each bench) from each control/graft were randomly selected. They were gathered into their replicate sets and turned out of their pots, upside down, on a tarpaulin. Photographs were taken where the roots had gathered in the pot base The photos were edited in Photoshop (Photoshop 22.5.1. Adobe ©, San Jose, CA, USA) to remove any perlite pieces. They were then opened in image analysis software, Image J [
31,
32], and a scale was defined from a plant tag of defined length that was included in each photograph. The image was converted to 8-bit black and white images, and then, the threshold was adjusted to a red-black image. A circular area that covered the bottom of the pot was drawn for each replicate, and the area of roots in mm was calculated in Image J [
31,
32].
2.4. Cannabinoid Quantification
Four biological replicates (one from each bench) from the four graft types (THC8r_CBD1s, THC9r_CBD1s, THC8r_2s, and THC9r_2s) and their respective non-grafted varieties, CBD1 and THC2, were analysed for the concentration of 17 cannabinoids according to the method described by Hewavitharana et al. [
33].
Three florets were randomly removed from the dried subsample flower material from each individual plant. From this, a further 0.1 g subsample was ground to a fine powder. Cannabinoids were extracted in 100% ethanol using sonication (SONICLEAN, Soniclean®, Dudley Park, SA, Australia) at 50/60 Hz. The ethanolic extract was analysed using High Performance Liquid Chromatography coupled to mass spectrometry (HPLC-MS, Agilent Technologies, Santa Clara, CA, USA). Separation was achieved using a reversed phase column (Agilent Poroshell, Agilent Technologies, Santa Clara, CA, USA) and methanol-water-acetonitrile mobile phase containing 0.1% formic acid (v/v). Cannabinoid peaks were identified according to their molar masses and quantified by calibration against known concentrations of commercially available cannabinoid standards. The method was validated in terms of detection and quantification limits, repeatability, and recoveries for all 17 cannabinoids before being used for quantification of cannabinoids in this study.
The total yield (mg plant
−1) of each cannabinoid was calculated as
All graphics and statistical analyses were performed in R 3.1 [
34].
4. Discussion
We have developed a grafting methodology for medicinal Cannabis that requires only a single cultivation step to connect a freshly cut donor scion to a freshly cut stem that will become the rootstock over only fourteen days. The method does not require any additional days of cultivation away from the usual propagation schedule and so does not reduce yield by increasing cultivation time. However, extra time is required to make the grafts compared to standard cloning. We estimate that grafting is three-fold slower than standard cloning. Another consideration is that mother plants for both the rootstock and scion would need to be maintained which increases labour demands and spacing requirements within the cultivation facility.
However, the potential benefits of grafting in medicinal Cannabis may extend beyond immediate yield advantages, as this method has been used in other crops to identify hormones, proteins, and RNAs that result in long-distance communication systems within the plant [
18,
19].
A prominent example are strigolactones, root-derived, actively transported [
35] phytohormones with profound effects on shoot architecture [
36]. While having multiple belowground and aboveground effects, including plant microbe interactions, strigolactones are best characterized for their negative regulation of lateral branching in response to nutrient availability, which is achieved in a complex interplay with other phytohormones [
37]. Strigolactones remained long elusive and were largely discovered and characterized through grafting experiments with biosynthesis and signal perception mutants in petunia (DAD mutants—decreased apical dominance], pea (RMS mutants—ramosus), and Arabidopis (MAX mutants– more axillary branching) [
38]. Strigolactone biosynthesis mutants showed a strong increased branching phenotype, which could be rescued by grafting mutant scions on wild type rootstock, which would then supply the signal. Contrastingly, strigolactone perception mutants could not be rescued via grafting.
It is likely that the branching and stunting phenotypes observed in our grafting experiments could at least in part be explained by differences between scion and rootstock in respect to strigolactone biosynthesis, perception, and or interaction with other phytohormones.
As a new crop to Australia, and one that produces medical products regulated by the Therapeutic Goods Act, there are currently no chemicals registered for use for the control of pests and diseases in medicinal Cannabis. Growers are therefore very vulnerable to outbreaks. Grafting may also be used as a scientific tool to identify novel methods of crop protection.
The main use of grafting in annual horticultural crops is for disease resistance to soil borne pathogens such as fusarium [
39] and verticillium [
40]. However, (currently) the main threats to medicinal Cannabis production are from foliar and flower pathogens, specifically botrytis (bud rot) and Golovinomyces Sp. (causing powdery mildew) [
41]. Long-distance transport/signalling mechanisms exist between roots and the aerial plant part, and these could provide a mechanism for inducing adaptations in the scion. For example, it has been demonstrated that grafting a powdery mildew (PM) susceptible scion on to a PM resistant rootstock reduced foliar infection in cucumber [
42]. The mechanism by which this occurs may have been via long-distance transport or through alteration of the metabolic profile of the scion [
43]. Differences in the expression of defence related transcripts or the proteome have also been observed in response to grafting in “Gala” apples and cucumber, respectively [
43,
44], and grafting may also promote induced systemic resistance by altering rhizobacterial populations [
45].
These examples raise the possibility that grafting could induce adaptive or resistance traits to foliar/floral pathogens into medicinal Cannabis directly or be a means to discover new pathways that can be manipulated to increase plant defences without chemicals.
The capacity of medicinal Cannabis to produce viable roots from defoliated, grafted, stem sections, with a high rate of success (>90% for THC9 rootstock grafts), in just 14 days, is remarkable. Indeed, we have observed that the scion and rootstock stem fuse within 72 h (data not shown). Willow (Salix Sp) also shows high regenerative rates and “willow water” extracted from the bark is used to promote root formation in other species, the active metabolite being salicylic acid [
46]. Characterisation of the cannabinoids and terpenes of Cannabis has been the subject of many publications [
3,
6,
47,
48,
49,
50], but the characterization and quantification of hormones has not; the underlying mechanism(s) behind this pronounced regenerative capacity remain to be discovered.
The presented method was capable of reducing height and changing stem morphology. Yields were also increased through increased flower biomass in one of our graft combinations. The outcomes from the two rootstocks used in our flowering trial were scion specific; one rootstock reduced plant height in one of the target scions but not the other, and another rootstock increased yield in one scion but not the other. The specificity of the rootstock–scion relationship has been previously reported in multiple crops [
17]. The poorer survival rate of clones grafted to the THC8 rootstock also shows that, as with other crops, all rootstocks are not equal, and experimentation is required to identify optimal combinations.
The rationale for using THC2 as a scion was that it showed variable root development during cloning which resulted in an unacceptably high failure rate (for commercial production). By grafting THC2 to THC9, this problem was ameliorated. However, the improvement that was observed in this graft combination cannot be explained by improved root area alone because the root area of THC8r_2s was also larger than non-grafted THC2, but there was no biomass yield advantage. Furthermore, the THC9r_CBD1s grafts had a rootstock less than half the area of CBD1 but showed no loss of yield, and the THC2 plants had the smallest root area of all lines but still produced exceptionally high yields. These findings show that a large root area is not necessary to produce high yields (indeed it can indicate the opposite). Why THC2 is impaired in root development has not been uncovered by this study but is not the result of inadequate signalling or transport/partitioning of carbon from the source organs, as both rootstocks grafted to this scion produced a root area that exceeded that of the non-grafted controls.
Overall, few changes in the concentration of cannabinoids were observed in the grafted scions. The main difference was that the concentration of CBDA was increased in one of the CBD1 grafted lines; however, as the biomass yield of this line was lower, the final yield of CBDA was not improved. A negative relationship between CBDA and biomass yield has been previously reported [
51]. In this cited example, the treatment applied (P fertilization) increased biomass yield but decreased CBDA concentration which the authors attributed to a diluting effect [
51]. Our observation may be the opposite response; a concentrating effect on CBDA from reduced biomass.
Cannabinol (CBN) was found in significantly higher quantities in one CBD1 graft type. This cannabinoid is a breakdown product from the degradation of THC, which is reportedly linked to aged, stored Cannabis [
52]. Both graft combinations of THC2 produced significantly higher concentrations of CBLA, which is again considered to be a breakdown product formed during storage or through exposure to environmental conditions. As all of our samples were stored and processed together, it is unclear why there should be increased concentrations of these cannabinoids. It is possible that the grafting process may have accelerated senescence, so the flowers were more mature at harvest. However, no differences in the % dry weight of the flowers or visual symptoms of accelerated maturity were observed.
The minimum number of changes induced by grafting suggests that the root has little influence on the cannabinoid concentrations, but through its effect on flower biomass, the grafting method presented can increase yields of THCA.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}