Mathematics of the Relationship between Plant Population and Individual Production of Maize (Zea mays L.)

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Statistical Analyses
2.4. Simulation Study
3. Results
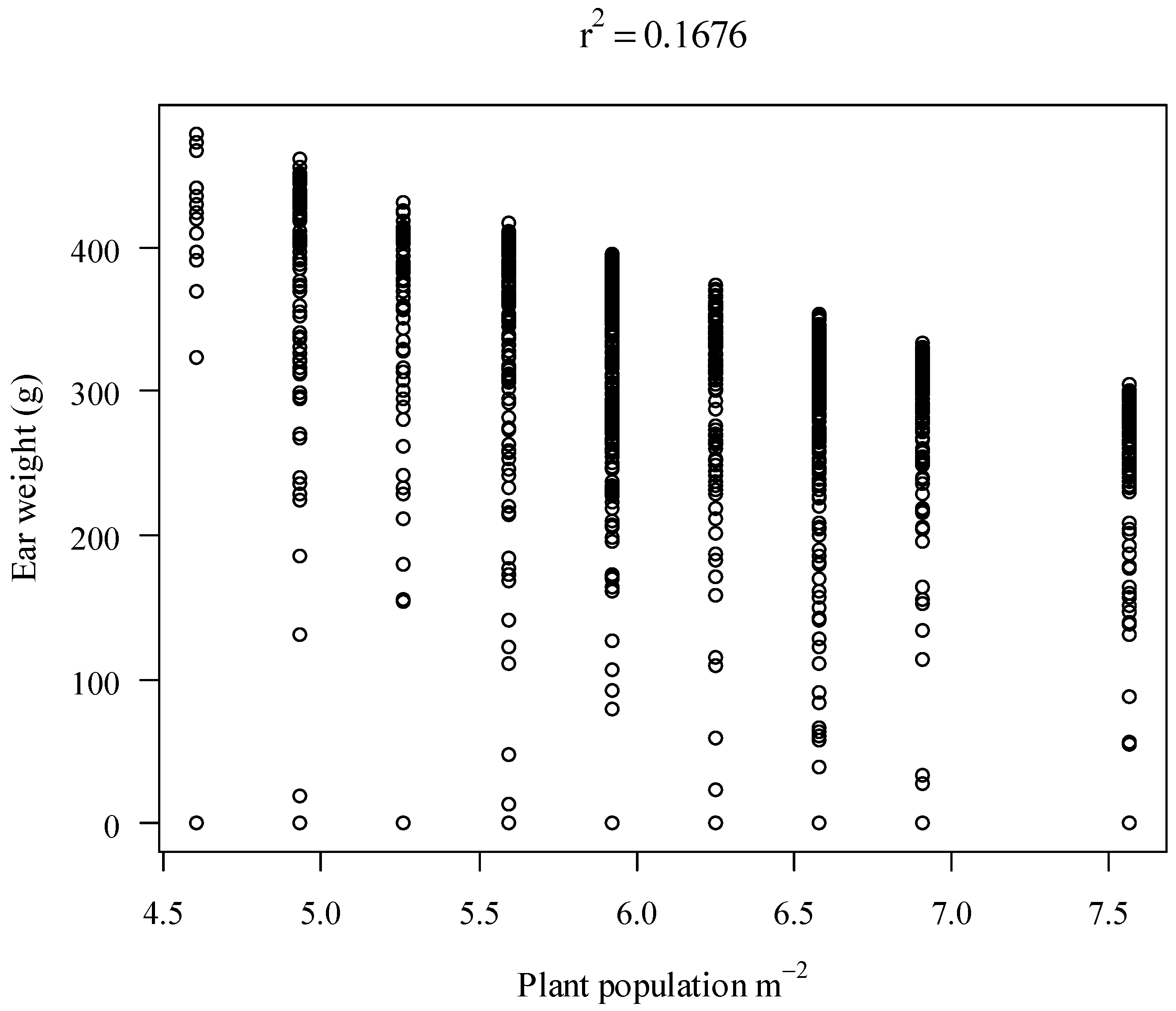
3.1. Regression Analysis for Different Mosaic Sizes within a Plot
3.2. Distributions of the Variables
3.3. Results of Simulation Modelling and Model Validation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organisation of the United Nations. Methodology for Estimation of Crop Area and Crop Yield under Mixed and Continuous Cropping. Tech. Rep. 2017, 5, 36–58. Available online: http://www.fao.org/3/ca6514en/ca6514en.pdf (accessed on 3 May 2021).
- Food and Agriculture Organisation of the United Nations. Handbook on Crop Statistics: Improving Methods for Measuring Crop Area and Production and Yield. Tech. Rep. 2018, 6, 67–79. Available online: https://www.fao.org/3/ca6408en/ca6408en.pdf (accessed on 3 May 2021).
- Tandzi, L.N.; Mutengwa, C.S. Estimation of maize (Zea mays L.) yield per harvest area: Appropriate methods. Agronomy 2020, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Assefa, Y.; Carter, P.; Hinds, M.; Bhalla, G.; Schon, R.; Jeschke, M.; Paszkiewicz, S.; Smith, S.; Ciampitti, I.A. Analysis of long term study indicates both agronomic optimal plant density and increase maize yield per plant contributed to yield gain. Sci. Rep. 2018, 8, 4937. [Google Scholar] [CrossRef] [Green Version]
- Ipsilandis, C.G.; Vafias, B.N. Plant density effects on grain yield per plant in maize: Breeding implications. Asian J. Plant Sci. 2005, 4, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Holliday, R. Plant density and crop grain yield. Nature 1960, 186, 22–24. [Google Scholar] [CrossRef]
- Tokatlidis, I.S.; Koutroubas, S.D. A review of maize hybrids’ dependence on high plant population and its implications for crop yield stability. Field Crops Res. 2004, 88, 103–114. [Google Scholar] [CrossRef]
- Haarhoff, S.J.; Swanepoel, P.A. Plant population and maize grain yield: A global systematic review of rainfed trials. Crop Sci. 2018, 58, 1819–1829. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Hara, T. On the relative yield of plants in two-species mixture. Oikos 1999, 85, 170–176. [Google Scholar] [CrossRef]
- Van Roekel, R.J.; Coulter, J.A. Agronomic responses of corn to planting date and plant density. Agron. J. 2011, 103, 1414–1422. [Google Scholar] [CrossRef]
- Assefa, Y.; Prasad, P.V.V.; Carter, P.; Hinds, M.; Bhalla, G.; Schon, R.; Jeschke, M.; Paszkiewicz, S.; Ciampitti, I.A. Yield responses to planting density for US modern corn hybrids: A synthesis-analysis. Crop Sci. 2016, 56, 2802–2817. [Google Scholar] [CrossRef]
- Mead, R. Plant density and crop yield. J. R. Stat. Soc. Ser. C (Appl. Stat.) 1970, 19, 64–81. Available online: www.jstor.org/stable/2346843 (accessed on 3 May 2021).
- Testa, G.; Reyneri, A.; Blandino, M. Maize grain yield enhancement through high plant density cultivation with different inter-row and intra-row spacing. Eur. J. Agron. 2016, 72, 28–37. [Google Scholar] [CrossRef]
- Sangoi, L.; Gracietti, M.A.; Rampazzo, C.; Bianchetti, P. Response of Brazilian maize hybrids from different eras to changes in plant density. Field Crop Res. 2002, 79, 39–51. [Google Scholar] [CrossRef]
- Qian, C.; Yu, Y.; Gong, X.; Jiang, Y.; Zhao, Y.; Yang, Z.; Hao, Y.; Li, L.; Song, Z.; Zhang, W. Response of grain yield to plant density and nitrogen rate in spring maize hybrids released from 1970 to 2010 in Northeast China. Crop J. 2016, 4, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Zhai, L.-C.; Xie, R.-Z.; Ming, B.; Li, S.-K.; Ma, D.-L. Evaluation and analysis of intraspecific competition in maize: A case study on plant density experiment. J. Integr. Agric. 2018, 17, 2235–2244. [Google Scholar] [CrossRef] [Green Version]
- Cazanga, R.; Ledent, J.F.; von Bennewith, E. Adaptation of CERES-Maize model for a better simulation of plant population density effects. Field Crop Res. 2019, 238, 27–34. [Google Scholar] [CrossRef]
- Sida, T.S.; Chamberlin, J.; Ayalew, H.; Kosmowski, F.; Craufurd, P. Implications of intra-plot heterogeneity for yield estimation accuracy: Evidence from smallholder maize systems in Ethiopia. Field Crops Res. 2021, 267, 108147. [Google Scholar] [CrossRef]
- Rossini, M.A.; Maddoni, G.A.; Otegui, M.E. Inter-plant competition for resources in maize crops grown under contrasting nitrogen supply and density: Variability in plant and ear growth. Field Crops Res. 2011, 121, 373–380. [Google Scholar] [CrossRef]
- Madonni, G.A.; Otegui, M.E. Intra-specific competition in maize: Early establishment of hierarchies among plant effects final kernel set. Field Crops Res. 2004, 85, 1–13. [Google Scholar] [CrossRef]
- Duncan, W.G. The relationship between corn population and yield. Agron. J. 1958, 50, 82–84. [Google Scholar] [CrossRef]
- Carmer, S.G.; Jackobs, J.A. An exponential model for predicting optimum plant density and maximum corn yield. Agron. J. 1965, 57, 241–244. [Google Scholar] [CrossRef]
- Farazdaghi, H.; Harris, P.M. Plant competition and crop yield. Nature 1968, 217, 289–290. [Google Scholar] [CrossRef]
- Willey, R.W.; Heath, S.B. The quantitative relationships between plant population and crop yield. Adv. Agron. 1969, 21, 281–321. [Google Scholar] [CrossRef]
- Tang, L.; Ma, W.; Noor, M.A.; Li, L.; Hou, H.; Zhang, X.; Zhao, M. Density resistance evaluation of maize varieties through new “Density–Yield Model” and quantification of varietal response to gradual planting density pressure. Sci. Rep. 2018, 8, 17281. [Google Scholar] [CrossRef]
- Shirzadifar, A.; Maharlooei, M.; Bajwa, S.G.; Oduor, P.G.; Nowatzki, J.F. Mapping crop stand count and planting uniformity using high resolution imagery in a maize crop. Biosyst. Eng. 2020, 200, 377–390. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2017 | 2018 | |||
|---|---|---|---|---|
| Row Number | Number of Plants | Mean Weight of Ear ± SD, g/Plant | Number of Plants | Mean Weight of Ear ± SD, g/Plant |
| 1 | 90 | 338.0 ± 49.2 | 92 | 351.0 ± 57.8 |
| 2 | 88 | 313.2 ± 61.0 | 93 | 341.9 ± 54.2 |
| 3 | 85 | 350.2 ± 61.8 | 94 | 318.8 ± 60.4 |
| 4 | 79 | 352.8 ± 58.8 | 99 | 293.6 ± 74.9 |
| 5 | 87 | 355.5 ± 70.3 | 96 | 298.4 ± 73.0 |
| 6 | 75 | 338.9 ± 78.7 | 99 | 331.2 ± 49.2 |
| 7 | 83 | 346.9 ± 67.8 | 98 | 294.2 ± 81.7 |
| 8 | 83 | 330.0 ± 53.4 | 103 | 311.1 ± 65.5 |
| 9 | 79 | 310.8 ± 57.7 | 101 | 307.8 ± 56.1 |
| 10 | 84 | 316.3 ± 64.8 | 98 | 322.0 ± 76.3 |
| 11 | 82 | 309.6 ± 79.7 | 93 | 323.8 ± 59.5 |
| 12 | 84 | 305.7 ± 70.3 | 94 | 343.6 ± 72.9 |
| Code | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| Number of row | 2 | 2 | 2 | 2 | 2 | 4 | 4 | 4 | 4 | 4 |
| Length (m) | 1 | 2 | 4 | 5 | 10 | 1 | 2 | 4 | 5 | 10 |
| Design Code | Number of Subplots | Area of a Subplot, m2 | RSE | r2 | p-Value | SE for the Number of Plants | SE for Ear Weight |
|---|---|---|---|---|---|---|---|
| 1 | – | – | 65.6 | 0.043 | <0.001 | 72.5 * | 67.0 * |
| 2 | 120 | 1.5 | 26.7 | 0.161 | <0.001 | 14.0 | 29.0 |
| 3 | 60 | 3.0 | 22.8 | 0.175 | <0.001 | 19.3 | 24.9 |
| 4 | 30 | 6.0 | 20.2 | 0.197 | 0.014 | 29.4 | 22.2 |
| 5 | 24 | 7.5 | 18.9 | 0.207 | 0.026 | 34.6 | 20.8 |
| 6 | 12 | 15.0 | 18.4 | 0.129 | 0.252 | 39.6 | 18.8 |
| 7 | 60 | 3.0 | 22.5 | 0.151 | 0.002 | 19.9 | 24.2 |
| 8 | 30 | 6.0 | 20.3 | 0.127 | 0.053 | 28.1 | 21.4 |
| 9 | 15 | 12.0 | 18.4 | 0.128 | 0.191 | 39.4 | 19.0 |
| 10 | 12 | 15.0 | 18.3 | 0.083 | 0.365 | 49.9 | 18.2 |
| 11 | 6 | 30.0 | 19.6 | 0.000 | 0.904 | 41.8 | 17.2 |
| Design Code | SD | CV | p | n | ||
|---|---|---|---|---|---|---|
| 2 | 8.5 | 1.372 | 0.161 | 0.221 | 0.8 | 11 |
| 3 | 17.0 | 1.832 | 0.108 | 0.197 | 0.8 | 22 |
| 4 | 34.0 | 2.828 | 0.083 | 0.235 | 0.8 | 44 |
| 5 | 42.5 | 3.349 | 0.079 | 0.264 | 0.8 | 53 |
| 6 | 85.0 | 3.790 | 0.045 | 0.169 | 0.8 | 106 |
| Design Code | SD | CV | ||
|---|---|---|---|---|
| 2 | 8.5 | 1.372 | 0.161 | 0.221 |
| 3 | 17.0 | 2.744 | 0.161 | 0.443 |
| 4 | 34.0 | 5.488 | 0.161 | 0.886 |
| 5 | 42.5 | 6.860 | 0.161 | 1.107 |
| 6 | 85.0 | 13.720 | 0.161 | 2.215 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huzsvai, L.; Bodnár, E.; Kovács, E.; Zsembeli, J.; Harsányi, E.; Juhász, C.; Szőke, S. Mathematics of the Relationship between Plant Population and Individual Production of Maize (Zea mays L.). Agronomy 2022, 12, 1602. https://doi.org/10.3390/agronomy12071602
Huzsvai L, Bodnár E, Kovács E, Zsembeli J, Harsányi E, Juhász C, Szőke S. Mathematics of the Relationship between Plant Population and Individual Production of Maize (Zea mays L.). Agronomy. 2022; 12(7):1602. https://doi.org/10.3390/agronomy12071602
Chicago/Turabian StyleHuzsvai, László, Emil Bodnár, Elza Kovács, József Zsembeli, Endre Harsányi, Csaba Juhász, and Szilvia Szőke. 2022. "Mathematics of the Relationship between Plant Population and Individual Production of Maize (Zea mays L.)" Agronomy 12, no. 7: 1602. https://doi.org/10.3390/agronomy12071602