
Identification of New Sources for Earliness and Low Grain Moisture at Harvest through Maize Landraces’ Test-Cross Performance

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
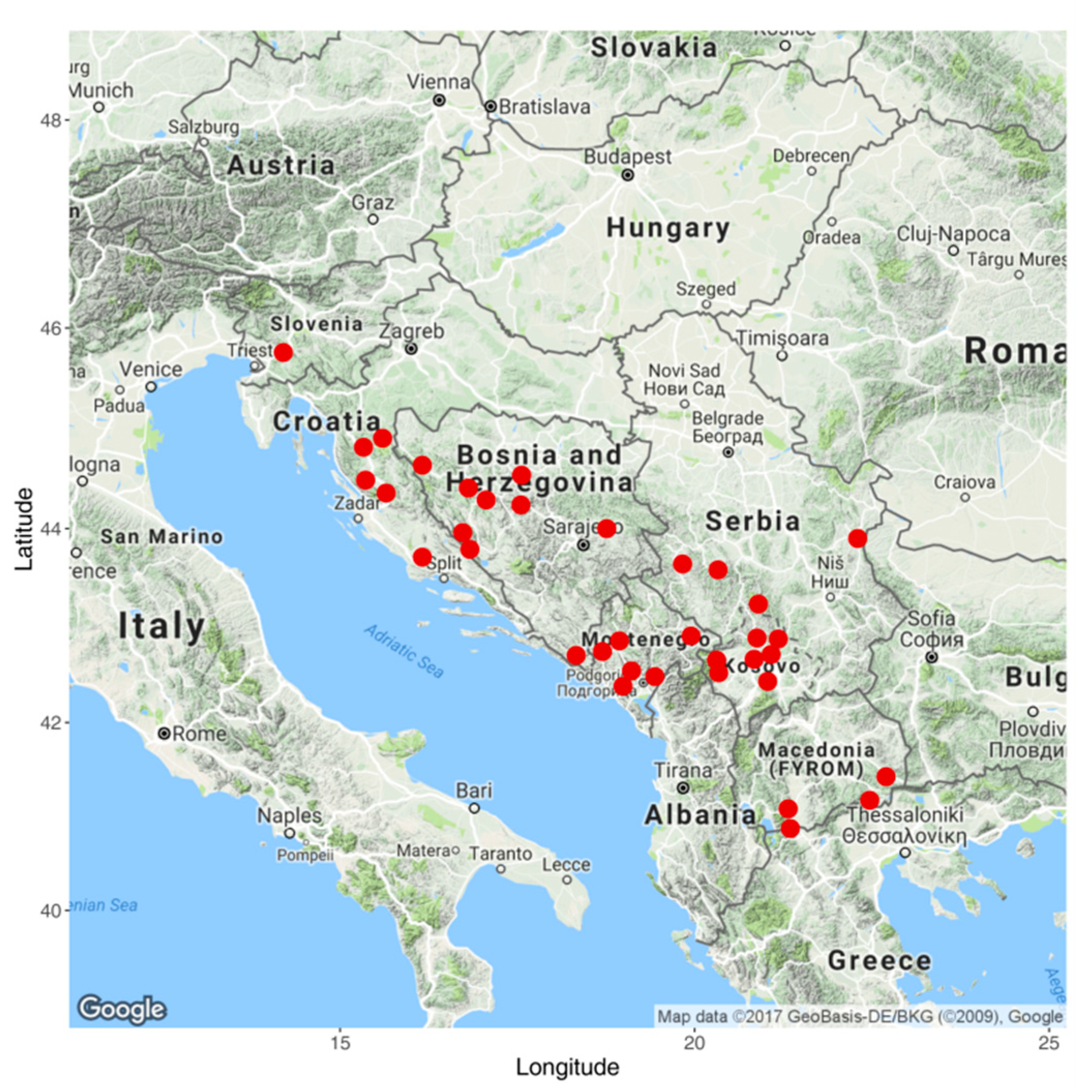
2.1. Plant Material
2.2. Field Experiment and Statistical Analysis
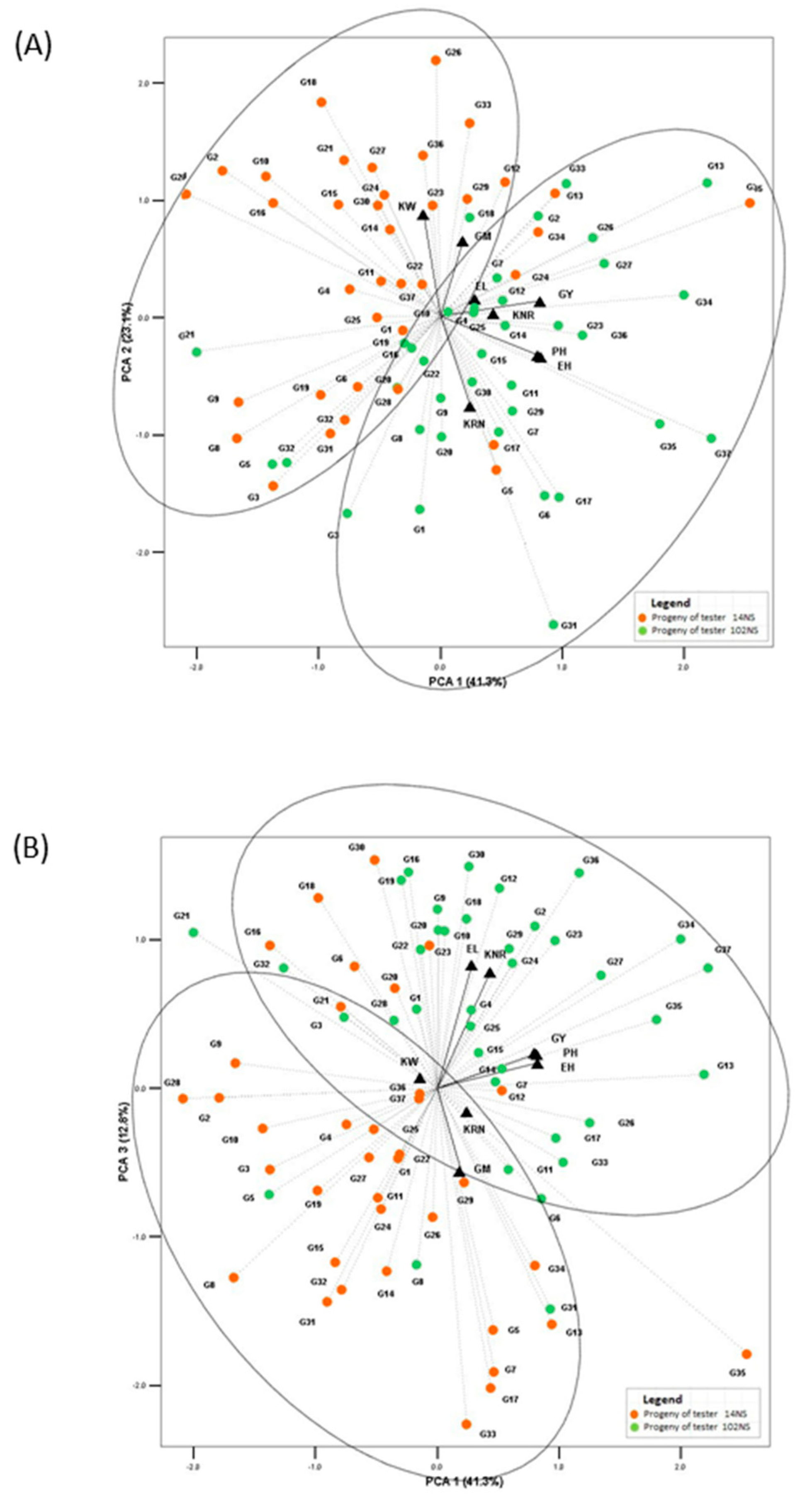
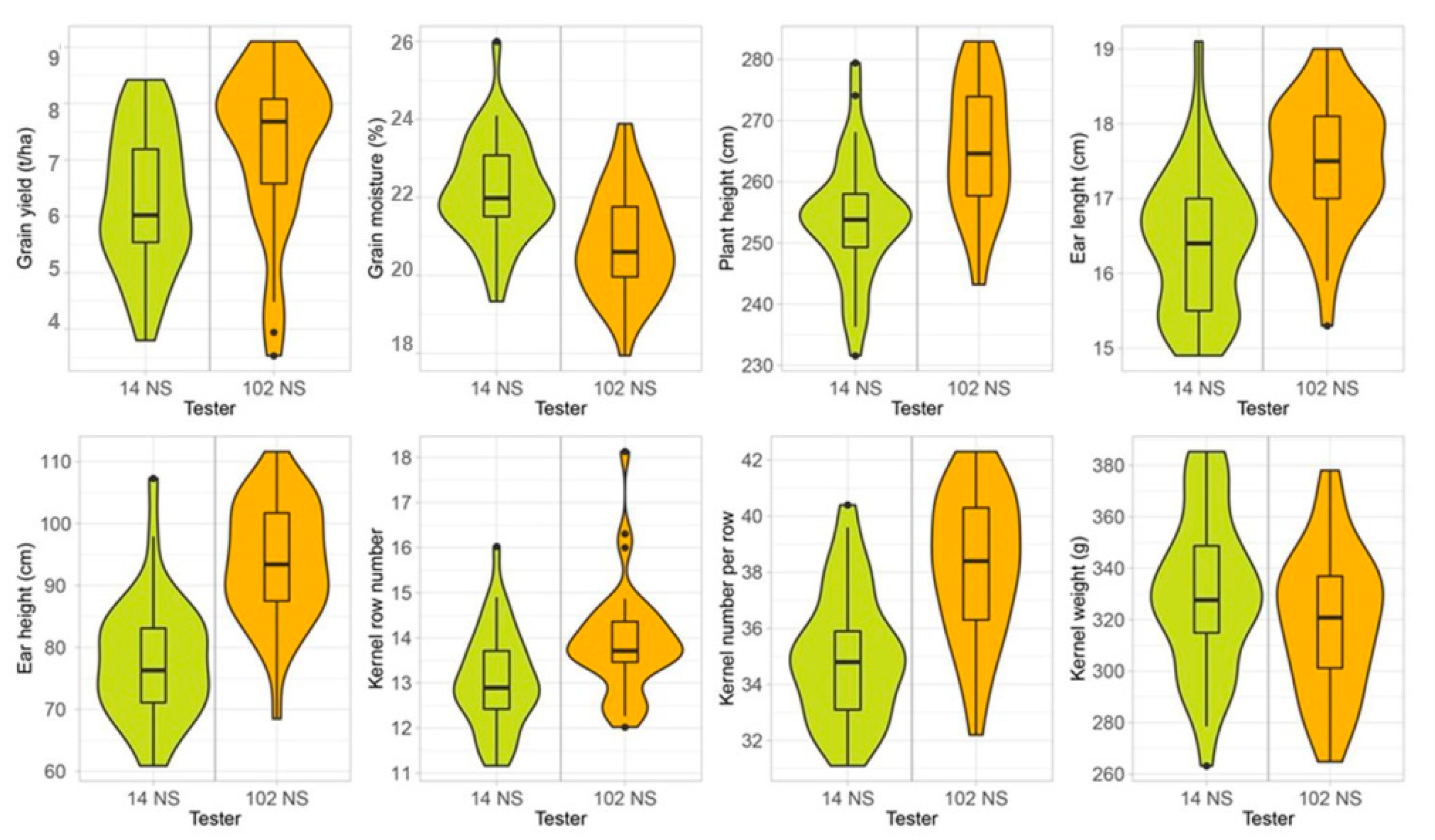
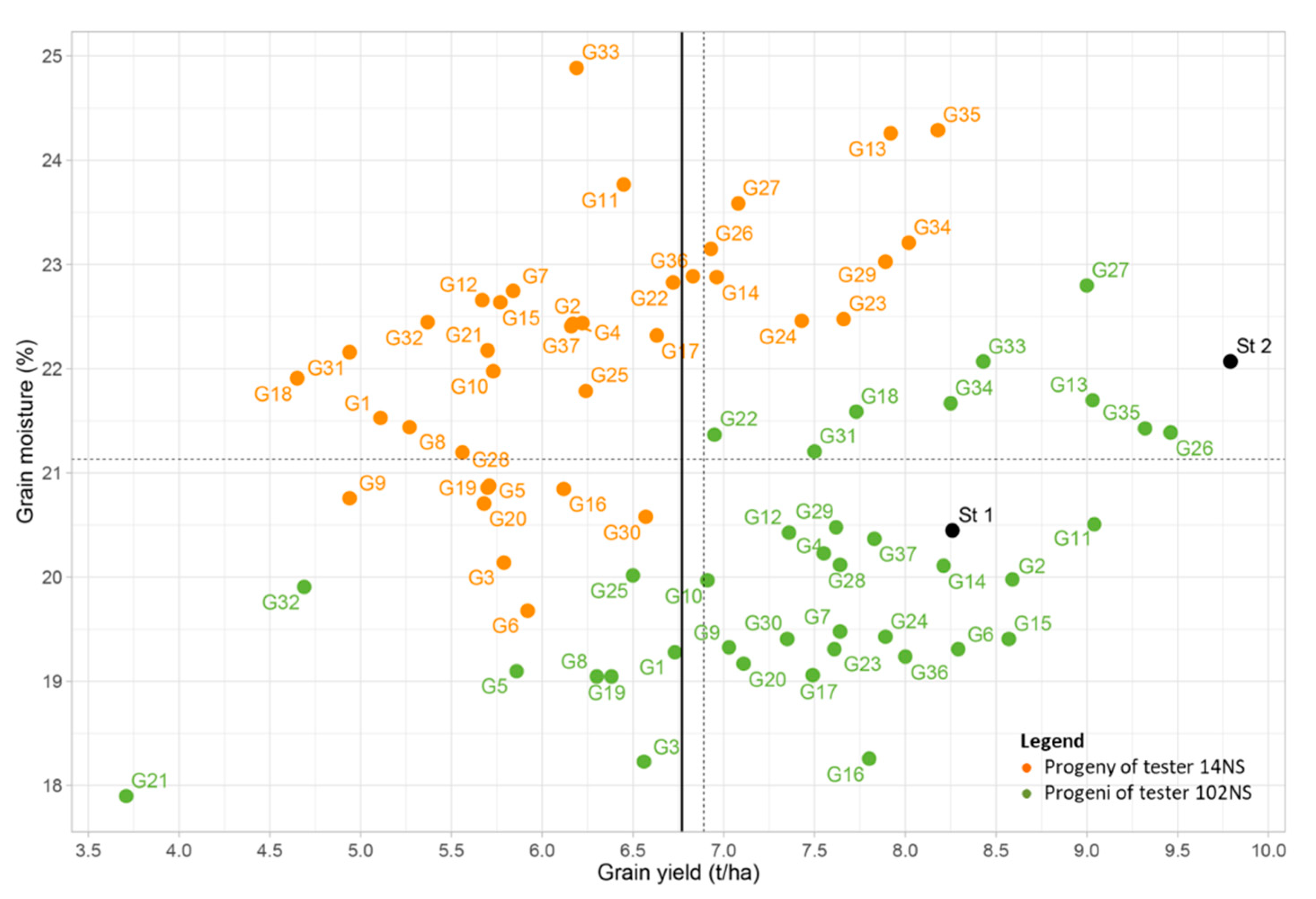
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiferaw, B.; Prasanna, B.M.; Hellin, J.; Bänziger, M. Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Secur. 2011, 3, 307–327. [Google Scholar] [CrossRef]
- Babić, V.; Pavlov, M.; Boćanski, J. Status and perspective of plant breeding and seed industry in Serbia. Sel. I Semen. Plant Breed. Seed Prod. 2016, 22, 19–27. [Google Scholar]
- Masuka, B.; Magorokosho, C.; Olsen, M.; Atlin, G.N.; Bänziger, M.; Pixley, K.V.; Vivek, B.S.; Labuschagne, M.; Matemba-Mutasa, R.; Burgueño, J.; et al. Gains in Maize Genetic Improvement in Eastern and Southern Africa: II. CIMMYT Open-Pollinated Variety Breeding Pipeline. Crop Sci. 2017, 57, 180–191. [Google Scholar] [CrossRef]
- Popović, A.; Kravić, N.; Prodanović, S.; Filipović, M.; Sečanski, M.; Babić, V.; Miriţescu, M. Characterisation and evaluation towards selection of maize landraces with the best per se performances. Rom. Agric. Res. 2020, 37, 49–58. Available online: https://www.incda-fundulea.ro/rar/nr37/rar37.7.pdf (accessed on 10 February 2021).
- Babić, V.; Andjelkovic, V.; Jovovic, Z.; Babic, M.; Vasic, V.; Kravic, N. Diversity Assessment of the Montenegrin Maize Landrace Gene Pool Maintained in Two Gene Banks. Plants 2021, 10, 1503. [Google Scholar] [CrossRef]
- Li, W.; Yu, Y.; Wang, L.; Luo, Y.; Peng, Y.; Xu, Y.; Liu, X.; Wu, S.; Jian, L.; Hu, J.; et al. The genetic architecture of the dynamic changes in grain moisture in maize. Plant Biotechnol. J. 2021, 19, 1195–1205. [Google Scholar] [CrossRef]
- De-Jager, B.; Roux, C.Z.; Kühn, H.C. An evaluation of two collections of south african maize (Zea mays L.) germplasm: 2. the genetic basis of dry-down rate. S. Afr. J. Plant Soil 2004, 21, 120–122. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Xu, C.; Tan, W.; Wang, P.; Meng, Q. Decreased kernel moisture in medium-maturing maize hybrids with high yield for mechanized grain harvest. Crop Sci. 2019, 59, 2794–2805. [Google Scholar] [CrossRef]
- Martinez-Feria, R.A.; Licht, M.A.; Ordóñez, R.A.; Hatfield, J.L.; Coulter, J.A.; Archontoulis, S.V. Evaluating maize and soybean grain dry-down in the field with predictive algorithms and genotype-by-environment analysis. Sci. Rep. 2019, 9, 7167. [Google Scholar] [CrossRef]
- Raymond, D.J.; Sessions, S.L.; Sobel, A.H.; Fuchs, Ž. The mechanics of gross moist stability. J. Adv. Model. Earth Syst. 2009, 1, 20. [Google Scholar] [CrossRef]
- Assefa, Y.; Carter, P.; Hinds, M.; Bhalla, G.; Schon, R.; Jeschke, M.; Paszkiewicz, S.; Smith, S.; Ciampitti, I.A. Analysis of long term study indicates both agronomic optimal plant density and increase maize yield per plant contributed to yield gain. Sci. Rep. 2018, 8, 4937. [Google Scholar] [CrossRef]
- Burton, A.B.; Kemanian, A.R. Maize yield in response to alternating low- and high-density rows of diverse hybrids. Eur. J. Agron. 2022, 135, 126472. [Google Scholar] [CrossRef]
- Lauer, S.; Hall, B.D.; Mulaosmanovic, E.; Anderson, S.R.; Nelson, B.; Smith, S. Morphological changes in parental lines of pioneer brand maize hybrids in the U.S. Corn Belt. Crop Sci. 2012, 52, 1033–1043. [Google Scholar] [CrossRef]
- Lindsey, A.J.; Thomison, P.R. Drought-tolerant corn hybrid and relative maturity yield response to plant population and planting date. Agron. J. 2016, 108, 229–242. [Google Scholar] [CrossRef]
- Brauner, P.C.; Schipprack, W.; Utz, H.F.; Bauer, E.; Mayer, M.; Schön, C.C.; Melchinger, A.E. Testcross performance of doubled haploid lines from European flint maize landraces is promising for broadening the genetic base of elite germplasm. Theor. Appl. Genet. 2019, 132, 1897–1908. [Google Scholar] [CrossRef]
- Vancetovic, J.; Bozinovic, S.; Ignjatovic-Micic, D.; Delic, N.; Kravic, N.; Nikolic, A. A diallel cross among drought tolerant maize populations. Euphytica 2015, 205, 1–16. [Google Scholar] [CrossRef]
- Van Hintum, T.J.L. Duplication within and between germplasm collections. III. A quantitative model. Genet. Resour. Crop Evol. 2000, 47, 507–513. [Google Scholar] [CrossRef]
- Babić, V.; Vančetović, J.; Prodanović, S.; Andjelković, V.; Babić, M.; Kravić, N. The identification of drought tolerant maize accessions by the two-step cluster analysis. Rom. Agric. Res. 2012, 29, 53–61. Available online: https://www.incda-fundulea.ro/rar/nr29/rar29.8.pdf (accessed on 1 March 2020).
- Babić, V.; Vančetović, J.; Prodanović, S.; Kravić, N.; Babić, M.; Anđelković, V. Numerical Classification of Western Balkan Maize Drought Tolerant Landraces. J. Agric. Sci. Technol. 2015, 17, 455–468. Available online: http://jast.modares.ac.ir/article_12232_8711c875192edd074d9cf7792c6a0ce2.pdf (accessed on 7 March 2021).
- Popović, A.; Kravić, N.; Babić, M.; Prodanović, S.; Sečanski, M.; Babić, V. Breeding potential of maize landraces evaluated by their testcross performance. Zemdirbyste 2020, 107, 153–160. [Google Scholar] [CrossRef]
- Metzger, M.J.; Brus, D.J.; Bunce, R.G.H.; Carey, P.D.; Gonçalves, J.; Honrado, J.P.; Jongman, R.H.G.; Trabucco, A.; Zomer, R. Environmental stratifications as the basis for national, European and global ecological monitoring. Ecol. Indic. 2013, 33, 26–35. [Google Scholar] [CrossRef]
- Ghosh, D.K.; Divecha, J. Two associate class partially balanced incomplete block designs and partial diallel crosses. Biometrika 1997, 84, 245–248. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Gilmour, A.R. Echidna Mixed Model Software. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 11–16 February 2018. [Google Scholar]
- Meier, U. A note on the power of Fisher’s least significant difference procedure. Pharm. Stat. 2006, 5, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Kosgey, J.R.; Moot, D.J.; Fletcher, A.L.; McKenzie, B.A. Dry matter accumulation and post-silking N economy of ‘stay-green’ maize (Zea mays L.) hybrids. Eur. J. Agron. 2013, 51, 43–52. [Google Scholar] [CrossRef]
- Antonietta, M.; Fanello, D.D.; Acciaresi, H.A.; Guiamet, J.J. Senescence and yield responses to plant density in stay green and earlier-senescing maize hybrids from Argentina. Field Crops Res. 2014, 155, 111–119. [Google Scholar] [CrossRef]
- Liu, W.; Liu, G.; Yang, Y.; Guo, X.; Ming, B.; Xie, R.; Liu, Y.; Wang, K.; Hou, P.; Li, S. Spatial variation of maize height morphological traits for the same cultivars at a large agroecological scale. Eur. J. Agron. 2021, 130, 126349. [Google Scholar] [CrossRef]
- Rana, M.; Sood, A.; Hussain, W.; Kaldate, R.; Sharma, T.R.; Gill, R.K.; Kumar, S.; Singh, S. Chapter 6—Gene Pyramiding and Multiple Character Breeding. In Lentils: Potential Resources for Enhancing Genetic Gains, 1st ed.; Singh, M., Ed.; Academic Press: London, UK, 2019; pp. 83–124. [Google Scholar] [CrossRef]
- Reif, J.C.; Hamrit, S.; Hechenberger, M.; Schipprack, W.; Maurer, H.P.; Bohn, M.; Melchinger, A.E. Trends in genetic diversity among European maize cultivars and their parental components during the past 50 years. Theor. Appl. Genet. 2005, 111, 838–845. [Google Scholar] [CrossRef]
- Wilde, K.; Burger, H.; Prigge, V.; Presterl, T.; Schmidt, W.; Ouzunova, M.; Geiger, H.H. Testcross performance of doubled-haploid lines developed from European flint maize landraces. Plant Breed. 2010, 129, 181–185. [Google Scholar] [CrossRef]
- Gouesnard, B.; Negro, S.; Laffray, A.; Glaubitz, J.; Melchinger, A.; Revilla, P.; Moreno Gonzalez, J.; Madur, D.; Combes, V.; Tollon Cordet, C.; et al. Genotyping-by-sequencing highlights original diversity patterns within a European collection of 1191 maize flint lines, as compared to the maize USDA genebank. Theor. Appl. Genet. 2017, 130, 2165–2189. [Google Scholar] [CrossRef]
- Böhm, J.; Schipprack, W.; Mirdita, V.; Utz, H.F.; Melchinger, A.E. Breeding potential of European flint maize landraces evaluated by testcross performance. Crop Sci. 2014, 54, 1665–1672. [Google Scholar] [CrossRef]
- Böhm, J.; Schipprack, W.; Utz, H.F.; Melchinger, A.E. Tapping the genetic diversity of landraces in allogamous crops with doubled haploid lines: A case study from European flint maize. Theor. Appl. Genet. 2017, 130, 861–873. [Google Scholar] [CrossRef]
- Melani, M.D.; Carena, M.J. Alternative maize heterotic patterns for the Northern Corn Belt. Crop Sci. 2005, 45, 2186–2194. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J.; Miranda Filho, J.B. Quantitative genetics in maize breeding. In Handbook of Plant Breeding, 3rd ed.; Prohens, J., Nuez, F., Carena, M.J., Eds.; Springer: New York, NY, USA, 2010; Volume 6, pp. 1–663. [Google Scholar] [CrossRef]
- Chassaigne-Ricciulli, A.A.; Mendoza-Onofre, L.E.; Córdova-Téllez, L.; Carballo-Carballo, A.; San Vicente-García, F.M.; Dhliwayo, T. Effective Seed Yield and Flowering Synchrony of Parents of CIMMYT Three-Way-Cross Tropical Maize Hybrids. Agriculture 2021, 11, 161. [Google Scholar] [CrossRef]
- Allier, A.; Teyssèdre, S.; Lehermeier, C.; Claustres, B.; Maltese, S.; Melkior, S.; Moreau, L.; Charcosset, A. Assessment of breeding programs sustainability: Application of phenotypic and genomic indicators to a North European grain maize program. Theor. Appl. Genet. 2019, 132, 1321–1334. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Munder, S.; Mauch, F.J.; Merdita, V.; Böhm, J.; Müller, J. High-throughput platform for automated sorting and selection of single seeds based on time-domain nuclear magnetic resonance (TD-NMR) measurement of oil content. Biosyst. Eng. 2017, 164, 213–220. [Google Scholar] [CrossRef]
- Allier, A.; Teyssèdre, S.; Lehermeier, C.; Moreau, L.; Charcosset, A. Optimized breeding strategies to harness genetic resources with different performance levels. BMC Genomics 2020, 21, 349. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GD | AN | CC | DT | DS | PH | EH | KRN | KNR | EL | KH | KD | KW | GY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | 1890 | CRO | 46 | 47 | 139.0 | 41.3 | 12.2 | 20.3 | 11.8 | 3.8 | 2.2 | 276 | 4.276 |
| G2 | 36 | MNE | 51 | 52 | 139.8 | 40.9 | 9.9 | 24.9 | 12.2 | 4.0 | 2.0 | 328 | 3.482 |
| G3 | 869 | SRB | 44 | 46 | 135.3 | 29.5 | 14.0 | 22.1 | 11.4 | 4.0 | 2.0 | 220 | 1.656 |
| G4 | 1962 | BIH | 46 | 47 | 151.4 | 47.3 | 13.0 | 23.0 | 12.9 | 3.8 | 2.2 | 310 | 3.678 |
| G5 | 466 | BIH | 47 | 46 | 149.0 | 41.0 | 14.3 | 20.7 | 10.2 | 3.6 | 2.4 | 260 | 2.766 |
| G6 | 392 | SRB | 48 | 49 | 172.4 | 68.8 | 13.2 | 24.2 | 11.9 | 4.0 | 2.0 | 242 | 3.325 |
| G7 | 594 | MAC | 48 | 50 | 167.4 | 58.6 | 14.8 | 28.3 | 14.0 | 3.8 | 2.2 | 227 | 3.038 |
| G8 | 789 | SRB | 50 | 51 | 149.4 | 42.9 | 12.8 | 18.1 | 8.9 | 4.0 | 2.0 | 262 | 2.926 |
| G9 | 1379 | BIH | 51 | 52 | 164.3 | 48.8 | 12.1 | 29.0 | 14.5 | 4.0 | 2.0 | 260 | 2.119 |
| G10 | 1320 | MNE | 53 | 53 | 169.8 | 56.7 | 11.4 | 24.0 | 11.7 | 3.7 | 2.3 | 354 | 4.058 |
| G11 | 142 | SRB | 56 | 58 | 145.4 | 69.8 | 14.0 | 23.9 | 10.9 | 4.0 | 2.0 | 238 | 3.349 |
| G12 | 625 | SRB | 56 | 58 | 160.5 | 54.0 | 13.5 | 28.3 | 13.0 | 3.0 | 3.0 | 240 | 3.852 |
| G13 | 1267 | MNE | 56 | 57 | 150.7 | 44.2 | 10.3 | 30.0 | 15.0 | 2.9 | 3.1 | 245 | 4.534 |
| G14 | 873 | MAC | 47 | 46 | 148.4 | 38.2 | 10.7 | 27.0 | 10.1 | 2.5 | 3.5 | 254 | 3.714 |
| G15 | 586 | MAC | 47 | 49 | 158.8 | 44.7 | 9.7 | 26.5 | 13.0 | 3.8 | 2.2 | 315 | 3.666 |
| G16 | 1781 | SRB | 47 | 48 | 157.7 | 33.5 | 8.8 | 28.0 | 13.5 | 3.9 | 2.1 | 328 | 4.124 |
| G17 | 13 | MNE | 50 | 53 | 156.8 | 50.3 | 15.2 | 21.8 | 9.5 | 4.0 | 2.0 | 183 | 3.138 |
| G18 | 64 | SRB | 53 | 59 | 154.8 | 42.3 | 8.2 | 32.3 | 17.1 | 4.0 | 2.0 | 342 | 3.040 |
| G19 | 1586 | BIH | 56 | 57 | 144.8 | 45.0 | 9.7 | 25.3 | 13.2 | 4.0 | 2.0 | 300 | 2.589 |
| G20 | 144 | SRB | 58 | 58 | 148.6 | 65.2 | 12.5 | 35.8 | 14.1 | 4.0 | 2.0 | 180 | 4.388 |
| G21 | 1883 | CRO | 45 | 46 | 152.0 | 40.5 | 10.4 | 22.9 | 12.7 | 3.4 | 2.6 | 382 | 3.142 |
| G22 | 1960 | BIH | 47 | 48 | 159.4 | 47.4 | 11.4 | 27.8 | 14.1 | 3.4 | 2.6 | 362 | 4.841 |
| G23 | 888 | MAC | 48 | 49 | 166.0 | 51.3 | 13.2 | 26.0 | 14.3 | 1.9 | 3.8 | 280 | 3.255 |
| G24 | 644 | SRB | 52 | 53 | 184.8 | 75.0 | 9.2 | 24.0 | 14.0 | 3.3 | 2.7 | 400 | 2.939 |
| G25 | 1367 | BIH | 53 | 54 | 184.4 | 69.2 | 11.4 | 26.7 | 12.2 | 2.5 | 3.5 | 328 | 3.554 |
| G26 | 1276 | MNE | 56 | 57 | 169.0 | 51.8 | 9.8 | 30.0 | 14.3 | 2.2 | 3.7 | 428 | 4.970 |
| G27 | 642 | SRB | 50 | 54 | 173.6 | 52.4 | 11.0 | 27.9 | 13.5 | 3.8 | 2.2 | 307 | 4.286 |
| G28 | 1961 | BIH | 45 | 46 | 164.3 | 50.0 | 11.2 | 29.9 | 14.4 | 3.2 | 2.8 | 342 | 3.930 |
| G29 | 1895 | CRO | 46 | 48 | 170.5 | 53.2 | 13.0 | 31.9 | 16.9 | 3.7 | 2.3 | 265 | 5.143 |
| G30 | 190 | SLO | 50 | 52 | 166.4 | 39.8 | 11.8 | 27.8 | 15.8 | 3.8 | 2.2 | 322 | 3.265 |
| G31 | 1931 | CRO | 49 | 51 | 153.7 | 51.7 | 19.3 | 22.6 | 9.8 | 3.9 | 2.1 | 238 | 3.084 |
| G32 | 1324 | MNE | 52 | 53 | 168.2 | 48.8 | 14.8 | 23.1 | 12.0 | 4.0 | 2.0 | 235 | 3.340 |
| G33 | 1972 | BIH | 52 | 53 | 191.2 | 65.0 | 12.7 | 23.9 | 15.1 | 4.0 | 2.0 | 428 | 5.503 |
| G34 | 1936 | CRO | 49 | 50 | 185.7 | 63.3 | 13.4 | 34.5 | 16.2 | 3.4 | 2.6 | 310 | 5.076 |
| G35 | 2249 | BIH | 54 | 56 | 231.8 | 86.7 | 14.3 | 41.5 | 18.1 | 2.3 | 3.7 | 360 | 7.738 |
| G36 | 1992 | SRB | 49 | 51 | 179.5 | 65.8 | 11.5 | 31.2 | 15.3 | 3.2 | 2.8 | 338 | 5.181 |
| G37 | 1945 | BIH | 50 | 51 | 195.5 | 68.3 | 13.4 | 34.2 | 15.8 | 3.6 | 2.4 | 288 | 4.445 |
| 102NS | 53 | 54 | 167.8 | 52.0 | 14.9 | 24.8 | 16.2 | 2.5 | 3.5 | 298 | 3.20 | ||
| 14NS | 50 | 51 | 135.0 | 39.0 | 11.8 | 18.3 | 14.3 | 4.0 | 2.0 | 205 | 2.18 |
| Trait | PCA Component | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| KRN | 0.246 | −0.767 | −0.154 |
| KNR | 0.435 | 0.026 | 0.781 |
| EL | 0.285 | 0.151 | 0.831 |
| KW | −0.138 | 0.869 | 0.073 |
| GY | 0.816 | 0.130 | 0.230 |
| PH | 0.800 | −0.332 | 0.241 |
| EH | 0.827 | −0.342 | 0.171 |
| GM | 0.186 | 0.644 | −0.555 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babic, V.; Stanisavljevic, D.; Zoric, M.; Mikic, S.; Mitrovic, B.; Andjelkovic, V.; Kravic, N. Identification of New Sources for Earliness and Low Grain Moisture at Harvest through Maize Landraces’ Test-Cross Performance. Agronomy 2022, 12, 1939. https://doi.org/10.3390/agronomy12081939
Babic V, Stanisavljevic D, Zoric M, Mikic S, Mitrovic B, Andjelkovic V, Kravic N. Identification of New Sources for Earliness and Low Grain Moisture at Harvest through Maize Landraces’ Test-Cross Performance. Agronomy. 2022; 12(8):1939. https://doi.org/10.3390/agronomy12081939
Chicago/Turabian StyleBabic, Vojka, Dusan Stanisavljevic, Miroslav Zoric, Sanja Mikic, Bojan Mitrovic, Violeta Andjelkovic, and Natalija Kravic. 2022. "Identification of New Sources for Earliness and Low Grain Moisture at Harvest through Maize Landraces’ Test-Cross Performance" Agronomy 12, no. 8: 1939. https://doi.org/10.3390/agronomy12081939
APA StyleBabic, V., Stanisavljevic, D., Zoric, M., Mikic, S., Mitrovic, B., Andjelkovic, V., & Kravic, N. (2022). Identification of New Sources for Earliness and Low Grain Moisture at Harvest through Maize Landraces’ Test-Cross Performance. Agronomy, 12(8), 1939. https://doi.org/10.3390/agronomy12081939