
Physiological Fitness Associated to ACCase Target-Site Resistance Enhances Growth and Reproduction in Phalaris brachystachys

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Source and Genetic Background Control
2.2. Evaluation of Growth
2.3. Determination of Physiological Parameters of S and R Sub-Populations
2.4. Determination of Physiological Parameters of S and R Populations
3. Results
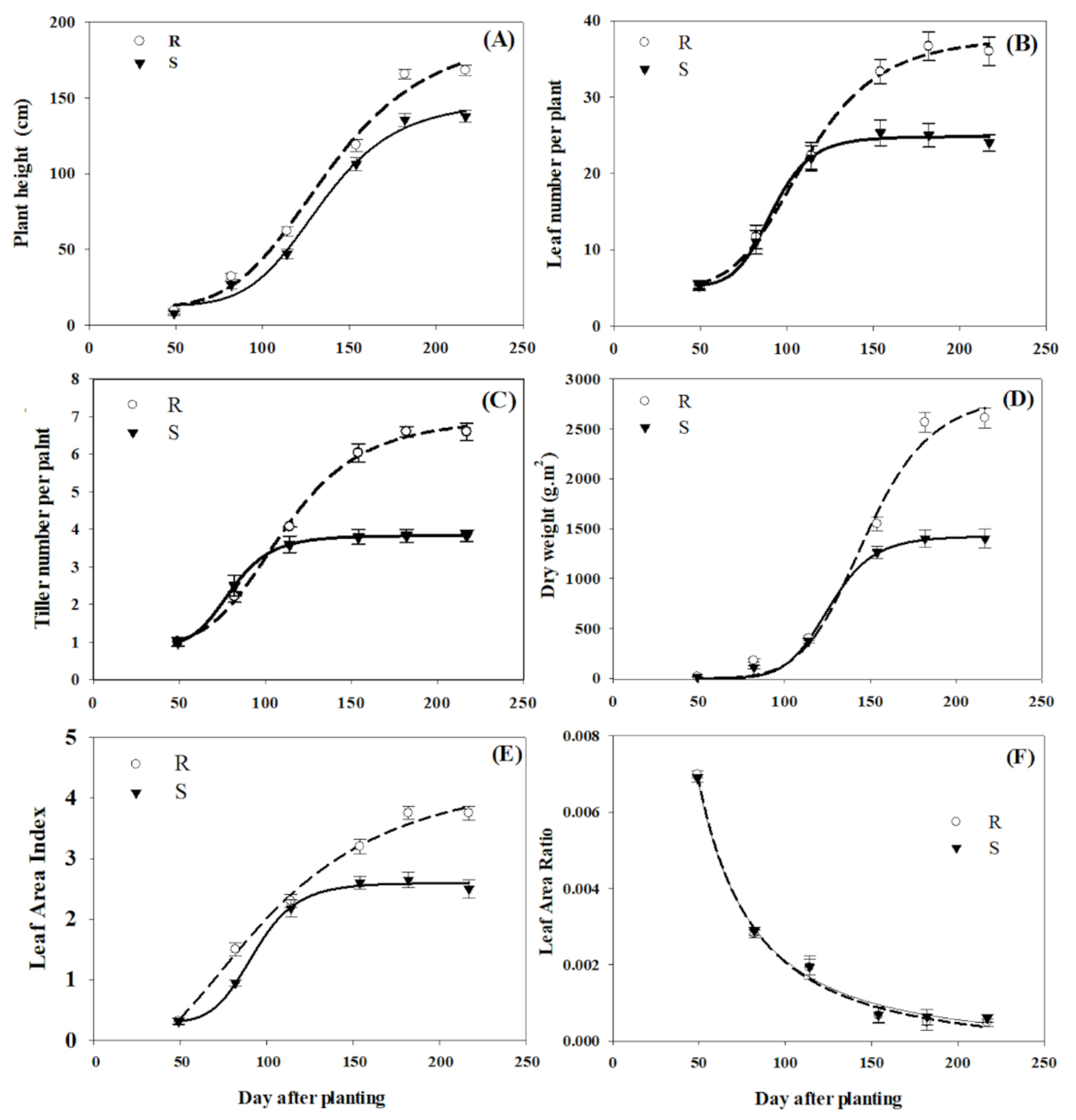
3.1. Comparative Growth Analysis
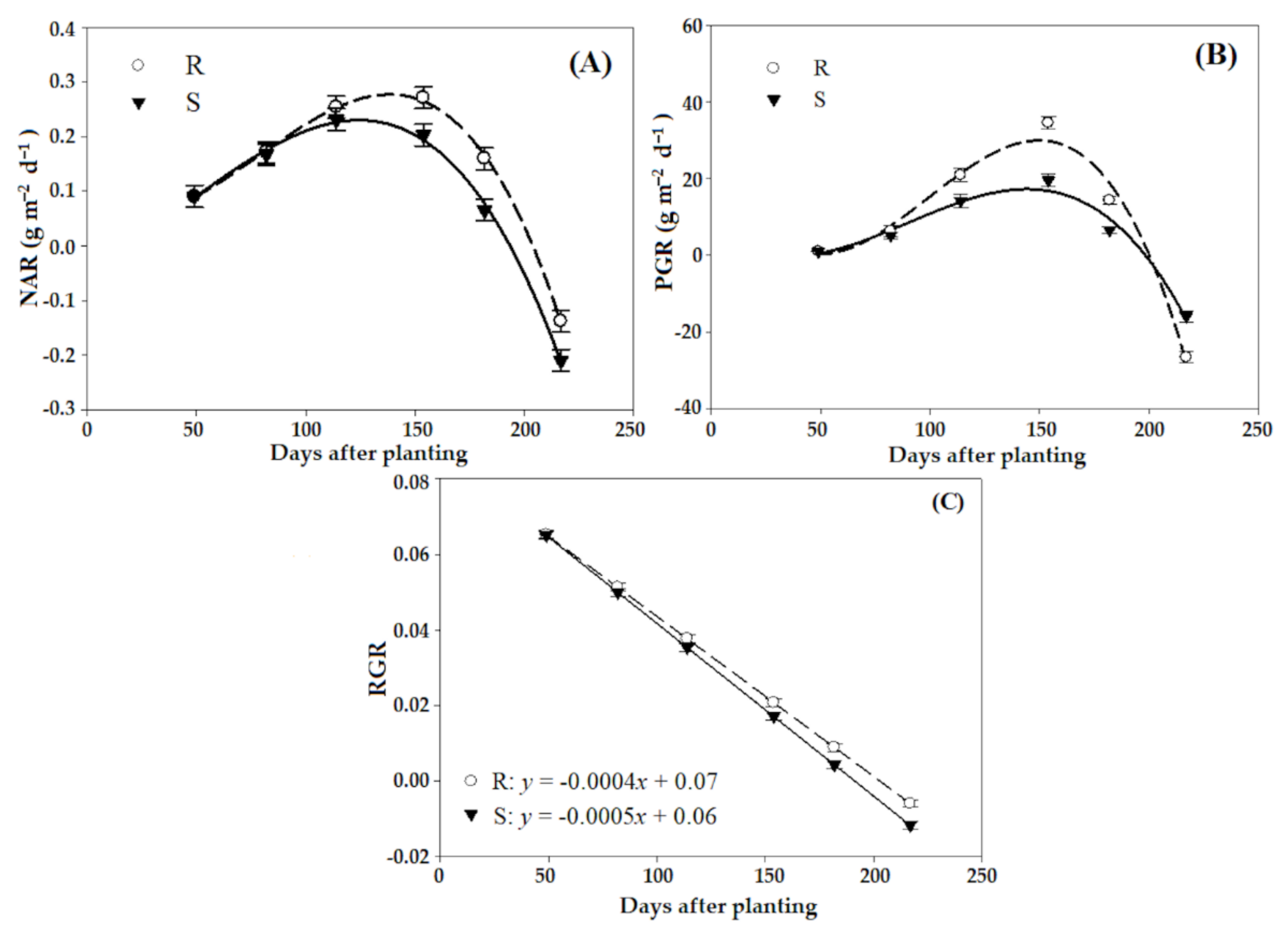
3.2. Physiological Parameters
3.3. Seed Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colbach, N.; Chauvel, B.; Darmency, H.; Délye, C.; Le Corre, V. Choosing the best cropping systems to target pleiotropic effects when managing single-gene herbicide resistance in grass weeds. A blackgrass simulation study. Pest Manag. Sci. 2016, 72, 1910–1925. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Marjanovic, J.; Gornicki, P. Resistance to herbicides caused by single amino acid mutations in acetyl-CoA carboxylase in resistant populations of grassy weeds. New Phytol. 2013, 197, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. International Survey of Herbicide Resistant Weeds. 2022. Available online: http://www.weedscience.org (accessed on 11 April 2022).
- Owen, M.J.; Michael, P.J.; Renton, M.; Steadman, K.J.; Powles, S.B. Towards large-scale prediction of Lolium rigidum emergence. II. Correlation between dormancy and herbicide resistance levels suggests an impact of cropping systems. Weed Res. 2011, 51, 133–141. [Google Scholar] [CrossRef]
- Panozzo, S.; Scarabel, L.; Rosan, V.; Sattin, M. A new Ala-122-Asn amino acid change confers decreased fitness to ALS-resistant Echinochloa Crus-Gall. Front. Plant Sci. 2017, 8, 2042. [Google Scholar] [CrossRef] [Green Version]
- Kaundun, S.S. Resistance to acetyl-CoA carboxylase-inhibiting herbicides. Pest Manag. Sci. 2014, 70, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Délye, C. Weed resistance to acetyl coenzyme A carboxylase inhibitors: An update. Weed Sci. 2005, 53, 728–746. [Google Scholar] [CrossRef]
- Busi, R.; Vila-Aiub, M.M.; Beckie, H.J.; Gaines, T.A.; Goggin, D.E.; Kaundun, S.S.; Lacoste, M.; Neve, P.; Nissen, S.J.; Norsworthy, J.K.; et al. Herbicide resistant weeds: From research and knowledge to future needs. Evol. Appl. 2013, 6, 1219–1221. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Picard, J.C.; Tian, X.; Darmency, H. A herbicide-resistant ACCase 1781 Setaria mutant shows higher fitness than wild type. Heredity 2010, 105, 394–400. [Google Scholar] [CrossRef]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Ann. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [Green Version]
- Neve, P.; Powles, S. Recurrent selection with reduced herbicide rates results in the rapid evolution of herbicide resistance in Lolium Rigidum. Theor. Appl. Genet. 2005, 110, 1154–1166. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, T.; Pan, L.; Wang, L.; Xu, H.; Dong, L. Germination requirements differ between fenoxaprop-P-ethyl resistant and susceptible Japanese foxtail (Alopecurus japonicus) biotypes. Weed Sci. 2016, 64, 653–663. [Google Scholar] [CrossRef]
- Frenkel, E.; Matzrafi, M.; Rubin, B.; Peleg, Z. Effects of environmental conditions on the fitness penalty in herbicide resistant Brachypodium Hybridum. Front. Plant Sci. 2017, 8, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Vila-Aiub, M.M.; Gundel, P.E.; Preston, C. Experimental methods for estimation of plant fitness costs associated with herbicide-resistance genes. Weed Sci. 2015, 63, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Kim, Y.; Tong, L. Mechanism for the inhibition of the carboxyltransferase domain of acetyl-coenzyme A carboxylase by pinoxaden. Proc. Natl. Acad. Sci. USA 2010, 107, 22072–22077. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Moreno, P.; Alcántara-de la Cruz, R.; Smeda, R.J.; De Prado, R. Differential resistance mechanisms to Glyphosate result in fitness cost for Lolium perenne and L. multiflorum. Front. Plant Sci. 2017, 8, 1796. [Google Scholar] [CrossRef] [Green Version]
- Vila-Aiub, M.M.; Neve, P.; Powles, S.B. Fitness costs associated with evolved herbicide resistance genes in plants. New Phytol. 2009, 184, 751–767. [Google Scholar] [CrossRef] [Green Version]
- Shergill, L.S.; Boutsalis, P.; Preston, C.; Gill, G.S. Fitness costs associated with 1781 and 2041 ACCase-mutant alleles conferring resistance to herbicides in Hordeum glaucum Steud. Crop Prot. 2016, 87, 60–67. [Google Scholar] [CrossRef]
- Travlos, I.S. Competition between ACCase-inhibitor resistant and susceptible sterile wild oat (Avena sterilis) biotypes. Weed Sci. 2013, 61, 26–31. [Google Scholar] [CrossRef]
- Keshtkar, E.; Mathiassen, S.K.; Kudsk, P. No vegetative and fecundity fitness cost associated with acetyl-coenzyme A carboxylase non-target-site resistance in a Black-grass (Alopecurus myosuroides Huds.) population. Front. Plant Sci. 2017, 8, 2011–2017. [Google Scholar] [CrossRef] [Green Version]
- Cousens, R.D.; Fournier-Level, A. Herbicide resistance costs: What are we actually measuring and why? Pest Manag. Sci. 2018, 74, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Darmency, H.; Menchari, Y.; Le Corre, V.; Délye, C. Fitness cost due to herbicide resistance may trigger genetic background evolution. Evolution 2015, 69, 271–278. [Google Scholar] [CrossRef]
- Li, M.; Yu, Q.; Han, H.; Vila-Aiub, M.; Powles, S.B. ALS herbicide resistance mutations in Raphanus raphanistrum: Evaluation of pleiotropic effects on vegetative growth and ALS activity. Pest Manag. Sci. 2013, 69, 689–695. [Google Scholar] [CrossRef]
- Kumar, V.; Jha, P. Differences in germination, growth, and fecundity characteristics of dicamba-fluroxypyr-resistant and susceptible Kochia Scoparia. PLoS ONE 2016, 11, e0161533. [Google Scholar] [CrossRef]
- Keshtkar, E.; Abdolshahi, R.; Sasanfar, H.; Zand, E.; Beffa, R.; Dayan, F. Assessing fitness costs from a herbicide-resistance management perspective: A review and insight. Weed Sci. 2019, 67, 137–148. [Google Scholar] [CrossRef]
- Gherekhloo, J.; Oveisi, M.; Zand, E.; De Prado, R. A review of herbicide resistance in Iran. Weed Sci. 2016, 64, 551–561. [Google Scholar] [CrossRef]
- Golmohammadzadeh, S.; Gherekhloo, J.; Rojano-Delgado, A.M.; Osuna-Ruíz, M.D.; Kamkar, B.; Ghaderi-Far, F.; De Prado, R. The first case of short-spiked canarygrass (Phalaris brachystachys) with cross-resistance to ACCase-inhibiting herbicides in Iran. Agronomy 2019, 9, 377. [Google Scholar] [CrossRef] [Green Version]
- Golmohammadzadeh, S.; Rojano-Delgado, A.M.; Vázquez-García, J.G.; Romano, Y.; Osuna, M.D.; Gherekhloo, J.; De Prado, R. Cross-resistance mechanisms to ACCase-inhibiting herbicides in short-spike canarygrass (Phalaris brachystachys). Plant Physiol. Biochem. 2020, 15, 681–688. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; Harington, K.C. Fitness costs associated with multiple resistance to dicamba and atrazine in Chenopodium Album. Planta 2019, 249, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Goudriaan, J.; Lantinga, E.A.; Vos, J.; Spiertz, H.J. A flexible sigmoid function of determinate growth. Ann. Bot. 2003, 91, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Simonyan, K.J.; Yiljep, Y.D.; Oyatoyan, O.B.; Bawa, G.S. Effect of moisture content on some physical properties of Lablab purpureus (L.) sweet seeds. Agric. Eng. Int. CIGR J. 2009, 5, 1279–1293. [Google Scholar]
- Niinemets, U. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The relationship between leaf area growth and biomass accumulation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Henckes, J.R.; Chechin, J.; Schemitz, M.F.; Piasecki, C.; Vargas, L.; Agostinetto, D. Fitness cost and competitive ability of ryegrass susceptible and with multiple resistance to glyphosate, iodosulfuron-methyl, and pyroxsulam. Planta Daninha 2019, 37, e019197532. [Google Scholar] [CrossRef]
- Cechin, J.; Vargas, L.; Agostinetto, D.; Zimmer, V.; Pertile, M.; Dal Magro, T. Fitness costs of susceptible and resistant radish biotypes to ALS-inhibitor herbicides. Comun. Sci. 2017, 8, 281–286. [Google Scholar] [CrossRef]
- Zhang, P.; Hefting, M.M.; Soons, M.B.; Kowalchuk, G.A.; Rees, M.; Hector, A.; Turnbull, L.A.; Zhou, X.; Guo, Z.; Chu, C.; et al. Fast and furious: Early differences in growth rate drive short-term plant dominance and exclusion under eutrophication. Ecol. Evol. 2020, 10, 10116–10129. [Google Scholar] [CrossRef]
- Lowry, C.J.; Smith, R.G. Weed control through crop plant manipulations. In Non-Chemical Weed Control, 1st ed.; Jabran, K., Chauhan, B.S., Eds.; Academic Press: London, UK, 2018; pp. 73–96. [Google Scholar]
- Koelewijn, H.P. Rapid change in relative growth rate between the vegetative and reproductive stage of the life cycle in Plantago coronopus. New Phytol. 2004, 163, 67–76. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Neve, P.; Steadman, K.J.; Powles, S.B. Ecological fitness of a multiple herbicide-resistant Lolium rigidum population: Dynamics of seed germination and seedling emergence of resistant and susceptible phenotypes. J. Appl. Ecol. 2005, 42, 288–298. [Google Scholar] [CrossRef]
- Sabet Zangeneh, H.; Mohammadust Chamanabad, H.R.; Zand, E.; Asghari, A.; Alamisaeid, K.; Travlos, I.S.; Alebrahim, M.T. Study of fitness cost in three rigid ryegrass populations susceptible and resistant to Acetyl-CoA Carboxylase Inhibiting herbicides. Front. Ecol. Evol. 2016, 4, 142. [Google Scholar] [CrossRef] [Green Version]
- Park, K.W.; Mallory-Smith, C.A.; Ball, D.A.; Mueller-Warrant, G.W. Ecological fitness of acetolactate synthase inhibitor-resistant and -susceptible downy brome (Bromus tectorum) biotypes. Weed Sci. 2004, 52, 768–773. [Google Scholar] [CrossRef]
- Menchari, Y.; Chauvel, B.; Darmency, H.; D’elye, C. Fitness costs associated with three mutant acetylcoenzyme a carboxylase alleles endowing herbicide resistance in black-grass Alopecurus myosuroides. J. Appl. Ecol. 2008, 45, 939–947. [Google Scholar] [CrossRef]
- Matzarfi, M.; Gerson, O.; Rubin, B.; Peleg, Z. Different mutations endowing resistance to Acetyl-CoA Carboxylase inhibitors results in changes in ecological fitness of Lolium rigidum populations. Front. Plant Sci. 2017, 8, 1078–1083. [Google Scholar] [CrossRef] [Green Version]
- Vila-Aiub, M.M.; Yu, Q.; Han, H.; Powles, S.B. Effect of herbicide resistance endowing Ile-1781-Leu and Asp-2078-Gly ACCase gene mutations on ACCase kinetics and growth traits in Lolium rigidum. J. Exp. Bot. 2015, 66, 4711–4718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassanpour-Bourkheili, S.; Gherekhloo, J.; Kamkar, B.; Ramezanpour, S.S. Comparing fitness cost associated with haloxyfop-R-methyl ester resistance in winter wild oat biotypes. Planta Daninha 2020, 38, e020213759. [Google Scholar] [CrossRef]
- Du, L.; Qu, M.; Jiang, X.; Li, X.; Ju, Q.; Lu, X.; Wang, J. Fitness costs associated with acetyl-coenzyme A carboxylase mutations endowing herbicide resistance in American sloughgrass (Beckmannia syzigachne Steud.). Ecol. Evol. 2019, 9, 2220–2230. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Traits | Population | Parameters 1 | R2 | p-Value * | ||
|---|---|---|---|---|---|---|
| Amax | b | t50 | ||||
| Plant height | R | 181.4 ± 6.5 a | 4.7 ± 1.7 a | 138.3 ± 2.9 a | 0.97 | <0.0001 |
| S | 135.2 ± 6.9 b | 6.13 ± 1.8 b | 134.0 ± 7.7 b | 0.99 | <0.0001 | |
| Leaf number per plant | R | 37.9 ± 2.5 a | 5.11 ± 1.0 a | 110.1 ± 4.2 a | 0.99 | <0.0001 |
| S | 24.8 ± 1.0 b | 8.3 ± 1.4 b | 91.3 ± 2.5 b | 0.99 | <0.0001 | |
| Number of tillers per plant | R | 6.6 ± 0.4 a | 6.8 ± 0.8 a | 110.2 ± 3.7 a | 0.99 | <0.0001 |
| S | 3.73 ± 0.1 b | 4.8 ± 0.1 b | 79.9 ± 1.1 b | 0.98 | <0.0001 | |
| Dry weight | R | 2848.2 ± 52.1 a | 9.7 ± 1.9 a | 147.4 ± 6.0 a | 0.99 | <0.0001 |
| S | 1425.9 ± 39.8 b | 7.8 ± 1.7 b | 126.0 ± 2.9 b | 0.99 | <0.0001 | |
| LAI | R | 3.2 ± 0.6 a | 7.5 ± 0.2 a | 102.7 ± 1.6 a | 0.98 | <0.0001 |
| S | 2.3 ± 0.1 b | 2.5 ± 1.2 b | 92.8 ± 2.6 b | 0.98 | <0.0001 | |
| LAR | R | 0.07 ± 0.01 a | 4.1 ± 0.6 a | 66.2 ± 1.1 a | 0.98 | <0.0001 |
| S | 0.07 ± 0.02 a | 4.0 ± 0.2 a | 65.7 ± 2.8 a | 0.98 | <0.0001 | |
| Trait | Population | Parameters 1 | R2 | p-Value * | |||
|---|---|---|---|---|---|---|---|
| Amax | Tb | Tc | Tp | ||||
| NAR | R | 0.33 ± 0.66 a | 59.5 ± 1.3 a | 204.3 ± 0.7 a | 138.1 ± 2.0 a | 0.97 | <0.0001 |
| S | 0.22 ± 0.28 b | 49.1 ± 1.4 b | 193.5 ± 1.2 b | 124.2 ± 2.9 b | 0.99 | <0.0001 | |
| PGR | R | 32.4 ± 2.4 a | 69.2 ± 2.5 a | 205.8 ± 1.4 a | 149.1 ± 2.8 a | 0.99 | <0.0001 |
| S | 18.4 ± 2.41 b | 57.8 ± 2.4 b | 196.6 ± 2.6 b | 139.2 ± 1.2 b | 0.99 | <0.0001 | |
| Population | Spikes per Plant | Spike Length (cm) | Seeds per Plant | 1000-Seed Weight (g) | Seed Area (mm2) |
|---|---|---|---|---|---|
| R | 6.0 ± 0.1 a | 8.4 ± 0.1 a | 1276 ± 17 a | 3.2 ± 0.4 a | 11.9 ± 0.5 a |
| S | 3.8 ± 0.1 b | 5.5 ± 0.6 b | 751 ± 21 b | 3.3 ± 0.2 a | 11.3 ± 0.6 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golmohammadzadeh, S.; Gherekhloo, J.; Osuna, M.D.; Ghaderi-Far, F.; Kamkar, B.; Alcántara-de la Cruz, R.; De Prado, R. Physiological Fitness Associated to ACCase Target-Site Resistance Enhances Growth and Reproduction in Phalaris brachystachys. Agronomy 2022, 12, 1206. https://doi.org/10.3390/agronomy12051206
Golmohammadzadeh S, Gherekhloo J, Osuna MD, Ghaderi-Far F, Kamkar B, Alcántara-de la Cruz R, De Prado R. Physiological Fitness Associated to ACCase Target-Site Resistance Enhances Growth and Reproduction in Phalaris brachystachys. Agronomy. 2022; 12(5):1206. https://doi.org/10.3390/agronomy12051206
Chicago/Turabian StyleGolmohammadzadeh, Sajedeh, Javid Gherekhloo, María Dolores Osuna, Farshid Ghaderi-Far, Behnam Kamkar, Ricardo Alcántara-de la Cruz, and Rafael De Prado. 2022. "Physiological Fitness Associated to ACCase Target-Site Resistance Enhances Growth and Reproduction in Phalaris brachystachys" Agronomy 12, no. 5: 1206. https://doi.org/10.3390/agronomy12051206
APA StyleGolmohammadzadeh, S., Gherekhloo, J., Osuna, M. D., Ghaderi-Far, F., Kamkar, B., Alcántara-de la Cruz, R., & De Prado, R. (2022). Physiological Fitness Associated to ACCase Target-Site Resistance Enhances Growth and Reproduction in Phalaris brachystachys. Agronomy, 12(5), 1206. https://doi.org/10.3390/agronomy12051206