Abstract
Gibberellins (GAs), a pivotal plant hormone, play fundamental roles in plant development, growth, and stress response. In rice, gibberellin-dioxygenases (GAoxes) are involved in the biosynthesis and deactivation of gibberellins. However, a comprehensive genome-wide analysis of GA oxidases in rice was not uncovered. Here, a total of 80 candidate OsGAox genes were identified and 19 OsGAox genes were further analyzed. Studies on those 19 OsGAox genes, including phylogenetic tree construction, analysis of gene structure, exploration of conserved motifs and expression patterns, were conducted. Results showed that the GAox genes in Arabidopsis and rice were divided into four subgroups and shared some common features. Analysis of gene structure and conserved motifs revealed that splicing phase and motifs were well conserved during the evolution of GAox genes in Arabidopsis and rice, but some special conserved motifs possessed unknown functions need to be further studied. Exploration of expression profiles from RNA-seq data indicated that each GAox gene had tissue-specific expression patterns, although they varied greatly. The expression patterns of these genes under GA3 treatment revealed that some genes, such as OsGA2ox1, OsGA2ox3, OsGA2ox4, OsGA2ox7, OsGA20ox1, and OsGA20ox4, may play a major role in regulating the level of bioactive GA. Taken together, our study provides a comprehensive analysis of the GAox gene family and will facilitate further studies on their roles in rice growth and development so that these genes can be better exploited.
1. Introduction
Bioactive gibberellins (GAs), a large group of diterpene plant hormones, play essential roles in the complete life cycle of rice, such as seed germination, stem elongation, root development, pollen development, and flower induction [1,2,3,4,5]. To date, about 136 GAs have been identified in higher plants, fungi, and bacteria [6], while most of them are non-bioactive and act as precursors for the bioactive forms or deactivated metabolites and only GA1, GA3, GA4, and GA7 are bioactive GAs [7]. In higher plants, the GA metabolism pathway has been extensively studied [8]. This pathway mainly involves three stages of reactions, according to the location and the enzymes involved. In the first stage, geranylgeranyl diphosphate (GGDP), a common C20 precursor for diterpenoids, is converted to the tetracyclic hydrocarbon intermediate ent-kaurene by two kinds of terpene synthases (TPSs) in the plastids, ent-copalyl diphosphate synthase (CPS) and ent-kaurene synthase (KS). In the second stage, GA12 and GA53 are synthesized from ent-kaurene by two types of cytochrome P450 monooxygenases (P450s) at the endoplasmic reticulum, ent-Kaurene oxidase (KO), and entkaurenoic acid oxidase (KAO). In the final stage of the pathway, bioactive GA synthesis is catalyzed by two kinds of soluble 2-oxoglutarate-dependent dioxygenases (2ODDs) known as GA 20-oxidase (GA20ox) and GA 3-oxidase (GA3ox) in the cytosol and the bioactive GAs or their immediate precursors are inactivated by a third 2ODD, GA 2-oxidase (GA2ox), including C19-GA2oxes and C20-GA2oxes [9].
The GA20ox, GA3ox, and GA2ox belong to the 2-ODDs superfamily and are each encoded by a multigene family [10], the members of which have different expression patterns and thus regulate GA metabolism in different plant developmental processes. Most GAox gene family members have been cloned and identified and their biological functions have also been studied in various plant species [11,12]. By manipulating the expression of GAox genes, the levels of endogenous active GAs can be regulated. For example, the deficiency of a rice semidwarfing gene (sd-1/OsGA20ox2) known as the “green revolution gene” causes reduction in endogenous GAs, thus affecting the plant height in rice [13]. Therefore, it is extremely important to identify and exploit GAox genes in plants.
Rice (Oryza sativa L.), one of the most important global food crops, is a primary source of food for over half of the world’s population [14]. Historically, the first green revolution resulted from the utilization of GAox genes and fertilizer greatly increased the production of crops, especially rice and wheat [3]. However, considering the deterioration of the environment caused by the use of fertilizer and the increasing of population, rice production should be increased more quickly via more scientific strategies. Grain yield in rice is a complex trait affected by multiple factors and major progress in increasing rice yield is on the basis of the exploitation of high yield varieties. Due to the key role that GA plays in regulating the yield of the rice, a lot of high yield varieties have been exploited by regulating genes involved in the GA biosynthetic pathways in rice [15,16,17]. In order to better manipulate the GAox genes to attain high yield varieties, further study of the GAox family genes needs to be conducted.
Here, we conducted a comprehensive genome-wide analysis of GAox genes in rice and further studied 19 OsGAox genes, including analysis of the phylogenetic relationship, gene structure, and motif identification. Furthermore, we investigated the gene expression profiles in different rice organs and alteration under GA3 treatment. These data suggested OsGAox genes had special expression patterns in various organs and played different roles under GA3 treatment in rice. This study will serve as a foundation into a comprehensive and deep study of GAox genes in future research.
2. Materials and Methods
2.1. Plant Materials and Treatments
Rice (Oryza sativa L. japonica cv. Nipponbare) seeds were grown in sterile water in conical flasks at 37 °C in an incubator. After 3 days, the seeds were germinated and transferred into hydroponic boxes. Seedlings were cultivated in greenhouse, and watered with nutrient solution every day; 7 days later, some of the seedlings were transferred into a paddy at Wuhan University under natural conditions, while the rest of them were still cultivated in greenhouse. The temperature of the greenhouse was 27 °C, and the relative humidity was 80%. Plant materials for analyzing expression patterns in different periods included: (i) stigma, ovary and root; (ii) young leaves, mature leaves; (iii) different developmental stages of panicles and seeds, which were sampled according to the rice growth stages in the paddy. For the GA3 treatment, the 14-day seedlings of rice cultivated in greenhouse were treated with stress condition: 100 μM GA3 (CAS:77-06-5, Sigma-Aldrich, St. Louis, MO, USA), which was dissolved in anhydrous ethanol and mixed with nutrient solution. For the Mock treatment, the 14-day seedlings of rice cultivated in greenhouse were still cultured with nutrient solution. At the same time as the GA3 treatment, the Mock treatment was also carried out, and samples were taken at the same time. During the time period of GA3 and Mock treatment, the leaves were collected at 0, 2, 4, 6, 8, 10, and 12 h after treatment and quickly frozen in liquid nitrogen for RNA extraction.
2.2. Sequence Retrieval and Identification and Analysis of GAox Genes in Rice
All sequences of 21 GAox genes were downloaded from Phytozome (The Plant Comparative Genomics portal of the Department of Energy’s Joint Genome Institute) [18], which was described in a previous report [17]. Finally, SMART (available online: http://smart.emblheidelberg.de/ (accessed on 15 October 2020)) were employed to further confirm the existence of both 2OG-FeII_Oxy (PF03171) and DIOX_N (PF14226) domains in all identified GAox gene proteins. All GAox genes’ loci and their information, including chromosome location, were searched in the RGAP and DAP-DB (the Rice Annotation Project Database). The map of genes distributed across chromosomes was created with Mapchart software.
2.3. Analysis of Phylogenetic Relationship and Sequences
Multiple sequence alignments of 19 OsGAox proteins from rice and 16 AtGAox proteins from Arabidopsis were performed by using the iTOL website [19]. A phylogenetic tree was conducted using the neighbor-joining (NJ) method with 1000 bootstrap replicates in TreeBeST (available online: https://github.com/Ensembl/treebest (accessed on 22 September 2021)) [20]. For gene structure analysis, the genomic DNA sequences with the corresponding cDNA sequences from the RADP and RAP-DB database, including the exons and introns structures of individual GAox genes, were displayed via the Gene Structure Display Server (available online: http://gsds.cbi.pku.edu.cn/ (accessed on 7 May 2021)) [21]. Conserved motifs analysis was performed by MEME analysis tool with the maximum number of motifs to identify set to 10 [22].
2.4. Analysis of GA Oxidase Family Gene Expression Patterns
To study expression patterns of OsGAox genes, the public RNA-seq data contained a wide range of rice developmental stages, downloaded from Rice Functional Genomic Express Database (available online: http://signal.salk.edu/cgi-bin/RiceGE (accessed on 23 March 2021)). Subsequently, the heat map was constructed with Heml software with the log-transformed values by reanalyzing the RNA-seq data [23].
2.5. RNA Extraction, qRT-PCR and RNA seq
Total RNA was isolated from collected samples using TRIzol reagent (Takara, Japan) and then treated with DNase I (New England Biolabs, Beijing, China) according to the manufacturers’ instructions. The RNA concentration was measured using RNA Nano6000 Assay Kit of the Bioanalyzer 2000 system (Thermo Scientific, Thermo Fisher Scientific Inc.1.6.198, Waltham, MA, USA). Approximate 1 μg of total RNA were reverse-transcribed using ABScript Ⅲ RT Master Mix for qPCR with gDNA Remover (ABclonal; cat.no.RK20429). Reverse transcription polymerase chain reaction (RT-PCR) was performed using the Hieff qPCR SYBR Green Master Mix (YEASEN; cat.no.11201ES08) in a CFX96 Real-Time System (BIO-RAD, CFX Manager Software 3.1, Hercules, CA 94547, USA). Specific primers (Supplementary Table S3) for qRT-PCR were designed by using the software Primer Premier 5 and synthesized by TSINGKE (Wuhan, China), and were then checked with the cDNA by PCR. The amplification length for each gene was restricted to 80–250 bp to ensure the efficiency of optimal polymerization. Quantitative real-time PCR (qRT-PCR) was performed with 1 μL cDNA, 0.4 μM gene specific primers, and 10 μL 2× mix (YEASEN), and water was used to supplement to 20 μL by a CFX96 Real-Time System (BIO-RAD, CFX Manager Software 3.1, Hercules, CA 94547, USA) according to the manufacturer’s instruction. The reaction program of qRT-PCR was performed under the following conditions: 95 °C for 5 min, followed by 40 cycles at 95 °C for 10 s, and 60 °C for 30 s. Three replicates were carried out for each sample and three biological replicates were also performed for each sample. The relative gene expression levels were calculated using a 2−ΔΔCt method [24] and the melting curve was carried out for each PCR product to avoid nonspecific amplification. The rice gene actin (LOC_Os03g50885) was used as an internal control to normalize the expression of related genes involved in GAs biosynthesis.
2.6. Statistical Analysis
All data were analyzed using the GraphPad Prism 7.00 statistics program (https://www.graphpad.com/, accessed on 23 March 2021) and the means were compared by Student’s t-test. Each assay was performed in three biological replicates and technical replications.
3. Results
3.1. Identification and Analysis of the GAox Family Genes in the Rice
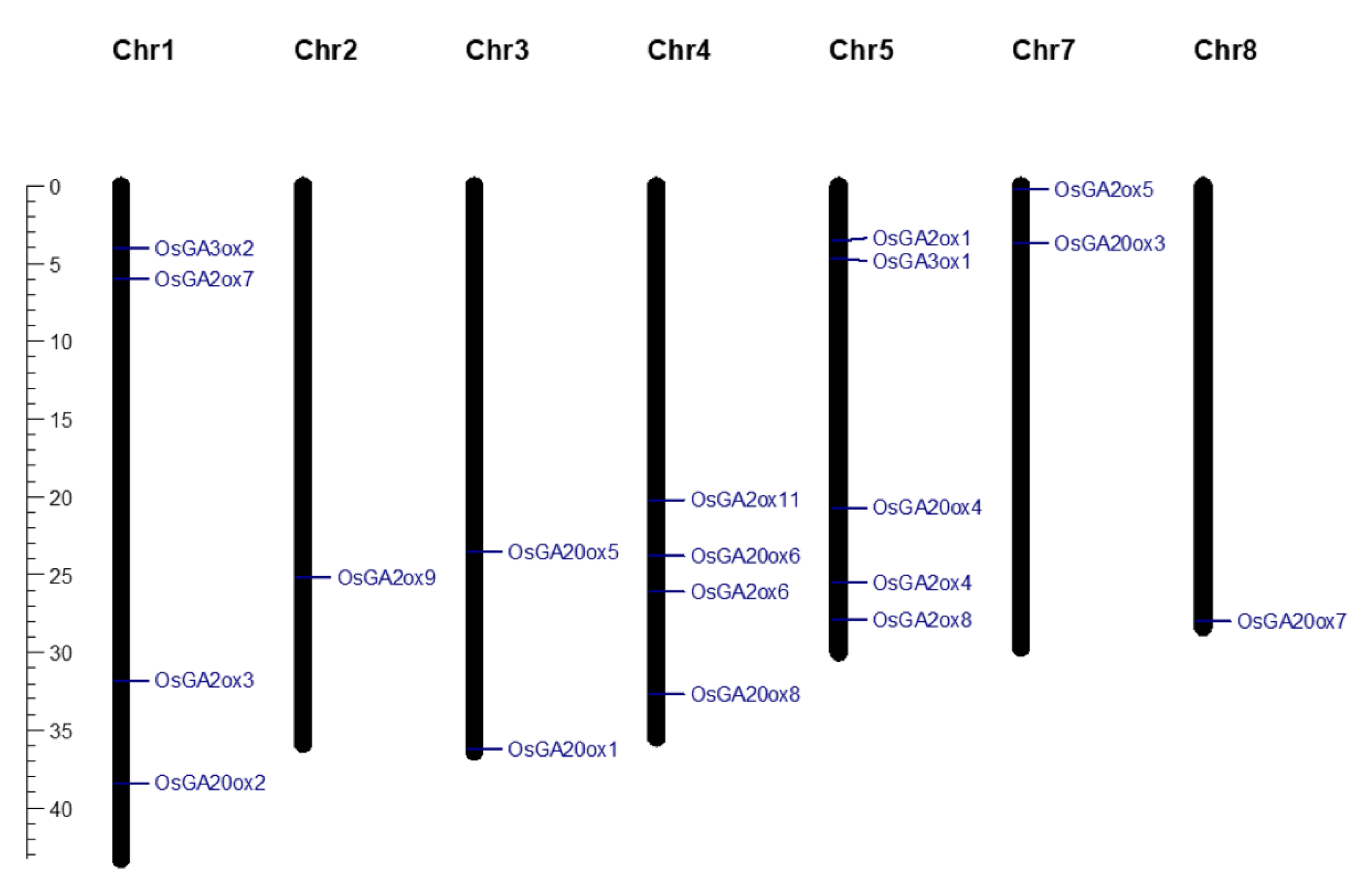
With genome wide analysis, we identified a total of 80 candidate OsGAox genes based on genome and transcriptome databases (Supplementary Table S1). A previous study reported about 21 OsGAox genes in rice [25]. Based on the results of alignment, two genes (OsGA2ox2 and OsGA2ox10) lacked a common domain DIOX_N (PF14226) were excluded in this study. To better understand the distribution of rice GAox genes on chromosomes, the map of genes distributed across chromosomes was created with Mapchart software (Figure 1). Our study showed that the 19 OsGAox genes were unevenly distributed on seven chromosomes. There were four OsGAox genes on both chromosome 1 and chromosome 4. Five OsGAox genes were located on chromosome 5. Two OsGAox genes were mapped on both chromosome 2 and chromosome 7. Only one OsGAox gene was found on chromosome 2 and chromosome 8. Interestingly, this result showed undiscovered GA biosynthesis genes in the other five chromosomes. The gene names, entry ID, number of deduced amino acid, molecular weights, predicted subcellular localizations, group classifications, and theoretical pI were also summarized in Table 1. Data showed that the protein length of the identified GAox family genes ranged from 301 (OsGA20ox6) to 446 (OsGA20ox4) amino acids (aa), with an average of 363 aa. The molecular mass ranged from 32.10 to 47.63 kDa, and the pI ranged from 5.25 (OsGA20ox8) to 7.44 (OsGA2ox6). Most of GAox family genes were predicted to be located in the nucleus and cytoplasm on WoLFPSORT [26] and TargetP [27], which was consistent with the previous findings [15,28]. Results also suggested some of them can be transported into mitochondria or chloroplasts, implied that these organelles might be also involved in the GA metabolism in plants.
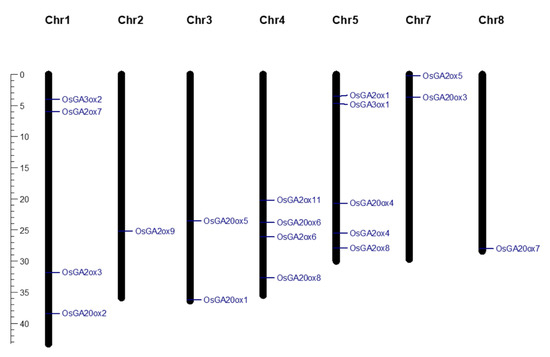
Figure 1.
The distribution of 19 GAox genes on rice chromosomes. The scale bar on the left represented the length (Mb) of rice chromosomes.

Table 1.
The 19 OsGAox family genes in the rice.
3.2. Phylogenetic Analysis of the GAox Gene Family
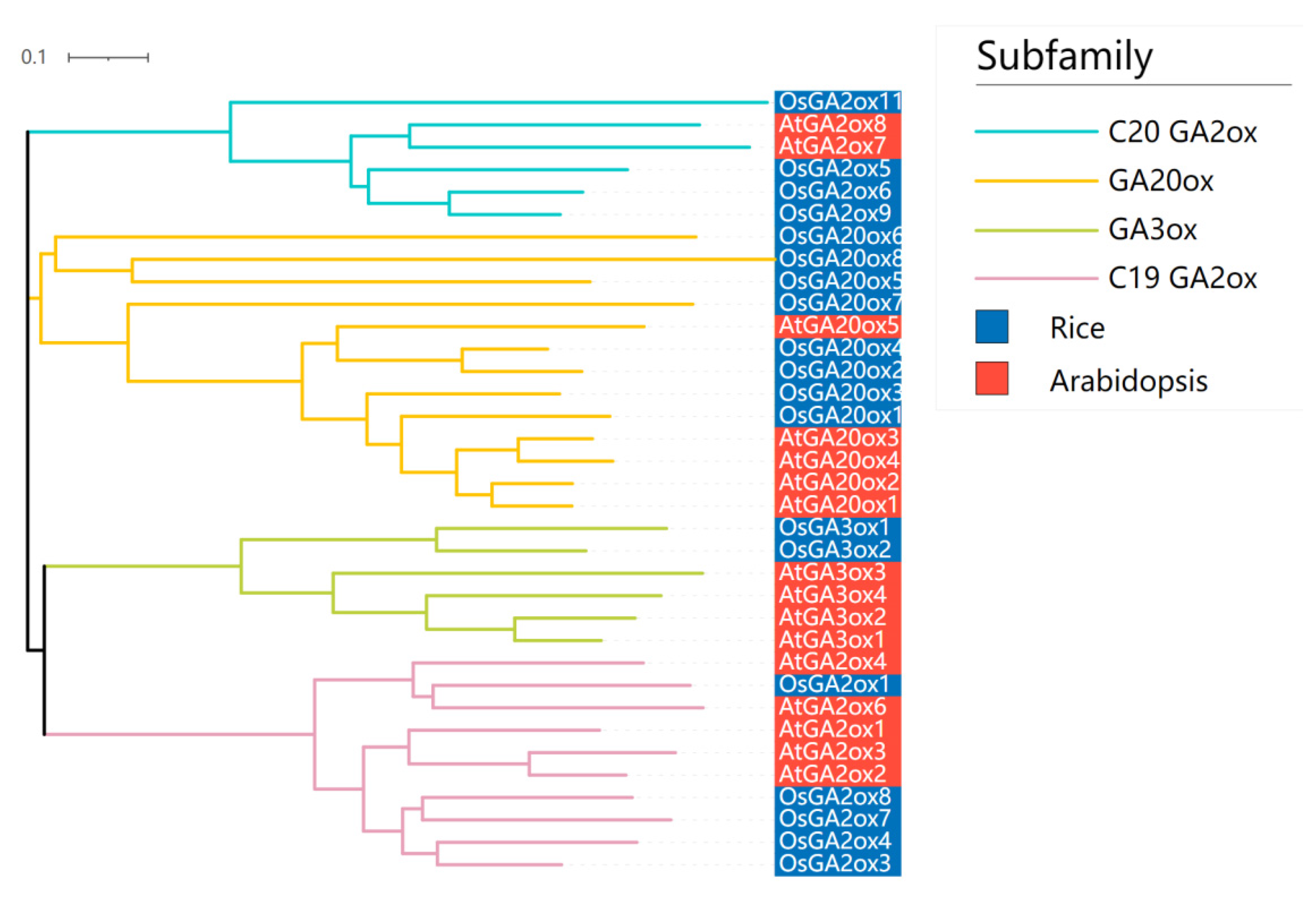
In Arabidopsis, sixteen GAox genes (seven GA2oxes, four GA3oxes and five GA20oxes) have been identified [9]. In order to determine evolutionary relationships of rice and Arabidopsis GAoxes, the phylogenetic tree was constructed using the neighbor-joining (NJ) method from alignments of the complete protein sequences from 16 AtGAox and 19 OsGAox (Figure 2). The phylogenetic tree generated four distinct subgroups, and also revealed that the phylogenetic representation of Arabidopsis and rice GAox proteins was diverse. Among the 35 proteins, five OsGAox and five AtGAox belonged to C19-GA2ox subfamily, eight OsGAox and five AtGAox belonged to GA20ox subfamily, two OsGAox and four AtGAox belonged to GA3ox subfamily, and four OsGAox and two AtGAox belonged to C20GA2ox subfamily. Four subfamilies (GA20ox, GA3ox, C19GA2ox, and C20GA2ox) were shared in both two species, suggesting that these four subfamilies might be widespread in plant GA metabolism. Furthermore, the numbers of GA20ox and GA2ox genes were greater than GA3ox in both two species, indicating that GA20ox and GA2ox had undergone a more dynamic evolutionary route than GA3ox and thus resulted in more functional redundancy. Overall, the GAox genes shared some common characteristics in monocots’ and dicots’ evolutionary relationship, so the related studies on them could interact and put each other forward.
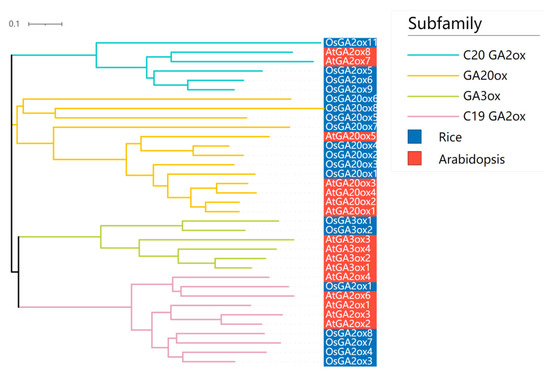
Figure 2.
Phylogenetic relationships in AtGAox and OsGAox genes. The phylogenetic tree was constructed based on the complete protein sequences of 16 AtGAox and 19 OsGAox proteins using MEGA 7.0 software. Arabidopsis and rice GAoxs were labeled by red and blue, respectively.
3.3. Gene Structure and Conserved Motif Analysis of GAox Gene Family
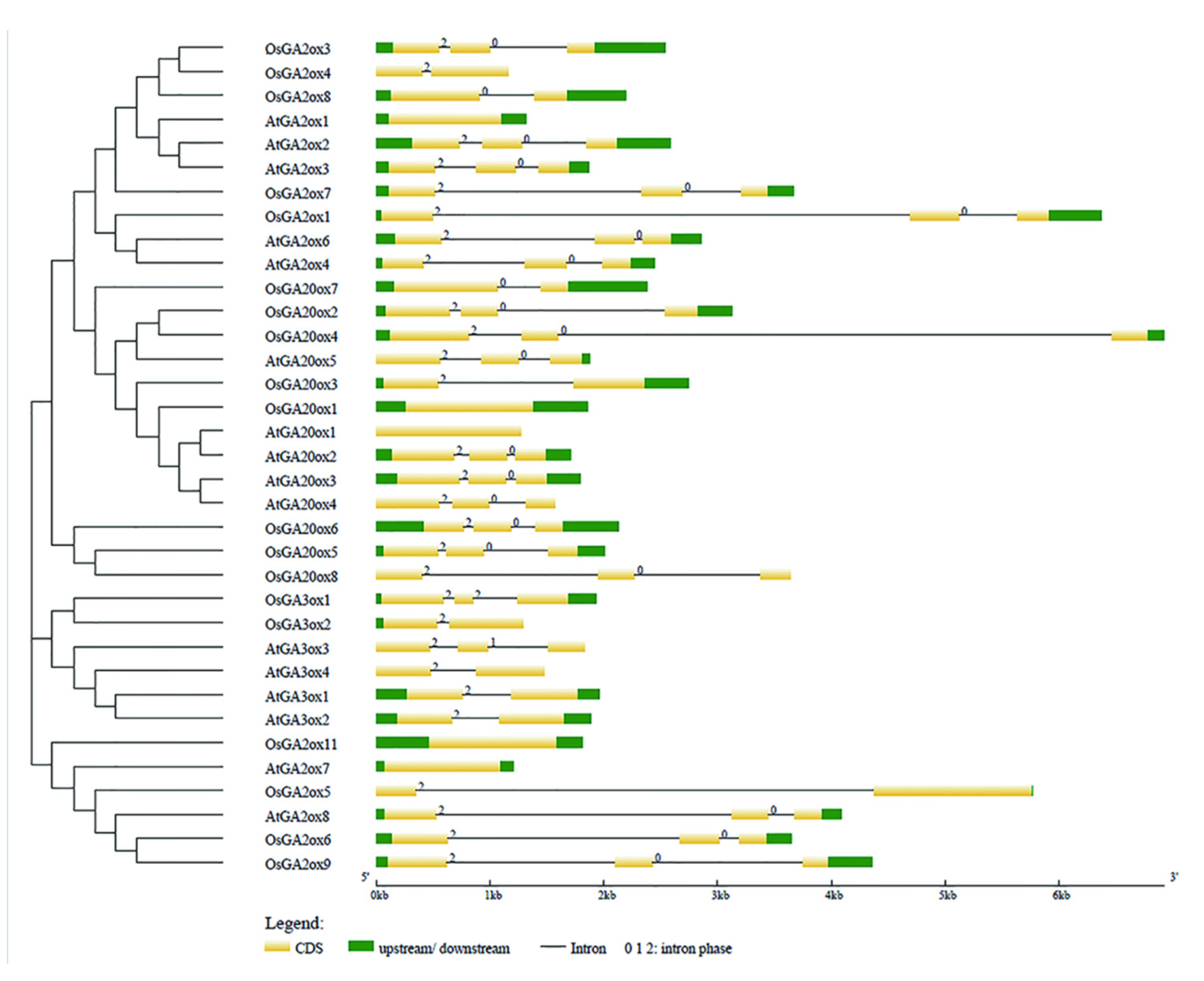
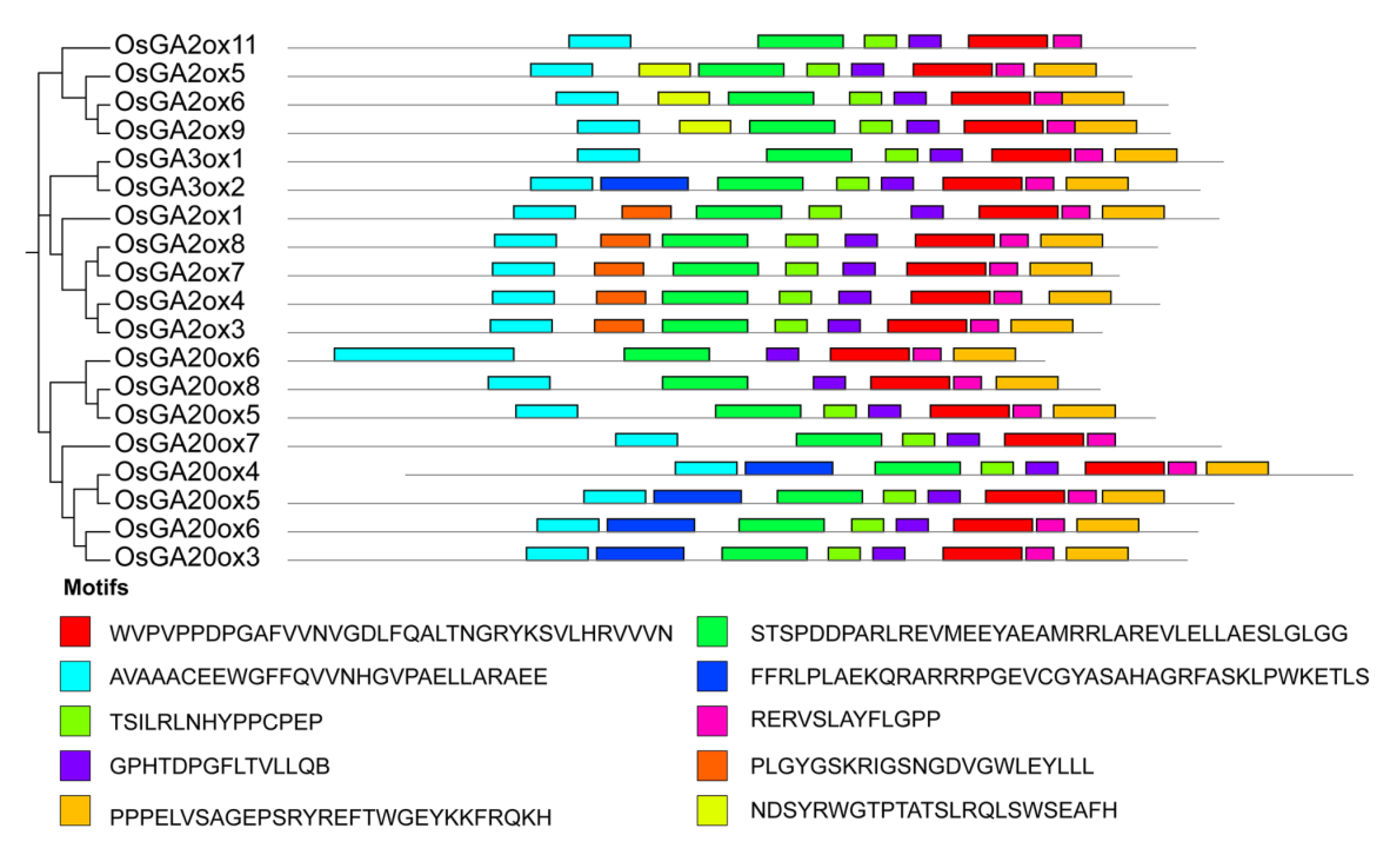
To support the phylogenetic analysis, we performed gene structure analysis of GAox family members from Arabidopsis and rice. As shown in Figure 3, the the number of exons was conserved, ranging from 1 to 3 exons in AtGAox and OsGAox genes. We also investigated intron phases with respect to codons. Most of the first intron was a phase 2 intron which meant that splicing occurred after the second nucleotide and the second intron is generally a phase 0 intron, which meant that splicing occurred after the third nucleotide. This result revealed that the splicing phase was also well conserved during the evolution of GAox genes in Arabidopsis and rice. Generally, the distribution is unequal in intron phases with a bias in favor of phase 0, which indicated that the ancient introns were dominantly of phase 0, so as to favor intron average length of GA oxidase genes in rice shuffling [29,30,31]. In order to better understand the structure of the GAox gene family, we further analyzed the cis-acting elements contained in the first 2000 bp of the 19 OsGAox genes (Figure S1). Most genes have roughly the same cis-acting elements, but different cis-acting elements are distributed in different positions of each gene, which may provide help for studying how to regulate GAox gene expression. To further investigate the relationship among GAox proteins with the same subfamily in rice, 10 different conserved motifs were identified and examined by using the MEME motif search tool in rice (Figure S2 and Figure 4). Motifs 1, 2, 3, 4, 5, 6, and 8 were shared by most of the GAox proteins. Interestingly, there were some specific absent motifs in specific subfamilies. Motif 7 was absent in all the members of C19-GA2ox except for GA2ox8 and motif 10 was shared by all C20-GA2ox and GA3ox. Motif 9 was only shared by proteins of the C19-GA2ox subfamily, which revealed that motif 9 may have a special function in C19-GA2ox subfamily. Differences among motif distributions explained sources of functional divergence in GA oxidases in evolutionary history. However, the function of these motifs in these proteins needed to be investigated further.
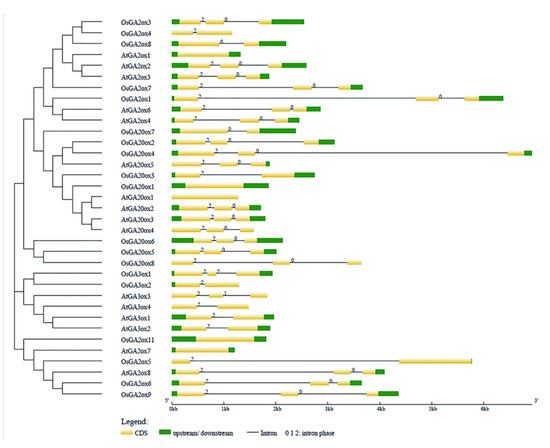
Figure 3.
Exon-intron structure of AtGAoxes and OsGAoxes. Exons and introns are shown by filled yellow boxes and thin black lines, respectively. UTRs are displayed by filled green boxes. Intron phases 0, 1, and 2 are indicated by numbers 0, 1, and 2.
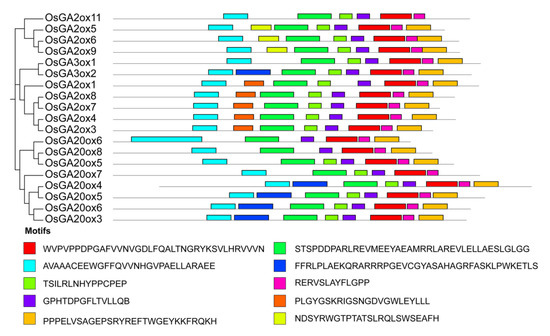
Figure 4.
Motif identification of OsGAox proteins. Different motifs are marked in numbers 1–10. The sequence logos and E values for each motif are given in Figure S2.
3.4. Expression Patterns of GAox Genes
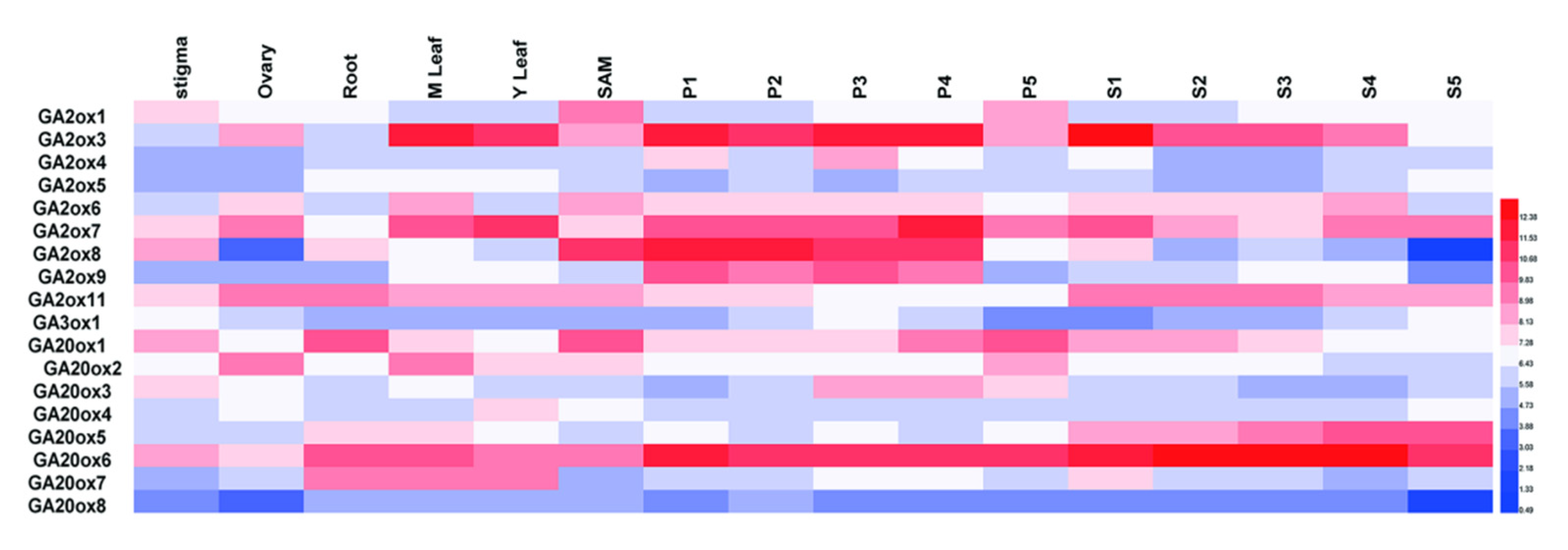
To provide clues for functional studies of GAox genes, we used FPKM values to represent their expression profiles in different organs of rice in this study [32]. Due to the lack of the corresponding probe of OsGA3ox2, we analyzed expression patterns of the other 18 genes (Supplementary Table S2). Our results showed that the OsGAox genes had different expression levels in various organs (Figure 5). It was worthy to note that some OsGAox genes, especially OsGA2ox3, OsGA2ox7, OsGA2ox8, and OsGA20ox6, were highly expressed in panicle and OsGA2ox7 and OsGA20ox6 had a high expression in all organs; however, OsGA20ox8 had a low expression in all organs. Based on the above results, we verified the expression pattern of these five genes from RNA-seq data by qRT-PCR. As shown the Supplementary Figure S3, those genes basically had the same pattern compared to RNA-seq results. Overall, those results represented that each OsGAox gene possessed special expression pattern in various organs, thus promoting the study on potential functions of these genes in different developmental stages of rice.
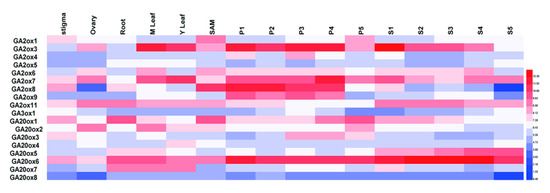
Figure 5.
Tissue-specific expression of OsGAoxes in various tissues. The expression values were normalized with logarithm with the base of 2 using the HemI software. The color bar in heat maps represents the expression values: blue represents low expression, white represents no significant difference in expression, and red means high expression. M Leaf: mature leaf; Y Leaf: young leaf; SAM means shoot apical meristem; P1 means young inflorescence of up to 3 cm; P2 means Inflorescence of 3–5 cm; P3 means Inflorescence of 5–10 cm; P4 means Inflorescence of 10–15 cm; P5 means 15–22 cm; S1 means seed of 0–2 DAP (Days After Pollination); S2 means seed of 3–4 DAP; S3 means seed of 5–10 DAP; S4 means seed of 11–20 DAP; S5 means seed of 21–29 DAP.
3.5. GAox Gene Expression Profiles under GA3 Treatment
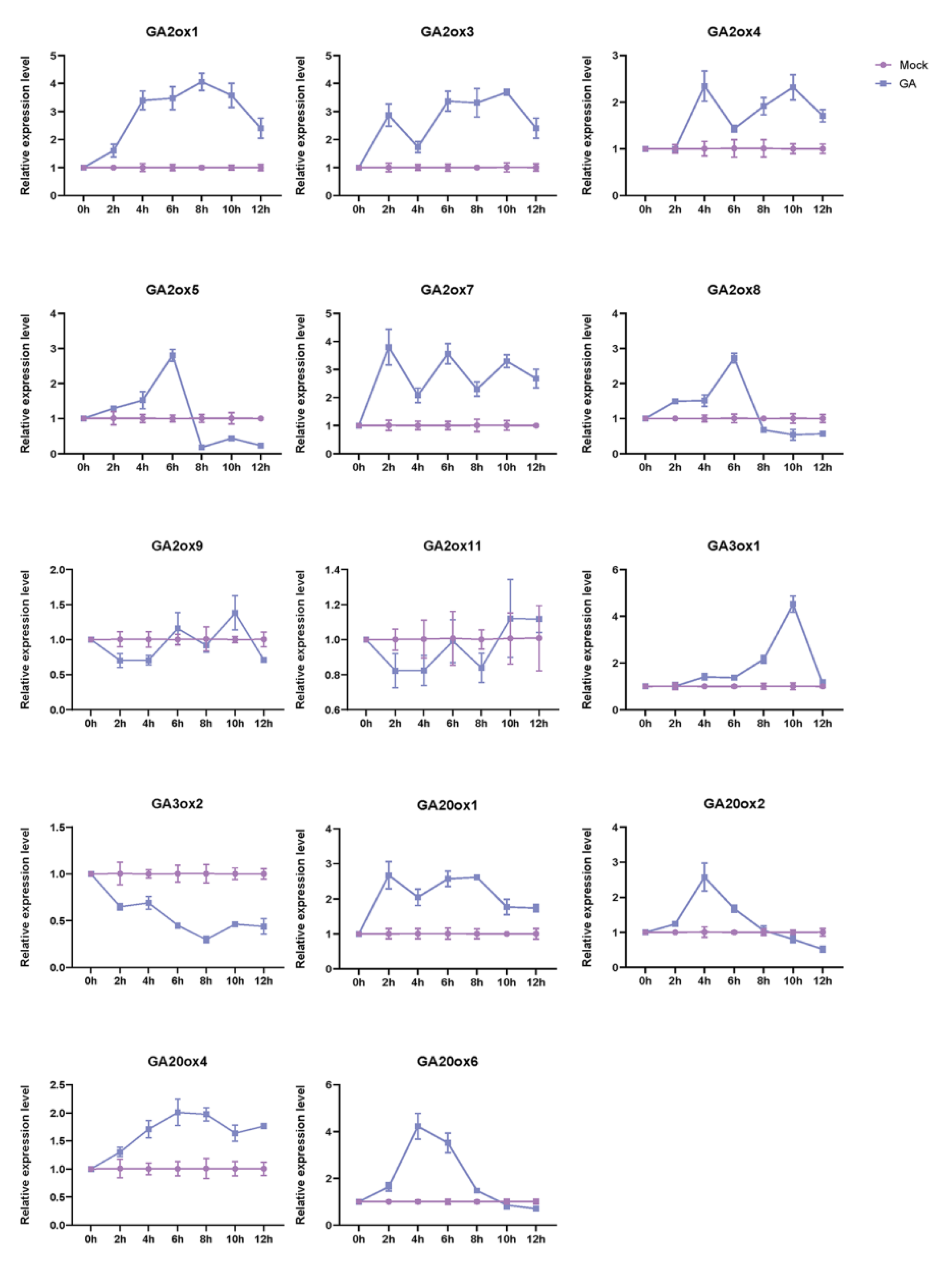
To explore the manner of these 19 OsGAox genes in response to exogenous GA, we treated the 14-day rice seedlings with GA3, then took the leaves of rice seedlings in different time periods and analyzed the expression profiles of 19 OsGAox genes under GA3 treatment by qRT-PCR. As expected, most of them exhibited different expression alteration under GA3 treatment (Figure 6). Results showed that the expression levels of OsGA2ox1, OsGA2ox3, OsGA2ox4, OsGA2ox7, OsGA20ox1, and OsGA20ox4 exhibited an upward trend after GA3 treatment, while the expression levels of OsGA2ox5, OsGA2ox8, OsGA3ox1, OsGA20ox2, and OsGA20ox6 increased first and then decreased. Furthermore, while the expression level of OsGA3ox2 was suppressed, the expression of OsGA2ox9 and OsGA2ox11 had no significant change. For the remaining GAox gene family members, because their Ct value is greater than 35, the test results cannot quantitatively analyze the gene expression, so they are not released. Overall, those results indicated that not all OsGAox genes were involved in the gibberellin homeostasis; some OsGAox genes are the main regulatory genes, but some genes which were not involved in the gibberellin homeostasis may play a role in other aspects of rice developmental progress.
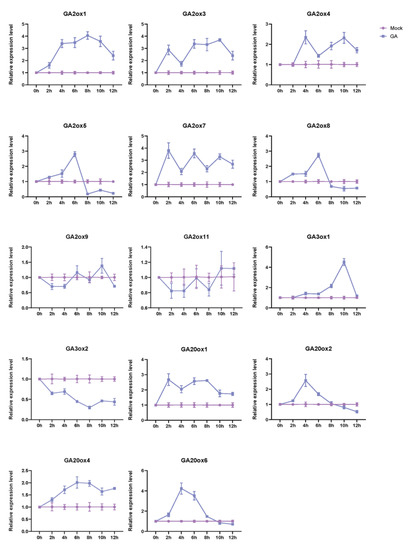
Figure 6.
The expression profiles of 19 OsGAox genes under GA3 treatment in 14-day rice seedlings. Mock represents controls (non-treated plants), GA represents GA3 treatment. The relative gene expression levels were calculated using the 2−ΔΔCt method and expressed as the fold change relative to expression of the Mock.
4. Discussion
The 2-ODDs superfamily is a large family that has been identified in many land plants, especially in crops, such as cucumber, soybean, and rice [25,33]. Previous research showed that 21 OsGAox genes have been named in rice [25], but most of researchers majorly focused on the cloning, function analysis, and molecular mechanism of one certain OsGAox gene in rice. Here, we identified 80 OsGAox genes in the rice genome and further analyzed 19 OsGAox genes with both 2OG-FeII_Oxy (PF03171) and DIOX_N(PF14226) domains for summarizing their common characteristics. The studies included phylogenetic tree construction, analysis of gene structure and conserved motifs, investigation of their expression patterns and exploration of their function of regulating the level of bioactive GA.
In our evolutionary analysis, GAox proteins family in Arabidopsis and rice can be divided into four subfamilies based on their protein sequences. Phylogenetic analysis revealed distinct differences between the two species, such as the number of each subfamily (Figure 2). In some studies, there was another subfamily in some species, GA7ox [33,34,35]. GA7ox, which oxidizes GA12-aldehyde to GA12 and possesses mono-oxygenase 7-oxidase activity, was reported in pumpkin and cucumber but has not been found in other species. So far, although three GA2ox, GA3ox, and GA20ox subfamilies have been found in most plant species, the identification of the GAox gene in other plant species still needs to be performed for finding GA7ox or the other subfamily so that we can have a more comprehensive understanding of the GAox genes’ evolutionary relationship.
In the present study, gene structure revealed that OsGAox and AtGAox gene structures were conserved and the ancient introns in rice were dominantly of phase 0 to favor intron average length of GAox genes in rice shuffling. Conserved motif analysis of the GAox proteins in rice revealed that most of motifs consisted in all GAox, while a few motifs were only possessed by a certain subfamily. To better understand the function of GAox genes, those special motifs’ biological functions are waiting for further investigating and exploring.
In this study, the expression patterns of 19 OsGAox genes were observed in different rice organs. The result showed that expression levels of these genes varied greatly in different organs. Four genes, OsGA2ox3, OsGA2ox7, OsGA2ox8, and OsGA20ox6, were highly expressed in panicle, indicating that they may play a key role in panicle development. OsGA20ox6 and OsGA20ox8 had a higher and lower expression level in all organs, respectively, revealing that they may have different roles in all developmental processes of rice. Gene expression patterns can generate fundamental new insights into their biological function in organisms in general. The OsGA20ox6 gene, for example, is essential for reproductive development, including anther dehiscence, pollen fertility, and seed initiation in rice [36], which is consistent with its expression patterns in various organs. From these results, the different expression levels in all rice organs indicated that certain OsGAox genes might play important roles in plant development and have unique functions in specific developmental stages. Now many researchers are cloning new GAox genes and exploring their functions, so our results can facilitate their research.
The levels of bioactive GAs in plants are maintained via feedback and feedforward regulation of GA metabolism [37], so almost all OsGAox genes expression level changed in response to GA3 and Mock treatment. We also examined OsGAox gene expression patterns in rice seedling under GA3 and Mock treatment. Notably, most OsGAox genes showed distinct changes after treatment. The expression levels of some genes were increased after GA3 treatment, but some of them were increased first and then decreased, which indicated that there might be feedback regulation to inhibit the expression of these genes after GA3 treatment for a period of time. OsGA3ox2 has been inhibited, which indicates that it may play a negative regulatory role in the GA synthesis pathway. Nevertheless, OsGA2ox9 and OsGA2ox11 have no significant change, which indicates that they may not participate in the response to exogenous GA. However, the other genes presented unexpected expression patterns which were inconsistent with our outstanding of the GA feedback and feedforward regulation (Figure 6). We speculated that some genes in the rice GAox gene family may maintain linkage relationships during the evolution process, and this linkage may have some advantages in selection under certain conditions, so we further analyzed the collinearity of these 19 OsGAox genes (Figure S4), which provided useful clues for studying the relationship between gene functions. GAs regulate plant growth and development and are closely related to the yield of other crops [1,2,3,4,5], and GA oxidases play pivotal roles in GAs’ biosynthesis and metabolism [7], but how to coordinate the regulation mechanism between the two has not yet been clear. Therefore, how all the GAox genes combine and coordinate to regulate the bioactive GA level should be further studied in the future. In addition, considering that the feasibility of genetic improvement of rice yield by manipulating the expression of OsGAox genes [38], our results can be for reference to improve the rice yield by changing one certain OsGAox gene expression.
5. Conclusions
In this study, we comprehensively analyzed 19 OsGAox genes in rice, which can be divided into four subgroups according to phylogenetic tree. Gene structure and conserved motif analysis showed that most GAox genes were conserved in two model plants, Arabidopsis and rice. We also analyzed their expression profiles in different organs in rice. The result suggested that various OsGAox genes played different roles in rice developmental stages. In addition, the expression patterns of those genes under GA3 treatment were also explored and from the result we found out not all OsGAox genes were involved in the gibberellin homeostasis. Taken together, our data will generate insight into the further study of OsGAox genes in rice and provide reference for exploitation of certain GAox genes to improve the rice yield and food security.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12071627/s1. Figure S1. The cis-acting elements contained in the −2000 bp of the 19 OsGAox genes. Figure S2. The sequence logos and E values for 10 conserved motifs among OsGAox proteins. Figure S3. Validation of expression patterns of 5 OsGAox genes by qRT-PCT. Figure S4. The collinearity analysis of the 19 OsGAox genes in rice. Table S1. 80 candidate OsGAox genes. Table S2. FPKM values of OsGAox genes in rice different organs. Table S3. Primers used in this study.
Author Contributions
J.H. (Jun Hu) designed the research; Y.H., W.L., Z.H., Q.Z., J.H. (Jishuai Huang) and Y.X. performed the molecular biology experiments; Z.H. performed the bioinformatics analyses; Y.H., W.L. and J.H. (Jun Hu) wrote the manuscript, and all authors read and approved it. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31871592), the Creative Research Groups of the Natural Science Foundation of Hubei Province (2020CFA009), and the Fundamental Research Funds for the Central Universities (2042022kf0015).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequences of genes and proteins of rice and Arabidopsis mentioned in our study are available for download from the public database mentioned above.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Peng, J.; Harberd, N.P. The role of GA-mediated signalling in the control of seed germination. Curr. Opin. Plant Biol. 2002, 5, 376–381. [Google Scholar] [CrossRef]
- Zhang, N.; Xie, Y.D.; Guo, H.J.; Zhao, L.S.; Xiong, H.C.; Gu, J.Y.; Li, J.H.; Kong, F.Q.; Sui, L.; Zhao, Z.W.; et al. Gibberellins regulate the stem elongation rate without affecting the mature plant height of a quick development mutant of winter wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2016, 1072, 28–236. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.F.; Yang, S.Y.; Chen, K.T.; Hsing, Y.I.; Zeevaart, J.A.; Chen, L.J.; Yu, S.M. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, T.; Oda, S.; Tsunaga, Y.; Shomura, H.; Kawagishi-Kobayashi, M.; Aya, K.; Saeki, K.; Endo, T.; Nagano, K.; Kojima, M.; et al. Reduction of gibberellin by low temperature disrupts pollen development in rice. Plant Physiol. 2014, 164, 2011–2019. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, E.; Atsmon, D.; Halevy, A.H. Adventitious staminate flower formation in gibberellin treated gynoecious cucumber plants. Plant Cell Physiol. 1977, 18, 1193–1201. [Google Scholar] [CrossRef]
- MacMillan, J. Occurrence of Gibberellins in Vascular Plants, Fungi, and Bacteria. J. Plant Growth Regul. 2001, 20, 387–442. [Google Scholar] [CrossRef]
- Hedden, P.; Thomas, S.G. Gibberellin biosynthesis and its regulation. Biochem. J. 2012, 444, 11–25. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, N.; Sun, T.P.; Gubler, F. Gibberellin signaling: Biosynthesis, catabolism, and response pathways. Plant Cell 2002, 14, S61–S80. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P. Gibberellin Metabolism and Its Regulation. J. Plant Growth Regul. 2001, 20, 317–318. [Google Scholar] [CrossRef]
- Arabidopsis Genome, I. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P.; Kamiya, Y. GIBBERELLIN BIOSYNTHESIS: Enzymes, Genes and Their Regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 484, 31–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carolis, E.; De Luca, V. 2-oxoglutarate-dependent dioxygenase and related enzymes: Biochemical characterization. Phytochemistry 1994, 36, 1093–1107. [Google Scholar] [CrossRef]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional cloning of rice semidwarfing gene, sd-1: Rice “green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. Dna Res. Int. J. Rapid Publ. Rep. Genes Genomes 2002, 9, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Tang, D.; Shen, Y.; Qin, B.; Hong, L.; You, A.; Li, M.; Wang, X.; Yu, H.; Gu, M.; et al. Activation of gibberellin 2-oxidase 6 decreases active gibberellin levels and creates a dominant semi-dwarf phenotype in rice (Oryza sativa L.). J. Genet. Genom. 2010, 37, 23–36. [Google Scholar] [CrossRef]
- Oikawa, T.; Koshioka, M.; Kojima, K.; Yoshida, H.; Kawata, M. A role of OsGA20ox1, encoding an isoform of gibberellin 20-oxidase, for regulation of plant stature in rice. Plant Mol. Biol. 2004, 55, 687–700. [Google Scholar] [CrossRef]
- Itoh, H.; Ueguchi-Tanaka, M.; Sentoku, N.; Kitano, H.; Matsuoka, M.; Kobayashi, M. Cloning and functional analysis of two gibberellin 3 beta -hydroxylase genes that are differently expressed during the growth of rice. Proc. Natl. Acad. Sci. USA 2001, 98, 8909–8914. [Google Scholar] [CrossRef] [Green Version]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucl. Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucl. Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Vilella, A.J.; Severin, J.; Ureta-Vidal, A.; Heng, L.; Durbin, R.; Birney, E. Ensembl Compara Gene Trees: Complete, duplication-aware phylogenetic trees in vertebrates. Genome Res. 2009, 19, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Guo, A.Y.; Zhu, Q.H.; Chen, X.; Luo, J.C. GSDS: A gene structure display server. Yi Chuan = Hered. 2007, 29, 1023–1026. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W. Noble WS: MEME SUITE: Tools for motif discovery and searching. Nucl. Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. HemI: A toolkit for illustrating heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Zhu, B. Evolutionary analysis of three gibberellin oxidase genes in rice, Arabidopsis, and soybean. Gene 2011, 473, 23–35. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucl. Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Mei, Z.; Duan, J.; Chen, H.; Feng, H.; Cai, W. OsGA2ox5, a gibberellin metabolism enzyme, is involved in plant growth, the root gravity response and salt stress. PLoS ONE 2014, 9, e87110. [Google Scholar] [CrossRef]
- Long, M.; Rosenberg, C.; Gilbert, W. Intron phase correlations and the evolution of the intron/exon structure of genes. Proc. Natl. Acad. Sci. USA 1995, 92, 12495–12499. [Google Scholar] [CrossRef] [Green Version]
- Kolkman, J.A.; Stemmer, W.P. Directed evolution of proteins by exon shuffling. Nat. Biotechnol. 2001, 19, 423–428. [Google Scholar] [CrossRef]
- Morgante, M.; Brunner, S.; Pea, G.; Fengler, K.; Zuccolo, A.; Rafalski, A. Gene duplication and exon shuffling by helitron-like transposons generate intraspecies diversity in maize. Nat. Genet. 2005, 37, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Abraham, Z.; Carbonero, P.; Diaz, I. Comparative phylogenetic analysis of cystatin gene families from arabidopsis, rice and barley. Mol. Genet. Genom. Mgg 2005, 273, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Pimenta Lange, M.J.; Liebrandt, A.; Arnold, L.; Chmielewska, S.M.; Felsberger, A.; Freier, E.; Heuer, M.; Zur, D.; Lange, T. Functional characterization of gibberellin oxidases from cucumber, Cucumis sativus L. Phytochemistry 2013, 906, 2–69. [Google Scholar] [CrossRef] [PubMed]
- Lange, T. Cloning gibberellin dioxygenase genes from pumpkin endosperm by heterologous expression of enzyme activities in Escherichia coli. Proc. Natl. Acad. Sci. USA 1997, 94, 6553–6558. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Cerezo, S.; Martinez-Montiel, N.; Garcia-Sanchez, J.; Perez, Y.T.R.; Martinez-Contreras, R.D. Gibberellin biosynthesis and metabolism: A convergent route for plants, fungi and bacteria. Microbiol. Res. 2018, 2088, 5–98. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Liu, X.; Zhang, X.; Liu, S.; Yu, X.; Ren, Y.; Zheng, X.; Zhou, K.; Jiang, L.; et al. A role for a dioxygenase in auxin metabolism and reproductive development in rice. Dev. Cell 2013, 27, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Sakamoto, T.; Morinaka, Y.; Ishiyama, K.; Kobayashi, M.; Itoh, H.; Kayano, T.; Iwahori, S.; Matsuoka, M.; Tanaka, H. Genetic manipulation of gibberellin metabolism in transgenic rice. Nat. Biotechnol. 2003, 21, 909–913. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).