Abstract
The development of varieties with strong tolerance is one of the important strategies to diminish the negative impact of chilling stress during heading on the spikelet fertility and yield formation of late-season rice. However, whether such genetic improvement has been made in inbred late rice lines in China is not clear. In the present study, three late-season inbred rice varieties, Xiangwanxian2 (XWX2, released in 1988), Xiangwanxian8 (XWX8, released in 1998) and Xiangwanxian17 (XWX17, released in 2008) were subjected to moderate (20 °C) and extreme (17 °C) chilling stress during heading, and the grain yield components and flowering-related traits of the three varieties in response to different temperature were investigated. The results showed that the newly released inbred late rice variety XWX17, demonstrated better chilling tolerance during heading than the early released varieties with respect to higher grain filling percentage. The improved grain filling percentage in XWX17 might be the results of increased spikelet fertility, which was attributed to the increase in pollen viability, anther dehiscence length and anther volume. In addition, the SPAD value and the chlorophyll a content of the flag leaf can be used as indicators to predict the rice spikelet fertility when suffering from chilling stress during heading. The present study provides evidence that the genetic approach has been made to improve the chilling tolerance of inbred late rice lines during heading; however, further research is needed to explore the physiological and molecular mechanism underlying the relationship between leaf characteristics and function with rice spikelet fertility.
1. Introduction
Rice (Oryza sativa L.) is the most important crop in the world and the main food for more than half of the population. It is also the main food crop in China. The Yangtze River Basin is the major rice production region in China, of which the double-season rice system is the dominant rice production system and is considered as an efficient system to improve multiple-crop index and total rice production. In the double-season rice system, the late-season rice is transplanted to the field in July after the harvesting of the early season rice, and it is flowering in the middle of September, during which the temperature is decreased rapidly. It has been reported that the daily mean temperature in the Yangtze River Basin during September is 21.8 °C, and the cold dew (the duration of low temperature that is below 20 °C and lasts for more than three days) frequently occurs in this area. The low temperature would cause chilling stress to the late-season rice, resulting in heading difficulty, poor spikelet fertility, which inhibits the transportation of photosynthetic products, and leads to the decrease in seed setting rate and yield reduction.
Rice originated in tropical regions and is extremely sensitive to temperature during heading and flowering. It has been reported that ambient temperature equal to or lower than 20 °C lasting for 3 days during flowering would seriously affect the seed setting rate for rice [1]. Jia et al. [2] reported that low-temperature treatment, at 17 °C, with cold water irrigation at the booting stage significantly reduced rice grain yield. The negative effects of low temperature during flowering on the grain filling percentage and grain yield were mainly attributed to the decreased spikelet fertility, which might be the results of short anther dehiscence, poor pollen grains, and low pollen germination on stigmas [3,4]. In addition, the reduced panicle exertion was also reported to be responsible for the decreased spikelet fertility under chilling stress [5]. In addition, the chilling stress might accelerate the production of reactive oxygen species (ROS) [6] and induce the fast degradation of chlorophyll in leaf [7,8], which ultimately reduce the production and translocation of photosynthetic products from the leaf to rice grain.
To diminish the negative effects of chilling stress during heading on the grain yield formation of late-season rice, it is of importance to select varieties with strong tolerance to low temperature stress. Several studies have reported the varietal differences in rice against temperature stress [9]. In general, indica species are more tolerant to high temperature; in contrast, japonica species are more tolerant to low temperature stress [10]. In recent years, the genetic approaches have been made to screen high chilling-tolerant germplasms and to introduce chilling-tolerant genes into modern varieties via hybrid rice breeding. Several hybrid late rice varieties that possess relatively high chilling resistance have been successfully bred and adopted in the Yangtze River Basin of China. Nevertheless, it is not clear whether the breeding progress of inbred late rice varieties has paid attention to enhancing the chilling resistance at the heading stage of late rice. In the present study, three late-season inbred rice varieties, Xiangwanxian2 (XWX2, released in 1988), Xiangwanxian8 (XWX8, released in 1998) and Xiangwanxian17 (XWX17, released in 2008) were subjected to moderate (20 °C) and extreme (17 °C) chilling stress during heading in a greenhouse experiment. The grain yield and flowering-related traits of the three varieties in response to different temperature were investigated. The objectives of the present study were to evaluate the variances in chilling tolerance among three inbred late rice varieties that were released from different decades regarding grain yield, grain filling percentage and flowering traits, and to explore whether the genetic improvement in inbred late rice has been focused on the chilling tolerance during heading.
2. Materials and Methods
2.1. Plant Material
Three inbred rice varieties, Xiangwanxian 17 (XWX17), Xiangwanxian8 (XWX8), and Xiangwanxian2 (XWX2), from the Yangtze River valley in China were selected as the test materials, which were provided by Hunan Germplasm Resources Bank. The detailed information for the tested varieties was shown in Table 1.

Table 1.
The detailed information for the tested varieties.
2.2. Experimental Design
A pot experiment was conducted in 2020 at the Rice Research Institute of Hunan Agricultural University. The rice seeds of the three varieties were sown in plastic seedling trays with wet paddy soil on 28 June 2020. Three-leaf seedlings were transplanted in to a plastic pot (22.5 cm height, 29 cm top diameter) on 23 July, which contains a mixture of 7 kg clay soil. Phosphate fertilizer (1.00 g P pot−1) in the form of calcium superphosphate and potassium fertilizer (1.60 g K pot−1) were applied as basal fertilizer. A total of 1.60 g N pot−1 was equally spilt three times as basal fertilizer, tillering fertilizer, and panicle fertilizer, respectively. The potted plants were placed in a plastic greenhouse with three replication, and each replication contained 10 pots. All the pots were watered with tap water, and the pests, diseases and weeds were strictly controlled during the entire growing season.
In each variety, when 50% of the rice panicle started exertion and the exerted length reached 1 cm, the exerted panicles of the rice plants were tagged, and the pots were then transferred to three individual growth chambers for temperature treatments. The daytime temperature was set as 22 °C, 19 °C and 25 °C from 6:00~18:00 for moderate chilling stress (MCS), extreme chilling stress (ECS) and normal temperature control (NT), respectively, whereas the corresponded nighttime temperature was set as 18 °C, 15 °C and 21 °C from 18:00~6:00, respectively. After 5 days of temperature treatment, all the pots were transferred to the plastic greenhouse and grown to maturity.
2.3. Observations
2.3.1. Spikelet Fertility and Yield Components
At maturity, three plants from three pots were harvested. After the panicle numbers were counted, all the grain were manually threshed and were then separated to filled grains, half-filled grains, and empty grains using tap water and an air-flow machine. The spikelet fertility, total spikelet numbers, grain filling percentage and 1000-grain weight was then determined according to the methods reported by [11].
2.3.2. Pollen Viability
At 0 and 5 days after temperature treatments, six spikelets were collected from the upper, middle and lower parts of one panicle and at least 20 panicles were selected in each replication. Pollen viability was observed by dyeing with 1% I2-Ki solution. Abnormal pollen grains with no staining, shallow staining, and partial staining, and shrunken or hollow were recognized as sterile pollen, and dark and full round pollen grains were recognized as fertile pollen [12].
2.3.3. Anther Characteristics
At 5 days after temperature treatments, at least 100 spikelets from each replication were sampled, and the anther length, anther width and dehiscence length were measured using a depth of field microscope (Zoom-Smart 5, Carl Zeiss AG, Oberkochen, Germany).
2.3.4. SPAD, Chlorophyll Assay
During the measurement, the instruments SPAD-502 Plus were employed, and flag leaves were selected for the SPAD measurement. Five flag leaves of rice were randomly selected to measure SPAD in each plot, and there were ten SPAD measurement data in the same period for each treatment [13].
The leaf samples were mixed with sodium phosphate buffer (50 mM, pH 6.8) and ground in an ice bath. The supernatant was mixed with 95% ethanol and kept in the dark for 30 min before being centrifuged at 1000× g under 4 °C for 15 min. The absorbance at wavelengths 665 nm and 649 nm was measured using a spectrophotometer (U-2900, Hitachi, Tokyo, Japan) [14].
2.4. Statistical Analysis
A one-way analysis of variance was conducted to test the effects of low temperature stress at flowering stage on flowering, fruiting, and physiology of three late rice varieties using IBM SPSS Statistics 20 (IBM, Inc., Chicago, IL, USA). Different alphabetical letters are used in figures and tables for showing significant differences.
3. Results
3.1. Yield Components
The variances in yield components of the three varieties in response to different temperature treatment are shown in Table 2. The chilling stress during flowering progressively decreased the grain filling percentage. When compared with normal temperature control, the grain filling percentage was decreased by 2.93–13.14% and 23.19–31.16% under 20 °C and 17 °C, respectively. Meanwhile, significant variances were found among different varieties responding to temperature treatments. The moderate chilling stress (20 °C) reduced the grain filling percentage of XWX2 and XWX8 when compared with normal temperature control (23 °C), but such effects were statistically insignificant (p > 0.05) in XWX17. In addition, although the extreme chilling stress (17 °C) significantly reduced the grain filling of the three varieties, such reduction was smaller in XWX17 than that in XW2 and XWX8. In addition, no variance was found on panicle numbers, spikelet number per panicle and grain weight among different varieties and temperature treatments.
Table 2.
The grain yield components of three inbred late varieties under different temperature treatments.
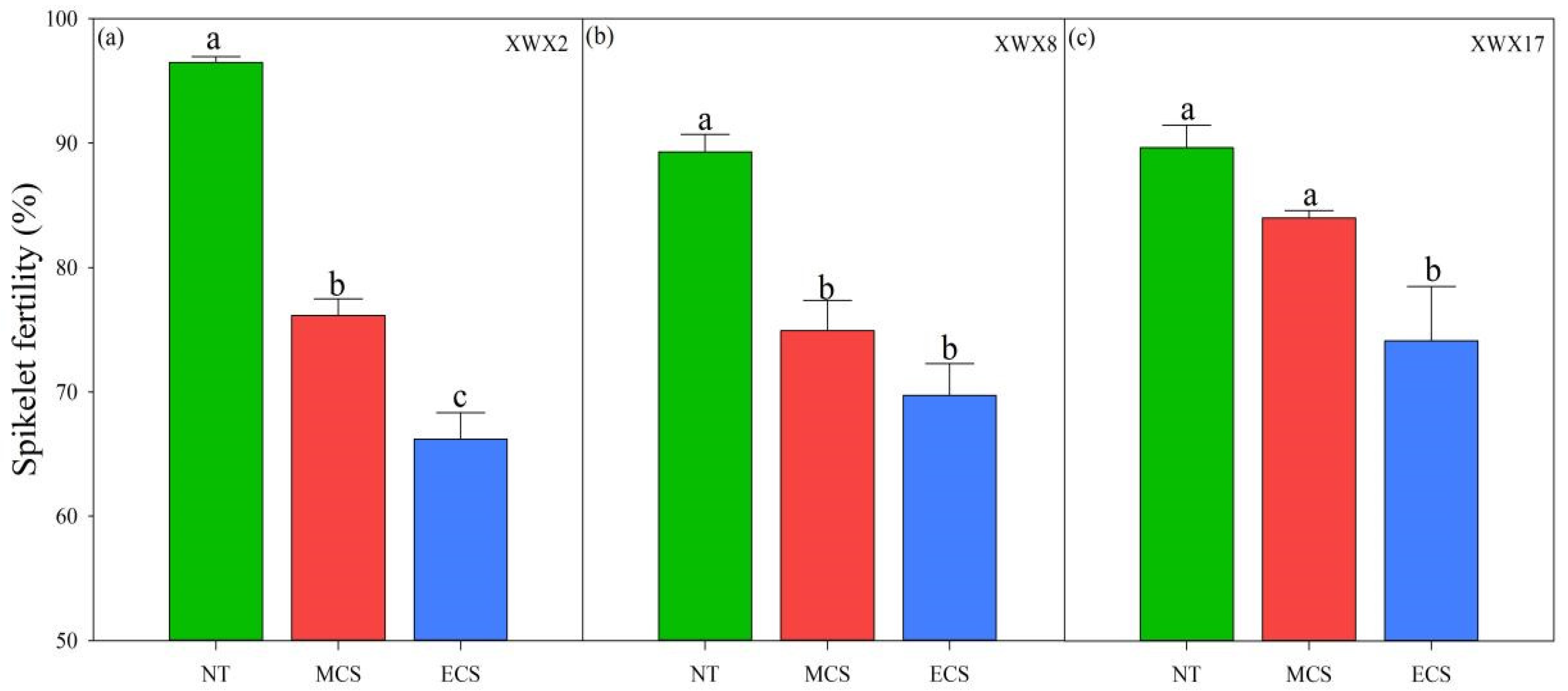
3.2. Spikelet Fertility
Significant variations in spikelet fertility were observed between varieties and temperature treatments (Figure 1). In XWX2 and XWX8, both moderate (20 °C) and extreme chilling stress significantly decreased the spikelet fertility. Compared with normal temperature control, the spikelet fertility was reduced by 6.29–21.07% and 17.32–31.39% at 20 °C and 17 °C, respectively. In contrast, the XWX17 exhibited stronger tolerance to moderate chilling stress, as the spikelet fertility at 20 °C showed no significant difference to that of 23 °C. Moreover, in addition, XWX17 showed a smaller decrease in spikelet fertility than XWX 2 and XWX8 under extreme chilling stress.
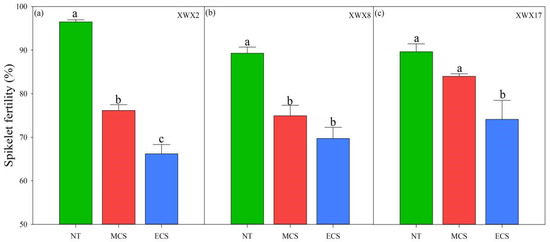
Figure 1.
The spikelet fertility of three inbred late varieties under different temperature treatments. (a) XWX2. (b) XWX8. (c) XWX17. XWX2: Xiangwanxian2; XWX8: Xiangwanxian8; XWX17: Xiangwanxian17; NT: Normal temperature control; MCS: Moderate chilling stress; ECS: Extreme chilling stress. Different lowercase letters denote statistical differences among treatments of a cultivar at the 5% level according to LSD test. Error bars above mean indicate standard error (n = 3).
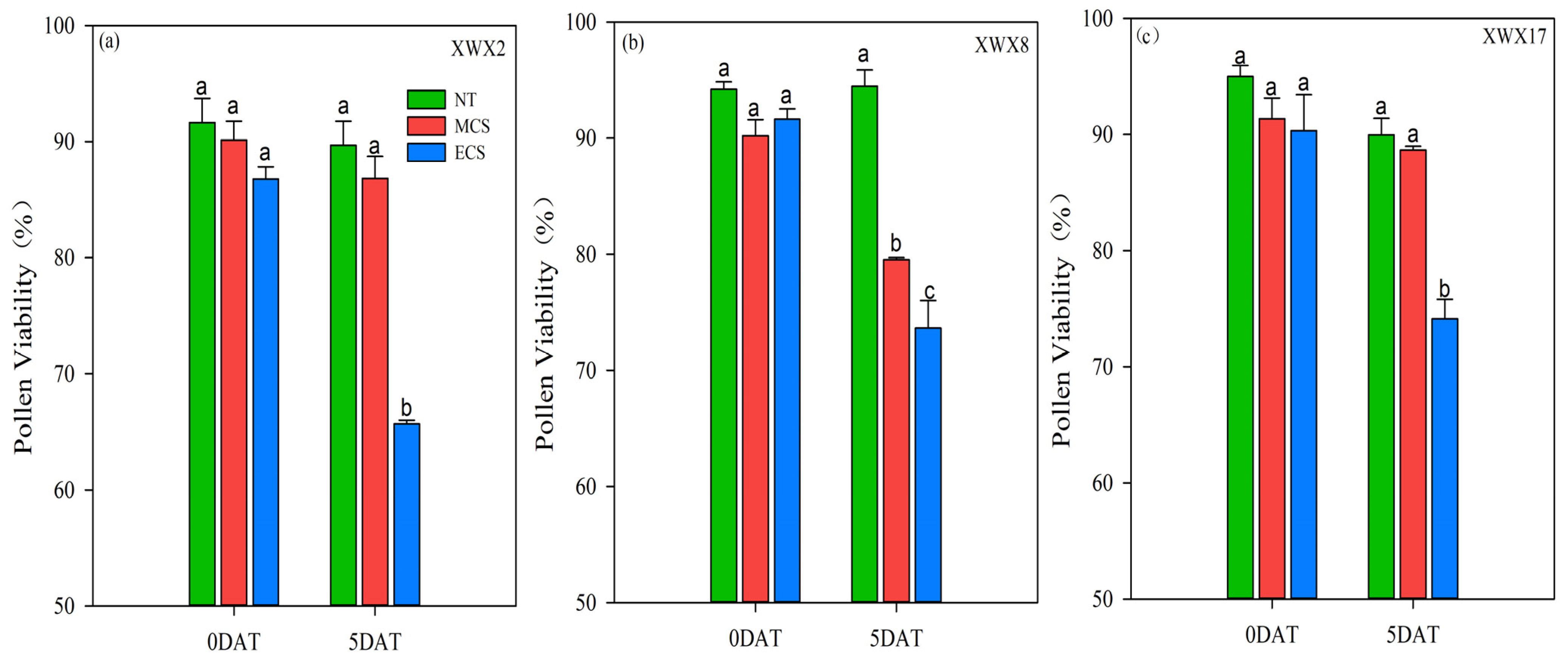
3.3. Pollen Viability
The pollen viability of the three late rice varieties was inhibited after 5 days of chilling stress treatments (Figure 2). When compared with NT, the pollen viability of XWX2, XWX8 and XWX17 was decreased by 3.67%, 11.83% and 2.96%, respectively, at MCS, and was decreased by 24.32%, 19.64% and 17.93%, respectively, at ECS. In comparison with XWX8, the XWX2 and XWX17 showed stronger resistance to MCS because the decrease in pollen viability did not reach a significant level. In addition, although the pollen viability of the three varieties was significantly reduced at ECS, the decreased level was lower in XWX17 than that in XWX2 and XWX8, as compared with NT.
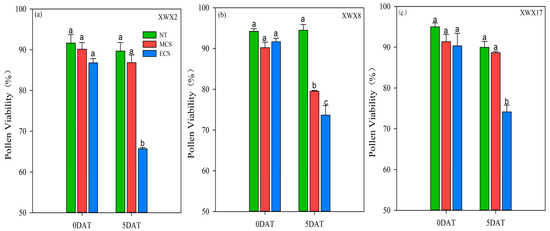
Figure 2.
The pollen viability of three inbred late varieties under different temperature treatments. (a) XWX2. (b) XWX8. (c) XWX17. DAT: Days after temperature treatments; XWX2: Xiangwanxian2; XWX8: Xiangwanxian8; XWX17: Xiangwanxian17; NT: Normal temperature control; MCS: Moderate chilling stress; ECS: Extreme chilling stress. Different lowercase letters denote statistical differences among treatments of a cultivar at the 5% level according to LSD test. Error bars above mean indicate standard error (n = 3).
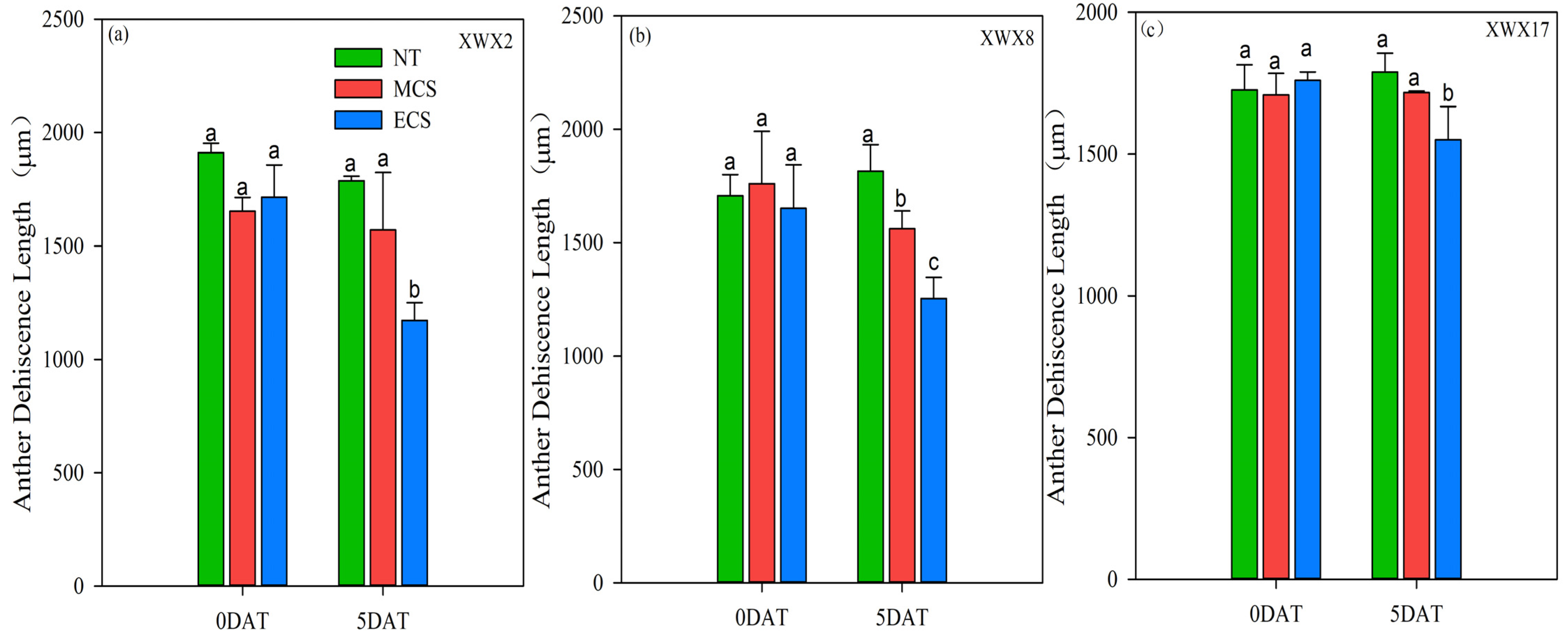
3.4. Anther Characters
The changes in anther characters of the three late rice varieties under different temperature treatments regarding anther width, anther length and anther dehiscence were shown in Figure 3 and Figure 4. The chilling stress reduced the anther dehiscence length of the three late rice varieties (Figure 3). The varietal differences were similar to that observed in spikelet fertility and pollen viability. The anther dehiscence length of XWX2 and XWX8 was significantly reduced by 5.00% and 11.31%, respectively, at MCS, as compared with NT, but such a reduction did not reach a significant level in XWX17. In addition, the anther dehiscence length of XWX17 was decreased by 11.91% at ECS, which was smaller than the decrease of 31.68% and 24.09% observed in XWX2 and XWX8, respectively, as compared with NT.
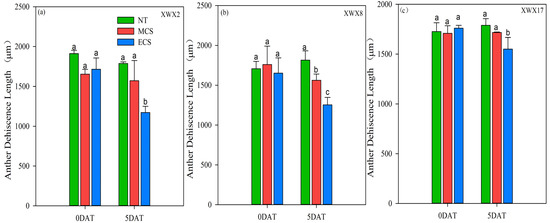
Figure 3.
The anther dehiscence length of three inbred late varieties under different temperature treatments. (a) XWX2. (b) XWX8. (c) XWX17. DAT: Days after temperature treatments; XWX2: Xiangwanxian2; XWX8: Xiangwanxian8; XWX17: Xiangwanxian17; NT: Normal temperature control; MCS: Moderate chilling stress; ECS: Extreme chilling stress. Different lowercase letters denote statistical differences among treatments of a cultivar at the 5% level according to LSD test. Error bars above mean indicate standard error (n = 3).
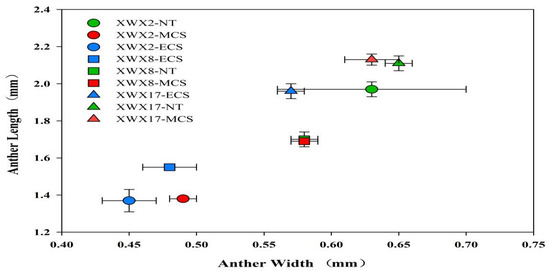
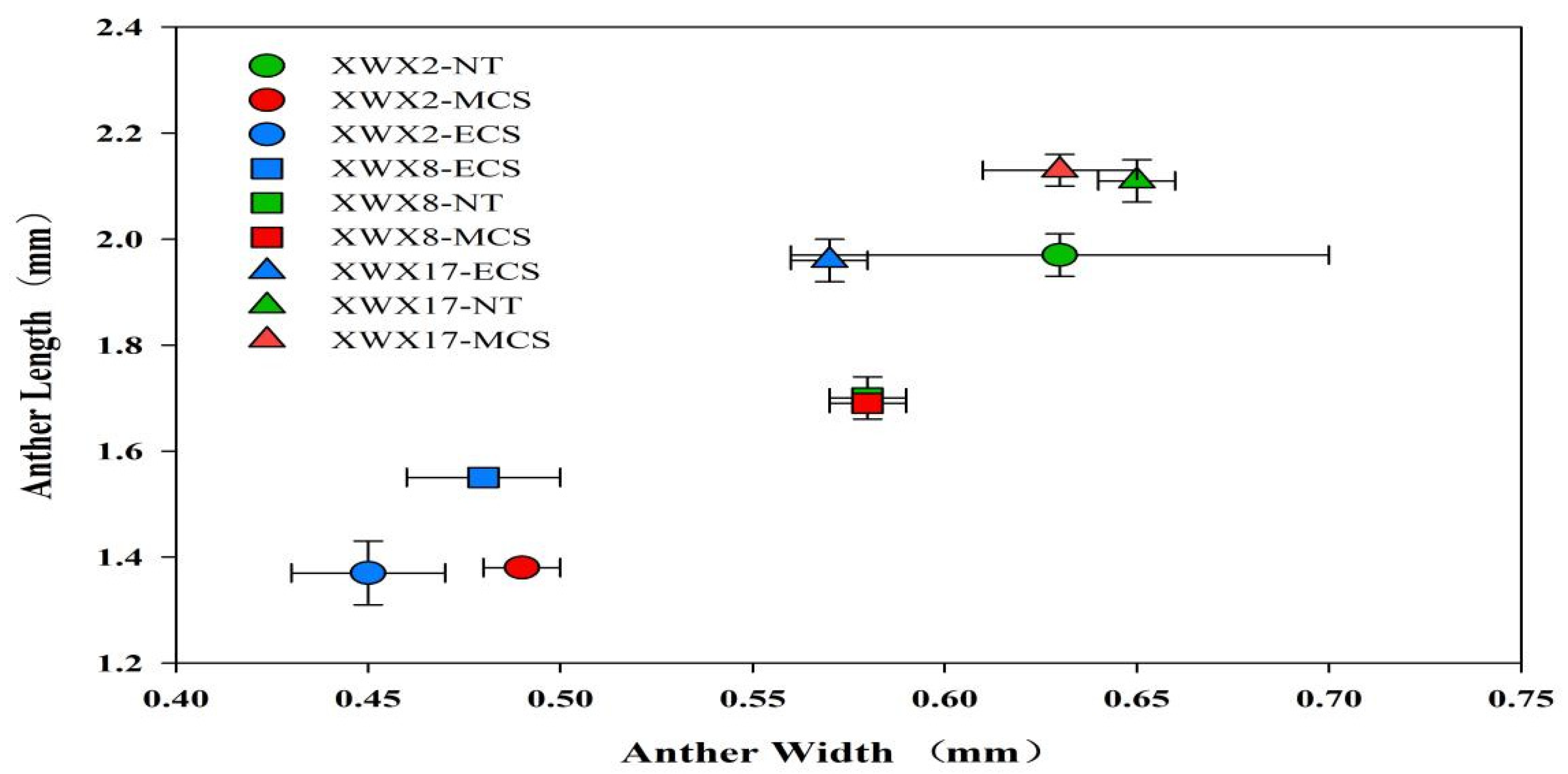
Figure 4.
The anther length and anther width of three inbred late varieties under different temperature treatments. XWX2: Xiangwanxian2; XWX8: Xiangwanxian8; XWX17: Xiangwanxian17; NT: Normal temperature control; MCS: Moderate chilling stress; ECS: Extreme chilling stress.
Similar trends were also observed in anther width and length. Chilling stress decreased the length and width of the anthers, and such effects was more pronounce in ECS than that MCS (Figure 4).
3.5. SPAD of Different Late Rice
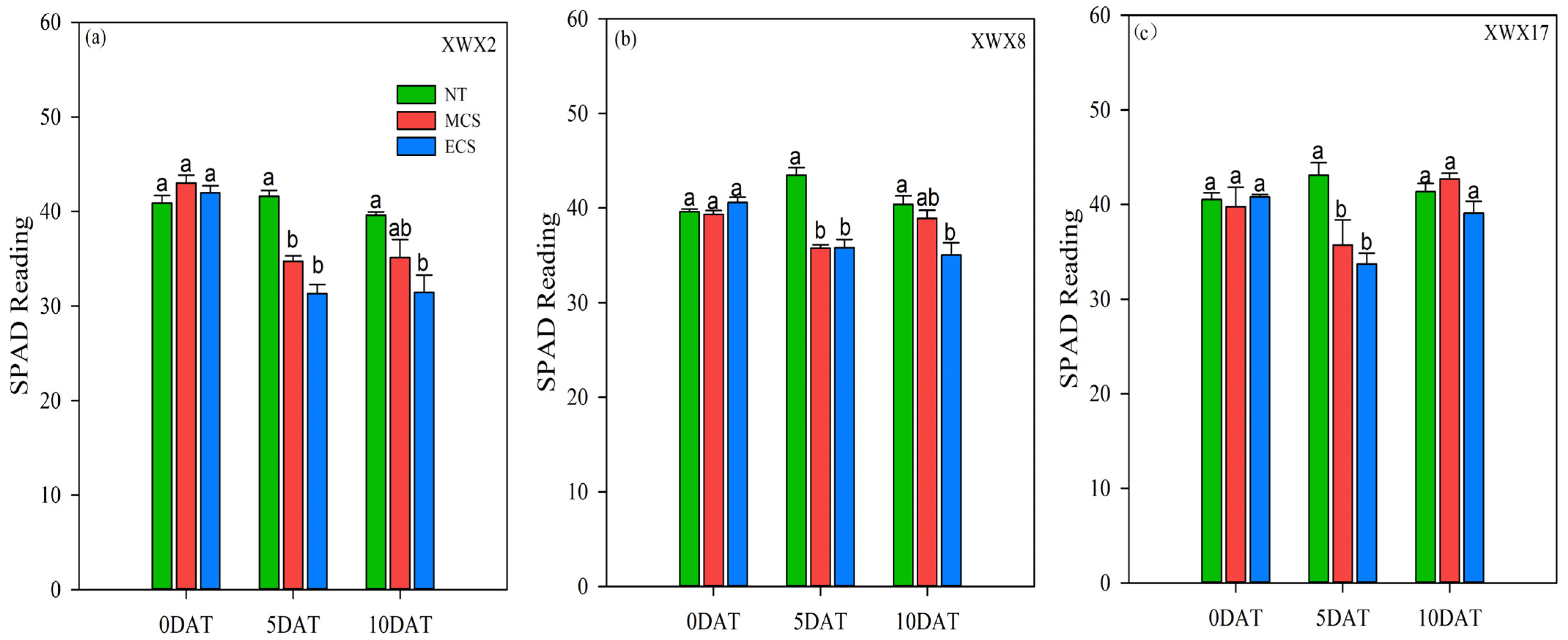
The SPAD of the three late rice varieties was descended after 5 days of chilling stress treatments (Figure 5). When compared with NT, the SPAD of XWX2, XWX8 and XWX17 was decreased by 19.26%, 9.15% and 10.21%, respectively, at MCS, and was decreased by 25.37%, 11.74% and 19.36%, respectively, at ECS. The SPAD of the three late rice varieties was recovered after 10 days of chilling stress treatments (Figure 4). When compared with XWX2, the SPAD of XWX8 and XWX17 was increased by 10.83%, 21.65%, respectively, at MCS, and was increased by 11.52%, 24.38%, respectively, at ECS. In comparison with XWX2 and XWX8, the XWX17 showed stronger resistance to MCS/ECS because the increase in SPAD did reach a significant level after 10 days of chilling stress treatments.
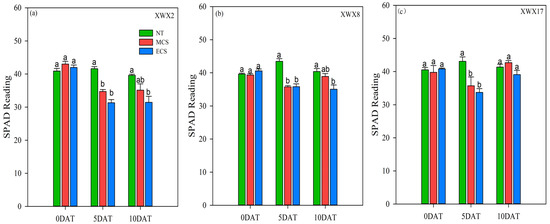
Figure 5.
The flag leaf SPAD reading of three inbred late varieties under different temperature treatments. (a) XWX2. (b) XWX8. (c) XWX17. DAT: Days after temperature treatments; XWX2: Xiangwanxian2; XWX8: Xiangwanxian8; XWX17: Xiangwanxian17; NT: Normal temperature control; MCS: Moderate chilling stress; ECS: Extreme chilling stress. Different lowercase letters denote statistical differences among treatments of a cultivar at the 5% level according to LSD test. Error bars above mean indicate standard error (n = 3).
3.6. Chlorophyll of Different Late Rices
The chlorophyll (chlorophyll a, chlorophyll b) of the three late rice varieties was descended after 5 days of chilling stress treatments (Table 3). When compared with NT, the chlorophyll a of XWX2, XWX8 and XWX17 was decreased by 14.70%, 10.51% and 7.57%, respectively, at MCS, and was decreased by 23.10%, 17.61% and 16.45%, respectively, at ECS. When compared with NT, the chlorophyll b of XWX2, XWX8 and XWX17 was decreased by 6.33%, 5.00% and 5.00%, respectively, at MCS, and was decreased by 8.86%, 8.75% and 6.25%, respectively, at ECS.
Table 3.
The leaf chlorophyll content of three inbred late varieties under different temperature treatments.
When compared with XWX2, the chlorophyll (chlorophyll a + b) of XWX17 was increased by 7.77% at MCS, and was increased by 8.22% at ECS. When compared with XWX8, the chlorophyll (chlorophyll a + b) of XWX17 was increased by 9.97% at MCS and was increased by 8.52% at ECS. In comparison with XWX2 and XWX8, XWX17 showed stronger resistance to MCS because the decrease in chlorophyll (chlorophyll a + b) did not reach a significant level. Compared with NT, although the chlorophyll (chlorophyll a + b) of the three varieties was significantly reduced at ECS, the decreased level was lower in XWX17 than that in XWX2 and XWX8.
4. Discussion
4.1. Chilling Stress at Heading Decreased the Grain Filling Percentage via Reducing Spikelet Fertility
Chilling is one of the major abiotic stresses limiting the growth and productivity of many field crops. Rice is cultivated in tropical and subtropical environments, and is extremely sensitive to chilling stress, particularly at flowering [15]. The results of the present study were consistent with previous research [16,17,18,19,20] showing that the exposure of rice plants to either moderate (20 °C) or extreme(17 °C) low temperature for 5 days during heading would irreversibly reduce grain filling percentage and grain yield. Moreover, such negative effects induced by chilling stress might mainly be attributed to the decrease in spikelet fertility, because a significantly positive correlation between grain filling percentage and spikelet fertility was observed (p < 0.01). Meanwhile, the grain weight showed no significant variance among NT, MCS and ECS, and the difference between spikelet fertility and grain filling percentage of the same variety did not vary across temperature treatments, suggesting that the chilling stress did not affect the grain filling process in present study. This result was in contrary with the research of Huang, M. et al. [18] which indicated that temperature stress before anthesis also had a significant impact on the final the grain weight. However, several studies denoted that the chilling stress at early stage of booting or flowering may not affect the grain filling process and grain weight [21,22]. The differences between studies might be attributed to the differences in chilling duration, chilling level and other factors including relative air humidity and crop managements, and the effects of chilling stress at heading on the grain filling attributes in late-season rice need to be explored in future studies.
4.2. The Variances in Chilling Tolerance among Three Inbred Late Varieties
The development of breeding varieties with strong tolerance is one of the important strategies to diminish the negative impact of chilling stress during heading on the spikelet fertility and yield formation of late-season rice. In recent years, the resistance of hybrid rice to abiotic stresses including drought [23,24,25], submergence [26,27,28], high temperature [29,30,31] and chilling has been intensively studied in China, and several hybrid rice varieties that possessed relative high chilling tolerance during flowering have been successfully bred and adopted in production. In contrary, little attention has been paid to the evaluation of stress tolerance in inbred lines. The results of the present study indicated that the genetic improvement has been achieved in inbred late rice varieties, as the grain filling percentage and the spikelet fertility of XWX17 (released in 2008) was significantly higher than that of XWX2 (released in 1988) and XWX8 (released in 1988) when suffering from moderate chilling stress during heading. In addition, although the grain filling percentage of all the three late rice varieties was significantly decreased under ECS, the decreased percentage was lower in XWX17 than in XWX2 and XWX8. Nevertheless, the present study evaluated the chilling tolerance of the three varieties without a chilling-susceptible or -tolerant control, which limited the comparability of the results. Therefore, further studies to evaluate the chilling tolerance of inbred late rice varieties in comparison with elite control varieties under both greenhouse and field conditions are desperately needed.
4.3. Morphological and Physiological Characteristics Underlying the Varietal Differences in Chilling Tolerance
The viability of the rice pollen grain is frequently affected by chilling stress [32,33,34]. It has been reported that the chilling stress during panicle initiation would significantly reduce pollen development, resulting in decreased pollen viability [35], while at the booting and heading stage, although the pollen development and filling is finished, the chilling-induced pollen sterility has been widely observed [36]. In the present study, the varietal differences in spikelet fertility in response to chilling stress can be partially explained by the variances in pollen viability. The pollen viability of XWX2 and XWX8 was reduced by both MCS and ECS, while the pollen viability of XWX17 was not reduced under MCS, and the deceased level under ECS was smaller than the other varieties, which suggested that the pollen grains of the newly developed variety, might possessed stronger chilling tolerance than the older ones. In addition to pollen viability, the anther development and characteristics also greatly contribute to the spikelet fertility. Anther dehiscence is essential for pollen spreading and pollination, which are tightly associated with spikelet fertility [9,15,36,37,38]. The ability of anther dehiscence is generally related with the anther development. The abnormal anther shape would greatly decrease anther dehiscence rate and length, resulting in decreased pollen germination and shedding, and thus resulting in decreased spikelet fertility [39]. The present study found that under MCS, the anther dehiscence length was significantly reduced in XWX2 and XWX8, but was not reduced in XWX17. Moreover, the anther volume of the XWX2 and XWX8 was reduced under ECS or MCS via decreased anther length and width; in contrast, the anther volume of XWX17 was not influenced by chilling stress even under ECS. In summary, the above results indicated that the better anther attributes in XWX17 regarding pollen viability, anther volume and anther dehiscence might contribute to the higher spikelet fertility under chilling stress, as compared with that of XWX2 and XWX8.
Several metabolic events including production and scavenging of reactive oxygen species (ROS), photosynthetic ability and leaf senescence are highly associated with plant stress tolerance and may affect the spikelet fertility of rice under chilling stress [40,41]. In the present study, the SPAD reading of the XWX17 was less influenced by chilling stress as compared with XWX2 and XWX8, suggesting that the leaf function in XWX17 was not severely reduced under chilling stress; such traits, which might be attributed to better ROS scavenging ability that reduced the degradation of leaf chlorophyll, greatly guaranteed the substrate transportation and photosynthesis of rice under chilling stress [42,43]. In addition, our study suggested that the chlorophyll a content and the SPAD reading of rice flag leaf can be used as indicators to predict spikelet fertility of late rice under chilling stress. In addition, several traits including the morphology of pollen [35], spikelet transpiration rate and internal spikelet temperature [44] were also reported to contribute to the variances in pollen viability and spikelet fertility, which might be manipulated by the expression levels of chilling-related genes and transcription factors such as Lsi1 [45], OsSAPK6 [46], and OsWRKY115 [47]. Nevertheless, the physiological and molecular mechanisms underlying the chilling responses of inbred late varieties were not fully examined and need to be addressed in future studies.
5. Conclusions
In summary, the newly released inbred late rice variety XWX17 demonstrated better chilling tolerance during heading than the early released varieties, with respect to higher grain filling percentage. The improved grain filling percentage in XWX17 might be the results of increased spikelet fertility, which was attributed to the increase in pollen viability, anther dehiscence length and anther volume. In addition, the SPAD value and the chlorophyll a content of the flag leaf can be used as indicators to predict the tolerance of rice spikelet fertility when suffering from chilling stress during heading. The present study provides evidence that the chilling tolerance of inbred lines at the heading stage was improved, However, the physiological and molecular mechanisms underlying the chilling responses of inbred late varieties were not fully examined and need to be addressed in future studies.
Author Contributions
Conceptualization, Q.T. and H.X.; data curation, M.R., W.W. and S.P.; formal analysis, M.R. and S.P.; funding acquisition, Q.T. and H.X.; investigation, S.P., H.X., W.S. and T.H.; methodology, M.R. and W.W.; project administration, Q.T. and H.X.; writing—original draft, M.R. and W.W.; writing—review and editing, Q.T. and H.X. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2017YFD0300409); the Project for China Agriculture Research System (CARS-01-27); the Key Projects of Regional Innovation and Development Joint Fund of China (U21A20184); the Natural Science Foundation of Hunan Province, China (2021JJ40248).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shinada, H.; Iwata, N.; Sato, T.; Fujino, K. Genetical and morphological characterization of cold tolerance at fertilization stage in rice. Breed. Sci. 2013, 63, 197–204. [Google Scholar] [CrossRef]
- Jia, Y.; Zou, D.; Wang, J.; Liu, H.; Inayat, M.A.; Sha, H.; Zheng, H.; Sun, J.; Zhao, H. Effect of low water temperature at reproductive stage on yield and glutamate metabolism of rice (Oryza sativa L.) in China. Field Crop. Res. 2015, 175, 16–25. [Google Scholar] [CrossRef]
- Matsui, T.; Omasa, K.; Horie, T. The difference in sterility due to high temperatures during the flowering period among japonica-rice varieties. Plant Prod Sci. 2001, 4, 90–93. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Boote, K.J.; Allen, L.H., Jr.; Sheehy, J.E.; Thomas, J.M.G. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crop. Res. 2006, 95, 98–411. [Google Scholar] [CrossRef]
- Rang, Z.W.; Jagadish, S.V.K.; Zhou, Q.M.; Craufurd, P.Q.; Heuer, S. Effect of high temperature and water stress on pollen germination and spikelet fertility in rice. Environ. Exp. Bot. 2011, 70, 58–65. [Google Scholar] [CrossRef]
- Song, Y.; Jiang, M.; Zhang, H.; Li, R. Zinc Oxide Nanoparticles Alleviate Chilling Stress in Rice (Oryza sativa L.) by Regulating Antioxidative System and Chilling Response Transcription Factors. Molecules 2021, 26, 2196. [Google Scholar] [CrossRef] [PubMed]
- Kudoh, H.; Sonoike, K. Irreversible damage to photosystemi by chilling in the light: Cause of the degradation of chlorophyll after returning to normal growth temperature. Planta 2002, 215, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; Chen, Y.; et al. Effect of Low Temperature on Chlorophyll Biosynthesis and Chloroplast Biogenesis of Rice Seedlings during Greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhang, Y.; Xiang, J.; Uphoff, N.T.; Pan, X.; Zhu, D. Effects of Low Temperature Stress on Spikelet-Related Parameters during Anthesis in Indica–Japonica Hybrid Rice. Front. Plant Sci. 2017, 8, 1350. [Google Scholar] [CrossRef] [PubMed]
- Farrell, T.C.; Fox, K.M.; Williams, R.L.; Fukai, S. Genotypic variation for cold tolerance during reproductive development in rice: Screening with cold air and cold water. Field Crop. Res. 2006, 98, 178–194. [Google Scholar] [CrossRef]
- Badawy, S.A.; Zayed, B.A.; Bassiouni, S.M.A.; Mahdi, A.H.A.; Majrashi, A.; Ali, E.F.; Seleiman, M.F. Influence of Nano Silicon and Nano Selenium on Root Characters, Growth, Ion Selectivity, Yield, and Yield Components of Rice (Oryza sativa L.) under Salinity Conditions. Plants 2021, 10, 1657. [Google Scholar] [CrossRef] [PubMed]
- Sim, J.; Kesawat, M.S.; Kumar, M.; Kim, S.; Mani, V.; Subramanian, P.; Hahn, B. Lack of the α1,3-fucosyltransferase gene (osfuct) affects anther development and pollen viability in rice. Int. J. Mol. Sci. 2018, 19, 1225. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, X.; Jiang, L.; Li, X.; Yang, S.; Li, Y. Hyperspectral Reflectance Characteristics of Rice Canopies under Changes in Diffuse Radiation Fraction. Remote Sens. 2022, 14, 285. [Google Scholar] [CrossRef]
- Wintermans, J.; De Mots, A. Spectrophotometric characteristics of chlorophylls a and b and their phenophytins in ethanol. Biochim. Biophys. Acta (BBA)-Biophys. Incl. Photosynth 1965, 109, 448–453. [Google Scholar] [CrossRef]
- Shi, Y.; Guo, E.; Wang, L.; Li, T.; Jiang, S.; Xiang, H.; Yang, X. Effects of chilling at the booting and flowering stages on rice phenology and yield: A case study in Northeast China. J. Agron. Crop Sci. 2022, 208, 197–208. [Google Scholar] [CrossRef]
- Ali, I.; Tang, L.; Dai, J.; Kang, M.; Mahmood, A.; Wang, W.; Zhu, Y. Responses of Grain Yield and Yield Related Parameters to Post-Heading Low-Temperature Stress in Japonica Rice. Plants 2021, 10, 1425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Peng, D.; Zhang, H.; Meng, C.; Zhang, X.; Hou, J.; Wei, C. Low soil temperature reducing the yield of drip irrigated rice in arid area by influencing anther development and pollination. J. Arid. Land 2019, 11, 419–430. [Google Scholar] [CrossRef]
- Huang, M.; Fang, S.; Shuanglü, S.; Zou, Y. Delayed transplanting reduced grain yield due to low temperature stress at anthesis in machine-transplanted late-season rice. Exp. Agric. 2019, 55, 843–848. [Google Scholar] [CrossRef]
- Siddik, M.A.; Zhang, J.; Chen, J.; Qian, H.; Jiang, Y.; Raheem, A.K.; Zhang, W. Responses of indica rice yield and quality to extreme high and low temperatures during the reproductive period. Eur. J. Agron. 2019, 106, 30–38. [Google Scholar] [CrossRef]
- .Pengyuan, X.; Dongxue, L.; Yuguang, L.; Xiaoli, Q. Research status and countermeasures of rice barrier chilling injury in cold region of china. IOP Conference Series. Earth Environ. Sci. 2018, 199, 022027. [Google Scholar] [CrossRef]
- Cai, C.; Yin, X.; He, S.; Jiang, W.; Si, C.; Struik, P.C.; Luo, W.; Li, G.; Xie, Y.; Xiong, Y.; et al. Responses of wheat and rice to factorial combinations of ambient and elevated CO2 and temperature in FACE experiments. Glob. Chang. Biol. 2016, 22, 856–874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hou, J.; Zhang, H.; Meng, C.; Zhang, X.; Wei, C. Low soil temperature inhibits yield of rice under drip irrigation. J. Soil Sci. Plant Nutr. 2019, 19, 228–236. [Google Scholar] [CrossRef]
- Wang, W.; Quan, C.; Zheng, S.; Yu, W.W.; Mo, Y.; Ma, C.; Chen, R. OsPM1 is a positive regulator of rice tolerance to drought stress but a negative regulator of rice tolerance to salt stress. J. Plant Interact. 2021, 16, 213–221. [Google Scholar] [CrossRef]
- Menghao, Z.; He, Y.; Mingqiang, Z.; Ayaz, A.; Xu, S.; Zijun, H.; Zhang, Z. ipa1 improves rice drought tolerance at seedling stage mainly through activating abscisic acid pathway. Plant Cell Rep. 2022, 41, 221–232. [Google Scholar] [CrossRef]
- Han, B.; Wang, J.; Li, Y.; Ma, X.; Jo, S.; Cui, D.; Han, L. Identification of quantitative trait loci associated with drought tolerance traits in rice (Oryza sativa L.) under PEG and field drought stress. Euphytica 2018, 214, 74. [Google Scholar] [CrossRef]
- Yang, S.; Wu, Y.; Chen, C.; Lai, M.; Yen, H.; Yang, C. Physiological and molecular responses of seedlings of an upland rice (‘Tung Lu 3’) to total submergence compared to those of a submergence-tolerant lowland rice (‘FR13A’). Rice 2017, 10, 42. [Google Scholar] [CrossRef]
- Tran, D.X.; Do, T.K. Effects of Exogenous Application of Protocatechuic Acid and Vanillic Acid to Chlorophylls, Phenolics and Antioxidant Enzymes of Rice (Oryza sativa L.) in Submergence. Molecules 2018, 23, 620. [Google Scholar] [CrossRef]
- Yong-Pei, W.; Wang, S.; Yu-Chi, C.; Ho, C.; Yu-Chia, H. Submergence Gene Sub1A Transfer into Drought-Tolerant japonica Rice DT3 Using Marker-Assisted Selection. Int. J. Mol. Sci. 2021, 22, 13365. [Google Scholar] [CrossRef]
- Guonan, F.; Shenglong, Y.; Banpu, R.; Chaolei, L.; Anpeng, Z.; Hongzhen, J.; Qian, Q. Isolation of TSCD11 Gene for Early Chloroplast Development under High Temperature in Rice. Rice 2020, 13, 49. [Google Scholar] [CrossRef]
- Luo, R.; Jiang, H.; Lv, Y.; Hu, S.; Sheng, Z.; Shao, G.; Wei, X. Chlorophyll deficient 3, Encoding a Putative Potassium Efflux Antiporter, Affects Chloroplast Development Under High Temperature Conditions in Rice (Oryza sativa L.). Plant Mol. Biol. Report. 2018, 36, 675–684. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, L.; Shujun, O.; Ruci, W.; Wang, Y.; Chengcai, C.; Shanguo, Y. Natural variations of SLG1 confer high-temperature tolerance in indica rice. Nat. Commun. 2020, 11, 5441. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, J.; Fang, J.; Guo, Z.; Lu, S. Down-regulation of S-adenosylmethionine decarboxylase genes results in reduced plant length, pollen viability, and abiotic stress tolerance. Plant Cell Tissue Organ Cult. 2014, 116, 311–322. [Google Scholar] [CrossRef]
- Liu, C.; Schläppi, M.R.; Mao, B.; Wang, W.; Wang, A.; Chu, C. The bZIP 73 transcription factor controls rice cold tolerance at the reproductive stage. Plant Biotechnol. J. 2019, 17, 1834–1849. [Google Scholar] [CrossRef]
- Jeong, B.Y.; Lee, Y.; Kwon, Y.; Kim, J.H.; Ham, T.-H.; Kwon, S.-W.; Lee, J. Genome-wide association study reveals the genetic basis of chilling tolerance in rice at the reproductive stage. Plants 2021, 10, 1722. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Wang, W.; Lu, Q.; Huang, J.; Peng, S.; Cui, K. Abnormal anther development leads to lower spikelet fertility in rice (Oryza sativa L.) under high temperature during the panicle initiation stage. BMC Plant Biol. 2021, 21, 1–17. [Google Scholar] [CrossRef]
- Koumoto, T.; Saito, N.; Aoki, N.; Iwasaki, T.; Kawai, S.; Yokoi, S.; Shimono, H. Effects of salt and low light intensity during the vegetative stage on susceptibility of rice to male sterility induced by chilling stress during the reproductive stage. Plant Prod. Sci. 2016, 19, 497–507. [Google Scholar] [CrossRef]
- Pereira, D.C.R.; Sperotto, R.A.; Cargnelutti, D.; Adamski, J.M.; de FreitasTerra, T.; Janette, P.F. Avoiding damage and achieving cold tolerance in rice plants. Food Energy Secur. 2013, 2, 96–119. [Google Scholar] [CrossRef]
- Dingkuhn, M.; Sow, A.; Ramantsoanirina, A.; Alpha, B.B.; Manneh, B.; Courtois, B.; .Radanielina, T. Field phenomics for response of a rice diversity panel to ten environments in senegal and madagascar. 2. chilling-induced spikelet sterility. Field Crops Res. 2015, 183, 282–293. [Google Scholar] [CrossRef]
- Xu, D.; Qu, S.; Tucker, M.R.; Zhang, D.; Liang, W.; Shi, J. Ostkpr1 functions in anther cuticle development and pollen wall formation in rice. BMC Plant Biol. 2019, 19, 104. [Google Scholar] [CrossRef]
- Sato, Y.; Masuta, Y.; Saito, K.; Murayama, S.; Ozawa, K. Enhanced chilling tolerance at the booting stage in rice by transgenic overexpression of the ascorbate peroxidase gene, OsAPXa. Plant Cell Rep. 2011, 30, 399–406. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, M.; Xu, W.; Wang, Y.; Huang, K.; Zhang, C.; Wen, J. Understanding the molecular mechanism of anther development under abiotic stresses. Plant Mol. Biol. 2021, 105, 1–10. [Google Scholar] [CrossRef]
- Gan, P.; Liu, F.; Li, R.; Wang, S.; Luo, J. Chloroplasts—Beyond energy capture and carbon fixation: Tuning of photosynthesis in response to chilling stress. Int. J. Mol. Sci. 2019, 20, 5046. [Google Scholar] [CrossRef]
- Sritama, M.; Abhishek, M.; Das, P.; Subhendu, B.; Debapriya, C.; Jolly, C.; Majumder, A.L. A salt-tolerant chloroplastic FBPase from Oryza coarctata confers improved photosynthesis with higher yield and multi-stress tolerance to indica rice. Plant Cell Tissue Organ Cult. 2021, 145, 561–578. [Google Scholar] [CrossRef]
- Wang, W.; Cui, K.; Hu, Q.; Wu, C.; Li, G.; Huang, J.; Peng, S. Response of spikelet water status to high temperature and its relationship with heat tolerance in rice. Crop J. 2020, 9, 1344–1356. [Google Scholar] [CrossRef]
- Li, Z.; Umar Khan, M.; Yan, X.; Mu, D.; Xie, Y.; Waqas, M.; Wu, X.; Letuma, P.; Fang, C.; Lin, W. Deciphering the Molecular Mechanisms of Chilling Tolerance in Lsi1-Overexpressing Rice. Int. J. Mol. Sci. 2022, 23, 4667. [Google Scholar] [CrossRef]
- Jia, M.; Meng, X.; Song, X.; Zhang, D.; Kou, L.; Zhang, J.; Jing, Y.; Liu, G.; Liu, H.; Huang, X.; et al. Chilling-induced phosphorylation of IPA1 by OsSAPK6 activates chilling tolerance responses in rice. Cell Discov. 2022, 8, 71. [Google Scholar] [CrossRef]
- Liu, H.; Yang, L.; Xu, S.; Lyu, M.-J.; Wang, J.; Wang, H.; Zheng, H.; Xin, W.; Liu, J.; Zou, D. OsWRKY115 on qCT7 links to cold tolerance in rice. Theor. Appl. Genet. 2022, 135, 2353–2367. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).