
Genome-Wide Identification, Characterization, and Expression Analysis of TUBBY Gene Family in Wheat (Triticum aestivum L.) under Biotic and Abiotic Stresses

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Wheat TUBBY Family Genes Identification
2.2. Phylogenetic Tree, Digital Expression, and Motif Analysis
2.3. Chromosomal Location and Protein-Protein Interaction of TUBBY Genes
2.4. Gene Structure and Conserved Motif Analysis
2.5. Gene Ontology and Cis-Elements Analysis of TUBBY Family Genes
2.6. RNA Isolation and cDNA Synthesis
2.7. Expression Analysis of Different Genes
3. Results
3.1. Identification and Analysis of TaTLPs Genes
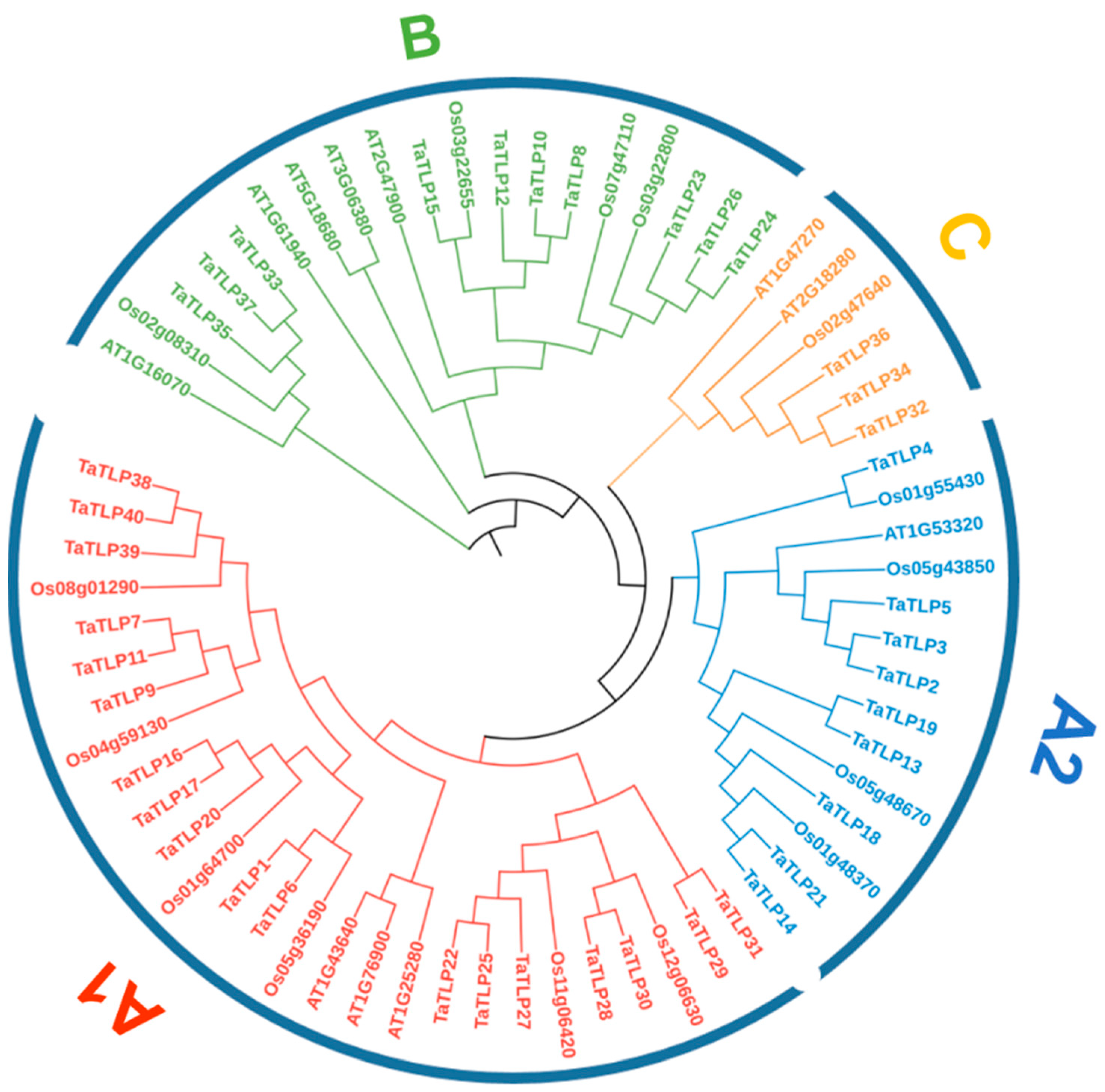
3.2. Phylogenetic Analysis of TaTLPs
3.3. Conserved Motif Analysis of TaTLPs-Genes
3.4. Gene Ontology of TaTLP Genes
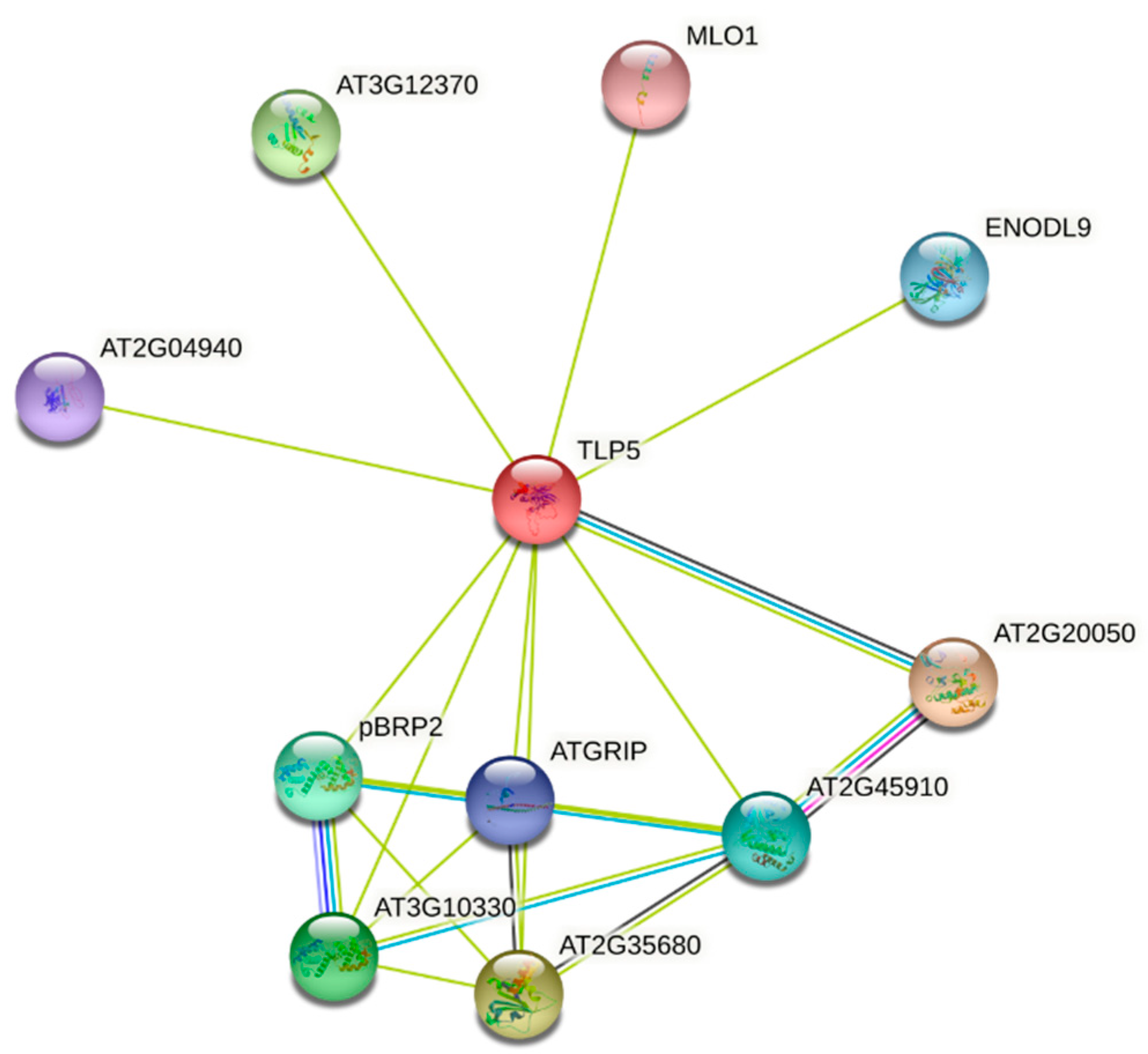
3.5. Protein-Protein Interaction of TaTLPs
3.6. TaTLPs Cis-Regulatory Elements
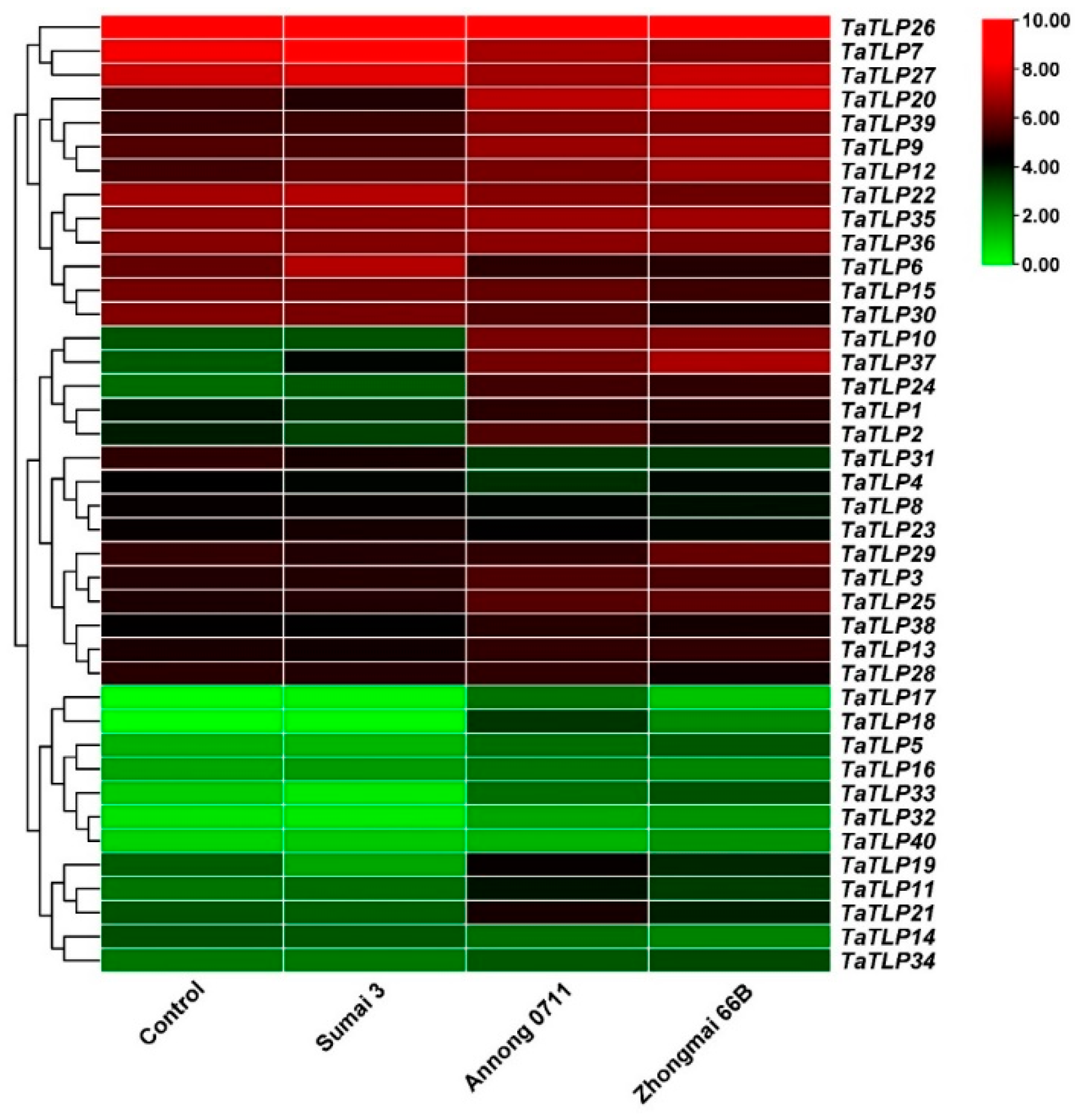
3.7. Expression Analysis of TaTLP Genes in Different Wheat Cultivars in Response to Fusarium graminum Stress
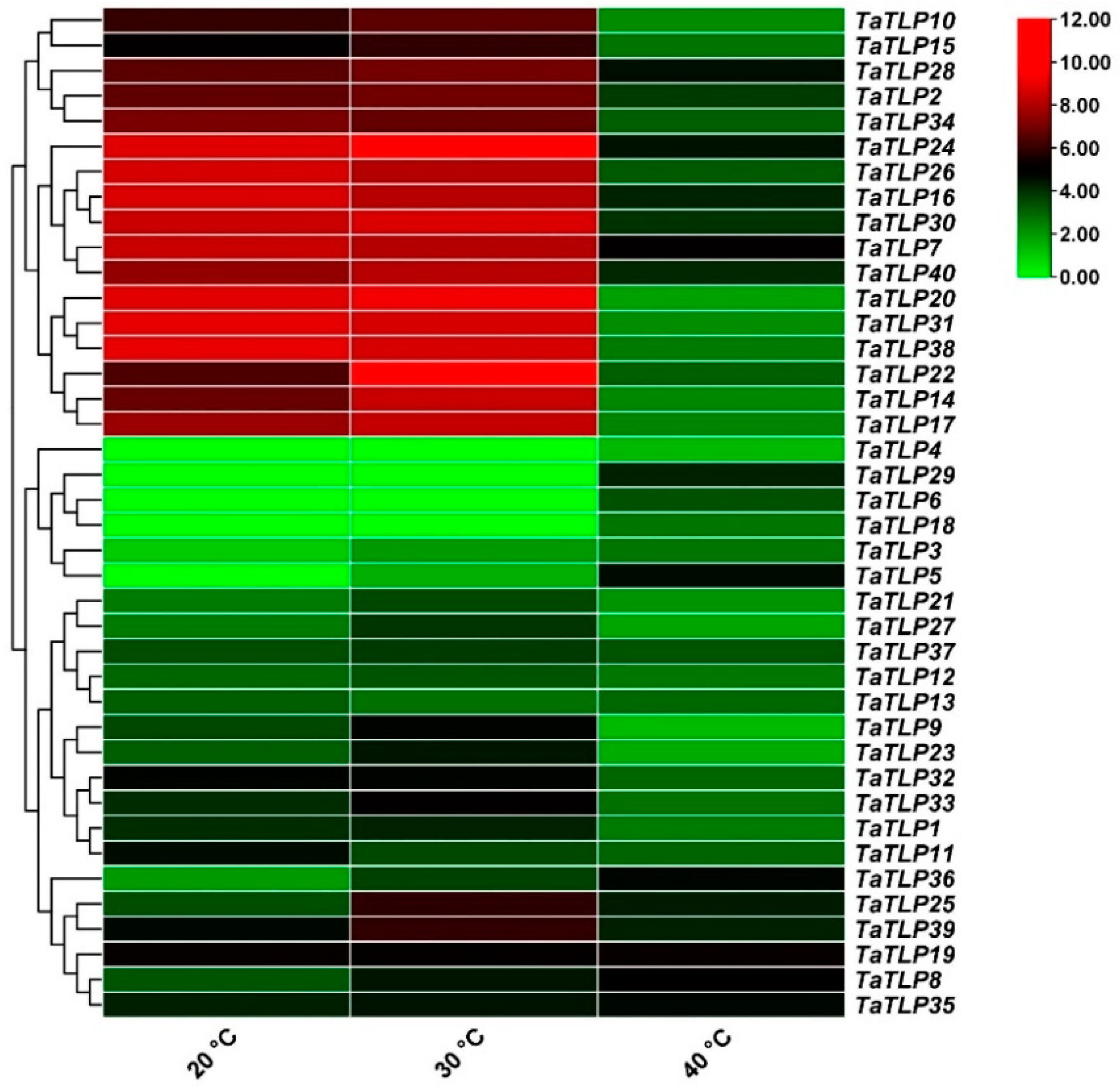
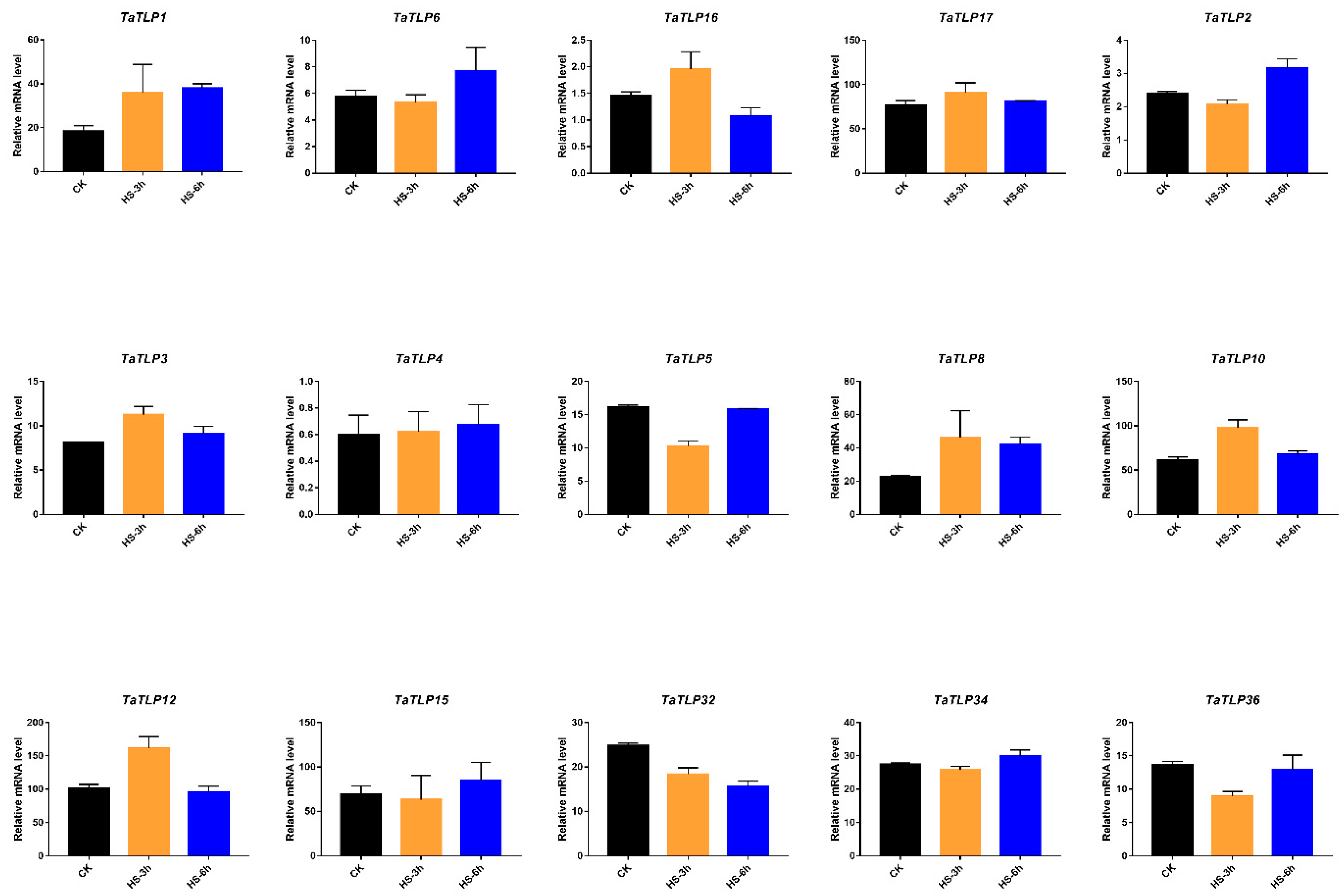
3.8. Expression Analysis of TaTLP Genes in Wheat in Response to Different Temperatures
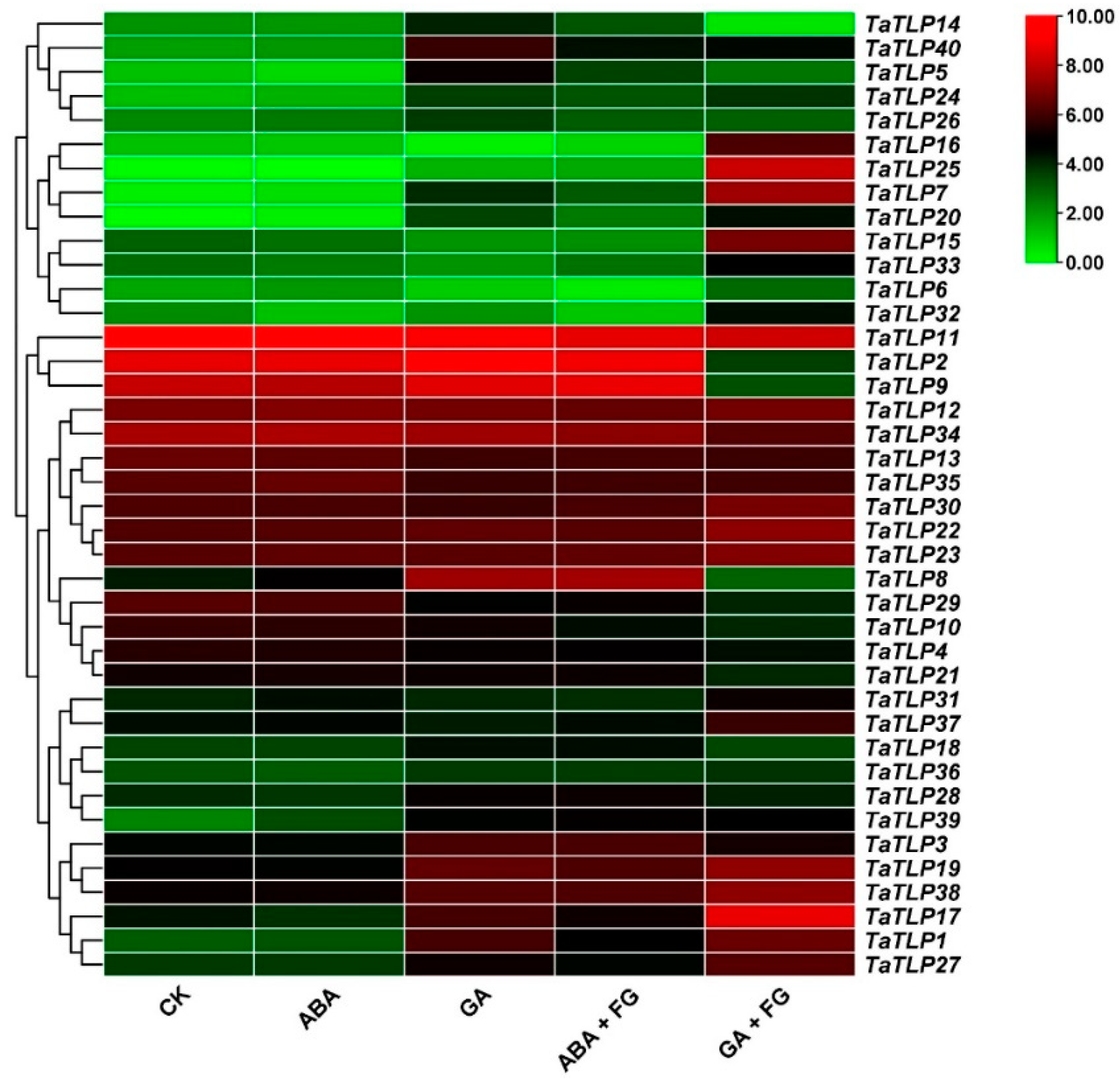
3.9. Expression Analysis of TaTLP Genes in Wheat Subjected to Hormonal Treatment
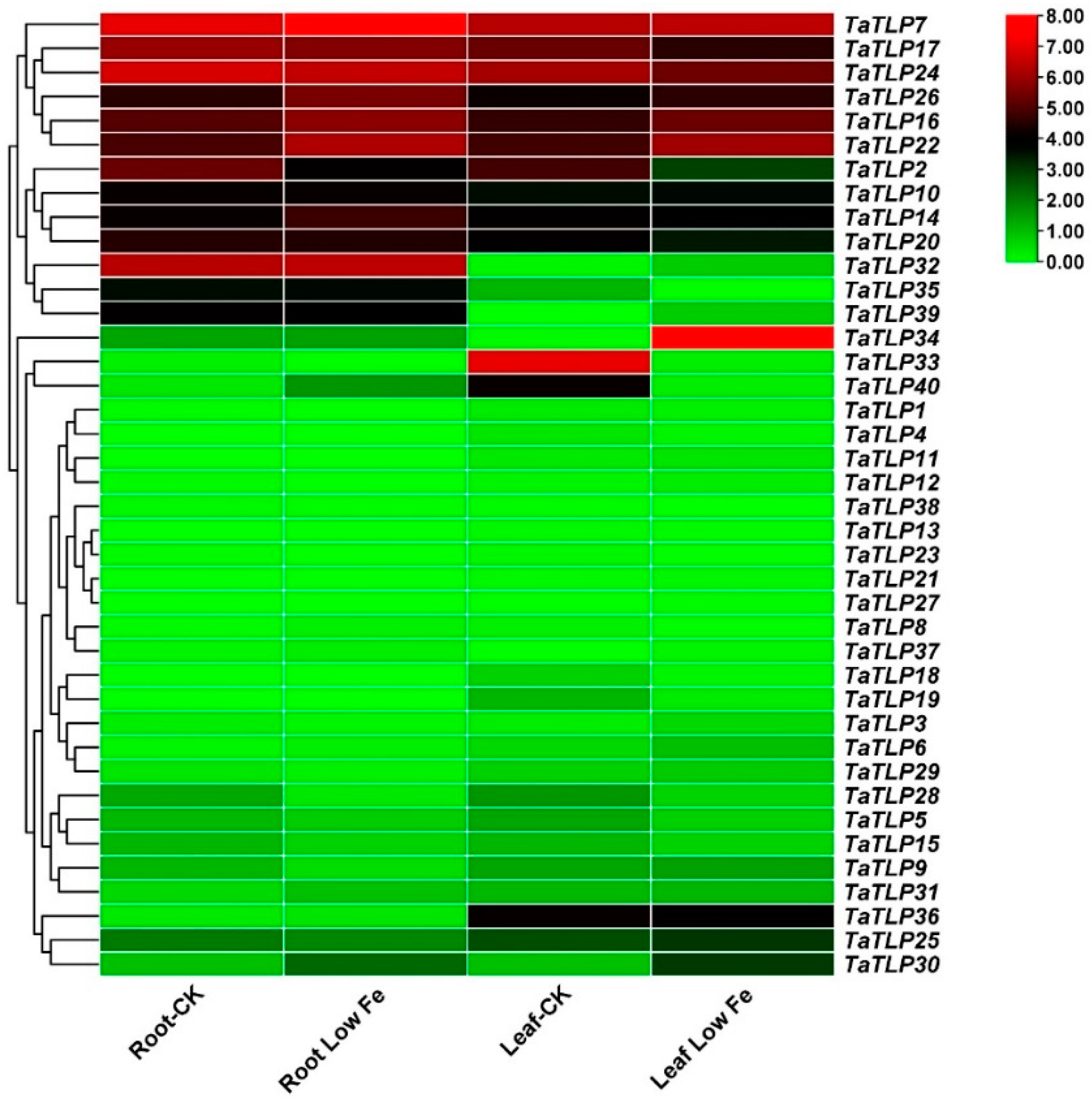
3.10. Expression Analysis of TaTLP Genes in Wheat in Response to Iron Deficiency Stress
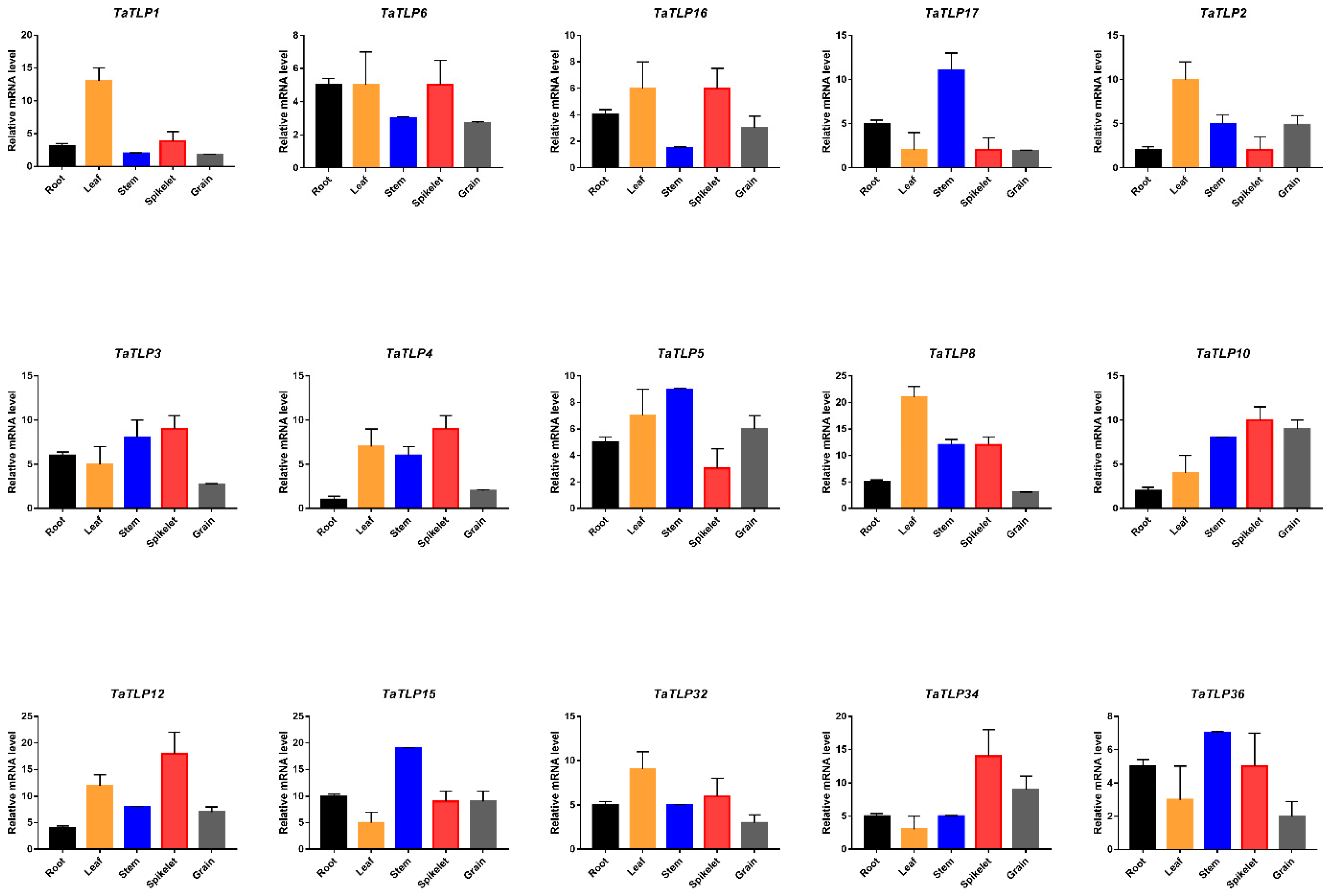
3.11. Expression Analysis of TaTLP Genes in Different Wheat Tissues
4. Discussion
4.1. TaTLP Genes Are Distributed Widely in the Wheat Genome
4.2. TaTLP Genes Are Thought to Be Involved in Critical Biological and Molecular Processes
4.3. TaTLP Genes Control Plant Response to Hormones and Abiotic and Biotic Stresses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carmo-Silva, A.E.; Gore, M.A.; Andrade-Sanchez, P.; French, A.N.; Hunsaker, D.J.; Salvucci, M.E. Decreased CO2 availability and inactivation of Rubisco limit photosynthesis in cotton plants under heat and drought stress in the field. Environ. Exp. Bot. 2012, 83, 1–11. [Google Scholar] [CrossRef]
- Sharif, R.; Xie, C.; Wang, J.; Cao, Z.; Zhang, H.; Chen, P.; Li, Y. Genome-wide identification, characterization and expression analysis of HD-ZIP gene family in Cucumis sativus L. under biotic and various abiotic stresses. Int. J. Biol. Macromol. 2020, 158, 502–520. [Google Scholar] [CrossRef] [PubMed]
- Altaf, A.; Zhu, X.; Zhu, M.; Quan, M.; Irshad, S.; Xu, D.; Aleem, M.; Zhang, X.; Gull, S.; Li, F. Effects of Environmental Stresses (Heat, Salt, Waterlogging) on Grain Yield and Associated Traits of Wheat under Application of Sulfur-Coated Urea. Agronomy 2021, 11, 2340. [Google Scholar] [CrossRef]
- Altaf, A.; Gull, S.; Zhu, X.; Zhu, M.; Rasool, G.; Ibrahim, M.E.H.; Aleem, M.; Uddin, S.; Saeed, A.; Shah, A.Z. Study of the effect of peg-6000 imposed drought stress on wheat (Triticum aestivum L.) cultivars using relative water content (RWC) and proline content analysis. Pak. J. Agric. Sci. 2021, 58, 357–367. [Google Scholar]
- Boggon, T.J.; Shan, W.S.; Santagata, S.; Myers, S.C.; Shapiro, L. Implication of tubby proteins as transcription factors by structure-based functional analysis. Science 1999, 286, 2119–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, K.; Gomez, C.; Shapiro, L. Tubby proteins: The plot thickens. Nat. Rev. Mol. Cell Biol. 2004, 5, 55–64. [Google Scholar] [CrossRef]
- Liu, Q. Identification of rice TUBBY-like genes and their evolution. FEBS J. 2008, 275, 163–171. [Google Scholar] [CrossRef]
- Xu, J.-N.; Xing, S.-S.; Zhang, Z.-R.; Chen, X.-S.; Wang, X.-Y. Genome-wide identification and expression analysis of the tubby-like protein family in the Malus domestica genome. Front. Plant Sci. 2016, 7, 1693. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhou, Y.; Wang, X.; Gu, S.; Yu, J.; Liang, G.; Yan, C.; Xu, C. Genomewide comparative phylogenetic and molecular evolutionary analysis of tubby-like protein family in Arabidopsis, rice, and poplar. Genomics 2008, 92, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Yulong, C.; Wei, D.; Baoming, S.; Yang, Z.; Qing, M. Genome-wide identification and comparative analysis of the TUBBY-like protein gene family in maize. Genes Genom. 2016, 38, 25–36. [Google Scholar] [CrossRef]
- Zhang, Y.; He, X.; Su, D.; Feng, Y.; Zhao, H.; Deng, H.; Liu, M. Comprehensive profiling of tubby-like protein expression uncovers ripening-related TLP genes in tomato (Solanum lycopersicum). Int. J. Mol. Sci. 2020, 21, 1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, N.; Fakhrah, S.; Mohanty, C.S.; Bag, S.K. Genome-Wide Identification and Evolutionary Analysis of Gossypium Tubby-Like Protein (TLP) Gene Family and Expression Analyses During Salt and Drought Stress. Front. Plant Sci. 2021, 12, 667929. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-P.; Lee, C.-L.; Chen, P.-H.; Wu, S.-H.; Yang, C.-C.; Shaw, J.-F. Molecular analyses of the Arabidopsis TUBBY-like protein gene family. Plant Physiol. 2004, 134, 1586–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, Y.; Song, W.-M.; Jin, Y.-L.; Jiang, C.-M.; Yang, Y.; Li, B.; Huang, W.-J.; Liu, H.; Zhang, H.-X. Characterization of Arabidopsis Tubby-like proteins and redundant function of AtTLP3 and AtTLP9 in plant response to ABA and osmotic stress. Plant Mol. Biol. 2014, 86, 471–483. [Google Scholar] [CrossRef]
- Wardhan, V.; Jahan, K.; Gupta, S.; Chennareddy, S.; Datta, A.; Chakraborty, S.; Chakraborty, N. Overexpression of CaTLP1, a putative transcription factor in chickpea (Cicer arietinum L.), promotes stress tolerance. Plant Mol. Biol. 2012, 79, 479–493. [Google Scholar] [CrossRef]
- Rad, R.N.; Kadir, M.A.; Jaafar, H.Z.; Gement, D. Physiological and biochemical relationship under drought stress in wheat (Triticum aestivum). Afr. J. Biotechnol. 2012, 11, 1574–1578. [Google Scholar]
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M. The Arabidopsis Information Resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, D1202–D1210. [Google Scholar] [CrossRef]
- Lukaszewski, A.J.; Alberti, A.; Sharpe, A.; Kilian, A.; Stanca, A.M.; Keller, B.; Clavijo, B.J.; Friebe, B.; Gill, B.; Wulff, B. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Yang, Y.; Luo, W.; Yang, C.; Ding, P.; Liu, Y.; Qiao, L.; Chang, Z.; Geng, H.; Wang, P. Genome-wide identification and analysis of the MADS-box gene family in bread wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0181443. [Google Scholar] [CrossRef] [Green Version]
- Borrill, P.; Ramirez-Gonzalez, R.; Uauy, C. expVIP: A customizable RNA-seq data analysis and visualization platform. Plant Physiol. 2016, 170, 2172–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, A.-Y.; Zhu, Q.-H.; Chen, X.; Luo, J.-C. GSDS: A gene structure display server. Yi Chuan = Hered. 2007, 29, 1023–1026. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Levy, A.A. Genome evolution due to allopolyploidization in wheat. Genetics 2012, 192, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, Z.; Kong, Y. The tubby-like proteins kingdom in animals and plants. Gene 2018, 642, 16–25. [Google Scholar] [CrossRef]
- Xu, J.; Xing, S.; Sun, Q.; Zhan, C.; Liu, X.; Zhang, S.; Wang, X. The expression of a tubby-like protein from Malus domestica (MdTLP7) enhances abiotic stress tolerance in Arabidopsis. BMC Plant Biol. 2019, 19, 60. [Google Scholar] [CrossRef] [Green Version]
- Gagne, J.M.; Downes, B.P.; Shiu, S.-H.; Durski, A.M.; Vierstra, R.D. The F-box subunit of the SCF E3 complex is encoded by a diverse superfamily of genes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2002, 99, 11519–11524. [Google Scholar] [CrossRef] [Green Version]
- Endler, A.; Kesten, C.; Schneider, R.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A mechanism for sustained cellulose synthesis during salt stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef] [Green Version]
- Durrant, W.E.; Rowland, O.; Piedras, P.; Hammond-Kosack, K.E.; Jones, J.D. cDNA-AFLP reveals a striking overlap in race-specific resistance and wound response gene expression profiles. Plant Cell 2000, 12, 963–977. [Google Scholar] [CrossRef]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Provart, N.J.; Glazebrook, J.; Katagiri, F.; Chang, H.-S.; Eulgem, T.; Mauch, F.; Luan, S.; Zou, G.; Whitham, S.A. Expression profile matrix of Arabidopsis transcription factor genes suggests their putative functions in response to environmental stresses. Plant Cell 2002, 14, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Karam, B.; Rhonda, C.; Luis, O. Transcription factors in plant defense and stress response. Curr Opin Plant. Biol 2002, 5, 430–436. [Google Scholar]
- He, Y.; Li, W.; Lv, J.; Jia, Y.; Wang, M.; Xia, G. Ectopic expression of a wheat MYB transcription factor gene, TaMYB73, improves salinity stress tolerance in Arabidopsis thaliana. J. Exp. Bot. 2012, 63, 1511–1522. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cui, X.; Meng, Z.; Huang, X.; Xie, Q.; Wu, H.; Jin, H.; Zhang, D.; Liang, W. Transcriptional regulation of Arabidopsis MIR168a and argonaute1 homeostasis in abscisic acid and abiotic stress responses. Plant. Physiol. 2012, 158, 1279–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Waterlogging response mechanisms in plants: Research progress and prospects. Front. Plant Sci. 2020, 11, 2319. [Google Scholar]
- Raza, A.; Tabassum, J.; Kudapa, H.; Varshney, R.K. Can omics deliver temperature resilient ready-to-grow crops? Crit. Rev. Biotechnol. 2021, 41, 1209–1232. [Google Scholar] [CrossRef]
- Gupta, R.; Chakrabarty, S. Gibberellic acid in plant: Still a mystery unresolved. Plant. Signal. Behav. 2013, 8, e25504. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P. The current status of research on gibberellin biosynthesis. Plant. Cell Physiol. 2020, 61, 1832–1849. [Google Scholar] [CrossRef]
- Yu, C.-S.; Cheng, C.-W.; Su, W.C.; Chang, K.-C.; Huang, S.-W.; Hwang, J.-K.; Lu, C.-H. CELLO2GO: A web server for protein subCELlular LOcalization prediction with functional gene ontology annotation. PLoS ONE 2014, 9, e99368. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus ID | Proteins | MW | PI | SL |
|---|---|---|---|---|---|
| TaTLP1 | Traes_1AL_399C1DBF5 | 269 | - | - | N |
| TaTLP2 | Traes_1AL_45AB9EF34 | 247 | 27,640.24 | 9.22 | EC |
| TaTLP3 | Traes_1BL_BDF2FCEC7 | 175 | 19,434.98 | 9.08 | N |
| TaTLP4 | Traes_1DL_106460E68 | 50 | 5544.55 | 9.42 | EC |
| TaTLP5 | Traes_1DL_9DE76F004 | 251 | 27,986.78 | 9.16 | EC |
| TaTLP6 | Traes_1DL_FAB396374 | 204 | 22,671.21 | 9.80 | N |
| TaTLP7 | Traes_2AL_436D234EE | 471 | 52,097.90 | 9.28 | N |
| TaTLP8 | Traes_2AS_11876C298 | 203 | 22,666.81 | 9.14 | N |
| TaTLP9 | Traes_2BL_181C4AA28 | 472 | 52,073.74 | 9.22 | N |
| TaTLP10 | Traes_2BS_9DCC9CC7A | 203 | 22,657.80 | 9.14 | N |
| TaTLP11 | Traes_2DL_79449BF6D | 347 | 38,850.30 | 9.73 | N |
| TaTLP12 | Traes_2DS_AC89AEF36 | 203 | 22,661.79 | 9.14 | N |
| TaTLP13 | Traes_3AL_108550E28 | 126 | 14,455.70 | 8.34 | EC |
| TaTLP14 | Traes_3AL_731AE8008 | 60 | 6858.86 | 10.08 | M |
| TaTLP15 | Traes_3AL_9B4AF9950 | 152 | 17,297.97 | 9.37 | N |
| TaTLP16 | Traes_3AL_C3AEDD333 | 82 | 9123.34 | 10.72 | EC |
| TaTLP17 | Traes_3B_021E89FE5 | 377 | - | - | N |
| TaTLP18 | Traes_3B_02CE045341 | 194 | 22,052.95 | 9.33 | N |
| TaTLP19 | Traes_3B_C967E97B9 | 180 | 20,367.29 | 9.30 | N |
| TaTLP20 | Traes_3DL_697C6F117 | 211 | 23,933.51 | 9.69 | N |
| TaTLP21 | Traes_3DL_C81B58D98 | 279 | 31,811.57 | 9.63 | EC |
| TaTLP22 | Traes_4AL_EDE236978 | 440 | 48,793.98 | 9.39 | N |
| TaTLP23 | Traes_4AS_0C8542099 | 402 | 44,497.04 | 9.43 | C |
| TaTLP24 | Traes_4BL_7E9BC637F | 559 | 61,782.42 | 9.73 | M |
| TaTLP25 | Traes_4BS_D5B5C14F6 | 440 | 48,817.02 | 9.39 | N |
| TaTLP26 | Traes_4DL_7D905B6BC | 404 | 44,726.34 | 9.29 | C |
| TaTLP27 | Traes_4DS_620432A0D | 440 | 48,833.02 | 9.39 | N |
| TaTLP28 | Traes_5AL_83533B97D | 361 | 40,470.04 | 9.54 | N |
| TaTLP29 | Traes_5AL_D38708404 | 64 | 7023.98 | 4.75 | N |
| TaTLP30 | Traes_5BL_ECCAFFEB4 | 440 | 49,017.86 | 9.34 | N |
| TaTLP31 | Traes_5DL_FA0200E13 | 439 | 48,856.72 | 9.25 | N |
| TaTLP32 | Traes_6AL_058D829C3 | 374 | - | - | N |
| TaTLP33 | Traes_6AS_FB1249AB4 | 321 | 35,747.87 | 9.70 | N |
| TaTLP34 | Traes_6BL_9F6ACF02D | 322 | 35,555.86 | 9.39 | N |
| TaTLP35 | Traes_6BS_5C281F303 | 177 | 20,314.47 | 9.98 | EC |
| TaTLP36 | Traes_6DL_E7A7DAE5C | 368 | 40,756.92 | 9.26 | N |
| TaTLP37 | Traes_6DS_D6AD8C3ED | 177 | 20,355.52 | 9.98 | N |
| TaTLP38 | Traes_7AL_52F3AE87C | 373 | 41,569.32 | 9.80 | N |
| TaTLP39 | Traes_7BL_8312EAB48 | 441 | - | - | N |
| TaTLP40 | Traes_7DL_037F2A4F4 | 238 | 26,443.41 | 9.69 | N |
| Gene Name | Protein Family | Putative Function | Interactive-Bit Score |
|---|---|---|---|
| ATG2G20050 | Protein phosphatase 2C and cyclic nucleotide-binding | Signal transduction, ATP binding, metal ion binding, protein serine phosphatase activity | 0.837 |
| ATG2G35680 | Phosphotyrosine protein phosphatase superfamily protein | Possess phosphate activity | 0.698 |
| AT3G12370 | EMB3136—Ribosomal protein L10 family protein | The function is a structural protein | 0.691 |
| AT3G10330 | Cyclin-like family protein | DNA-templated transcription, initiation, transcription preinitiation complex assembly | 0.637 |
| pBRP2 | Plant-specific TFIIB-related protein 2 | Regulation of endosperm proliferation, DNA-templated transcription, initiation | 0.637 |
| AT2G45910 | U-box domain-containing protein kinase family protein | Cellular response to oxygen-containing compound, defense response to the bacterium, flower development, | 0.633 |
| ENDOL9 | Early nodulin-like protein 9 | electron carrier activity | 0.623 |
| ATGRIP | Golgi-localized grip domain-containing protein | Involved in Golgi protein trafficking. AtARL1 binds directly to the GRIP domain of AtGRIP in a GTP-dependent manner. | 0.616 |
| AT2G04940 | scramblase-like protein | plasma membrane phospholipid scrambling | 0.603 |
| MLO1 | Transmembrane domain protein | barely mildew resistance | 0.588 |
| Category | Cis-Elements | Annotations |
|---|---|---|
| Hormone | ABRE | Cis-acting element involved in the abscisic acid responsiveness. |
| TCA | Cis-acting element involved in salicylic acid responsiveness. | |
| TATC-Box | Gibberellin-responsive element. | |
| AuxRR-Core | Auxin-responsive element. | |
| CGTCA | Cis-acting regulatory element involved in the MeJA-responsiveness. | |
| TGACG | Cis-acting regulatory element involved in the MeJA-responsiveness. | |
| Stress and Growth | ARE | Stimulate mRNA decay. |
| ACE | Cis-acting element involved in light responsiveness. | |
| G-Box | Cis-acting element involved in light responsiveness. | |
| LTR | Long-terminal repeat. | |
| CAT-Box | Cis-acting element involved in meristem development. | |
| O2-Site | Cis-acting regulatory element involved in zein metabolism regulation. | |
| MSA-Like | Cis-acting regulatory element involved in the cell cycle. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altaf, A.; Zada, A.; Hussain, S.; Gull, S.; Ding, Y.; Tao, R.; Zhu, M.; Zhu, X. Genome-Wide Identification, Characterization, and Expression Analysis of TUBBY Gene Family in Wheat (Triticum aestivum L.) under Biotic and Abiotic Stresses. Agronomy 2022, 12, 1121. https://doi.org/10.3390/agronomy12051121
Altaf A, Zada A, Hussain S, Gull S, Ding Y, Tao R, Zhu M, Zhu X. Genome-Wide Identification, Characterization, and Expression Analysis of TUBBY Gene Family in Wheat (Triticum aestivum L.) under Biotic and Abiotic Stresses. Agronomy. 2022; 12(5):1121. https://doi.org/10.3390/agronomy12051121
Chicago/Turabian StyleAltaf, Adil, Ahmad Zada, Shahid Hussain, Sadia Gull, Yonggang Ding, Rongrong Tao, Min Zhu, and Xinkai Zhu. 2022. "Genome-Wide Identification, Characterization, and Expression Analysis of TUBBY Gene Family in Wheat (Triticum aestivum L.) under Biotic and Abiotic Stresses" Agronomy 12, no. 5: 1121. https://doi.org/10.3390/agronomy12051121
APA StyleAltaf, A., Zada, A., Hussain, S., Gull, S., Ding, Y., Tao, R., Zhu, M., & Zhu, X. (2022). Genome-Wide Identification, Characterization, and Expression Analysis of TUBBY Gene Family in Wheat (Triticum aestivum L.) under Biotic and Abiotic Stresses. Agronomy, 12(5), 1121. https://doi.org/10.3390/agronomy12051121