Abstract
Salicylic acid (SA) plays an important role in regulating salt stress tolerance in plants. However, there are no studies on the effect of exogenous SA on Saponaria officinalis under salt stress. To study the effectiveness of SA on mitigating salt stress, S. officinalis were used in a pot experiment of salt stress simulated with an NaCl solution (100, 200, and 300 mmol L−1), while an SA solution (0, 0.2, 0.4, 0.6, 0.8, 1.0 mmol L−1) was sprayed on leaves. Under salt stress, spraying SA caused an increase in the salt damage index, electrolyte leakage, and a reduction in malondialdehyde and Na+ content, but an increase in the rate of photosynthesis, chlorophyll, soluble sugar, soluble protein, free proline, K+, Mg2+, Ca2+ content, the K+/Na+ ratio, superoxide dismutase, peroxidase, catalase, ascorbate peroxidase activity, and the comprehensive score. The results show that SA improves the salt tolerance capacity of S. officinalis by modulating its photosynthetic rate, osmoprotectants, antioxidant levels, and ion homeostasis. However, the effectiveness of SA was not linearly related to its concentration. In summary, our findings reveal the protective roles of SA against salinity in S. officinalis and suggest that the use of 0.6 mmol L−1 of SA in salt stress conditions could be an effective approach to reduce the damage caused by saline soil in S. officinalis.
1. Introduction
Salinization is one of the top ten threats to the world’s soil resources. Salinity has affected more than 3600 Mha of soil, and the total area is expanding by around 1.5 Mha of land each year [1]. As an example, more than 70% of Northeast China is currently salinized land [2]. Improving the level of salinity in soil using physical or chemical methods is costly, of limited effectiveness, and can cause secondary soil salinity [3]. Plant repair is one of the best ways to improve the salt levels in soil. Saponaria officinalis is widespread in its distribution in the saline areas of northeast area of China, and has the potential to become a plant material for improved soil salinity. Additionally, the leaves of S. officinalis can be used to extract active surface agents, creating economic value [4]. However, salt stress may produce harmful effects on S. officinalis by osmotic stress, peroxidative damage, and ion toxicity [5]. This will not only affect the improvement of the saline soil, but also produce economic losses. Under such situations, it is necessary to explore ways to improve the salt tolerance capacity of S. officinalis.
The use of exogenous hormones to alleviate the harmful effects of salt stress on plants is providing new ideas for the use of various plants in saline soil [6]. Salicylic acid (SA) is a widely found phytohormone that stimulates various responses in plant cells under adverse conditions [7]. Researchers believe that SA is a potential growth regulator that can improve plant salt tolerance [7,8]. The positive effects of SA may stem from its involvement in activating the osmotic regulation system and antioxidant system [8,9], which reduce the adverse effects of salt stress, water deficiency, structural cell damage (disrupted chlorophyll, proteins, and lipids), disordered metabolic activity (blocked photosynthesis), and maintain ion homeostasis [10]. However, there are currently no studies on the effect of exogenous SA on S. officinalis under salt stress, and no reports of SA application on S. officinalis under salt stress. Whether or not SA can improve salt tolerance capacity and how SA improves salt tolerance capacity of S. officinalis are still unknown.
We speculated that SA could improve the salt tolerance capacity of S. officinalis by modulating the photosynthetic rate, osmoprotectants, antioxidant levels, and ion homeostasis. Thus, the following physiological indexes of S. officinalis were determined after SA treatment and the exposure to salt stress. The comprehensive score, salt damage index, electrolyte leakage, and malondialdehyde content were used to evaluate the degree of salt toxicity; the photosynthesis rate and chlorophyll content were used to evaluate the photosynthetic capabilities; soluble sugar, soluble protein, and free proline content were used to evaluate the accumulation of osmoprotectants; superoxide dismutase, peroxidase, catalase, and ascorbate peroxidase activity were used to evaluate the antioxidant levels; Na+, K+, Mg2+, Ca2+ content and the K+/Na+ ratio were used to evaluate ion homeostasis. We hope that exploring the effectiveness of SA in the mitigation of the adverse effects of salinity and investigating the possible mechanisms of SA enhancement of salt tolerance in the S. officinalis can provide a feasible method and an effective concentration to promote the growth of S. officinalis under salt stress.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
The experiment was conducted in a randomized complete block design. Each group had 24 pots of two-year-old S. officinalis. An amount of 1000 g dray mixture of cultivated soil (N, 2.8%; P2O5, 1.9%; K2O, 0.8%; organic matter, 60%; humic acid, 35%) and vermiculite at 3:1 (v:v) was packed in a plastic pot (170 × 123 × 153 mm). The experiment was started 30 days after S. officinalis were transplanted. The experiment start date was defined as day 0. Six treatments were set up (SA; 0, 0.2, 0.4, 0.6, 0.8, and 1.0 mmol L−1). SA (Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China) was dissolved with a small amount of absolute ethanol (Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China) and diluted with distilled water to specific concentrations (final solutions contained 0.05% pure ethanol). After the SA solutions were sprayed evenly (one drop of Tween-20 was added to each liter of the solution to ensure that the SA solution was evenly attached to the leaf surface; 15 mL to each pot; Tween-20, Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China), 250 mL of NaCl solution (100, 200, 300 mmol L−1; Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China) was injected into each pot to simulate salt stress (mild, moderate, and severe). As a control, distilled water (contained 0.05% pure ethanol; 15 mL to each pot) was sprayed on the leaf surface, and 250 mL of distilled water was injected into the pots. Leaves of S. officinalis were harvested on day 30 under the above-mentioned conditions to determine various physiological and biochemical attributes.
2.2. Determination of Assays and Contents
A grade scale for salt damage index (SDI) was outlined as follows [11]. Level 1: No damage; Level 2: the leaf edge and leaf tip of old leaves turn yellow and dry, while the new leaves show no symptoms; Level 3: 1/3 to 2/3 leaves show yellowing and drying symptoms; Level 4: more than 2/3 of the leaves appear yellowing and drying symptoms; Level 5: Severe damage, almost all leaves have withered or plant death occurs (Figure 1 and Figure 2).
SDI (%) = ∑ (X × N)/(5N) × 100%

Figure 1.
The effects of salinity and salicylic acid on Saponaria officinalis.

Figure 2.
The representative images of the leaves considered for calculating the salt damage index.
X (index of each damage grade), N (number of each damage grade).
Fresh samples were placed in 20 mL of deionized water in a closed vial and incubated at 25 °C for 24 h. The conductivity of the liquid was measured with a DDS-11A conductivity meter (Shanghai INESA Scientific Instrument Co., Ltd., Shanghai, China). The conductivity of the liquid was measured again after being heated at 100 °C for 20 min.
EL (%) = (C1/C2) × 100%
C1 (electrical conductivity before heating), C2 (electrical conductivity after heating).
Fresh samples were used to determine malondialdehyde (MDA) content according to the method of thiobarbituric acid [12]. The optical density was measured with a Cary 60 UV-Vis Spectrophotometer (Agilent Technologies Inc., Santa Clara, CA, USA).
Chlorophyll (Chl) content was measured using the acetone extraction method and calculated using the optical density [12]. An amount of 0.3 g of leaves was homogenized with 10 mL of 80% pure acetone (Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China), and the homogenate was centrifuged (4 °C, 14,000 r min−1, 15 min) to obtain the supernatant solution. The absorbance at 663 and 645 nm was recorded. Photosynthesis rate (Pn) was measured with a Li-6400 Portable Photosynthesis System (LI-COR Biotechnology, Lincoln, NE, USA). The conditions in the leaf chamber included a light intensity of 1000 μmol m−2 s−1, a leaf temperature of 25 °C, the CO2 mole fraction of 450 μmol mol−1, and a flow rate of 500 mL min−1, with a relative humidity of 50–70% [13].
The quick-frozen samples were used to determinate the osmoregulatory material content. Soluble sugar (SS) content was quantified by the anthrone sulphuric acid method [14]. Soluble protein content (SP) content was quantified by the Coomassie brilliant blue method. The leaves were homogenized in 10 mL distilled water, and the supernatant was obtained after centrifugation. The supernatant was mixed with 5 mL Coomassie brilliant blue G-250 (Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China), and the change in absorbance at 595 nm was recorded. Bovine serum protein was used to obtain the standard curve by the same method [15]. Free proline (FP) content was determined by the sulfosalicylic acid method [12].
An amount of 0.3 g leaves were homogenized with 10 mL of 0.05 mol L−1 precooled phosphate buffer (pH 7.8; Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China). The homogenate was centrifuged (4 °C, 5000 r min−1, 10 min) to obtain the supernatant enzyme solution. The enzyme solution (0.05 mL) was mixed with 0.05 mol L−1 phosphate buffer (1.5 mL), 130 mmol L−1 Met (0.3 mL; Sigma-Aldrich, Shanghai, China), 750 μmol L−1 NBT (0.3 mL; Sigma-Aldrich, Shanghai, China), 100 μmol L−1 EDTA-Na2 (0.3 mL; Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China), 2.0 μmol L−1 riboflavin (0.3 mL; Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China), and distilled water (0.25 mL) to obtain a superoxide dismutase (SOD) reaction mixture. This reaction mixture was exposed to light for a color reaction, and the absorbance at 560 nm was recorded [16]. The enzyme solution (0.1 mL) was mixed with 0.05 mol L−1 phosphate buffer (1.5 mL), 0.05 mol L−1 guaiacol solution (1 mL; Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China), and 2% hydrogen peroxide (H2O2; 1 mL; Tianjin Yongda Chemical Reagent Co., Ltd., Tianjin, China) to obtain a peroxidase (POD) reaction mixture. The change in absorbance at 470 nm was recorded [15]. The enzyme solution (0.1 mL) was placed in the test tube. An amount of 0.05 mol L−1 phosphate buffer (1 mL) was added to distilled water (1.7 mL) to obtain a catalase (CAT) reaction mixture. The mixture rested at 25 °C for 3 min. Then, 200 mM H2O2 (0.2 mL) was added to the test tube. The change in absorbance at 240 nm was recorded [17]. The enzyme solution (0.1 mL) was mixed with 0.05 mol L−1 phosphate buffer (2.7 mL), 15 mmol L−1 ASA (1 mL), and H2O2 (0.1 mL) to produce the ascorbate peroxidase (APX) reaction mixture. Then, the change in absorbance at 290 nm was recorded [18].
Na+, K+, Mg2+, and Ca2+ content were determined from oven-dried leaf samples, after wet-digesting the samples in an HNO3–HClO4 acid mixture (4:1 v/v), by ICP-OES Optima 8300 (Perkins Elmer, Norwalk, CT, USA) [19].
2.3. Statistical Analysis
The statistical chart was made with Origin 2019 (OriginLab, Northampton, MA, USA). Data analysis was performed with SPSS 22.0 (Statistical Product and Service Solutions, IBM, Amonk, NY, USA). A two-way analysis and one-way analysis of variance (ANOVA) was performed, and the differences between the mean values were analyzed for significance following the Tukey method. Correlation analysis was performed by the Pearson method. Data were standardized according to the correlation analysis results. When the metric is positively correlated with the SDI, standardized data = (X − Xmin)/(Xmax − Xmin); When the metric is negatively correlated with the SDI, standardized data = (Xmax − X)/(Xmax − Xmin). Factor analysis was performed on the standardized data.
3. Results
3.1. Salt Damage
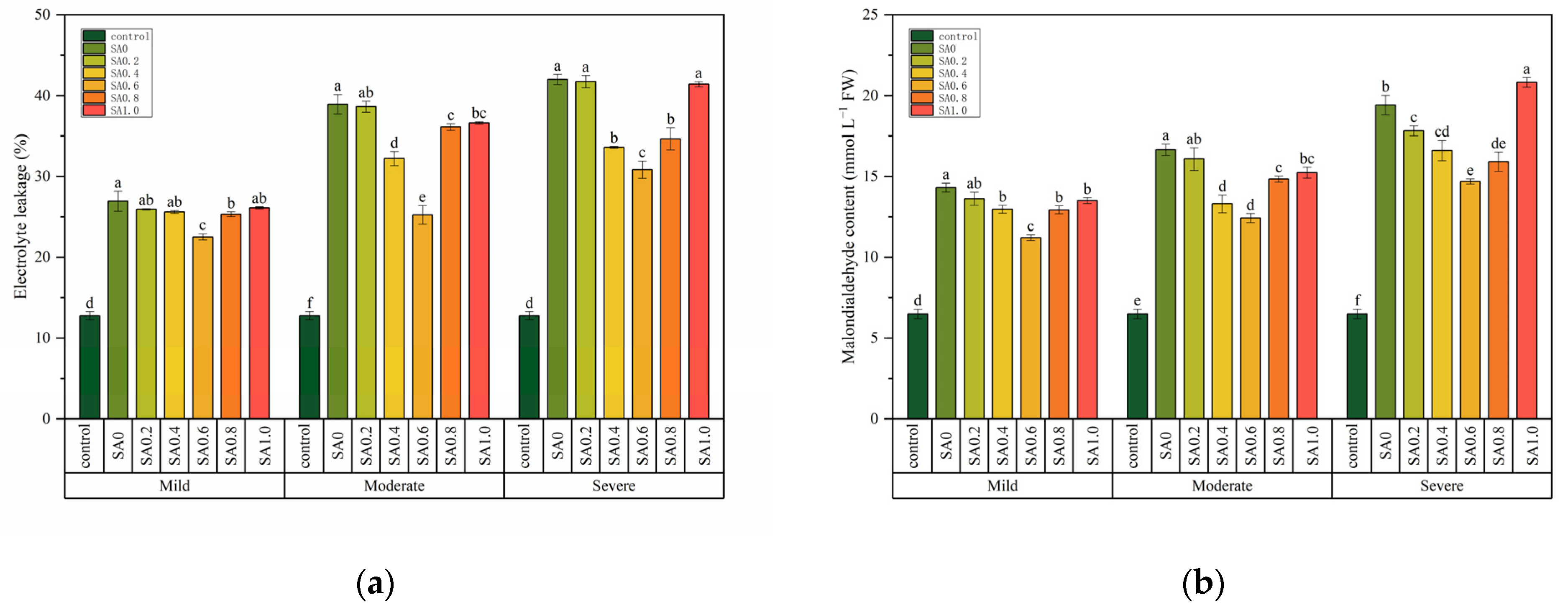
Two-way ANOVA tests indicated that the salt data index (SDI) was affected by salt stress and SA treatment but was not affected by the interaction of these two factors, while electrolyte leakage (EL) and malondialdehyde (MDA) content were affected by salt stress and SA treatment and the interaction of these two factors (Table 1). The SDI increased with the increase of salt stress. For example, the SDI increased by 75% when compared with the control under severe salt stress (Table 2). A similar trend was also found for the EL and MDA content (Figure 3). The SDI of the SA treatment were lower than SA0 at each salt stress, but the difference was insignificant (p > 0.05, Table 2). However, the EL and MDA content of SA0.6 were significantly lower than for SA0 under each salt stress (p < 0.05, Figure 3). When compared with SA0, the EL of SA0.6 decreased by 16.33, 35.20, and 26.58% respectively, and the MDA content decreased by 21.72, 25.45, and 24.40% respectively. Notably, the EL of SA0.2 and SA1.0 was not significantly different from that of SA0 (p > 0.05, Figure 3a). The MDA content of SA1.0 was even higher than that of SA0 under severe salt stress (Figure 3b).

Table 1.
Two-way analysis of variance (ANOVA) of the effects of salinity (S), salicylic acid (SA), and their interactions on the salt damage index, electrolyte leakage, malondialdehyde content, chlorophyll content, photosynthesis rate, soluble sugar content, soluble protein content, free proline content, superoxide dismutase activity, peroxidase activity, catalase activity, ascorbate peroxidase activity, Na+ content, K+ content, Mg2+ content, Ca2+ content, the and K+/Na+ ratio of Saponaria officinalis.
Table 2.
The effects of salinity and salicylic acid on the salt damage index of Saponaria officinalis.

Figure 3.
The effects of salinity and salicylic acid on the electrolyte leakage (a) and malondialdehyde content (b) of Saponaria officinalis. One-way analysis of variance (ANOVA) was used to test the differences between different treatments under mild, moderate, and severe salt stress. The differences were considered significant when p < 0.05, as indicated by the different letters (n = 3). FW, fresh weight.
3.2. Chlorophyll Content and Photosynthesis Rate
Two-way ANOVA tests indicated that the chlorophyll (Chl) content and photosynthesis rate (Pn) were affected by salt stress and SA treatments, but the interactions of these two factors were significant only for Chl (Table 1). The Chl content and Pn decreased with an increase in salt stress, and SA treatment was shown to alleviate this reduction (Figure 4). For example, the Chl of SA0.6 was increased by 14.72, 18.94, and 0.25% when compared with that of SA0 at mild, moderate, and severe salt stresses, respectively (Figure 4a). The Pn of SA0.6 was significantly higher than SA0 at mild and moderate salt stress (p > 0.05), but the difference was not significant between SA0.6 and SA0 at severe salt stress (p < 0.05, Figure 4b).
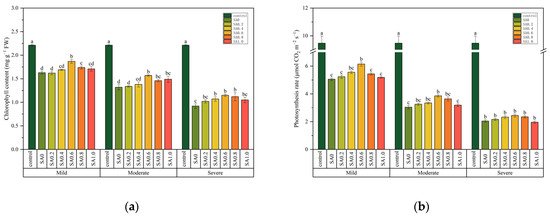
Figure 4.
The effects of salinity and salicylic acid on the chlorophyll content (a) and photosynthesis rate (b) of Saponaria officinalis. One-way analysis of variance (ANOVA) was used to test the differences between different treatments under mild, moderate, and severe salt stresses. The differences were considered significant when p < 0.05, as indicated by the different letters (n = 3). FW, fresh weight.
3.3. Osmoregulatory Material Content
Two-way ANOVA tests indicated that the soluble sugar (SS), soluble protein (SP), and free proline (FP) content were affected by salt stress and SA treatment, but the interactions of these two factors were significant only for SP and FP content (Table 1). The SS, SP and FP content increased with the increase of salt stress, and SA treatment was shown to alleviate this increase (Table 3). There were few differences between the SS content of SA treatment and that of SA0 (Table 3). However, when compared with SA0, the SP and FP content were higher. For example, the SP of SA0.6 was increased by 10.89, 5.47, and 10.94%, and the FP content was increased by 11.28, 4.66, and 8.34% when compared with SA0 (Table 3).
Table 3.
The effects of salinity and salicylic acid on the soluble sugar, soluble protein, and free proline content of Saponaria officinalis.
3.4. Antioxidase Activity
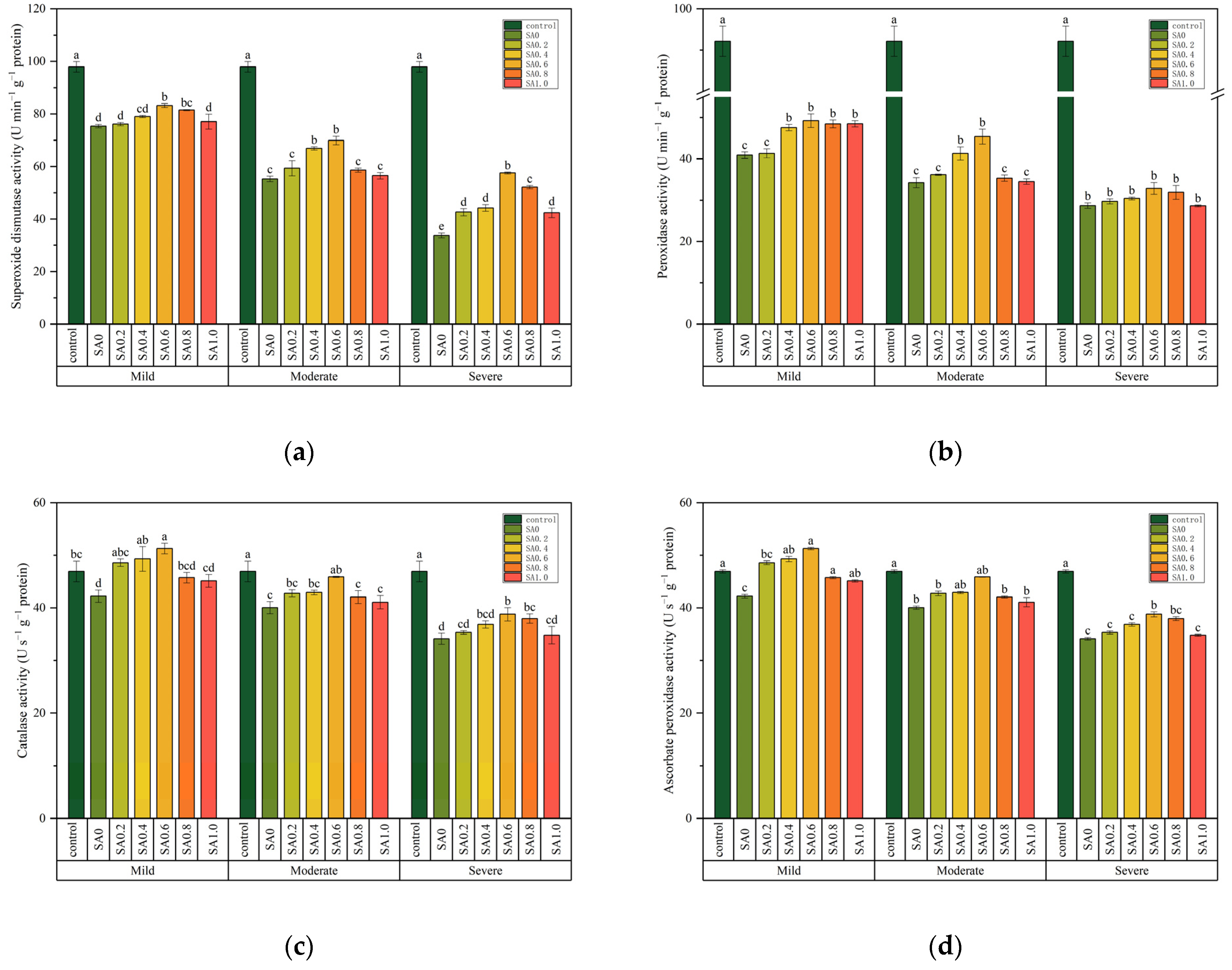
Two-way ANOVA tests indicated that superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX) activity were affected by salt stress and SA treatment and the interaction of these two factors (Table 1). The SOD, POD, CAT, and APX activity decreased with the increase in salt stress. For example, the SOD, POD, CAT, and APX activity decreased by 65.50, 68.86, 27.28, and 21.14% when compared with the control under severe salt stress, respectively (Figure 5). After application of the SA spray, the SOD, POD, CAT, and APX activity was higher than that of SA0. The SOD, POD, CAT, and APX activity of SA0.6 was significantly higher than that of SA0 at each salt stress (p < 0.05, Figure 5). However, under mild salt stress, the difference of SOD and POD activity between SA0.2, SA1.0, and SA0 did not reach a significant level ((p > 0.05, Figure 5a,b).

Figure 5.
The effects of salinity and salicylic acid on superoxide dismutase activity (a), Peroxidase activity (b), catalase activity (c), ascorbate peroxidase activity (d) of Saponaria officinalis. One-way analysis of variance (ANOVA) was used to test the differences between different treatments under mild, moderate, and severe salt stresses. The differences were considered significant when p < 0.05, as indicated by the different letters (n = 3).
3.5. Ion Homeostasis
Two-way ANOVA tests indicated that Na+, K+, Mg2+, and Ca2+ content was affected by salt stress, SA treatment, and the interaction of these two factors (Table 1). The Na+ content increased with the increase in salt stress, while K+, Mg2+, Ca2+ content, and the K+/Na+ ratio decreased with the increase in salt stress (Table 4). The K+, Mg2+, Ca2+ content, and the K+/Na+ ratio was higher than that of SA0, and the Na+ content was lower than that of SA0 after SA application. The Na+, K+, Mg2+, Ca2+ content, and K+/Na+ ratio of SA0.6 were significantly different from that of SA0 at each salt stress (p < 0.05, Table 4). However, under severe salt stress, the difference of Na+, Mg2+, Ca2+ content, and the K+/Na+ ratio between SA0.2, SA1.0, and SA0 did not reach a significant level (p > 0.05, Table 4).
Table 4.
The effects of salinity and salicylic acid on Na+ content, K+ content, Mg2+ content, Ca2+ content, and the K+/Na+ ratio of Saponaria officinalis.
3.6. Factor Analysis
We speculate that those plants with more osmolators, a higher antioxidant enzyme activity, and a stable ionic concentration suffer less from salinity under equivalent salt stress. However, we could not accurately evaluate the degree of salt damage in plants by a single indicator. For example, the SDI differences among the individual treatment groups did not reach significant levels under salt stress (Table 2). In addition, under severe salt stress, in contrast to the SA0, SA1.0 had less SDI, but a higher MDA content, less SP content, but a higher SOD activity (Table 2 and Table 3, Figure 3 and Figure 5). Therefore, we considered a comprehensive evaluation of the salt damage of each treatment group by factor analysis. The results of the factor analysis indicate that there was no significant difference between SA1.0 and SA0 under severe salt stress (p> 0.05, Table 5). Additionally, SA0.2, SA0.4, SA0.6, SA0.8, and SA1.0 were significantly different from the SA0 (p <0.05) under the three kinds (mild, moderate, and severe) of salt stress (Table 5). Moreover, the combined score of SA0.6 was the smallest among all three conditions (Table 5).
Table 5.
Comprehensive score of Saponaria officinalis in different treatment groups.
4. Discussion
Based on the extrinsic morphology of plants, a grade scale for SDI can be used to simply describe the salt damage in plants, while Pn also reflects plant physiological activities to some extent. The results achieved in this work showed that salinity makes the SDI of S. officinalis increase while SA (0.4, 0.6, 0.8 mmol L−1) treatment makes the SDI stay at relatively low levels (Table 2). The opposite trend was observed for Pn (Figure 4).The increasing of SDI and the decrease of Pn can be attributed to cellular oxidative damage and Chl decomposition (Figure 3 and Figure 4). EL and the cellular peroxidation product MDA can reflect the extent of oxidative damage in plant cells [20]. Changes in Chl content are associated with visual symptoms and photosynthetic efficiency changes during the disease incidence [21]. SA may improve the salt tolerance capacity of S. officinalis by alleviating cellular oxidative damage and avoiding Chl breakdown, since the S. officinalis treated with SA had a lower EL, lower MDA content, and higher Chl content (Figure 3 and Figure 4). Similarly, SA (100 μmol L−1) reduced the MDA content in Oryza sativa under salt conditions (200 mmol L−1) [5]. SA leaf treatment (0.5 and 1.0 mM) reduced the electrolyte leakage in Ocimum basilicum under different levels of NaCl stress (60, 120 mmol L−1) [22]. SA treatment significantly alleviated the decrease of photosynthetic pigment content in Trigonella foenum-graecum caused by salt stress [23]. Meanwhile, the application of 2 mmol L−1 SA to the leaves of Raphanus sativus significantly enhanced the levels of photosynthetic pigments under salt stress [12]. SA also promoted the Pn of Brassica juncea [24] and cucumber (Cucumis sativus cv. ‘Zhongnong 26’) [25] under salt stress. It seems no doubt that SA can alleviate oxidative damage, avoid Chl breakdown, and promote photosynthesis. Therefore, SA may improve the salt tolerance capacity of S. officinalis by modulating the photosynthetic rate.
SA may mitigate the oxidative damage of S. officinalis by maintaining high antioxidant enzyme activity (Figure 5). S. officinalis sprayed with SA (0.4, 0.6 mmol L−1) also has higher SOD, POD, CAT, and FP activity when compared with S. officinalis that is sprayed with distilled water (Figure 5). In Brassica parachinensis under salt stress, treatment with 1 mmol L−1 SA enhanced the SOD, CAT, and APX activity [26]. Additionally, SA treatment increased the SOD and POD activity and reduced lipid peroxidation levels, subsequently reducing the toxicity of salt stress in Cucumis sativus [27]. Faghih et al. found that SA enhanced the APX, POD, and SOD activity of Lycopersicon esculentum under salt stress [28]. It is worth mentioning that protective effect of SA on Chl may be achieved by enhancing the activity of APX, since APX is present in the chloroplasts (Figure 4 and Figure 5). Therefore, SA may improve the salt tolerance capacity of S. officinalis by modulating antioxidant levels.
Salinity affects plant growth and development through osmotic stress, while plants resist salt stress by accumulating osmotic regulatory substances. In this study, S. officinalis leaves accumulated a lot of Na+ under salt stress (Table 3 and Table 4). Although the accumulation of Na+ is able to increase the cellular water potential, excessive Na+ can cause ion toxicity in plants [13]. We found that the K+ content and K+/Na+ ratio of S. officinalis were decreased (Table 4), and the reduction in K+ caused by Na+ is a well-known competitive process in plant tissues [29]. Furthermore, we found that salt stress decreased the Mg2+ and Ca2+ contents in S. officinalis (Table 4). SA may maintain the ionic balance within plant leaves by limiting the excessive accumulation of Na+ (Table 4). SA promotes the absorption of K+ and Ca2+ because SA has an inhibitory effect on the excess accumulation of Na+, alleviating the competitive effect of Na+ on K+ [29], and keeping the excess Na+ from replacing Ca2+ on the membrane binding site [30]. However, the promoting effects of SA on Mg2+ need to be further studied. Studies on Mentha pulegium revealed that 1 mmol L−1 SA reduced the accumulation of Na+ in the leaves under salt stress, promoted the accumulation of K+ in leaves, and improved the K+/Na+ ratio [31]. It also found that SA contrasted the massive entry of Na+, in favor of K+, Ca2+, and Mg2+ accumulation in maize [32]. The protective effect of SA on Chl may be also related to promoting the accumulation of Mg2+, since Mg2+ is an important constituent element of Chl (Table 4). Therefore, SA may improve the salt tolerance capacity of S. officinalis by modulating ion homeostasis.
In addition to inorganic salt ions, organic matter such as SS and SP and FP have important roles in plant osmotic regulation. In this study, S. officinalis sprayed with SA (0.4, 0.6, 0.8 mmol L−1) also had higher SS, SP, and FP content when compared with the S. officinalis that was sprayed with distilled water, but the difference in the SS content was not significant (Table 3). The results show that for S. officinalis, SA mainly alleviates salt damage by regulating its accumulation of FP and SP. Similarly, 0.5 mmol L−1 SA increased the FP content of lentils under 100 mmol L−1 NaCl stress [33]. However, Studies in O. basilicum showed that foliar treatment with SA at 0.5 and 1.0 mmol L−1 concentrations under saline soil conditions (60 and 120 mM) increased the SP content but decreased the FP content [20]. The above phenomenon shows that, under salt stress, SA may alleviate osmotic damage by inducing plants to accumulate more organic matter. However, the change rules of different osmotic substances are different according to different plant species. SA may act via the accumulation of osmosis modulators, which is achieved by regulating the activity of certain enzymes. For example, SA promotes the activity of sucrose synthase and enhances the content of soluble sugars in plant organs [34]. Meanwhile, SA leaf spray improved pyrroline-5-carboxylic acid reductase activity while it inhibited the proline oxidase activity in Mentha pulegium under saline soil conditions [35]. Since FP can protect Rubisco (the key enzyme that determines carbon assimilation rate in photosynthesis), promoting the accumulation of FP is also one of the reasons why SA promotes Pn under saline stress [35]. In summary, SA may improve the salt tolerance capacity of S. officinalis by modulating osmoprotectants.
The effects of SA on plant physiological and biochemical properties vary depending on the experimental conditions, such as the intensity and stress duration and SA application method [7]. Some scholars have suggested that the effects of SA depend on the application dose; low concentrations of SA reduce plant damage while high concentrations exacerbate plant damage [7]. The absorption and utilization of SA varied based on the plant species, genotype, developmental stage, and stress strength [35]. It was found in this study that high concentrations of SA (1.0 mmol L−1) had little effect on the antioxidant enzymes of S. officinalis (Figure 5), whereas 0.1 mol L−1 SA slightly promoted APX and SOD activity and decreased the POX activity of Strawberry [28]. Moderate concentrations of SA (0.6 mmol L−1) were more effective in alleviating the harmful effects on S. officinalis under salt stress, while higher (1.0 mmol L−1) concentrations of SA were not as effective in alleviating harmful effects on S. officinalis under saline stress (Table 5). The positive effect of SA was not enhanced with its increased concentration.
5. Conclusions
Exogenous SA can effectively alleviate the harmful effect of salt stress on S. officinalis, improving the osmotic stress, oxidative damage, and ion poisoning caused by salinity. (1) SA promotes the growth of S. officinalis by modulating the photosynthetic rate; (2) SA enhances the osmoregulatory ability by promoting SP and FP synthesis in S. officinalis; (3) SA enhances the antioxidant capacity by maintaining SOD, POD, CAT, and APX activity; (4) The positive effect of SA was not enhanced with increasing concentrations. Moderate concentrations of SA (0.6 mmol L−1) are more effective in alleviating the harmful effects on S. officinalis under salt stress. Thus, the external administration of SA can be used as a feasible method to improve plant salt tolerance, but the concentration of SA needs to be standardized based on the degree of salt damage and the plant type.
Author Contributions
Data curation, H.C., T.Z. and Y.D.; Writing—original draft, L.X.; Writing—review & editing, J.Y. and L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D program of China under grant (2021YFD1500600); Fundamental Research Funds for the Central Universities under grant (2572021BK01); National Natural Science Foundation of China under grant (31800546).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jie, W.; Peng, X.; Hua, X.; Yan, X.; Zhou, Z.; Lin, J. Physiological Adaptive Strategies of Oil Seed Crop Ricinus communis Early Seedlings (Cotyledon vs. True Leaf) Under Salt and Alkali Stresses: From the Growth, Photosynthesis and Chlorophyll Fluorescence. Front. Plant Sci. 2019, 9, 1939. [Google Scholar] [CrossRef] [PubMed]
- Kulak, M.; Gul, F.; Sekeroglu, N. Changes in growth parameter and essential oil composition of sage (Salvia officinalis L.) leaves in response to various salt stresses. Ind. Crops Prod. 2020, 145, 112078. [Google Scholar] [CrossRef]
- Smulek, W.; Zdarta, A.; Pacholak, A.; Zgola-Grzeskowiak, A.; Marczak, L.; Jarzebski, M.; Kaczorek, E. Saponaria officinalis L. extract: Surface active properties and impact on environmental bacterial strains. Colloids Surf. B-Biointerfaces 2017, 150, 209–215. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M.; Tran, L.-S.P. Nitric oxide mediates hydrogen peroxide- and salicylic acid-induced salt tolerance in rice (Oryza sativa L.) seedlings. Plant Growth Regul. 2015, 77, 265–277. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M. Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by up-regulating antioxidative and glyoxalase systems. Ecotoxicology 2013, 22, 959–973. [Google Scholar] [CrossRef]
- Horvath, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Irfan, M.; Ahmad, A. Effect of exogenous salicylic acid under changing environment: A review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Abdoli, S.; Ghassemi-Golezani, K.; Alizadeh-Salteh, S. Responses of ajowan (Trachyspermum ammi L.) to exogenous salicylic acid and iron oxide nanoparticles under salt stress. Environ. Sci. Pollut. Res. 2020, 27, 36939–36953. [Google Scholar] [CrossRef]
- Oueslati, S.; Karray-Bouraoui, N.; Attia, H.; Rabhi, M.; Ksouri, R.; Lachaal, M. Physiological and antioxidant responses of Mentha pulegium (Pennyroyal) to salt stress. Acta Physiol. Plant. 2010, 32, 289–296. [Google Scholar] [CrossRef]
- Zhong, Y.-p.; Qi, X.-j.; Chen, J.-y.; Li, Z.; Bai, D.-f.; Wei, C.-g.; Fang, J.-b. Growth and physiological responses of four kiwifruit genotypes to salt stress and resistance evaluation. J. Integr. Agric. 2019, 18, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Bukhat, S.; Manzoor, H.; Athar, H.-u.-R.; Zafar, Z.U.; Azeem, F.; Rasul, S. Salicylic Acid Induced Photosynthetic Adaptability of Raphanus sativus to Salt Stress is Associated with Antioxidant Capacity. J. Plant Growth Regul. 2020, 39, 809–822. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Y.; Peng, X.; Yan, J.; Yan, X.; Zhou, Z.; Lin, J. Cotyledon removal decreases salt tolerance during seedling establishment of Ricinus communis, an oilseed energy crop species. Ind. Crops Prod. 2019, 142, 111857. [Google Scholar] [CrossRef]
- Bai, J.-h.; Liu, J.-h.; Zhang, N.; Yang, J.-h.; Sa, R.-l.; Wu, L. Effect of Alkali Stress on Soluble Sugar, Antioxidant Enzymes and Yield of Oat. J. Integr. Agric. 2013, 12, 1441–1449. [Google Scholar] [CrossRef]
- Tuna, A.L.; Kaya, C.; Dikilitas, M.; Yokas, I.; Burun, B.; Altunlu, H. Comparative effects of various salicylic acid derivatives on key growth parameters and some enzyme activities in salinity stressed maize (Zea mays L.) plants. Pak. J. Bot. 2007, 39, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Seckin, B.; Sekmen, A.H.; Turkan, I. An Enhancing Effect of Exogenous Mannitol on the Antioxidant Enzyme Activities in Roots of Wheat Under Salt Stress. J. Plant Growth Regul. 2009, 28, 12–20. [Google Scholar] [CrossRef]
- Nakano, Y.N.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Souana, K.; Taibi, K.; Abderrahim, L.A.; Amirat, M.; Achir, M.; Boussaid, M.; Mulet, J.M. Salt-tolerance in Vicia faba L. is mitigated by the capacity of salicylic acid to improve photosynthesis and antioxidant response. Sci. Hortic. 2020, 273, 109641. [Google Scholar] [CrossRef]
- Gunes, A.; Inal, A.; Alpaslan, M.; Eraslan, F.; Bagci, E.G.; Cicek, N. Salicylic acid induced changes on some physiological parameters symptomatic for oxidative stress and mineral nutrition in maize (Zea mays L.) grown under salinity. J. Plant Physiol. 2007, 164, 728–736. [Google Scholar] [CrossRef]
- Selem, E.E.; Hamed, R.E.A.; Kamel, H.A.; Hegazy, H.S. Physiological and Biochemical Response of Gamma Irradiated Sesamum indicum L. Seed Grown in Heavy Metal Contaminated Soil. Biosci. Res. 2018, 15, 1063–1072. [Google Scholar]
- Elhindi, K.M.; Al-Amri, S.M.; Abdel-Salam, E.M.; Al-Suhaibani, N.A. Effectiveness of salicylic acid in mitigating salt-induced adverse effects on different physio-biochemical attributes in sweet basil (Ocimum basilicum L.). J. Plant Nutr. 2017, 40, 908–919. [Google Scholar] [CrossRef]
- Abdelhameed, R.E.; Abdel Latef, A.A.H.; Shehata, R.S. Physiological Responses of Salinized Fenugreek (Trigonella foenum-graecum L.) Plants to Foliar Application of Salicylic Acid. Plants 2021, 10, 657. [Google Scholar] [CrossRef]
- Nazar, R.; Umar, S.; Khan, N.A. Exogenous salicylic acid improves photosynthesis and growth through increase in ascorbate-glutathione metabolism and S assimilation in mustard under salt stress. Plant Signal. Behav. 2015, 10, e1003751. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; Luo, X.; Gao, X.; Wang, W.; Li, B.; Hou, L. Exogenous salicylic acid alleviates salt stress by improving leaf photosynthesis and root system architecture in cucumber seedlings. Sci. Hortic. 2020, 272, 109577. [Google Scholar] [CrossRef]
- Kamran, M.; Xie, K.; Sun, J.; Wang, D.; Shi, C.; Lu, Y.; Gu, W.; Xu, P. Modulation of growth performance and coordinated induction of ascorbate-glutathione and methylglyoxal detoxification systems by salicylic acid mitigates salt toxicity in choysum (Brassica parachinensis L.). Ecotoxicol. Environ. Saf. 2020, 188, 109877. [Google Scholar] [CrossRef]
- Shi, Q.; Zhu, Z. Effects of exogenous salicylic acid on manganese toxicity, element contents and antioxidative system in cucumber. Environ. Exp. Bot. 2008, 63, 317–326. [Google Scholar] [CrossRef]
- Faghih, S.; Ghobadi, C.; Zarei, A. Response of Strawberry Plant cv. ‘Camarosa’ to Salicylic Acid and Methyl Jasmonate Application Under Salt Stress Condition. J. Plant Growth Regul. 2017, 36, 651–659. [Google Scholar] [CrossRef]
- Botella, M.A.; Martinez, V.; Pardines, J.; Cerda, A. Salinity induced potassium deficiency in maize plants. J. Plant Physiol. 1997, 150, 200–205. [Google Scholar] [CrossRef]
- Cachorro, P.; Ortiz, A.; Cerda, A. Implications of Calcium Nutrition on the Response of Phaseolus vulgaris L. to Salinity. Plant Soil 1994, 159, 205–212. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Farhadi, N. The Efficacy of Salicylic Acid Levels on Photosynthetic Activity, Growth, and Essential Oil Content and Composition of Pennyroyal Plants Under Salt Stress. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Sultan, I.; Khan, I.; Chattha, M.U.; Hassan, M.U.; Barbanti, L.; Calone, R.; Ali, M.; Majid, S.; Ghani, M.A.; Batool, M.; et al. Improved salinity tolerance in early growth stage of maize through salicylic acid foliar application. Ital. J. Agron. 2021, 16, 1810. [Google Scholar] [CrossRef]
- Misra, N.; Saxena, P. Effect of salicylic acid on proline metabolism in lentil grown under salinity stress. Plant Sci. 2009, 177, 181–189. [Google Scholar] [CrossRef]
- Sha, H.; Liu, H.; Wang, J.; Jia, Y.; Wang, X.; Zou, D.; Zhao, H. Physiological mechanism of salicylic acid regulating salt tolerance of crops. J. Northeast. Agric. Univ. 2017, 48, 80–88. [Google Scholar] [CrossRef]
- Farhadi, N.; Ghassemi-Golezani, K. Physiological changes of Mentha pulegium in response to exogenous salicylic acid under salinity. Sci. Hortic. 2020, 267, 109325. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).