
First Report of Helicoverpa armigera (Lepidoptera: Noctuidae) Resistance to Flubendiamide in Brazil: Genetic Basis and Mechanisms of the Resistance

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Characterization of Flubendiamide Resistance
2.3. Inheritance of Flubendiamide Resistance
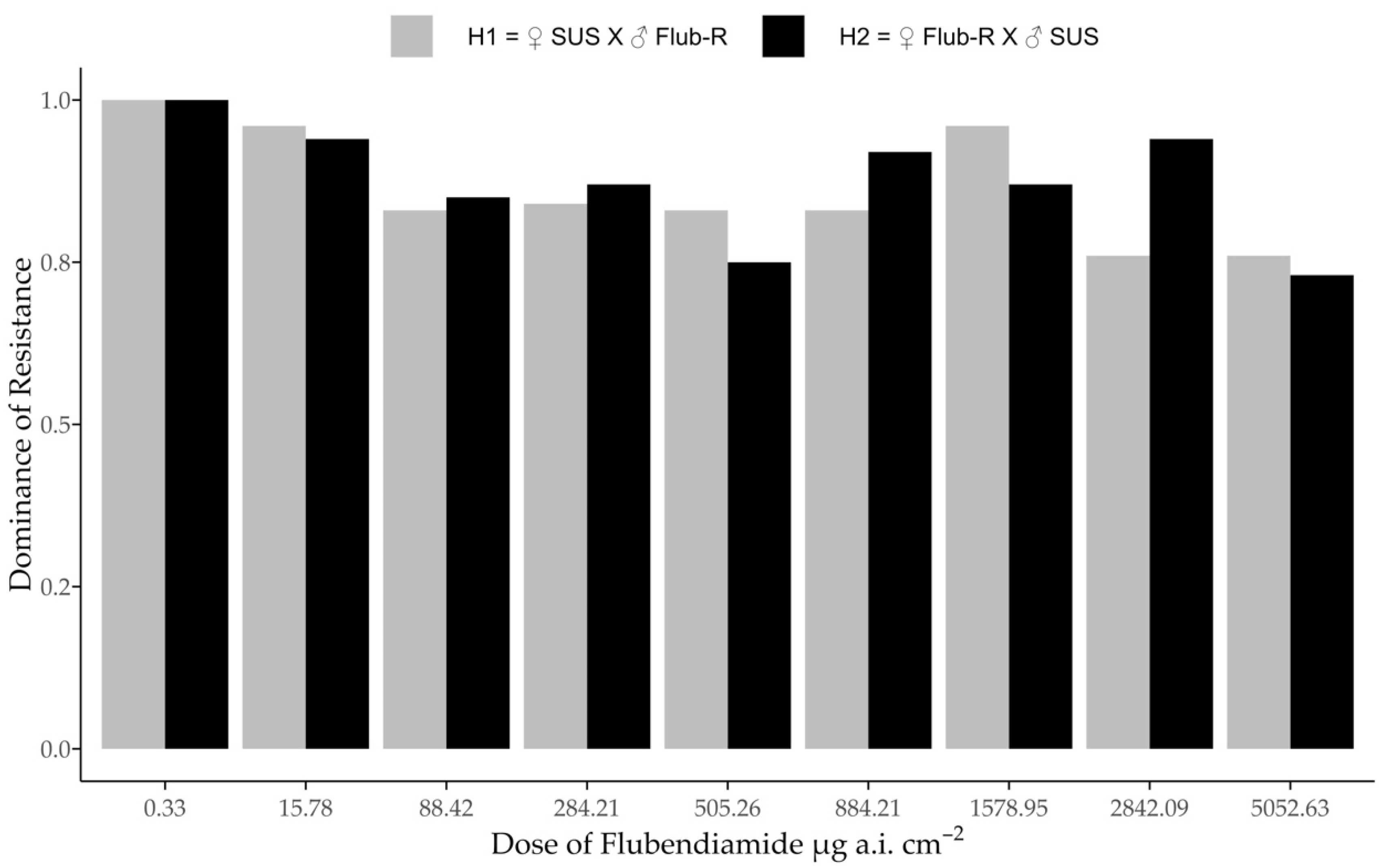
2.4. Dominance of Resistance
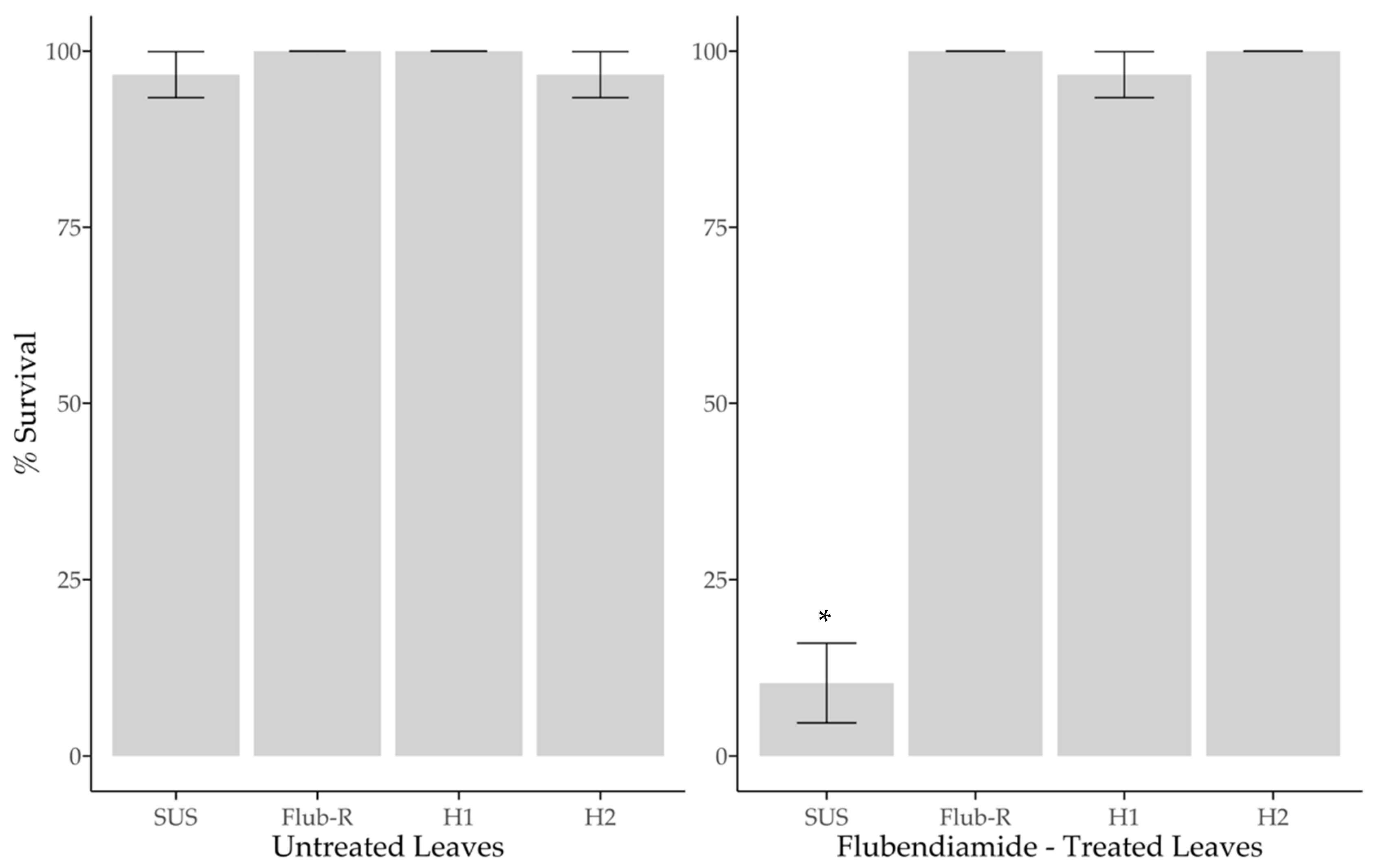
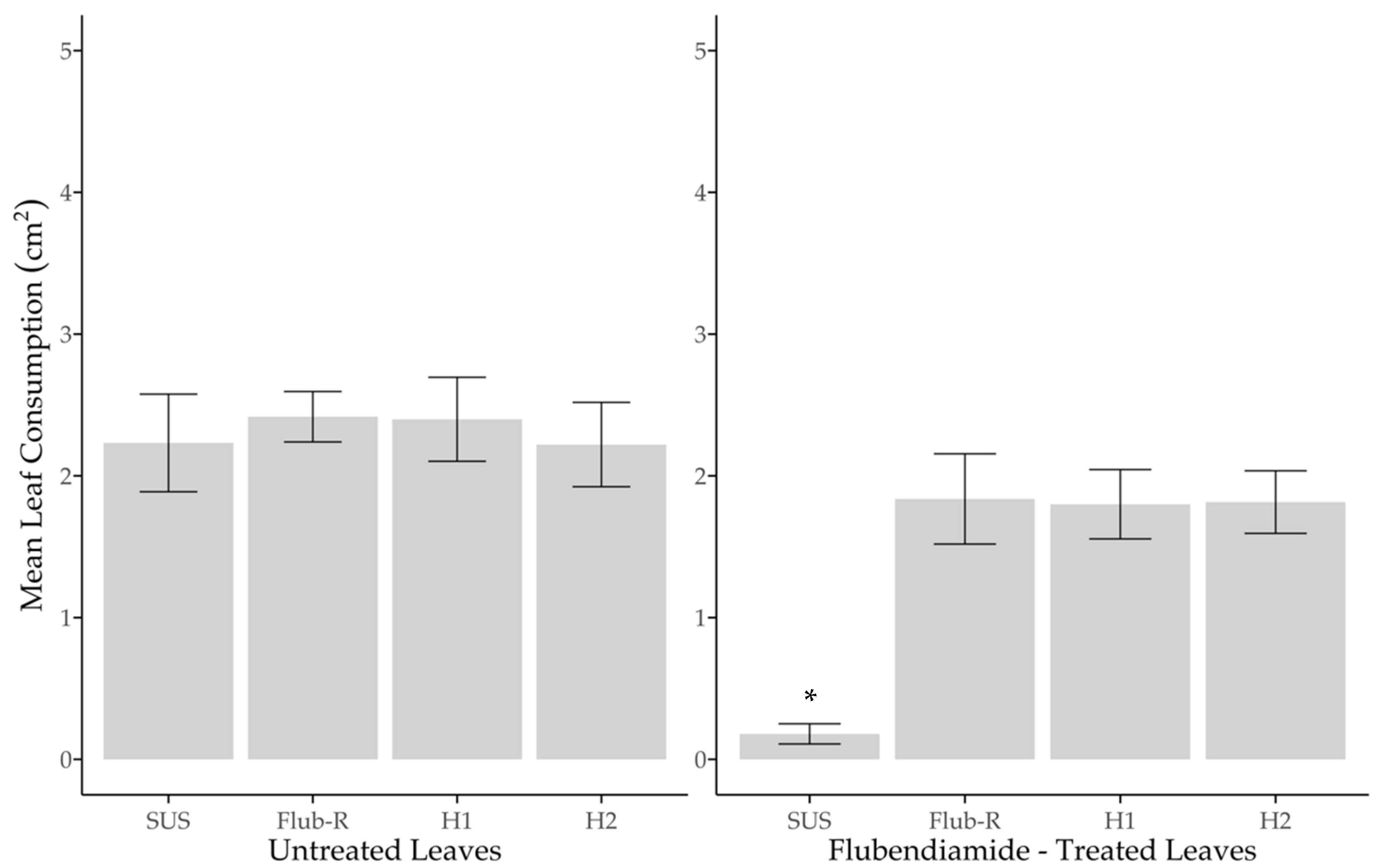
2.5. Functional Dominance of Resistance and Leaf Consumption
2.6. Cross Resistance
2.7. Synergist Bioassays
2.8. Sequencing Partial RyR
2.9. Data Analysis
3. Results
3.1. Characterization of Flubendiamide Resistance
3.2. Inheritance of Flubendiamide Resistance
3.3. Dominance of Resistance
3.4. Functional Dominance of Resistance and Leaf Consumption
3.5. Cross Resistance
3.6. Synergist Bioassays
3.7. Sequencing Partial RyR
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Czepak, C.; Albernaz, K.C.; Vivan, L.M.; Guimarães, H.O.; Carvalhais, T. First reported occurrence of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) in Brazil. Pesq. Agropecu. Trop. 2013, 43, 110–113. [Google Scholar]
- Tay, W.T.; Soria, M.F.; Walsh, T.; Thomazoni, D.; Silvie, P.; Behere, G.T.; Anderson, C.; Downes, S. A brave new world for an old world pest: Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. PLoS ONE 2013, 8, e80134. [Google Scholar]
- Fitt, G.P. The ecology of Heliothis species in relation to agroecosystems. Ann. Rev. Entomol. 1989, 66, 17–52. [Google Scholar]
- Tay, W.T.; Gordon, K.H.J. Going global—Genomic insights into insect invasions. Curr. Opin. Insect Sci. 2019, 31, 123–130. [Google Scholar]
- Murúa, M.G.; Scalora, F.S.; Navarro, F.R.; Cazado, L.E.; Casmuz, A.; Villagrán, M.E.; Lobos, E.; Gastaminza, G. First record of Helicoverpa armigera (Lepidoptera: Noctuidae) in Argentina. Fla. Entomol. 2014, 97, 854–856. [Google Scholar]
- Arnemann, J.A.; James, W.J.; Walsh, T.K.; Guedes, J.V.C.; Smagghe, G.; Castiglioni, E.; Tay, W.T. Mitochondrial DNA COI characterization of Helicoverpa armigera (Lepidoptera: Noctuidae) from Paraguay and Uruguay. Gen. Mol. Res. 2016, 15, gmr8292. [Google Scholar]
- Castiglioni, E.; Perini, C.R.; Chiaravalle, W.; Arnemann, J.A.; Ugalde, G.; Guedes, J.V.C. Primer registro de ocurrencia de Helicoverpa armigera (Hübner, 1808) (Lepidoptera: Noctuidae) en soja, en Uruguay. Agrociencia 2016, 20, 31–35. Available online: http://www.scielo.edu.uy/scielo.php?script=sci_arttext&pid=S2301-15482016000100005&nrm=iso (accessed on 30 May 2022).
- Gilligan, T.M.; Goldstein, P.Z.; Timm, A.E.; Farris, R.; Ledezma, L.; Cunningham, A.P. Identification of Heliothine (Lepidoptera: Noctuidae) larvae intercepted at U.S. ports of entry from the new world. J. Econ. Entomol. 2019, 112, 603–615. [Google Scholar]
- Cunningham, J.P.; Zalucki, M.P. Understanding Heliothine (Lepidoptera: Heliothinae) pests: What is a host plant? J. Econ. Entomol. 2014, 107, 881–896. [Google Scholar]
- Widmer, M.W.; Schofield, P. Heliothis: Dispersal and Migration; Tropical Development and Research Institute: London, UK, 1983; Volume 2, p. 41. [Google Scholar]
- Mota-Sanchez, D.; Wise, J.C. The Arthropod Pesticide Resistance Database; Michigan State University: East Lansing, MI, USA, 2022; Available online: http://www.pesticideresistance.org (accessed on 27 February 2022).
- Jeanguenat, A. The story of a new insecticidal chemistry class: The diamides. Pest Manag. Sci. 2013, 69, 7–14. [Google Scholar]
- Cordova, D.; Benner, E.A.; Sacher, M.D.; Rauh, J.J.; Sopa, J.S.; Lahm, G.P.; Selby, T.P.; Stevenson, T.M.; Flexner, L.; Gutteridge, S.; et al. Anthranilic diamides: A new class of insecticides with a novel mode of action, ryanodine receptor activation. Pestic. Biochem. Physiol. 2006, 84, 196–214. [Google Scholar]
- Kato, K.; Kiyonaka, S.; Sawaguchi, Y.; Tohnishi, M.; Masaki, T.; Yasokawa, N.; Mizuno, Y.; Mori, E.; Inoue, K.; Hamachi, I.; et al. Molecular characterization of flubendiamide sensitivity in the lepidopterous ryanodine receptor Ca2+ release channel. Biochemistry 2009, 48, 10342–10352. [Google Scholar]
- Richardson, E.B.; Troczka, B.J.; Gutbrod, O.; Davies, T.G.E.; Nauen, R. Diamide resistance: 10 years of lessons from lepidopteran pests. J. Pest Sci. 2020, 93, 911–928. [Google Scholar]
- Zuo, Y.; Wang, H.; Xu, Y.; Huang, J.; Wu, S.; Wu, Y.; Yang, Y. CRISPR/Cas9 mediated G4946E substitution in the ryanodine receptor of Spodoptera exigua confers high levels of resistance to diamide insecticides. Insect Biochem. Mol. Biol. 2017, 89, 79–85. [Google Scholar]
- Roditakis, E.; Steinbach, D.; Moritz, G.; Vasakis, E.; Stavrakaki, M.; Ilias, A.; García-Vidal, L.; Martínez-Aguirre, M.R.; Bielza, P.; Morou, E.; et al. Ryanodine receptor point mutations confer diamide insecticide resistance in tomato leafminer, Tuta absoluta (Lepidoptera: Gelechiidae). Insect Biochem. Mol. Biol. 2017, 80, 11–20. [Google Scholar]
- Troczka, B.J.; Williamson, M.S.; Field, L.M.; Davies, T.G.E. Rapid selection for resistance to diamide insecticides in Plutella xylostella via specific amino acid polymorphisms in the ryanodine receptor. Neuro Toxicol. 2017, 60, 224–233. [Google Scholar]
- Bolzan, A.; Padovez, F.E.; Nascimento, A.R.; Kaiser, I.S.; Lira, E.C.; Amaral, F.S.; Kanno, R.H.; Malaquias, J.B.; Omoto, C. Selection and characterization of the inheritance of resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to chlorantraniliprole and cross-resistance to other diamide insecticides. Pest Manag. Sci. 2019, 75, 2682–2689. [Google Scholar]
- Troczka, B.; Zimmer, C.T.; Elias, J.; Schorn, C.; Bass, C.; Davies, T.G.E.; Field, L.M.; Williamson, M.S.; Slater, R.; Nauen, R. Resistance to diamide insecticides in diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae) is associated with a mutation in the membrane-spanning domain of the ryanodine receptor. Insect Biochem. Mol. Biol. 2012, 42, 873–880. [Google Scholar]
- Zuo, Y.; Ma, H.; Lu, W.; Wang, X.; Wu, S.; Nauen, R.; Wu, Y.; Yang, Y. Identification of the ryanodine receptor mutation I4743M and its contribution to diamide insecticide resistance in Spodoptera exigua (Lepidoptera: Noctuidae). Insect Sci. 2020, 27, 791–800. [Google Scholar]
- Douris, V.; Papapostolou, K.M.; Ilias, A.; Roditakis, E.; Kounadi, S.; Riga, M.; Nauen, R.; Vontas, J. Investigation of the contribution of RyR target-site mutations in diamide resistance by CRISPR/Cas9 genome modification in Drosophila. Insect Biochem. Mol. Biol. 2017, 87, 127–135. [Google Scholar]
- Pereira, R.M.; Abbade-Neto, D.; Amado, D.; Durigan, M.R.; Franciscatti, R.A.; Mocheti, M.; Omoto, C. Baseline susceptibility and frequency of resistance to diamide insecticides in Helicoverpa armigera (Lepidoptera: Noctuidae) populations in Brazil. Crop Prot. 2020, 137, 105266. [Google Scholar]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. Velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar]
- Stone, B.F. A formula for determining degree of dominance in cases of monofactorial inheritance of resistance to chemicals. Bull. World Health Organ 1968, 38, 325–326. [Google Scholar]
- Lehmann, E.L.; Romano, J.P. Testing Statistical Hypotheses, 3rd ed.; Springer: New York, NY, USA, 2005. [Google Scholar]
- Robertson, J.L.; Jones, M.M.; Olguin, E.; Alberts, B. Bioassays with Arthropods, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Bourguet, D.; Genissel, A.; Raymond, M. Insecticide resistance and dominance levels. J. Econ. Entomol. 2000, 93, 1588–1595. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Rigby, R.A.; Stasinopoulos, D.M. Generalized additive models for location, scale and shape. J. R. Stat. Soc. 2005, 54, 507–554. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Soft. 2016, 69, 1–33. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality. Biometrika 1965, 52, 591–611. [Google Scholar]
- Bartlett, M.S. Properties of sufficiency and statistical tests. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 1937, 160, 268–282. [Google Scholar]
- Durbin, J.; Watson, G.S. Testing for serial correlation in least squares regression. Biometrika 1971, 58, 1. [Google Scholar]
- Zeileis, A.; Hothorn, T. Diagnostic checking in regression relationships. R News 2002, 2, 7–10. [Google Scholar]
- Silva, J.E.; Ribeiro, L.M.S.; Vinasco, N.; Guedes, R.N.C.; Siqueira, H.Á.A. Field-evolved resistance to chlorantraniliprole in the tomato pinworm Tuta absoluta: Inheritance, cross-resistance profile, and metabolism. J. Pest Sci. 2019, 92, 1421–1431. [Google Scholar]
- Wang, X.; Khakame, S.K.; Ye, C.; Yang, Y.; Wu, Y. Characterization of field-evolved resistance to chlorantraniliprole in the diamondback moth, Plutella xylostella, from China. Pest Manag. Sci. 2013, 69, 661–665. [Google Scholar]
- Roush, R.T.; Mckenzie, J.A. Ecological genetics of insecticide and acaricide resistance. Ann. Rev. Entomol. 1987, 32, 361–380. [Google Scholar]
- Tabashnik, B.E.; Croft, B.A. Managing pesticide resistance in crop-arthropod complexes: Interactions between biological and operational factors 1. Environ. Entomol. 1982, 11, 1137–1144. [Google Scholar]
- Roditakis, E.; Vasakis, E.; Grispou, M.; Stavrakaki, M.; Nauen, R.; Gravouil, M.; Bassi, A. First report of Tuta absoluta resistance to diamide insecticides. J. Pest Sci. 2015, 88, 9–16. [Google Scholar]
- Li, X.; Li, R.; Zhu, B.; Gao, X.; Liang, P. Overexpression of cytochrome P450 CYP6BG1 may contribute to chlorantraniliprole resistance in Plutella xylostella (L.). Pest Manag. Sci. 2018, 74, 1386–1393. [Google Scholar]
- Zhao, J.; Xu, L.; Sun, Y.; Song, P.; Han, Z. UDP-Glycosyltransferase genes in the striped rice stem borer, Chilo suppressalis (Walker), and their contribution to chlorantraniliprole resistance. Int. J. Mol. Sci. 2019, 20, 1064. [Google Scholar]
- Yin, F.; Lin, Q.; Wang, X.; Li, Z.; Feng, X.; Shabbir, M.Z. The glutathione S-transferase (PxGST2L) may contribute to the detoxification metabolism of chlorantraniliprole in Plutella xylostella (L.). Ecotoxicology 2021, 30, 1007–1016. [Google Scholar]
- Ma, R.; Haji-Ghassemi, O.; Ma, D.; Jiang, H.; Lin, L.; Yao, L.; Samurkas, A.; Li, Y.; Wang, Y.; Cao, P.; et al. Structural basis for diamide modulation of ryanodine receptor. Nat. Chem. Biol. 2020, 16, 1246–1254. [Google Scholar]
- Boaventura, D.; Bolzan, A.; Padovez, F.E.; Okuma, D.M.; Omoto, C.; Nauen, R. Detection of a ryanodine receptor target-site mutation in diamide insecticide resistant fall armyworm, Spodoptera frugiperda. Pest Manag. Sci. 2019, 76, 47–54. [Google Scholar]
- Jouraku, A.; Kuwazaki, S.; Miyamoto, K.; Uchiyama, M.; Kurokawa, T.; Mori, E.; Mori, M.X.; Mori, Y.; Sonoda, S. Ryanodine receptor mutations (G4946E and I4790K) differentially responsible for diamide insecticide resistance in diamondback moth, Plutella xylostella L. Insect Biochem. Mol. Biol. 2020, 118, 103308. [Google Scholar]
- Kim, J.; Nam, H.Y.; Kwon, M.; Choi, J.H.; Cho, S.R.; Kim, G.-H. Novel diamide resistance-linked mutation in Korean Spodoptera exigua and a LAMP assay based on a mutation-associated intronic InDel. J. Pest Sci. 2021, 94, 1017–1029. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′-3′ | Amplification Length (bp) |
|---|---|---|
| HA1-F | CTCAAGAGAGGATGGAAGC | 728 |
| HA1-R | AGAAGGAAGTCTTCTCCATGC | |
| HA2-F | TCGCTCGTAAGCTGGAAT | 756 |
| HA2-R | AGACCCTGGAGAATAGCG | |
| HA3-F | TTCCGCAAGTTCTACGTGC | 510 |
| HA3-R | TTCGGAAACAATCTCCCACG |
| Strain | n a | Slope (±SE) | LD50 (95% CI) b | χ2 (p) c | d.f. d | RR e |
|---|---|---|---|---|---|---|
| SUS | 864 | 1.98 (±0.12) | 0.017 (0.015–0.019) | 8.70 (0.88) | 6 | - |
| Flub-R | 662 | 1.58 (±0.11) | 871.84 (744.74–1020.62) | 1.96 (0.14) | 5 | >50,000 |
| H1 (♀ SUS × ♂ Flub-R) | 707 | 1.14 (±0.08) | 509.43 (412.26–629.48) | 10.68 (0.90) | 6 | 29,966.47 |
| H2 (♀ Flub-R × ♂ SUS) | 709 | 1.09 (±0.08) | 504.36 (403.78–630.00) | 10.67 (0.90) | 6 | 29,647.06 |
| Insecticide | Strain | n a | Slope (±SE) | LD50 (CI 95%) b | χ2 (p) c | d.f. d | RR e |
|---|---|---|---|---|---|---|---|
| Chlorantraniliprole | SUS | 499 | 2.28 (±0.22) | 0.020 (0.016–0.025) | 7.19 (0.79) | 5 | - |
| Flub-R | 767 | 1.59 (±0.11) | 0.090 (0.077–0.106) | 1.78 (0.12) | 5 | 4.50 | |
| Cyantraniliprole | SUS | 497 | 1.51 (±0.12) | 0.039 (0.032–0.047) | 3.65 (0.40) | 5 | - |
| Flub-R | 637 | 1.29 (±0.09) | 0.193 (0.158–0.237) | 4.25 (0.48) | 5 | 4.94 | |
| Cyclaniliprole | SUS | 475 | 1.54 (±0.12) | 0.003 (0.002–0.004) | 7.11 (0.78) | 5 | - |
| Flub-R | 461 | 2.00 (±0.14) | 0.004 (0.003–0.005) | 2.60 (0.23) | 5 | 1.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbade-Neto, D.; Amado, D.; Pereira, R.M.; Basso, M.; Spineli-Silva, S.; Gonçalves, T.M.; Corrêa, A.S.; Omoto, C. First Report of Helicoverpa armigera (Lepidoptera: Noctuidae) Resistance to Flubendiamide in Brazil: Genetic Basis and Mechanisms of the Resistance. Agronomy 2022, 12, 1664. https://doi.org/10.3390/agronomy12071664
Abbade-Neto D, Amado D, Pereira RM, Basso M, Spineli-Silva S, Gonçalves TM, Corrêa AS, Omoto C. First Report of Helicoverpa armigera (Lepidoptera: Noctuidae) Resistance to Flubendiamide in Brazil: Genetic Basis and Mechanisms of the Resistance. Agronomy. 2022; 12(7):1664. https://doi.org/10.3390/agronomy12071664
Chicago/Turabian StyleAbbade-Neto, Dyrson, Douglas Amado, Rogério Machado Pereira, Murilo Basso, Sandy Spineli-Silva, Thaini Mariane Gonçalves, Alberto Soares Corrêa, and Celso Omoto. 2022. "First Report of Helicoverpa armigera (Lepidoptera: Noctuidae) Resistance to Flubendiamide in Brazil: Genetic Basis and Mechanisms of the Resistance" Agronomy 12, no. 7: 1664. https://doi.org/10.3390/agronomy12071664
APA StyleAbbade-Neto, D., Amado, D., Pereira, R. M., Basso, M., Spineli-Silva, S., Gonçalves, T. M., Corrêa, A. S., & Omoto, C. (2022). First Report of Helicoverpa armigera (Lepidoptera: Noctuidae) Resistance to Flubendiamide in Brazil: Genetic Basis and Mechanisms of the Resistance. Agronomy, 12(7), 1664. https://doi.org/10.3390/agronomy12071664