
Herbage and Silage Quality Improved More by Mixing Barley and Faba Bean Than by N Fertilization or Stage of Harvest

 ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Analyses and Variables Measured on Both the Herbage and the Silage
2.3. Computations and Statistical Analyses
3. Results
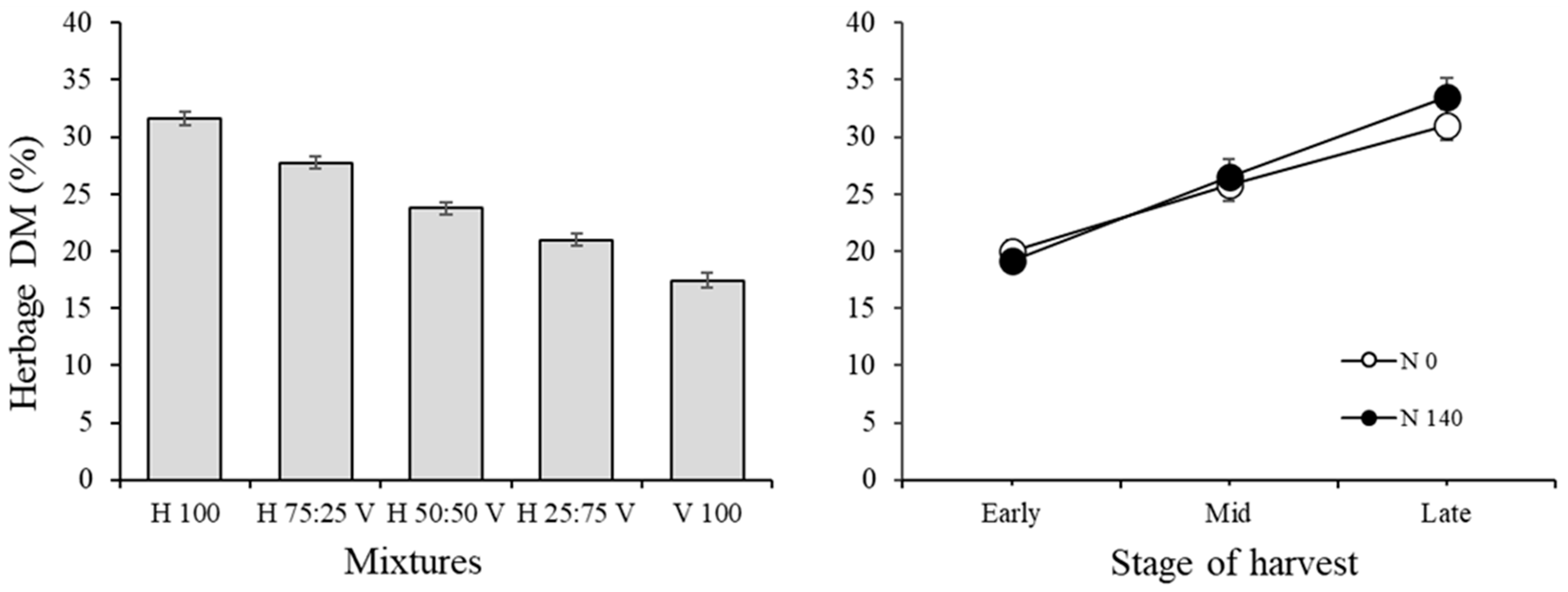
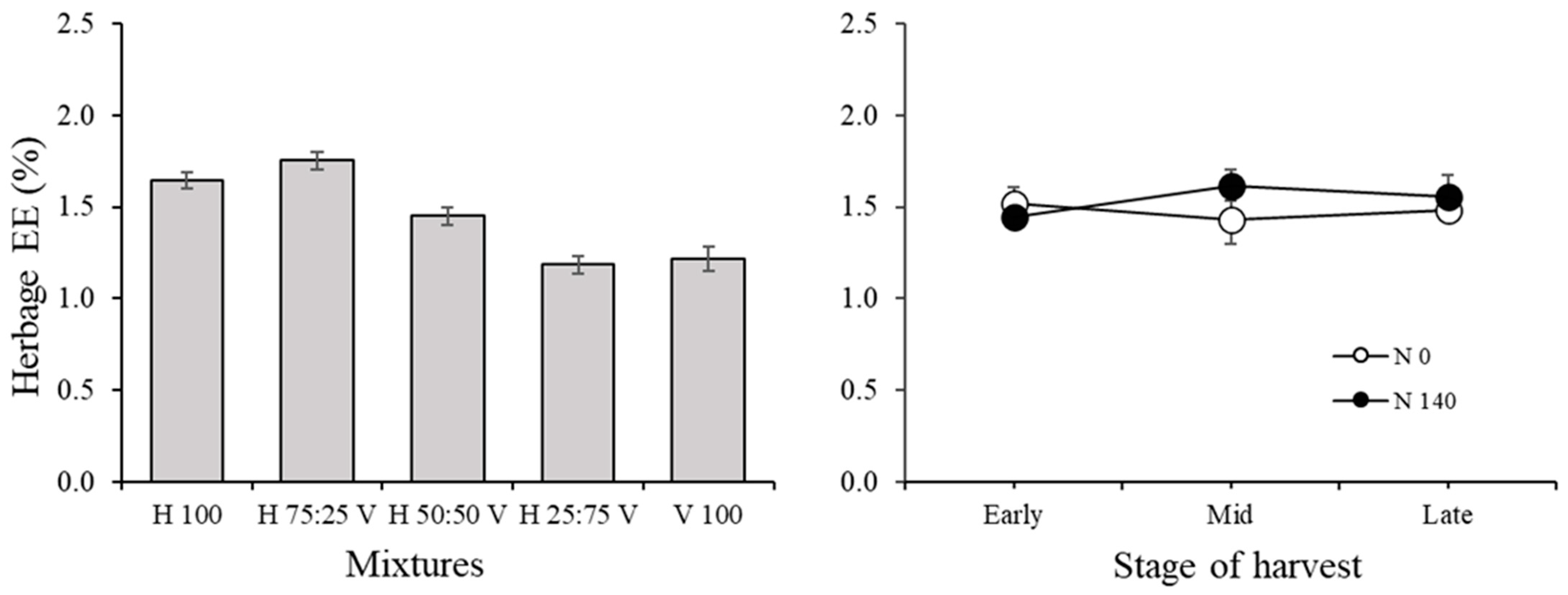
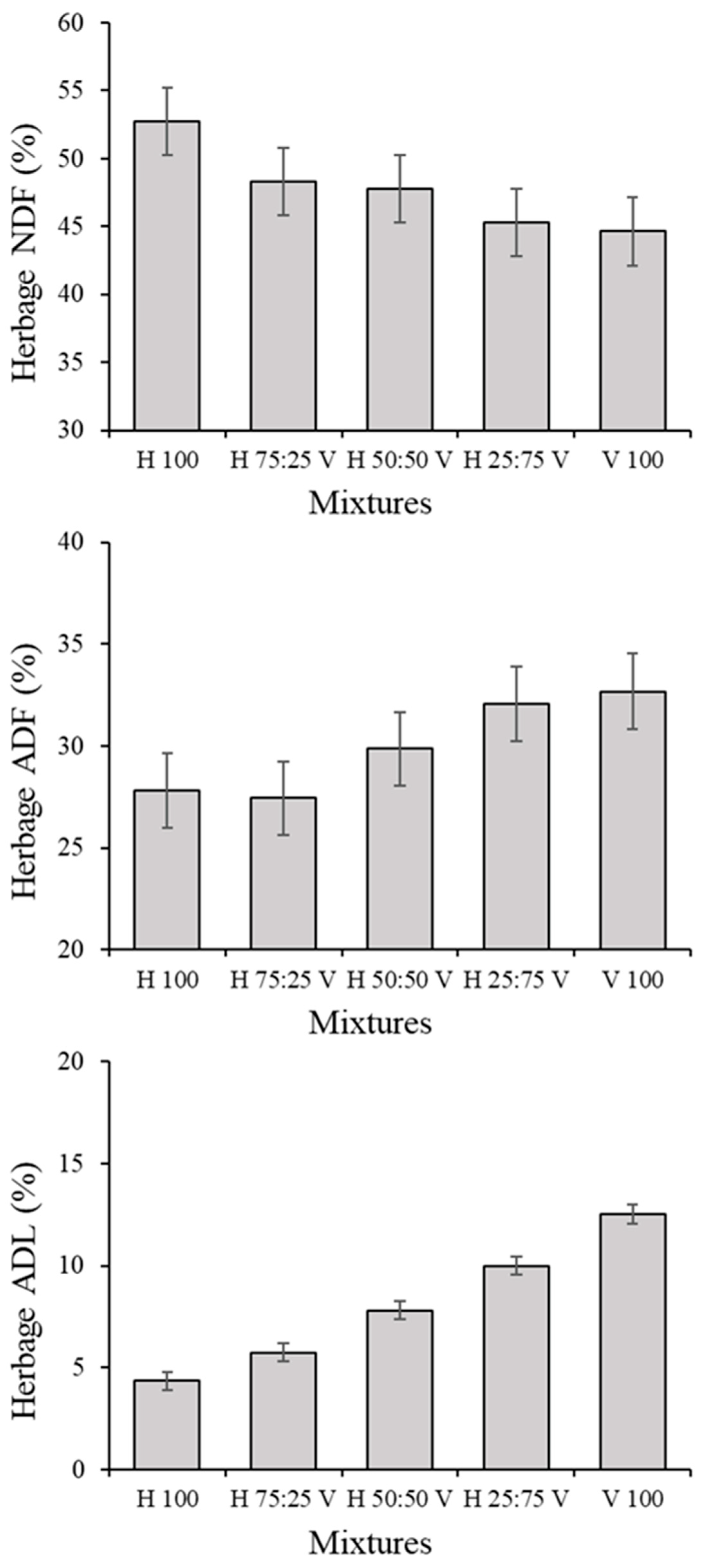
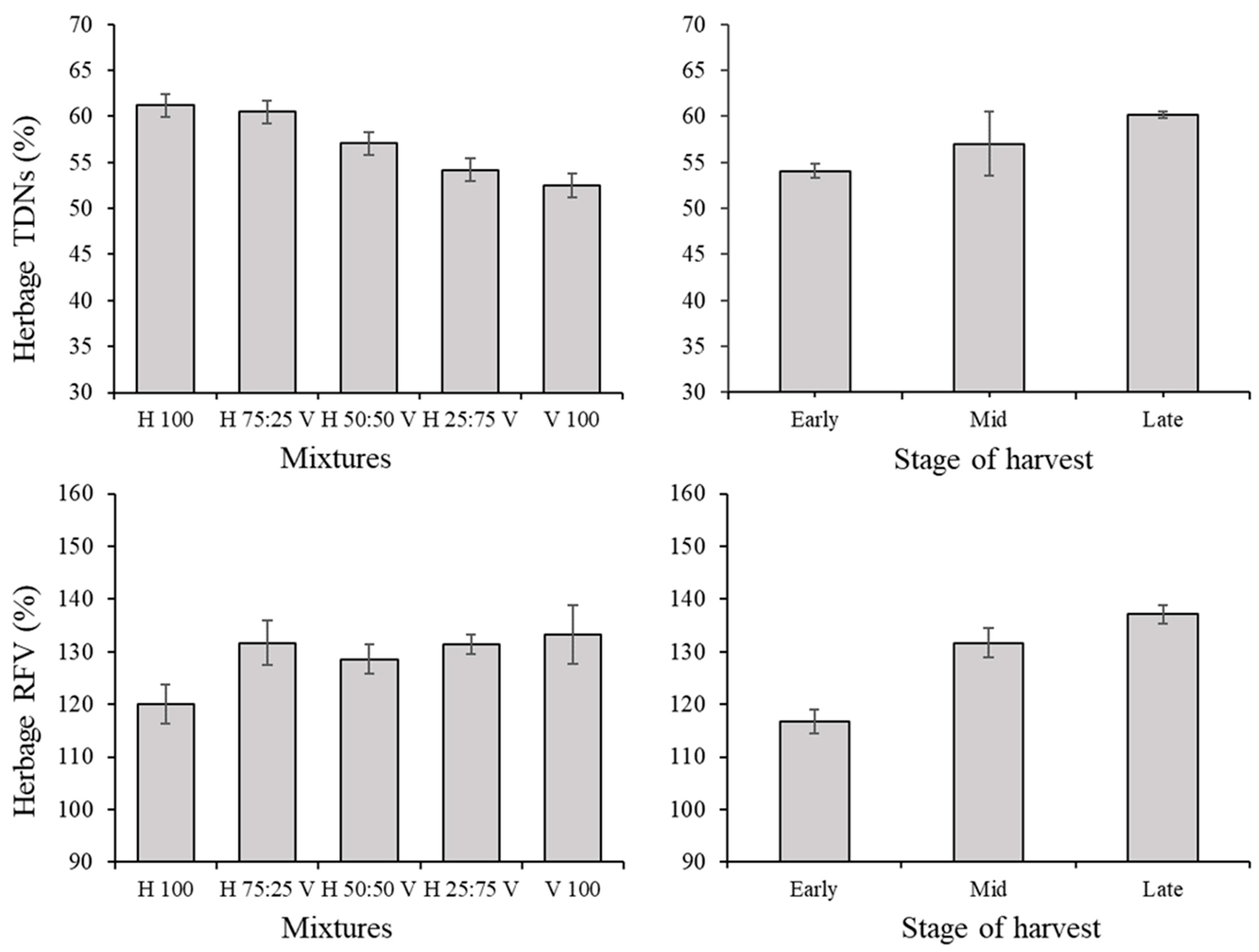
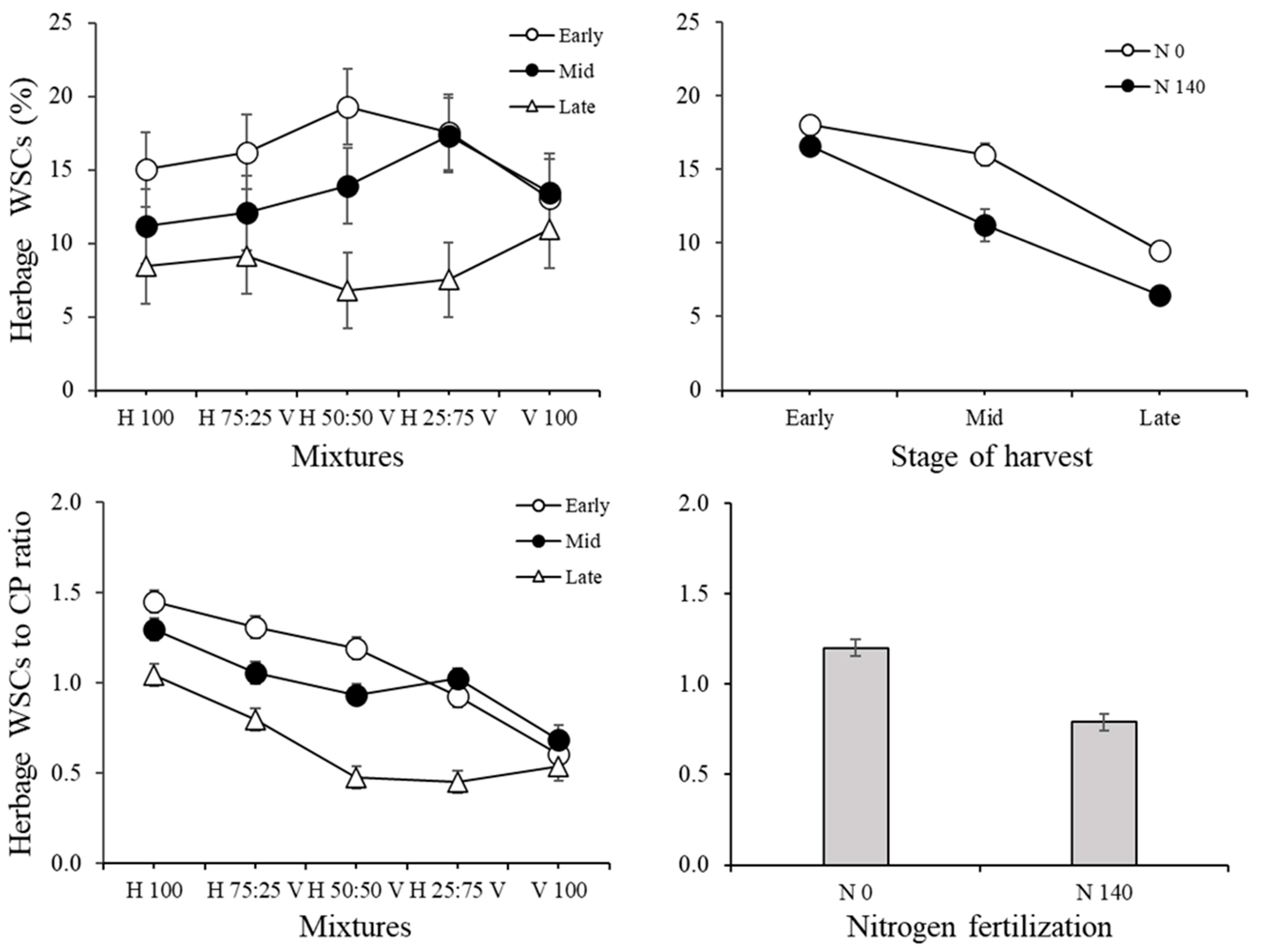
3.1. Herbage Traits and Correlations among Variables in the Herbage
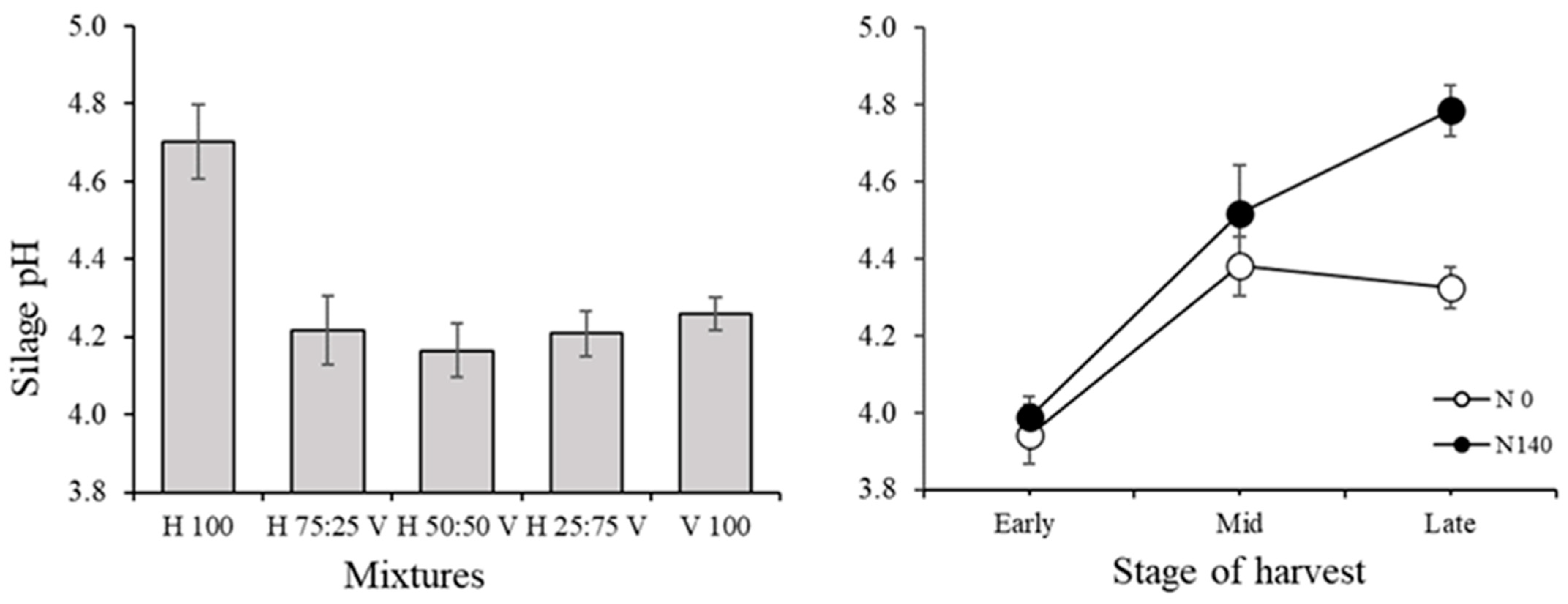
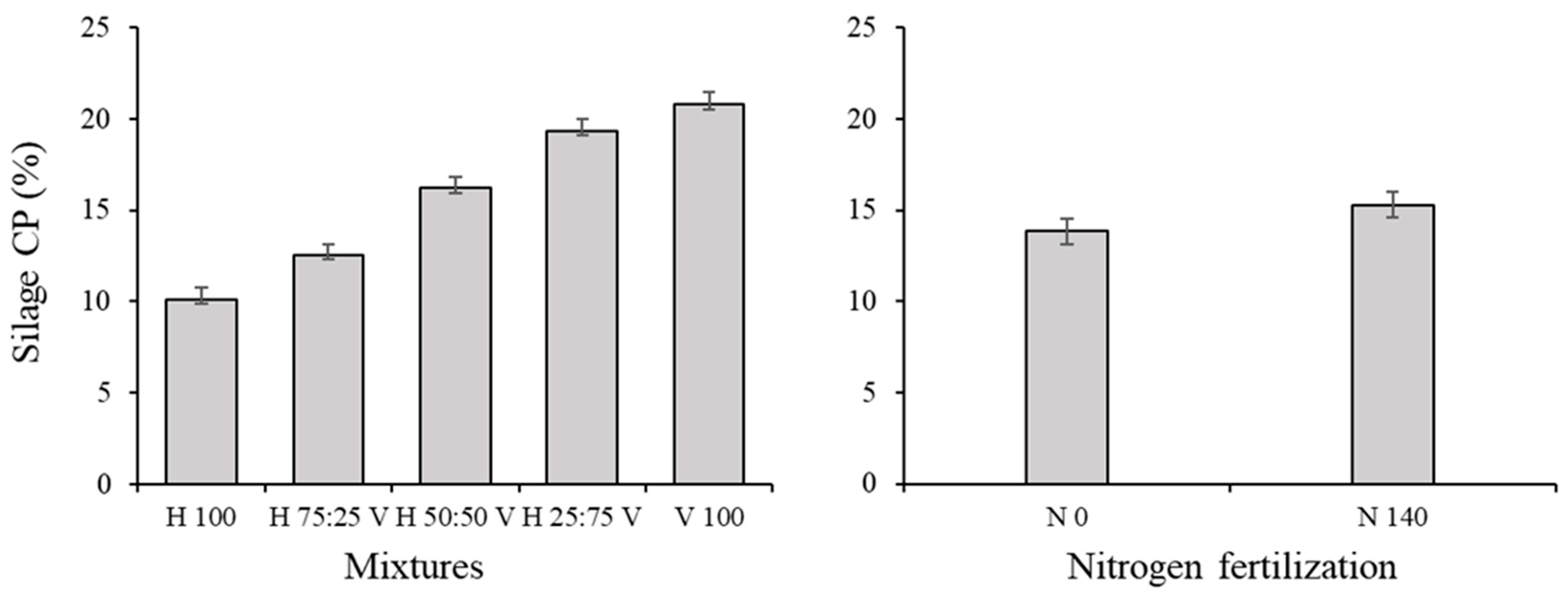
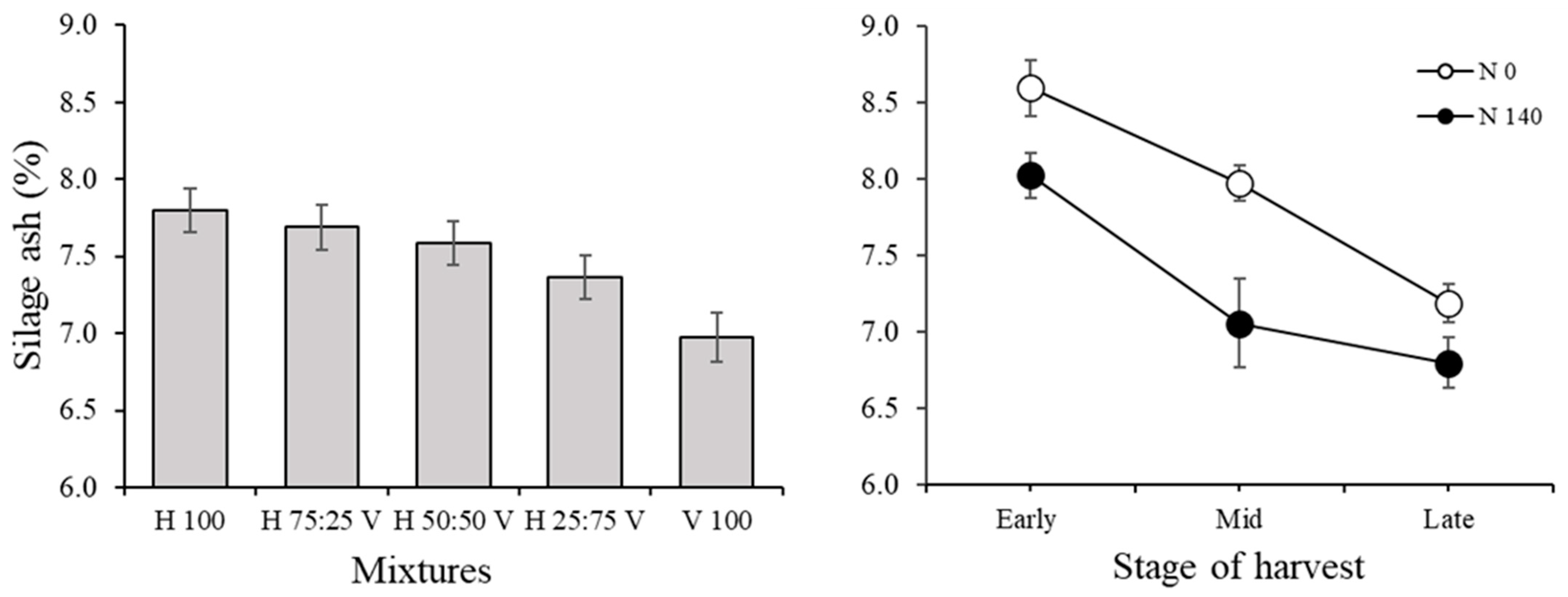
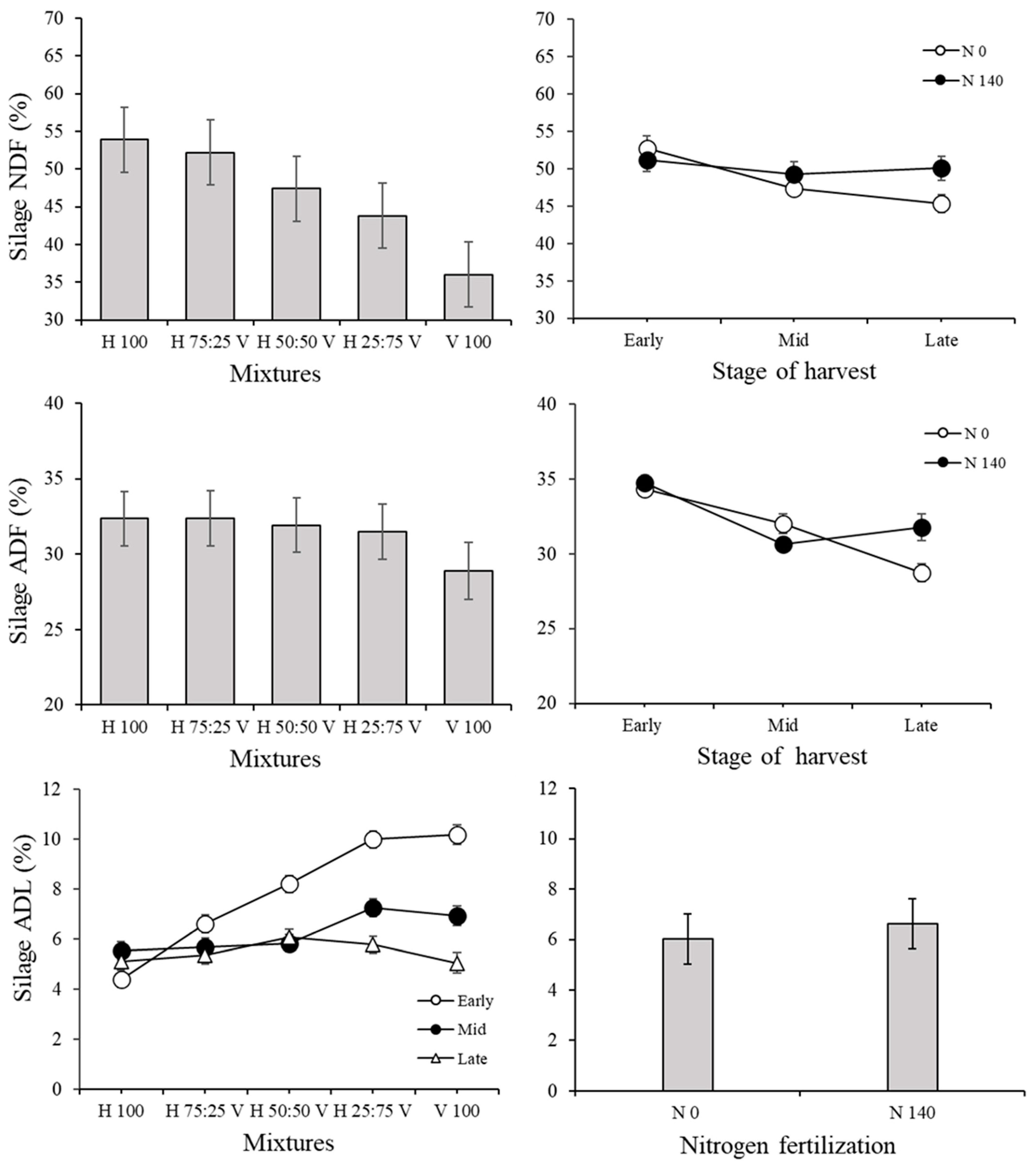
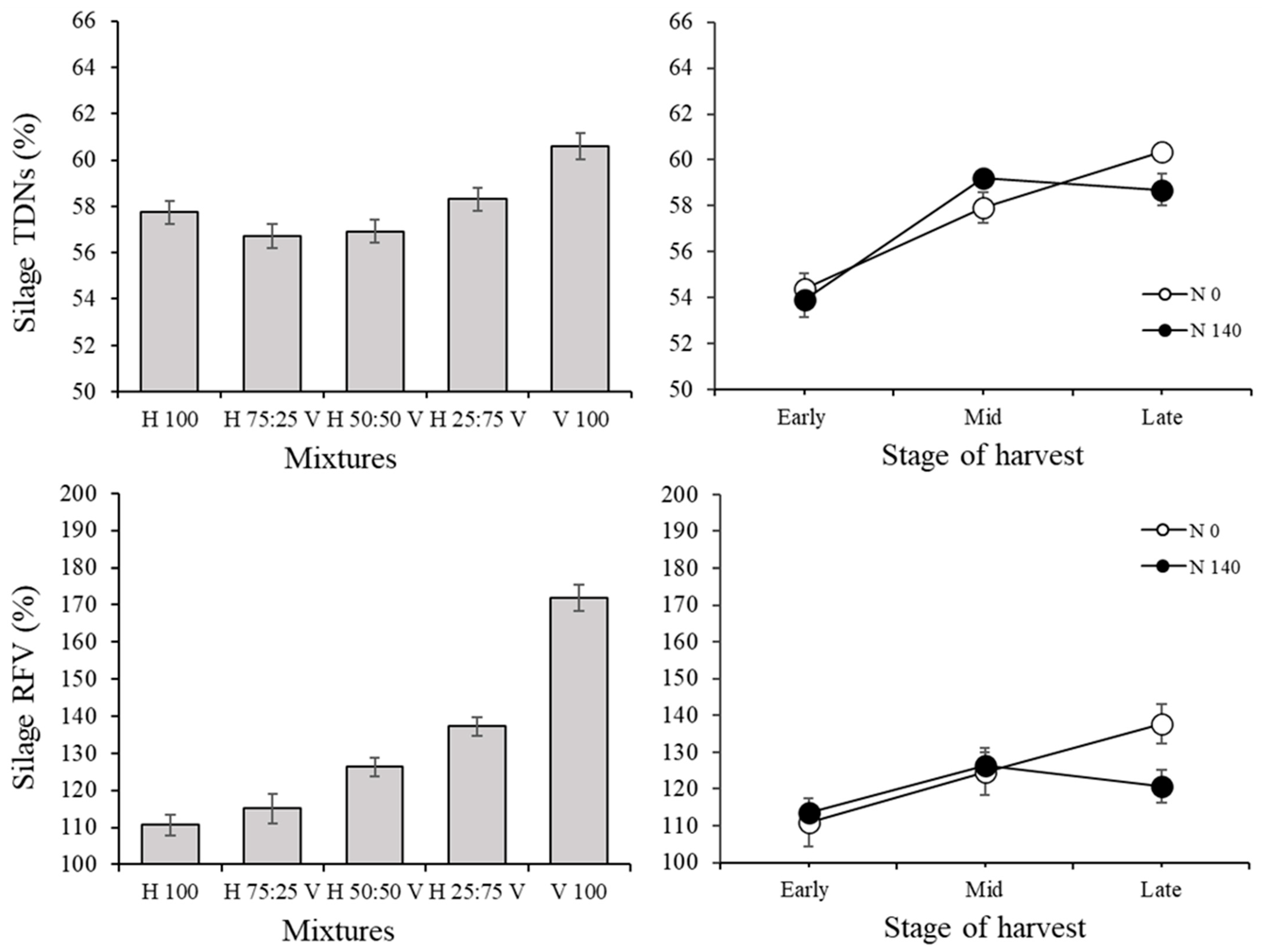
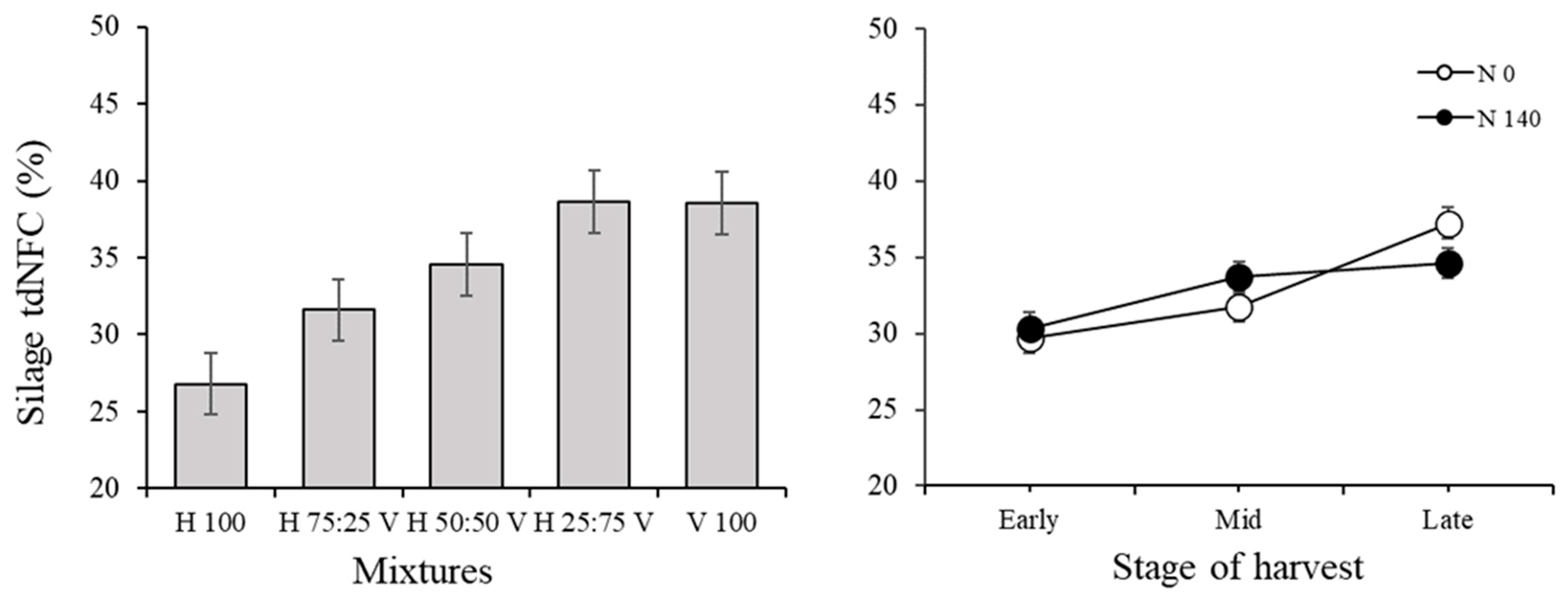
3.2. Silage Traits and Correlations among Variables in the Silage
3.3. Variation of Traits from Herbage to Silage
3.4. Choice of the Variables and Results from the Canonical Discriminant Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, Y.; Stomph, T.-J.; Makowski, D.; van der Werf, W. Temporal niche differentiation increases the land equivalent ratio of annual intercrops: A meta-analysis. Field Crop. Res. 2015, 184, 133–144. [Google Scholar] [CrossRef]
- Yu, Y.; Makowski, D.; Stomph, T.-J.; van der Werf, W. Robust Increases of Land Equivalent Ratio with Temporal Niche Differentiation: A Meta-Quantile Regression. Agron. J. 2016, 108, 2269–2279. [Google Scholar] [CrossRef]
- Saia, S.; Urso, V.; Amato, G.; Frenda, A.S.; Giambalvo, D.; Ruisi, P.; Di Miceli, G. Mediterranean forage legumes grown alone or in mixture with annual ryegrass: Biomass production, N2 fixation, and indices of intercrop efficiency. Plant Soil 2016, 402, 395–407. [Google Scholar] [CrossRef]
- Yu, Y.; Stomph, T.-J.J.; Makowski, D.; Zhang, L.; van der Werf, W. A meta-analysis of relative crop yields in cereal/legume mixtures suggests options for management. Field Crop. Res. 2016, 198, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Lithourgidis, A.S.; Dordas, C.A.; Damalas, C.A.; Vlachostergios, D.N. Annual intercrops: An alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]
- Willey, R.W. Resource use in intercropping systems. Agric. Water Manag. 1990, 17, 215–231. [Google Scholar] [CrossRef]
- Olesen, J.E.; Trnka, M.; Kersebaum, K.C.; Skjelvåg, A.O.; Seguin, B.; Peltonen-Sainio, P.; Rossi, F.; Kozyra, J.; Micale, F. Impacts and adaptation of European crop production systems to climate change. Eur. J. Agron. 2011, 34, 96–112. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Change 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Elferink, S.O.; Driehuis, F.; Gottschal, J.; Spoelstra, S. Silage fermentation processes and their manipulation. In Proceedings of the FAO Electronic Conference on Tropical Silage. Silage Making in the Tropics with Particular Emphasis on Smallholders. 1 September–15 December 1999; Mannetje, L., Ed.; FAO Plant Production and Protection Paper; Food and Agriculture Organization: Rome, Italy, 2000; Available online: http://www.fao.org/docrep/005/x8486e/x8486e09.htm (accessed on 28 July 2022).
- Anil, L.; Park, J.; Phipps, R.H.; Miller, F.A. Temperate intercropping of cereals for forage: A review of the potential for growth and utilization with particular reference to the UK. Grass Forage Sci. 1998, 53, 301–317. [Google Scholar] [CrossRef]
- Eskandari, H.; Ghanbari, A.; Javanmard, A. Intercropping of cereals and legumes for forage production. Not. Sci. Biol. 2009, 1, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Kung, L.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef]
- Buxton, D.R.; O’Kiely, P. (Eds.) Preharvest plant factors affecting ensiling. In Silage Science and Technology; American Society of Agronomy, Madison (US); American Society of Agronomy: Madison, WI, USA, 2015; pp. 199–250. ISBN 9780891182344. [Google Scholar]
- Berkenkamp, B.; Meeres, J. Mixtures of Annual Crops for Forage in Central Alberta. Can. J. Plant Sci. 1987, 67, 175–183. [Google Scholar] [CrossRef]
- Kirchhof, S.; Eisner, I.; Gierus, M.; Südekum, K.-H. Variation in the contents of crude protein fractions of different forage legumes during the spring growth. Grass Forage Sci. 2010, 65, 376–382. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, Q.; Ge, G.; Han, G.; Jia, Y. Influence of drought stress on afalfa yields and nutritional composition. BMC Plant Biol. 2018, 18, 13. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Andreuccetti, V.; Arduini, I.; Minieri, S.; Pampana, S. Field bean for forage and grain in short-season rainfed mediterranean conditions. Ital. J. Agron. 2018, 13, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Steinshamn, H. Effect of forage legumes on feed intake, milk production and milk quality–A review. Anim. Sci. Pap. Rep. 2010, 28, 195–206. [Google Scholar]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage; Chalcombe Publications: Southampton, UK, 1991; ISBN 0948617225. [Google Scholar]
- Mohler, C.L.; Liebman, M. Weed Productivity and Composition in Sole Crops and Intercrops of Barley and Field Pea. J. Appl. Ecol. 1987, 24, 685–699. [Google Scholar] [CrossRef]
- Frenda, A.S.; Ruisi, P.; Saia, S.; Frangipane, B.; Di Miceli, G.; Amato, G.; Giambalvo, D. The Critical Period of Weed Control in Faba Bean and Chickpea in Mediterranean Areas. Weed Sci. 2013, 61, 452–459. [Google Scholar] [CrossRef]
- Herbert, S.J.; Putnam, D.H.; Poos-Floyd, M.I.; Vargas, A.; Creighton, J.F. Forage Yield of Intercropped Corn and Soybean in Various Planting Patterns1. Agron. J. 1984, 76, 507–510. [Google Scholar] [CrossRef]
- Strydhorst, S.M.; King, J.R.; Lopetinsky, K.J.; Harker, K.N. Forage Potential of Intercropping Barley with Faba Bean, Lupin, or Field Pea. Agron. J. 2008, 100, 182–190. [Google Scholar] [CrossRef]
- Pampana, S.; Arduini, I.; Andreuccetti, V.; Mariotti, M. Fine-Tuning N Fertilization for Forage and Grain Production of Barley–Field Bean Intercropping in Mediterranean Environments. Agronomy 2022, 12, 418. [Google Scholar] [CrossRef]
- Mariotti, M.; Masoni, A.; Ercoli, L.; Arduini, I. Forage potential of winter cereal/legume intercrops in organic farming. Ital. J. Agron. 2006, 1, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Masoni, A.; Ercoli, L.; Arduini, I. Optimizing forage yield of durum wheat/field bean intercropping through N fertilization and row ratio. Grass Forage Sci. 2012, 67, 243–254. [Google Scholar] [CrossRef]
- Liponi, G.B.; Casini, L.; Gatta, D. Valutazione nutrizionale del favino (vicia faba minor) quale principale fonte proteica in diete per ovini: Risultati preliminari. Ann. Della Fac. Di Med. Vet. 2004, 57, 291–296. [Google Scholar]
- Louw, A.W. The Nutritive Value of Faba Bean Silage for Lactating Dairy Cows. Master’s Thesis, University of Stellenbosch, Stellenbosch, South Africa, 2009. Available online: http://hdl.handle.net/10019.1/4051 (accessed on 28 July 2022).
- Pursiainen, P.; Tuori, M. Effect of ensiling field bean, field pea and common vetch in different proportions with whole-crop wheat using formic acid or an inoculant on fermentation characteristics. Grass Forage Sci. 2008, 63, 60–78. [Google Scholar] [CrossRef]
- Borreani, G.; Chion, A.R.; Colombini, S.; Odoardi, M.; Paoletti, R.; Tabacco, E. Fermentative profiles of field pea (Pisum sativum), faba bean (Vicia faba) and white lupin (Lupinus albus) silages as affected by wilting and inoculation. Anim. Feed Sci. Technol. 2009, 151, 316–323. [Google Scholar] [CrossRef]
- Pisponen, A.; Pajumägi, S.; Mootse, H.; Karus, A.; Poikalainen, V. The lactose from Ricotta cheese whey: The effect of pH and concentration on size and morphology of lactose crystals. Dairy Sci. Technol. 2013, 93, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, M.; Fratini, F.; Cerri, D.; Andreuccetti, V.; Giglio, R.; Angeletti, F.G.S.; Turchi, B. Use of fresh scotta whey as an additive for alfalfa silage. Agronomy 2020, 10, 365. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, A.; Hill, J.; Leaver, J.D. Effect of stage of growth on the chemical composition, nutritive value and ensilability of whole-crop barley. Anim. Feed Sci. Technol. 2009, 152, 50–61. [Google Scholar] [CrossRef]
- Kung, L.; Ranjit, N.K. The Effect of Lactobacillus buchneri and Other Additives on the Fermentation and Aerobic Stability of Barley Silage. J. Dairy Sci. 2001, 84, 1149–1155. [Google Scholar] [CrossRef]
- Acosta, Y.M.; Stallings, C.C.; Polan, C.E.; Miller, C.N. Evaluation of Barley Silage Harvested at Boot and Soft Dough Stages. J. Dairy Sci. 1991, 74, 167–176. [Google Scholar] [CrossRef]
- Raffaella, T.; Calabrò, S.; Terzi, V.; Piccolo, V.; Guglielmelli, A.; Infascelli, F. In vitro fermentation of ten cultivars of barley silage. Ital. J. Anim. Sci. 2009, 8, 343–345. [Google Scholar] [CrossRef]
- Seppälä, A.; Rinne, M.; Huuskonen, A. Efficacy of different additives in ensiling faba bean and field pea based whole crop silages. Agric. Food Sci. 2019, 28, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Nadeau, E.; Hallin, O.; Jansson, J. Whole-crop legume/barley silages ensiled with different additives. In Proceedings of the XVth International Silage Conference, Madison, WI, USA, 27–29 July 2009; pp. 27–29. [Google Scholar]
- McAllister, T.A.; Hristov, A.N. The Fundamentals of Making Good Quality Silage. Adv. Dairy Technol. 2000, 12, 381–399. [Google Scholar]
- Preiti, G.; Calvi, A.; Romeo, M.; Badagliacca, G.; Bacchi, M. Seeding Density and Nitrogen Fertilization Effects on Agronomic Responses of Some Hybrid Barley Lines in a Mediterranean Environment. Agronomy 2021, 11, 1942. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants; BBCH Monograph, Federal Biological Research Centre for Agriculture and Forestry: Bonn, Germany; Julius Kühn-Institut (JKI): Quedlinburg, Germany, 2018; ISBN 978-3-95547-071-5. [Google Scholar]
- Griggs, T.C. Determining Forage Dry Matter Concentration with a Microwave Oven; AG/Forage Pasture/2005-01; Coop. Extention, Utah State University: Logan, UT, USA, 2005; pp. 1–2. [Google Scholar]
- Martillotti, F.; Antongiovanni, M.; Rizzi, L.; Santi, E.B.G. Metodi di analisi per gli alimenti d’impiego zootecnico. In Quaderni Metodologici; CNR-IPRA, Angeli: Milano, Italy, 1987; Volume 8. [Google Scholar]
- NRC. Nutrient Requirements of Dairy Cattle: Seventh Revised Edition, 2001; The National Academies Press: Washington, DC, USA, 2001; ISBN 978-0-309-06997-7. [Google Scholar]
- Rohweder, D.A.; Barnes, R.F.; Jorgensen, N. Proposed Hay Grading Standards Based on Laboratory Analyses for Evaluating Quality. J. Anim. Sci. 1978, 47, 747–759. [Google Scholar] [CrossRef]
- European Commission. Commission regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union 2009, 54, 2–54. [Google Scholar]
- Canale, A.; Valente, M.E.; Ciotti, A. Determination of volatile carboxylic acids (C1–C5i) and lactic acid in aqueous acid extracts of silage by high performance liquid chromatography. J. Sci. Food Agric. 1984, 35, 1178–1182. [Google Scholar] [CrossRef]
- Suardi, A.; Saia, S.; Alfano, V.; Rezaei, N.; Cetera, P.; Bergonzoli, S.; Pari, L. Pruning harvesting with modular towed chipper: Little effect of the machine setting and configuration on performance despite strong impact on wood chip quality. PLoS ONE 2021, 16, e0261810. [Google Scholar] [CrossRef]
- Bergonzoli, S.; Brambilla, M.; Romano, E.; Saia, S.; Cetera, P.; Cutini, M.; Toscano, P.; Bisaglia, C.; Pari, L. Feeding Emitters for Microirrigation with a Digestate Liquid Fraction up to 25 % Dilution Did Not Reduce Their Performance. Agronomy 2020, 10, 1150. [Google Scholar] [CrossRef]
- Pengelly, B.C.; Maass, B.L. Lablab purpureus (L.) Sweet—Diversity, potential use and determination of a core collection of this multi-purpose tropical legume. Genet. Resour. Crop Evol. 2001, 48, 261–272. [Google Scholar] [CrossRef]
- Goeser, J.P.; Heuer, C.R.; Crump, P.M. Forage fermentation product measures are related to dry matter loss through meta-analysis. Prof. Anim. Sci. 2015, 31, 137–145. [Google Scholar] [CrossRef] [Green Version]
- García-Chávez, I.; Meraz-Romero, E.; Castelán-Ortega, O.; Zaragoza Esparza, J.; Osorio Avalos, J.; Robles Jimenez, L.E.; González-Ronquillo, M. Corn Silage, Meta-Analysis of The Quality and Yield of Different Regions in the World. Preprints 2020, 2020100094. [Google Scholar] [CrossRef]
- Raseduzzaman, M.; Jensen, E.S. Does intercropping enhance yield stability in arable crop production? A meta-analysis. Eur. J. Agron. 2017, 91, 25–33. [Google Scholar] [CrossRef]
- de Rezende, R.P.; de Oliveira Golin, H.; de Abreu, V.L.D.S.; de Faria Theodoro, G.; Franco, G.L.; Brumatti, R.C.; Fernandes, P.B.; de Lucca Bento, A.L.; Rocha, R.F.A.T. Does intercropping maize with forage sorghum effect biomass yield, silage bromatological quality and economic viability? Res. Soc. Dev. 2020, 9, e46942818. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, A.G.; Dear, B.S.; Morris, S.G. An evaluation of the yield and quality of oat–legume and ryegrass–legume mixtures and legume monocultures harvested at three stages of growth for silage. Aust. J. Exp. Agric. 2007, 47, 25–38. [Google Scholar] [CrossRef]
- Shoaib, M.; Ayub, M.; Shehzad, M.; Akhtar, N.; Tahir, M.; Arif, M. Dry matter yield and forage quality of oat, barley and canola mixture. Pakistan J. Agric. Sci. 2014, 51, 443–449. [Google Scholar]
- Jacobs, J.L.; Ward, G.N. Effect of intercropping forage peas (Pisum sativum L.) with winter wheat (Tritium vulgare L.) or triticale (Triticale hexaploide Lart.) on DM yield, nutritive characteristics when harvested at different stages of growth. Anim. Prod. Sci. 2012, 52, 949–958. [Google Scholar] [CrossRef]
- Hernández-Ortega, M.; Heredia-Nava, D.; Espinoza-Ortega, A.; Sánchez-Vera, E.; Arriaga-Jordán, C.M. Effect of silage from ryegrass intercropped with winter or common vetch for grazing dairy cows in small-scale dairy systems in Mexico. Trop. Anim. Health Prod. 2011, 43, 947–954. [Google Scholar] [CrossRef]
- Maxin, G.; Andueza, D.; Le Morvan, A.; Baumont, R. Effect of intercropping vetch (Vicia sativa L.), field pea (Pisum sativum L.) and triticale (X Triticosecale) on dry-matter yield, nutritive and ensiling characteristics when harvested at two growth stages. Grass Forage Sci. 2017, 72, 777–784. [Google Scholar] [CrossRef]
- Prithiviraj, B.; Carruthers, K.; Fe, Q.; Cloutier, D.; Martin, R.C.; Smith, D.L. Intercropping of corn with soybean and lupin for silage: Effect of seeding date on yield and quality. J. Agron. Crop Sci. 2000, 185, 129–136. [Google Scholar] [CrossRef]
- Brown, A.N.; Ferreira, G.; Teets, C.L.; Thomason, W.E.; Teutsch, C.D. Nutritional composition and in vitro digestibility of grass and legume winter (cover) crops. J. Dairy Sci. 2018, 101, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Titterton, M.; Maasdorp, B.V. Nutritional improvement of maize silage for dairying: Mixed crop silages from sole and intercropped legumes and a long season variety of maize. 2. Ensilage. Anim. Feed Sci. Technol. 1997, 69, 263–270. [Google Scholar] [CrossRef]
- Soufan, W.; Al-Suhaibani, N.A. Optimizing yield and quality of silage and hay for pea–barley mixtures ratio under irrigated arid environments. Sustainability 2021, 13, 13621. [Google Scholar] [CrossRef]
- Wang, S.; Yuan, X.; Dong, Z.; Li, J.; Shao, T. Effect of ensiling corn stover with legume herbages in different proportions on fermentation characteristics, nutritive quality and in vitro digestibility on the Tibetan Plateau. Grassl. Sci. 2017, 63, 236–244. [Google Scholar] [CrossRef]
- Goyal, M.; Tiwana, U.S. Ensiling Legume with Cereal Fodder Influences Quality of Silage Mixtures. Indian J. Anim. Nutr. 2016, 33, 228–232. [Google Scholar] [CrossRef]
- Halmemies-Beauchet-Filleau, A.; Rinne, M.; Lamminen, M.; Mapato, C.; Ampapon, T.; Wanapat, M.; Vanhatalo, A. Review: Alternative and novel feeds for ruminants: Nutritive value, product quality and environmental aspects. Animal 2018, 12, s295–s309. [Google Scholar] [CrossRef] [Green Version]
- Pampana, S.; Masoni, A.; Mariotti, M.; Ercoli, L.; Arduini, I. Nitrogen fixation of grain legumes differs in response to nitrogen fertilisation. Exp. Agric. 2018, 54, 66–82. [Google Scholar] [CrossRef]
- Galanopoulou, K.; Lithourgidis, A.S.; Dordas, C.A. Intercropping of faba bean with barley at various spatial arrangements affects dry matter and N Yield, Nitrogen Nutrition Index, and interspecific competition. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 1116–1127. [Google Scholar] [CrossRef]
- Guevara-Oquendo, V.H.; Christensen, D.A.; Refat, B.; Rodriguez-Espinosa, M.E.; Feng, X.; Yu, P. Production performance and metabolic characteristics of cows fed whole plant faba bean silage in comparison with barley and corn silage. Can. J. Anim. Sci. 2022, 102, 145–154. [Google Scholar] [CrossRef]
- Dewhurst, R. Milk production from silage: Comparison of grass, legume and maize silages and their mixtures. Agric. Food Sci. 2013, 22, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Huuskonen, A.; Pesonen, M. Performance and meat quality of Nordic Red and Aberdeen Angus bulls offered faba bean or field pea based whole crop legumecereal silages. Agric. Food Sci. 2016, 25, 1–12. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; Dickhoefer, U. Effects of tropical legume silages on intake, digestibility and performance in large and small ruminants: A review. Grass Forage Sci. 2018, 73, 26–39. [Google Scholar] [CrossRef]
- Li, P.; Zhang, Y.; Gou, W.; Cheng, Q.; Bai, S.; Cai, Y. Silage fermentation and bacterial community of bur clover, annual ryegrass and their mixtures prepared with microbial inoculant and chemical additive. Anim. Feed Sci. Technol. 2019, 247, 285–293. [Google Scholar] [CrossRef]
- Wang, S.; Li, J.; Zhao, J.; Dong, Z.; Shao, T. Exploring the ensiling characteristics and bacterial community of red clover inoculated with the epiphytic bacteria from temperate gramineous grasses. J. Appl. Microbiol. 2022, 132, 177–188. [Google Scholar] [CrossRef]
- Li, H.; Zeng, T.; Du, Z.; Dong, X.; Xin, Y.; Wu, Y.; Huang, L.; Liu, L.; Kang, B.; Jiang, D.; et al. Assessment on the Fermentation Quality and Bacterial Community of Mixed Silage of Faba Bean With Forage Wheat or Oat. Front. Microbiol. 2022, 13, 875819. [Google Scholar] [CrossRef]
- Wang, S.; Shao, T.; Li, J.; Zhao, J.; Dong, Z. Fermentation Profiles, Bacterial Community Compositions, and Their Predicted Functional Characteristics of Grass Silage in Response to Epiphytic Microbiota on Legume Forages. Front. Microbiol. 2022, 13, 830888. [Google Scholar] [CrossRef]
- Nykänen, A.; Jauhiainen, L.; Rinne, M. Biomass production and feeding value of whole-crop cereal-legume silages. Agron. Res. 2009, 7, 684–690. [Google Scholar]



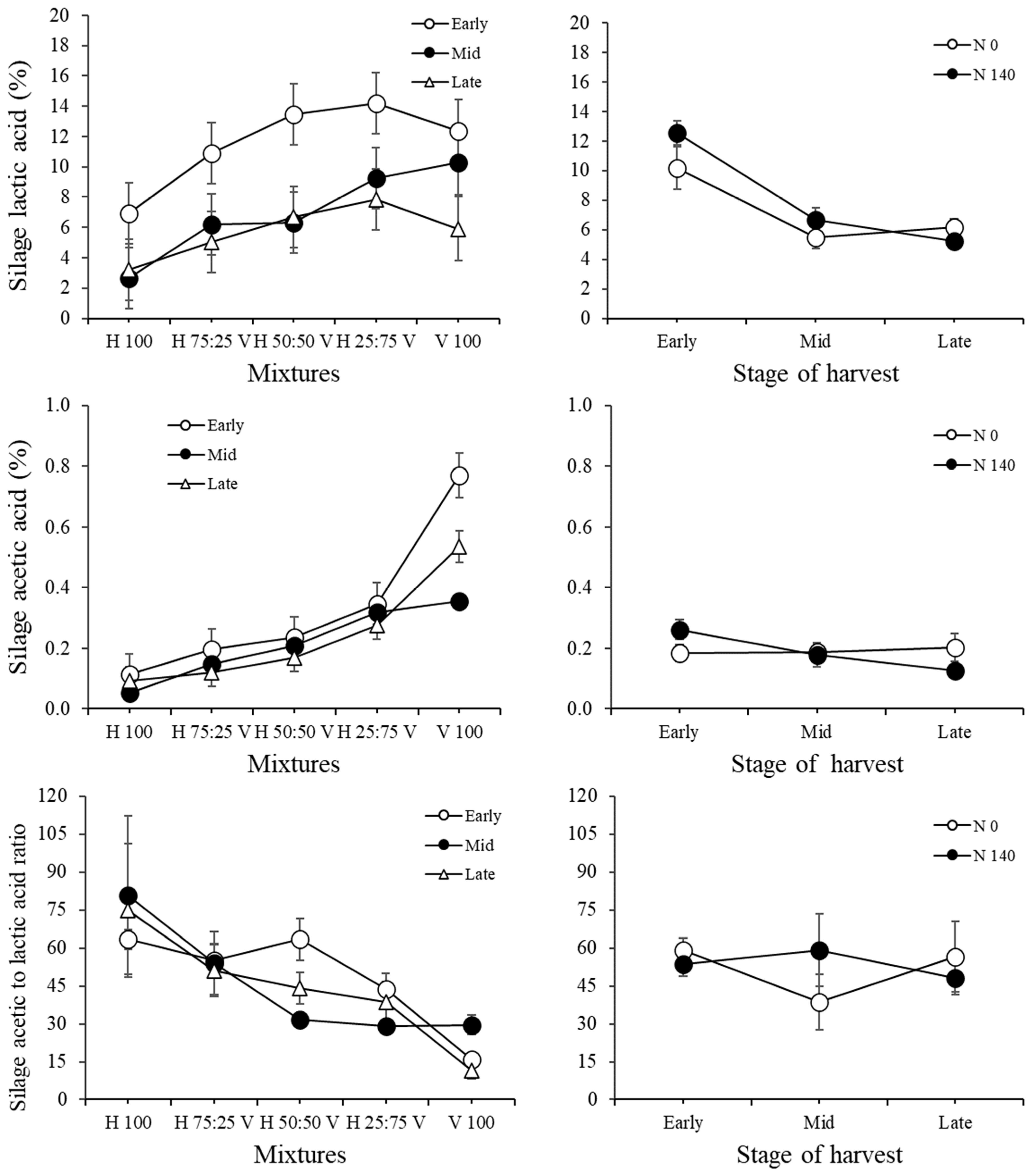



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SH | MX | SH × MX | NF | SH × NF | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | |
| Dry matter (DM) (g (100 g FM)−1) | 42.05 | <0.0001 | 488.67 | <0.0001 | 22.63 | <0.0001 | 4.36 | 0.004 | 6.58 | <0.0001 |
| Crude protein (CP) (g (100 g DM)−1) | 8.23 | 0.0008 | 794.19 | <0.0001 | 2.69 | 0.0145 | 31.72 | <0.0001 | 10.98 | <0.0001 |
| Ether extracts (EE) (g (100 g DM)−1) | 0.11 | 0.9002 | 30.92 | <0.0001 | 2.06 | 0.0562 | 1.59 | 0.1892 | 3.98 | 0.0009 |
| Ash (g (100 g DM)−1) | 19.1 | <0.0001 | 6.98 | 0.0001 | 1.92 | 0.0754 | 19.81 | <0.0001 | 6.06 | <0.0001 |
| Neutral-detergent fiber (NDF) (g (100 g DM)−1) | 0.47 | 0.6281 | 29.76 | 0.0002 | 9.69 | <0.0001 | 1.31 | 0.2826 | 2.06 | 0.0649 |
| Acid-detergent fiber (ADF) (g (100 g DM)−1) | 0.58 | 0.5629 | 27.66 | <0.0001 | 8.73 | <0.0001 | 2.72 | 0.0389 | 1.71 | 0.1178 |
| Acid-detergent lignin (ADL) (g (100 g DM)−1) | 2.47 | 0.0942 | 141.78 | <0.0001 | 13.52 | <0.0001 | 2.85 | 0.0343 | 1.74 | 0.1144 |
| Total digestible nutrients (TDNs) (g (100 g DM)−1) | 28.34 | <0.0001 | 61.04 | <0.0001 | 5.77 | <0.0001 | 2.5 | 0.0533 | 2.37 | 0.0288 |
| Relative Feed Value (RFV) (%) | 0 | 1 | 6.68 | 0.0002 | 7.77 | <0.0001 | 1.71 | 0.1618 | 1.91 | 0.0768 |
| Water-soluble carbohydrates (WSCs) (g (100 g DM)−1) | 2.79 | 0.0705 | 3.95 | 0.0192 | 10.85 | <0.0001 | 22.4 | <0.0001 | 3.07 | 0.007 |
| WSC to CP ratio | 76.69 | 0.0724 | 33.04 | <0.0001 | 5.86 | <0.0001 | 70.74 | <0.0001 | 6.65 | <0.0001 |
| Hemicellulose (g (100 g DM)−1) | 0.33 | 0.718 | 166.44 | <0.0001 | 5.5 | <0.0001 | 1.44 | 0.2326 | 0.86 | 0.5583 |
| Cellulose (g (100 g DM)−1) | 0.21 | 0.8109 | 13.69 | <0.0001 | 9.35 | <0.0001 | 2.33 | 0.0680 | 1.57 | 0.1551 |
| Truly digestible non-fibrous carbohydrates (tdNFC) (g (100 g DM)−1) | 0.86 | 0.4307 | 5.53 | 0.0069 | 9.25 | <0.0001 | 1.92 | 0.1239 | 2.72 | 0.0154 |
| SH | MX | SH × MX | NF | SH × NF | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | |
| pH | n.a | <0.0001 | n.a | <0.0001 | n.a | <0.0001 | 35.69 | <0.0001 | 41.1 | <0.0001 |
| Dry matter (DM) (g (100 g FM)−1) | 13.8 | <0.0001 | 157.11 | <0.0001 | 4.41 | 0.0004 | 5.73 | 0.0007 | 3.68 | 0.002 |
| Crude protein (CP) (g (100 g DM)−1) | 1.35 | 0.267 | 307.53 | <0.0001 | 2.63 | 0.0164 | 10.68 | <0.0001 | 2.32 | 0.0321 |
| Ether extracts (EE) (g (100 g DM)−1) | 4.63 | 0.014 | 12.26 | <0.0001 | 2.16 | 0.0451 | 0.88 | 0.4795 | 0.49 | 0.8581 |
| Ash (g (100 g DM)−1) | 16.54 | 0.1713 | 8.18 | 0.0011 | 18.53 | <0.0001 | 35.21 | <0.0001 | 5.96 | <0.0001 |
| Neutral-detergent fiber (NDF) (g (100 g DM)−1) | 0.07 | 0.9286 | 87.6 | <0.0001 | 2.1 | 0.0521 | 3.61 | 0.0111 | 2.81 | 0.0112 |
| Acid-detergent fiber (ADF) (g (100 g DM)−1) | 0.53 | 0.5899 | 6.34 | 0.001 | 1.45 | 0.2002 | 4.27 | 0.0047 | 2.96 | 0.0085 |
| Acid-detergent lignin (ADL) (g (100 g DM)−1) | 16.59 | <0.0001 | 73.41 | <0.0001 | 28.9 | <0.0001 | 7.52 | <0.0001 | 1.87 | 0.0837 |
| Total digestible nutrients (TDNs) (g (100 g DM)−1) | 0.32 | 0.7265 | 6.34 | 0.001 | 1.45 | 0.2002 | 4.27 | 0.0047 | 2.96 | 0.0085 |
| Relative Feed Value (RFV) (%) | 0 | 1 | 84.44 | <0.0001 | 1.14 | 0.354 | 4.08 | 0.0059 | 2.55 | 0.0197 |
| Water-soluble carbohydrates (WSCs) (g (100 g DM)−1) | 0.08 | 0.9193 | 4.16 | 0.0053 | 5.06 | 0.0001 | 6.54 | 0.0002 | 1.42 | 0.2109 |
| WSC to CP ratio | 4.42 | 0.0168 | 19.45 | <0.0001 | 6.32 | <0.0001 | 11.32 | <0.0001 | 1.92 | 0.0762 |
| Hemicellulose (g (100 g DM)−1) | 0.01 | 0.9864 | 119.16 | <0.0001 | 3.52 | 0.0026 | 1.86 | 0.1321 | 5.95 | <0.0001 |
| Cellulose (g (100 g DM)−1) | 0.04 | 0.9626 | 21.25 | <0.0001 | 5.6 | <0.0001 | 3.68 | 0.0106 | 2.45 | 0.0257 |
| Truly digestible non-fibrous carbohydrates (tdNFC) (g (100 g DM)−1) | 0.32 | 0.726 | 19.69 | <0.0001 | 4.41 | 0.0004 | 6.72 | 0.0002 | 2.9 | 0.0092 |
| Lactic acid (g (100 g DM)−1) | 2.66 | 0.0792 | 32.6 | <0.0001 | 3.16 | 0.0056 | 5.41 | 0.0011 | 7.66 | <0.0001 |
| Acetic acid (g (100 g DM)−1) | 1.36 | 0.4516 | 69.88 | <0.0001 | 7.25 | <0.0001 | 4.09 | 0.0063 | 6.99 | <0.0001 |
| Propionic acid (g (100 g DM)−1) | 28.05 | 0.1323 | 3.16 | 0.0542 | 7.01 | <0.0001 | 0.31 | 0.8677 | 3.3 | 0.0046 |
| Butyric acid (g (100 g DM)−1) | 26.31 | <0.0001 | 15.97 | <0.0001 | 2.13 | 0.0494 | 1.26 | 0.298 | 3.48 | 0.0029 |
| Isovaleric acid (g (100 g DM)−1) | 33.34 | <0.0001 | 49.66 | <0.0001 | 25.42 | <0.0001 | 10.62 | <0.0001 | 2.25 | 0.0376 |
| Valeric acid (g (100 g DM)−1) | 324.83 | <0.0001 | 16.33 | <0.0001 | 9.2 | <0.0001 | 0.89 | 0.4775 | 1.34 | 0.2443 |
| Acetic Acid to Lactic Acid ratio | 0 | 1 | 7.84 | <0.0001 | 0.88 | 0.5361 | 3.7 | 0.0098 | 3.59 | 0.0021 |
| Total Volatile Fatty Acids (TVFA) (g (100 g DM)−1) | 2.36 | 0.104 | 33.02 | <0.0001 | 2.36 | 0.0306 | 4.92 | 0.002 | 6.7 | <0.0001 |
| Treatment | Butyric Acid | Isovaleric Acid | Valeric Acid |
|---|---|---|---|
| Mixtures | |||
| H 100 | 0.622 b | 0.244 a | 0.026 d. |
| H 75:25 V | 0.628 b | 0.189 b | 0.033 bc |
| H 50:50 V | 0.712 b | 0.050 c | 0.032 c. |
| H 25:75 V | 0.907 a | 0.026 c | 0.037 ab |
| V 100 | 0.958 a | 0.032 c | 0.040 a. |
| Stage of harvest | |||
| Early | 0.896 a | 0.181 a | 0.050 a |
| Mid | 0.849 a | 0.084 b | 0.037 b |
| Late | 0.551 b | 0.059 b | 0.014 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angeletti, F.G.S.; Mariotti, M.; Tozzi, B.; Pampana, S.; Saia, S. Herbage and Silage Quality Improved More by Mixing Barley and Faba Bean Than by N Fertilization or Stage of Harvest. Agronomy 2022, 12, 1790. https://doi.org/10.3390/agronomy12081790
Angeletti FGS, Mariotti M, Tozzi B, Pampana S, Saia S. Herbage and Silage Quality Improved More by Mixing Barley and Faba Bean Than by N Fertilization or Stage of Harvest. Agronomy. 2022; 12(8):1790. https://doi.org/10.3390/agronomy12081790
Chicago/Turabian StyleAngeletti, Francesco G. S., Marco Mariotti, Beatrice Tozzi, Silvia Pampana, and Sergio Saia. 2022. "Herbage and Silage Quality Improved More by Mixing Barley and Faba Bean Than by N Fertilization or Stage of Harvest" Agronomy 12, no. 8: 1790. https://doi.org/10.3390/agronomy12081790
APA StyleAngeletti, F. G. S., Mariotti, M., Tozzi, B., Pampana, S., & Saia, S. (2022). Herbage and Silage Quality Improved More by Mixing Barley and Faba Bean Than by N Fertilization or Stage of Harvest. Agronomy, 12(8), 1790. https://doi.org/10.3390/agronomy12081790