Abstract
Nitrogen (N) deficiency causes a morphological disequilibrium between shoots and roots, favoring hypogeal growth. Our hypothesis is that short-term nitrogen deficiency could induce an increase in vine resilience, inducing root growth before the appearance of significant dysfunctions in the epigeal part of the plant. Then, nitrogen availability could be promptly restored through a well-thought-out fertilization plan. In this experiment, two table grape cultivars grafted onto three different rootstocks were studied in three hydroponic growing conditions characterized by different nitrogen availability. After 53 days of treatment, plant morphologies were described, and leaf photosynthetic pigments were quantified. The short-term nitrogen deficiency induced root growth, leading to significantly different proportions among the hypogeal and the epigeal parts, despite no significant differences were recorded concerning shoot growth. Some small differences in the leaf pigmentations indicated an incipient symptom of nitrogen deficiency, and significant differences in the phytomer morphology indicated the possibility of developing low-cost indices for prompt identification of incipient symptoms. This will allow the exploitation of the advantages of short-term nitrogen deficiency while avoiding negative effects on shoot growth by promptly restoring the nitrogen availability.
1. Introduction
Grapevines are a major crop worldwide [1,2,3]. They can be grown with limited water supplies, occupying also dry and semidry lands [4,5]. However, climate change could exacerbate drought stress in many viticultural regions, causing serious problems in grape and wine production [6,7].
A strategy commonly suggested for resilience improvement in viticulture is related to plant selection. A wide number of Vitis species are available in nature [8,9] and could be used for crossing programs to develop new rootstocks [10,11]. However, cultivar selection programs last many years and the rootstocks available for winegrowers are usually limited to a small number of traditional genotypes propagated by nurseries. Thus, other strategies of adaptation should be taken into consideration to complement variety choice.
It has been shown that vegetative growth and grape quality can be strongly affected by vineyard management in drought conditions [12,13]. Thus, new strategies should be explored to improve vineyard resilience to drought environments.
Growing conditions significantly affect the vine phenotype, including the plant morphology [14]. For sustainable water management, it is fundamental to consider the relationship between root-water uptake and leaf transpiration [15], keeping in mind the specific roles of the different plant organs in water use. Generally, increased root development is related to increased canopy water demand [16]. Nevertheless, adjustment in the roots’ growth could be related to the water needs for leaf transpiration, causing modifications in the proportions among roots and leaf biomasses, with different root depths [17] depending on the plant biomass [18].
From a morphological perspective, lower transpiration should be expected in plants with less expanded canopies [19], because vines lose water through their leaves. If water availability represents a limitation for transpiration, the plant adapts its morphology to decrease the water loss by reducing the leaf area. In drought conditions, it has been shown that there is a significantly lower leaf area due to a reduction in size of the shoot leaves and a smaller number of leaves situated on lateral shoots [13].
On the other hand, vines absorb water through their roots. In general, root elongation and the increase of soil scouting play a central role in adaptation to abiotic stresses [20,21,22,23,24], including a higher tolerance to drought conditions [25,26]. In fact, it has been demonstrated that a morphological adaptation strategy against drought is the production of deeper roots that are able to search for wetter soils [27]. Furthermore, the proliferation of roots, by increasing the number and branching connections, increases the water conductance [28].
It is worth noting that roots represent the interface between the plant and the soil, and being exposed to many abiotic stresses, different factors can affect their growth [29,30]. In fact, roots can adjust their development strategy, playing an essential role in plant performance during abiotic stresses [31,32]. Nitrogen is the most abundant nutrient and it directly affects grapevine growth [33,34,35,36,37]. It is widely known that an important adaptive strategy for plants subjected to nitrogen deficiency is root elongation. Compared to sufficient N supply conditions, it has been shown that crops grown under low N availability can increase their axial root length by about 40%, and the elongation of axial roots begins to increase after the reduction of nitrogen nutrition, demonstrating a cause–effect relationship [20].
Thus, our hypothesis is that a short-term nitrogen deficiency in young vines could induce an increase in plant resilience by modifying plant morphology (root/shoot growth ratio). We want to understand if an induction of root growth could be achieved before the appearance of evident symptoms of dysfunction in the shoots. Nitrogen is the most mobile and dynamic element in soil; it can be leached, denitrified, lost, or transferred [38]. Thus, keeping in mind that N availability can be promptly restored through a well-thought-out fertilization plan, we want to check the possibility of developing a fast and low-cost morphological method for nitrogen nutritional status detection. Finally, as N uptake also depends on the genotype [39,40], in this work, the effects of short-term nitrogen deficiency on plant morphology have been studied on different combinations of scion-rootstock vines, to increase the reliability of our results.
The active growth of roots under low N availability is related to the translocation of substances from old leaves and permanent organs to the tips of the root. In this way, the plants exposed to nutrient stress increase root growth and reduce canopy expansion [41,42], which is also responsible for water loss through transpiration. On the other hand, the growing capacity of the root system depends on the organs of reserve and transport, which is supported by the leaf apparatus [43], and N deficiency negatively affects shoot growth and the plant’s photosynthetic performances [41,44,45]. Grafted rooted cuttings that are commercially available are expected to have high quality woody organs with high quantities of reserves [46]. However, short-term nitrogen deficiency during root growth stimulation should not excessively impact the photosynthetic performance of the new leaves. Thus, in this experiment, leaf photosynthetic pigments were also quantified.
2. Materials and Methods
2.1. Plant Material and Experimental Plan
The experiment was carried out in 2021 at the National Research & Development Institute for Biotechnologies in Horticulture Stefanesti Arges (Stefanesti, Romania). The plant material was provided by the Research and Development Institute for Viticulture and Oenology Valea Calugareasca (Valea Calugareasca, Romania), and it respected the commercial standards for Romanian viticulture. Two table grape cultivars (Afuz Ali and Italia) were grafted on the most used rootstocks in Romania: Teleki 4 Sel. Oppenheim 4-4Bl (SO4-4), Kobber 5BB, and Teleki 8 BB Sel. Craciunel 71 (C71). At the beginning of the experiment, the plants were pruned following local traditions for new vineyard plantation: three buds were left in the scions and about 10 cm of roots were left in the rootstocks. Three hydroponic growing conditions were obtained by modifying the nitrogen availability and five plant replications were used for each grafting combination. Thus, for this experiment, a total of 90 grafted cuttings were studied.
2.2. Hydroponic Growing Conditions
Three plastic tanks (110/90/65 cm) were used for hydroponic cultivation. Each tank was filled with 500 L of a nutrient solution with different nitrogen availability, as described below. Air was pumped into the tanks for 45 min/h with a timer (Emos Mecanische Schaltsteckdose) connected to an air pomp (Ocean Free ULTRAIR 1000) linked to two plastic air distributors (type T) with six channels, two air curtains, and eight air stones. A perforated floating platform made by extruded polystyrene (thickness 5 cm) was used as plant support. Figure 1 shows the experimental conditions.

Figure 1.
Photo of the experimental plants.
Each tank was filled with a different nutrient solution (Table 1), characterized by different nitrogen availability: full-nitrogen (standard concentrations), half-nitrogen, and no-nitrogen. The solutions were prepared by adapting the protocols reported in Rustioni et al. [40]. To study the effects of a short-term nitrogen deficiency on plant growth, vines were kept in these growing conditions for 53 days, renewing the nutrient solutions every 10 days.

Table 1.
Mineral composition of nutrient solutions. Concentrations are reported in mM.
2.3. Analytical Methods and Data Elaborations
At the end of the experiment, plants were observed and analyzed.
The phenological stage was measured by counting the unfolded leaves. New formed roots and shoot growth were measured by using two KERN scales (EHA 500-2 and TGD 50-3C). Total leaf area was measured with the smartphone application “Easy leaf Area Free” [47]. These data were also used to calculate the ratios among roots and shoots or roots and leaf area. Phytomer morphology was described following the method reported previously by Dinu et al. [14]. The third, fourth, and fifth phytomers were considered. The lengths of the internodes and petioles were measured with a tape line, and diameters of the internodes and petioles were obtained with a digital caliper (INSIZE 1108-150W). Areas were measured with the smartphone application “Easy leaf Area Free” [47].
The photosynthetic pigments were quantified using the method described by Holm [48]. Briefly, 100 mg of leaves were finely crushed with a mortar and extracted with 25 mL of 85% acetone. After filtration through a filter paper of MN 615-Ø 110 mm, the absorbance of the solution was recorded at 440.5, 662, and 644 nm. The concentrations of the pigments were calculated by using the formulas:
En = absorbance recorded at n wavelength.
The ratios of Chlorophyll a and Chlorophyll b were also calculated, as well as the total concentrations of photosynthetic pigments (sum of all the different pigment types).
All statistical analyses were done in SPSS (IBM Corporation, Armonk, NY, USA). Significances of the differences among groups of samples was quantified by analysis of variance (ANOVA) and Duncan post-hoc. A principal component analysis (PCA) was performed on all direct and derivative variables considered for phytomer morphology.
3. Results
3.1. Morphological Adaptation to the Short-Term Nitrogen Deficiency
The short-term nitrogen deficiency lasted 53 days. In this time, no significant effects of nitrogen availability, cultivar, or rootstock were observed concerning the growth of the shoots quantified as total weight of the green organs. The shoot weight was 14.2 ± 7 g. In general, the average phenological stage reached by the shoot growth was 11 ± 3 unfolded leaves.
On the contrary, the short-term nitrogen deficiency significantly increased the root growth (Figure 2). The determinations for the evaluation of the growth of the root system proved that a high concentration of nitrogen induced a reduction of the growth rate of the roots and of the formation of new roots. The average newly formed roots weight in full nitrogen conditions was 0.19 ± 0.24 g. Reducing the amount of nitrogen by half and eliminating nitrogen from the nutrient solution were shown to be similarly effective in stimulating the growth of the root system and the formation of new roots (the new formed roots weights were 1.96 ± 1.16 g and 1.64 ± 1.25, respectively).
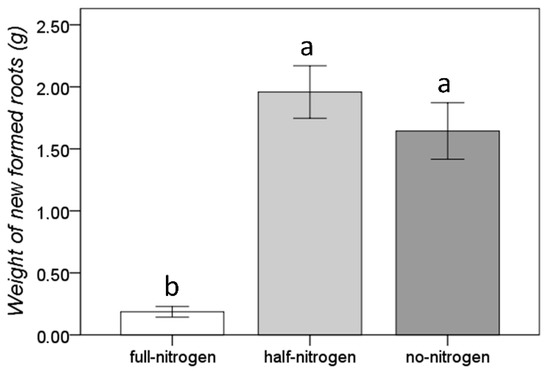
Figure 2.
Effect of the short-term nitrogen deficiency on root growth. Bars represents the standard error; different letters indicate significant differences among groups resulted found by the Duncan post-hoc test.
No significant differences in the root growth were ascribable to the vine cultivar, but significant differences were related to the rootstock, with higher values in SO4 with respect to Kober 5BB and C71 (average data for each grafting combinations are available in Supplementary Materials Table S1).
This differential root growth led to significantly different proportions among the hypogeal and the epigeal parts (Figure 3). Considering both the ratio between root and shoot growth and the ratio between root growth and leaf area, a significant increase in the values was recorded in both the total and partial nitrogen deficiency conditions. This indicates that the short-term nitrogen deficiency induced an adaptive response of the plants to this type of stress and an over-expression of root system growth.
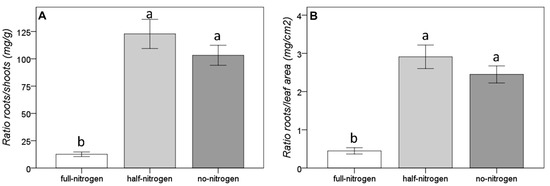
Figure 3.
Effect of the short-term nitrogen deficiency on the proportions of the hypogeal and the epigeal parts ((A): ratio roots/shoots; (B): ratio roots/leaf area). Bars represent the standard error; different letters indicate significant differences among group results found by the Duncan post-hoc test.
Once again, no significant differences in the proportions among the hypogeal and the epigeal parts were ascribable to the grape cultivar, but significant differences were related to the rootstock, with higher values in SO4 with respect to Kober 5BB and C71 (both the ratios among roots and shoots or roots and leaf area). Average data for each grafting combinations are available in Supplementary Materials (Table S1) and photos of the plants are available in Supplementary Materials (Figure S1).
Some differences in the leaf pigmentation indicated an incipient symptom of nitrogen deficiency, especially in the without nitrogen condition (Figure 4). In detail, the absence of nitrogen induced a decrease in the total pigments due to a significant decrease of chlorophyll a, chlorophyll b, and carotenoids. However, the proportion between the two chlorophylls remained unaltered. On the other hand, when the nitrogen availability was halved, only the concentration of chlorophyll b was significantly affected, while other pigments maintained concentrations similar to those recorded in the control plants.
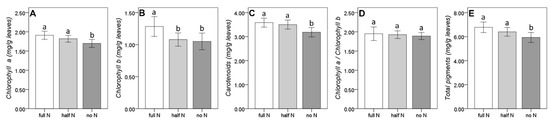
Figure 4.
Effect of the short-term nitrogen deficiency on the leaf pigmentation ((A): chlorophyll a; (B): chlorophyll b; (C): carotenoids; (D): chlorophyll a/chlorophyll b; (E): total pigments). Bars represent the standard error; different letters indicate significant differences among group results found by the Duncan post-hoc test.
Regarding the varieties, we observed significantly higher concentrations of all chlorophyll pigments in Italia, while the ratio of chlorophyll a/chlorophyll b was significantly higher in Afuz Ali. The rootstock significantly affected only chlorophyll a, with significant differences between the higher concentrations observed in SO4-4 and the lower ones of Kober 5BB (C71 showed intermediate values). Average data for each grafting combinations are available in Supplementary Materials (Table S1).
3.2. Shoot Morphology Indexes of Incipient Nitrogen Deficiency
The principal component analysis of the 16 shoot morphology parameters produced five functions with Eigenvalues higher than 1, explaining, respectively, 33.9%, 30.7%, 17.4%, 9.3%, and 6.5% of the total observed variance. Details concerning the matrix of the five main principal component functions are reported in Supplementary Materials (Table S2). Briefly, the first principal component is mainly related to the leaf area and to the proportional distribution among the green surfaces (mainly leaf and internodal stem). The second principal component is mainly linked to the internode development (stem surface and length). The third principal component is related to the petiole proportion with respect to the leaf and total green surface. The stem diameter is the parameter mainly affecting the fourth principal component, while the ratio between petiole length and diameter is the parameter mainly affecting the fifth principal component.
Focusing on the first two components, it appears that the first is related to nitrogen availability (Figure 5A) and the second is more related to the cultivar characteristics (Figure 5B).
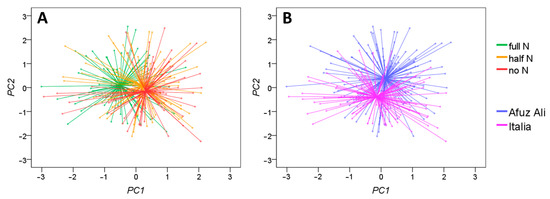
Figure 5.
Principal component analysis (PCA) of the 16 shoot morphological parameters, ordering the samples according to nitrogen availability (A) and the cultivar (B). The first two components (representative of the 64.6% of the variance) are reported.
Despite the short-term nitrogen deficiency not producing strong differences in the total epigeal growth and pigmentation, the shoot morphology underwent significant modifications in relation to the nitrogen availability. Table 2 reports the significances of the effects of N availability, variety, rootstock, and internode (and all their possible interactions) on the regression factor scores of the five functions with Eigenvalues higher than 1 (altogether representing 97.8% of the morphological variability detected in this work).
Table 2.
Results of the ANOVA of the effects of N availability, variety, rootstock, and internode (and all their possible interactions) on the regression factor scores of the five functions with Eigenvalues higher than 1 (PC = principal component).
Considering the principal effects, it is worth noting the significant impacts of the variety and the internode position on all the regression factor scores. The nitrogen availability also showed significant effects on most of the principal components (except PC2), while the rootstock was the least impactful variable. Some significant interaction effects were highlighted as well (Table 2).
4. Discussion
It is well known that nitrogen deficiency induces dysfunctions in plants’ green organs, causing a reduced growth of shoots [41]. However, in our experiment, we observed that a short-term nitrogen deficiency did not induce significant modifications in the total weight of the green shoots. This is likely due to the nitrogen reserves stored into the young plants grown in nursery. In fact, nitrogen is accumulated in woody tissues. Nitrogen is almost equally distributed in the perennial tissues of the plant, with a slightly higher concentration in roots. During dormancy, 75% of the N reserves are located in the roots and this concentration decreases moderately during the spring growth [49,50].
In conditions of nitrogen deficiency, plants induce root elongation, increasing soil scouting [20]. It is known that the root elongation and the increase of soil scouting induce abiotic stresses tolerance [16,17,18,19,20], including a higher tolerance to drought [25,26]. Our data indicates that the hypogeal response is fast, occurring before the appearance of evident deficiency symptoms in the shoot growth. Thus, an incipient deficiency related to a short-term treatment could also induce this morphological adaptation. Furthermore, both the without nitrogen condition and the half nitrogen condition had significantly higher values than the full nitrogen availability concerning the new formed roots, as well as both the ratios among roots and shoots or roots and leaf area. In nitrogen deficiency conditions, slightly better (not significant) performances were observed where some nitrogen was available. This is probably due to the better photosynthetic performances of the plants grown in this condition due to the lower impact on leaf pigmentation. In general, we can suppose that a more realistic partial nitrogen deficiency (half N) is sufficient to induce the root growth and that this adaptive mechanism appears before the occurrence of significant shoot dysfunctions. Thus, a short-term nitrogen deficiency of few weeks could be a useful tool to induce root elongation in the first period after vineyard plantation, improving the vines’ resilience to severe abiotic constraints.
Of course, to exploit the advantages of a short-term nitrogen deficiency and avoid negative effects on the plant growth, it is important to promptly restore the nitrogen availability to avoid plant damages. Nitrogen is a mobile element in the soil that can be leached, denitrificated, lost, and transferred [38]. To maximize the response of the plants to nitrogen restoration, different techniques able to rapidly supply nitrogen to the plants could be taken into consideration, such as fertigation and foliar fertilization [33,51,52]. Further experiments in soil will be necessary to develop a realistic fertilization strategy for short-term nitrogen deficiency management.
To allow prompt nitrogen fertilization able to restore the plant growth equilibrium before the occurrence of damages, it is important to promptly recognize the incipient symptoms of deficiency in the visible shoot growth. Leaf element analysis could support the fertilization management [53]. However, the analytical costs and the laboratory equipment necessary for this analysis (not widely available for winegrowers) could make the frequent check over time difficult during the short-term deficiency treatment.
Different simple methods are available for pigment quantification, both with sample extraction [48,54] or with non-invasive approaches [55,56,57,58,59]. Despite the fact that the ratio of chlorophyll a and chlorophyll b was not significantly affected by the nitrogen availability, the only pigment significantly reduced in the half nitrogen condition was chlorophyll b. This result, in agreement with the observations of Rustioni et al. [44], suggests that concentration in chlorophyll b could be a good index of incipient nitrogen deficiency. On the other side, many biotic and abiotic stresses modify the leaves’ pigmentation, inducing chlorosis [44,59,60,61,62]. Thus, other observations could be useful to complete the diagnosis, allowing a more reliable decision.
References are available in literature, concerning the shoot morphological modifications in relation to nitrogen deficiency. It is known that reducing the photosynthesis and the leaf area affects plant development [45,63,64], causing an evident decrease of the shoot growth [65].
In our case, we need to find indices that are able to promptly highlight incipient deficiency, since if the symptoms are already evident in the shoots, the plant could have already undergone damage. Dinu et al. [14] demonstrated the possibility of using shoot morphological descriptors to discriminate the grapevine growing conditions in the absence of evident dysfunctions. The same parameters were measured in this work, and the data elaboration confirmed this result. In fact, the described morphological variability was significantly affected by the nitrogen availability.
5. Conclusions
Short-term nitrogen deficiency could be a useful tool to induce root growth. Thus, a new fertilization strategy in vineyard planting could be developed in the near future to improve young vineyard resilience through a temporary nitrogen deficiency. For example, soil amendments with a high C/N ratio could be used to cause a temporary nitrogen deficiency for plants due to the microorganisms’ competitive interactions in the soil.
With this research, we demonstrated that a brief deficiency could obtain this effect before the appearance of dysfunctions in the shoots. By changing the shoot/root growth ratio, an improvement of resilience to severe abiotic constraints (such as drought) should be expected; however, further studies will be necessary to confirm this response in field conditions and to optimize the length of the short-term deficiency, as well as to validate the drought tolerance modulation.
To promptly restore the nitrogen availability before the appearance of plant dysfunctions, low-cost and easy analysis based on plant morphology and leaf pigmentation could be developed as a tool to manage fertilization. This will allow the interruption of the short-term nitrogen deficiency before shoot damages. Further studies will be devoted to the development of these indices during the course of the induced nitrogen deficiency. Finally, a well-thought-out fertilization strategy could be set up to promptly restore the nitrogen availability, taking into consideration techniques such as fertigation and foliar fertilization.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12061355/s1, Table S1: average data recorded for each grafting combinations; Table S2: matrix of the five main principal component functions; Figure S1: photos of the plant growth at the end of the experiment. New formed roots are recognizable by the lighter color.
Author Contributions
Conceptualization L.R. and D.G.D.; methodology, L.R., D.G.D. and C.F.P.; analysis, D.G.D., C.F.P., A.E.M. and L.M.P.; resources, C.F.P. and D.I.S.; data elaboration, L.R., D.G.D. and L.M.P.; writing—original draft preparation, D.G.D. and L.R.; writing—review and editing, D.G.D., C.F.P., D.I.S., A.E.M., L.M.P. and L.R.; supervision, L.R., C.F.P. and D.I.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Rough data can be requested from the corresponding author.
Acknowledgments
We thank our colleagues from the National Research & Development Institute for Biotechnologies in Horticulture Stefanesti Arges. A special thanks to Badulescu Adriana, Vizitiu Diana Elena, Sardarescu Ionela Daniela, Stan Ramona Onache Anca, Nedelea Gina, and Anghel Gheorghe. We also acknowledge Marian Ion, Elena Branduse, and Burlacu Cristian from the Research and Development Institute for Viticulture and Oenology Valea Calugareasca (Romania) for providing the plant material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rustioni, L.; Cola, G.; Maghradze, D.; Abashidze, E.; Argiriou, A.; Aroutiounian, R.; Brazão, J.; Chipashvili, R.; Cocco, M.; Cornea, V.; et al. Description of the Vitis vinifera L. phenotypic variability in eno-carpological traits by a Euro-Asiatic collaborative network among ampelographic collections. Vitis 2019, 58, 37–46. [Google Scholar]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Bisson, L.F.; Waterhouse, A.L.; Ebeler, S.E.; Walker, M.A.; Lapsley, J.T. The present and future of the international wine industry. Nature 2002, 418, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Cifre, J.; Bota, J.; Escalona, J.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.): An open gate to improve water-use efficiency? Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Chalmers, Y.; Kelly, G.; Krstic, M. Partial Rootzone Drying of Vitis Vinifera cv. ‘shiraz’ Winegrapes in a Semi-Arid Climate. Acta Hortic. 2004, 664, 133–138. [Google Scholar] [CrossRef]
- Gambetta, G.; Manuck, C.M.; Drucker, S.T.; Shaghasi, T.; Fort, K.; Matthews, M.A.; Walker, M.A.; McElrone, A.J. The relationship between root hydraulics and scion vigour across Vitis rootstocks: What role do root aquaporins play? J. Exp. Bot. 2012, 63, 6445–6455. [Google Scholar] [CrossRef]
- de Orduña, R.M. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Padgett-Johnson, M.; Williams, L.E.; Walker, M.A. Vine water relations, gas exchange, and vegetative growth of seventeen Vitis species grown under irrigated and nonirrigated con conditions in California. J. Am. Soc. Hortic. Sci. 2003, 128, 269–276. [Google Scholar] [CrossRef]
- Rustioni, L.; Ciacciulli, A.; Grossi, D.; Brancadoro, L.; Failla, O. Stem Xylem Characterization for Vitis Drought Tolerance. J. Agric. Food Chem. 2016, 64, 5317–5323. [Google Scholar] [CrossRef]
- Bianchi, D.; Grossi, D.; Tincani, D.T.; Di Lorenzo, G.S.; Brancadoro, L.; Rustioni, L. Multi-parameter characterization of water stress tolerance in Vitis hybrids for new rootstock selection. Plant Physiol. Biochem. 2018, 132, 333–340. [Google Scholar] [CrossRef]
- Bianchi, D.; Grossi, D.; Di Lorenzo, G.S.; Ying, Y.Z.; Rustioni, L.; Brancadoro, L. Phenotyping of the “G series” Vitis hybrids: First screening of the mineral composition. Sci. Hortic. 2020, 264, 109155. [Google Scholar] [CrossRef]
- Ortega-Farias, S.; Fereres, E.; Sadras, V.O. Special issue on water management in grapevines. Irrig. Sci. 2012, 30, 335–337. [Google Scholar] [CrossRef][Green Version]
- Gómez-Del-Campo, M.; Ruiz, C.; Baeza, P.; Lissarrague, J.R. Drought adaptation strategies of four grapevine cultivars (Vitis vinifera L.): Modification of the properties of the leaf area. OENO One 2003, 37, 131–143. [Google Scholar] [CrossRef]
- Dinu, D.G.; Bianchi, D.; Mamasakhlisashvili, L.; Quarta, C.; Brancadoro, L.; Maghradze, D.; Cardinale, M.; Rustioni, L. Effects of genotype and environmental conditions on grapevine (Vitis vinifera L.) shoot morphology. Vitis 2021, 60, 85–91. [Google Scholar] [CrossRef]
- Green, S.R.; Kirkham, M.; Clothier, B.E. Root uptake and transpiration: From measurements and models to sustainable irrigation. Agric. Water Manag. 2006, 86, 165–176. [Google Scholar] [CrossRef]
- Gambetta, G.A. Water Stress and Grape Physiology in the Context of Global Climate Change. J. Wine Econ. 2016, 11, 168–180. [Google Scholar] [CrossRef]
- Jackson, R.B.; Sperry, J.S.; Dawson, T.E. Root water uptake and transport: Using physiological processes in global predictions. Trends Plant Sci. 2000, 5, 482–488. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.B.; Mooney, H.A.; Sala, O.; Schulze, E.-D. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Archer, E.; Strauss, H. The Effect of Vine Spacing on Some Physiological Aspects of Vitis vinifera L. (cv. Pinot noir). South Afr. J. Enol. Vitic. 2017, 11, 76–87. [Google Scholar] [CrossRef][Green Version]
- Sun, X.; Chen, F.; Yuan, L.; Mi, G. The physiological mechanism underlying root elongation in response to nitrogen deficiency in crop plants. Planta 2020, 251, 84. [Google Scholar] [CrossRef]
- Mariani, L.; Ferrante, A. Agronomic Management for Enhancing Plant Tolerance to Abiotic Stresses—Drought, Salinity, Hypoxia, and Lodging. Horticulturae 2017, 3, 52. [Google Scholar] [CrossRef]
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.M.; Testerink, C. Roots Withstanding their Environment: Exploiting Root System Architecture Responses to Abiotic Stress to Improve Crop Tolerance. Front. Plant Sci. 2016, 7, 1335. [Google Scholar] [CrossRef] [PubMed]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Mi, G.; Chen, F.; Wu, Q.; Lai, N.; Yuan, L.; Zhang, F. Ideotype root architecture for efficient nitrogen acquisition by maize in intensive cropping systems. Sci. China Life Sci. 2010, 53, 1369–1373. [Google Scholar] [CrossRef]
- Köse, B.; Karabulut, B.; Ceylan, K. Effect of Rootstock on Grafted Grapevine Quality Europ. J. Hort. Sci. 2014, 79, 197. [Google Scholar]
- Corso, M.; Bonghi, C. Grapevine rootstock effects on abiotic stress tolerance. Plant Sci. Today 2014, 1, 108–113. [Google Scholar] [CrossRef]
- Bauerle, T.L.; Smart, D.R.; Bauerle, W.L.; Stockert, C.; Eissenstat, D.M. Root foraging in response to heterogeneous soil moisture in two grapevines that differ in potential growth rate. New Phytol. 2008, 179, 857–866. [Google Scholar] [CrossRef]
- Alsina, M.M.; Smart, D.R.; Bauerle, T.; De Herralde, F.; Biel, C.; Stockert, C.; Negron, C.; Save, R. Seasonal changes of whole root system conductance by a drought-tolerant grape root system. J. Exp. Bot. 2010, 62, 99–109. [Google Scholar] [CrossRef]
- Franco, J.A.; Bañón, S.; Vicente, M.J.; Miralles, J.; Martínez-Sánchez, J.J. Review Article: Root development in horticultural plants grown under abiotic stress conditions–A review. J. Hortic. Sci. Biotechnol. 2011, 86, 543–556. [Google Scholar] [CrossRef]
- Wells, C.E.; Eissenstat, D.M. Beyond the Roots of Young Seedlings: The Influence of Age and Order on Fine Root Physiology. J. Plant Growth Regul. 2002, 21, 324–334. [Google Scholar] [CrossRef]
- Maurel, C.; Simonneau, T.; Sutka, M. The significance of roots as hydraulic rheostats. J. Exp. Bot. 2010, 61, 3191–3198. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-J.; Yang, H.-Q.; Wang, J.-X. Arginine metabolism in roots and leaves of apple (Malus domestica Borkh.): The tissue-specific formation of both nitric oxide and polyamines. Sci. Hortic. 2009, 119, 147–152. [Google Scholar] [CrossRef]
- Vilanova, M.; Fandiño, M.; Frutos-Puerto, S.; Cancela, J.J. Assessment fertigation effects on chemical composition of Vitis vinifera L. cv. Albariño. Food Chem. 2018, 278, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.P.; Merkt, N.; Zörb, C. Different nitrogen (N) forms affect responses to N form and N supply of rootstocks and grafted grapevines. Plant Sci. 2018, 277, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Álvarez, E.P.; Martínez-Vidaurre, J.M.; Martín, I.; García-Escudero, E.; Peregrina, F. Relationships among Soil Nitrate Nitrogen and Nitrogen Nutritional Status, Yield Components, and Must Quality in Semi-arid Vineyards from Rioja AOC, Spain. Commun. Soil Sci. Plant Anal. 2013, 44, 232–242. [Google Scholar] [CrossRef]
- Akin, A.; Dardeniz, A.; Ates, F.; Celik, M. Effects of various crop loads and leaf fertilizer on grapevine yield and quality. J. Plant Nutr. 2012, 35, 1949–1957. [Google Scholar] [CrossRef]
- Coipel, J.; Lovelle, B.R.; Sipp, C.; Van Leeuwen, C. “Terroir” effect, as a result of enviromental stess, depends more on soil depth than on soil type (Vitis vinifera L. cv. Grenache Noir, Côtes du Rhône, France, 2000). OENO One 2006, 40, 177–185. [Google Scholar] [CrossRef]
- Delgado, J.A.; Kowalski, K.; Tebbe, C. The first Nitrogen Index app for mobile devices: Using portable technology for smart agricultural management. Comput. Electron. Agric. 2013, 91, 121–123. [Google Scholar] [CrossRef]
- Xu, X.-J.; Li, Q.-Y.; Song, X.-H.; Qi-Rong, S.; Cai-Xia, D. Dynamic Regulation of Nitrogen and Organic Acid Metabolism of Cherry Tomato Fruit as Affected by Different Nitrogen Forms. Pedosphere 2012, 22, 67–78. [Google Scholar] [CrossRef]
- Roubelakis-Angelakis, K.A.; Kliewer, W.M. Nitrogen Metabolism in Grapevine. Hortic. Rev. 1992, 14, 407–452. [Google Scholar] [CrossRef]
- Grechi, I.; Vivin, P.; Hilbert, G.; Milin, S.; Robert, T.; Gaudillère, J.-P. Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine. Environ. Exp. Bot. 2007, 59, 139–149. [Google Scholar] [CrossRef]
- Robinson, D. Compensatory Changes in the Partitioning of Dry Matter in Relation to Nitrogen Uptake and Optimal Variations in Growth. Ann. Bot. 1986, 58, 841–848. [Google Scholar] [CrossRef]
- Keller, M.; Koblet, W. Dry matter and leaf area partitioning, bud fertility and second season growth of Vitis vinifera L: Responses to Nitrogen supply and limiting irradiance. Vitis 1995, 34, 77–83. [Google Scholar]
- Rustioni, L.; Grossi, D.; Brancadoro, L.; Failla, O. Iron, magnesium, nitrogen and potassium deficiency symptom discrimination by reflectance spectroscopy in grapevine leaves. Sci. Hortic. 2018, 241, 152–159. [Google Scholar] [CrossRef]
- Metay, A.; Magnier, J.; Guilpart, N.; Christophe, A. Nitrogen supply controls vegetative growth, biomass and nitrogen allocation for grapevine (cv. Shiraz) grown in pots. Funct. Plant Biol. 2015, 42, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Somkuwar, R.G.; Satisha, J.; Ramteke, S.D. Berry weight, quality and cane biochemistry changes in relation to cane thickness of own-rooted and grafted ‘Tas-A-Ganesh’ grape. J. Hortl. Sci. 2013, 8, 30–34. [Google Scholar]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated digital image analysis for rapid and accurate measurement of leaf area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef]
- Holm, G. Chlorophyll Mutations in Barley. Acta Agric. Scand. 1954, 4, 457–471. [Google Scholar] [CrossRef]
- Zapata, C.; Deléens, E.; Chaillou, S.; Magné, C. Partitioning and mobilization of starch and N reserves in grapevine (Vitis vinifera L.). J. Plant Physiol. 2004, 161, 1031–1040. [Google Scholar] [CrossRef]
- Najar, A.; Landhäusser, S.M.; Whitehill, J.G.A.; Bonello, P.; Erbilgin, N. Reserves Accumulated in Non-Photosynthetic Organs during the Previous Growing Season Drive Plant Defenses and Growth in Aspen in the Subsequent Growing Season. J. Chem. Ecol. 2013, 40, 21–30. [Google Scholar] [CrossRef]
- Stefanello, L.O.; Schwalbert, R.; Schwalbert, R.A.; De Conti, L.; Kulmann, M.S.D.S.; Garlet, L.P.; Silveira, M.L.R.; Sautter, C.K.; de Melo, G.W.B.; Rozane, D.E.; et al. Nitrogen supply method affects growth, yield and must composition of young grape vines (Vitis vinifera L. cv Alicante Bouschet) in southern Brazil. Sci. Hortic. 2019, 261, 108910. [Google Scholar] [CrossRef]
- Lacroux, F.; Trégoat, O.; Van Leeuwen, C.; Pons, A.; Tominaga, T.; Lavigne-Cruège, V.; Dubourdieu, D. Effect of foliar nitrogen and sulphur application on aromatic expression of Vitis vinifera L. cv. Sauvignon blanc. OENO One 2008, 42, 125–132. [Google Scholar] [CrossRef]
- Romero, I.; Benito, A.; Domínguez, N.; García-Escudero, E.; Martín, I. Leaf blade and petiole nutritional diagnosis for Vitis vinifera L. cv. Tempranillo by deviation from optimum percentage method. Span. J. Agric. Res. 2014, 12, 206. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids–Measurement and Characterisation by UV-VIS. In Current Protocols in Food Analytical Chemistry (CPFA), (Supplement 1), F4.3.1–F 4.3.8; John Wiley: New York, NY, USA, 2001. [Google Scholar]
- Rustioni, L.; Rocchi, L.; Di Meo, F.; Failla, O.; Trouillas, P. Advances in grape phenotyping: Pigment characterization by reflectance and theoretical chemistry. Vitis 2015, 54, 173–174. [Google Scholar]
- Rocchi, L.; Rustioni, L.; Failla, O. Chlorophyll and carotenoid quantifications in white grape (Vitis vinifera L.) skins by reflectance spectroscopy. VITIS-J. Grapevine Res. 2016, 55, 11–16. [Google Scholar] [CrossRef]
- Steele, M.R.; Gitelson, A.A.; Rundquist, D.C. A Comparison of Two Techniques for Nondestructive Measurement of Chlorophyll Content in Grapevine Leaves. Agron. J. 2008, 100, 779–782. [Google Scholar] [CrossRef]
- Shaahan, M.M.; El-Sayed, A.; El-Nour, E.A. Predicting nitrogen, magnesium and iron nutritional status in some perennial crops using a portable chlorophyll meter. Sci. Hortic. 1999, 82, 339–348. [Google Scholar] [CrossRef]
- Setshedi, K.J.; Mutingwende, N.; Ngqwala, N.P. The Use of Artificial Neural Networks to Predict the Physicochemical Characteristics of Water Quality in Three District Municipalities, Eastern Cape Province, South Africa. Int. J. Environ. Res. Public Health 2021, 18, 5248. [Google Scholar] [CrossRef]
- Rustioni, L.; Grossi, D.; Brancadoro, L.; Failla, O. Characterization of iron deficiency symptoms in grapevine (Vitis spp.) leaves by reflectance spectroscopy. Plant Physiol. Biochem. 2017, 118, 342–347. [Google Scholar] [CrossRef]
- Carter, G.A.; Knapp, A.K. Leaf optical properties in higher plants: Linking spectral characteristics to stress and chlorophyll concentration. Am. J. Bot. 2001, 88, 677–684. [Google Scholar] [CrossRef]
- Bavaresco, L. Relationship between chlorosis occurrence and mineral composition of grapevine leaves and berries. Commun. Soil Sci. Plan 1997, 28, 13–21. [Google Scholar] [CrossRef]
- Mu, X.; Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant Physiol. Biochem. 2020, 158, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.A.; Louarn, G.; Peña, J.P.; Ojeda, H.; Simonneau, T.; Lebon, E. A leaf gas exchange model that accounts for intra-canopy variability by considering leaf nitrogen content and local acclimation to radiation in grapevine (Vitis vinifera L.). Plant Cell Environ. 2012, 35, 1313–1328. [Google Scholar] [CrossRef] [PubMed]
- Vrignon-Brenas, S.; Aurélie, M.; Romain, L.; Shiva, G.; Alana, F.; Myriam, D.; Gaëlle, R.; Anne, P. Gradual responses of grapevine yield components and carbon status to nitrogen supply. OENO One 2019, 53, 289–306. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).