
Genome-Wide Identification of Long Non-Coding RNAs in Pearl Millet (Pennisetum glaucum (L.)) Genotype Subjected to Drought Stress


 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Data Pre-Preprocessing
2.2. Candidate lncRNA Prediction
2.3. Identification of Differentially Expressed lncRNAs, Their Characterization and Annotation
2.4. Identification of miRNAs Targeting the lncRNAs and Their Target mRNAs
2.5. Development of Pearl Millet lncRNAs Database (PMDlncRDB)
3. Results
3.1. Data Pre-Preprocessing
3.2. Candidate lncRNA Prediction
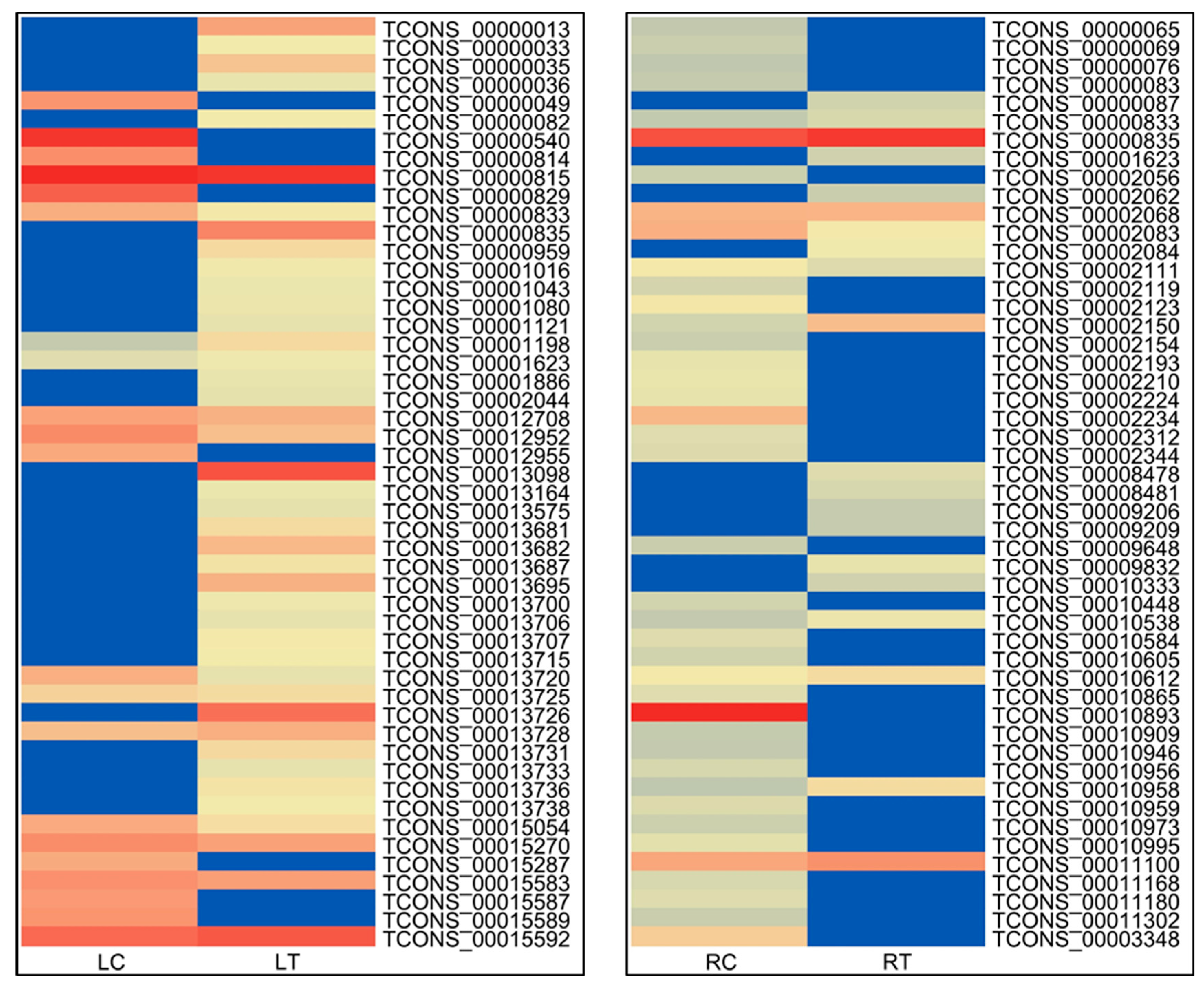
3.3. Identification of Differentially Expressed lncRNAs, Their Characterization and Annotation
3.4. Identification of miRNAs Targeting the lncRNAs
3.5. Development of Pearl Millet lncRNAs Database (PMDlncRDB)
4. Discussion
4.1. Data Pre-Processing
4.2. Candidate lncRNA Prediction
4.3. Identification of Differentially Expressed lncRNAs and Their Annotation
4.4. Identification of miRNAs Targeting the lncRNAs
4.5. lncRNA-miRNA-mRNA Interaction
4.6. Development of Pearl Millet lncRNAs Database (PMDlncRDB)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattick, J.S. RNA regulation: A new genetics? Nat. Rev. Genet. 2004, 5, 316. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Amaral, P.P.; Clark, M.B.; Gascoigne, D.K.; Dinger, M.E.; Mattick, J.S. lncRNAdb: A reference database for long noncoding RNAs. Nucleic Acids Res. 2010, 39 (Suppl. S1), D146–D151. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Sunwoo, H.; Spector, D.L. Long noncoding RNAs: Functional surprises from the RNA world. Genes Dev. 2009, 23, 1494–1504. [Google Scholar] [CrossRef] [Green Version]
- Amor, B.B.; Wirth, S.; Merchan, F.; Laporte, P.; d’Aubenton-Carafa, Y.; Hirsch, J.; Maizel, A.; Mallory, A.; Lucas, A.; Deragon, J.M.; et al. Novel long non-protein coding RNAs involved in Arabidopsis differentiation and stress responses. Genome Res. 2009, 19, 57–69. [Google Scholar] [CrossRef] [Green Version]
- De Lucia, F.; Dean, C. Long non-coding RNAs and chromatin regulation. Curr. Opin. Plant Biol. 2011, 14, 168–173. [Google Scholar] [CrossRef]
- Song, X.; Sun, L.; Luo, H.; Ma, Q.; Zhao, Y.; Pei, D. Genome-wide identification and characterization of long non-coding RNAs from mulberry (Morus notabilis) RNA-seq Data. Genes 2016, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.L.; Carmichael, G.G. Decoding the function of nuclear long non-coding RNAs. Curr. Opin. Cell Biol. 2010, 22, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Jung, C.; Xu, J.; Wang, H.; Deng, S.; Bernad, L.; Chua, N.H. Genome-wide analysis uncovers regulation of long intergenic noncoding RNAs in Arabidopsis. Plant Cell 2012, 24, 4333–4345. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.C.; Liao, J.Y.; Li, Z.Y.; Yu, Y.; Zhang, J.P.; Li, Q.F.; Chen, Y.Q. Genome-wide screening and functional analysis identify a large number of long noncoding RNAs involved in the sexual reproduction of rice. Genome Biol. 2014, 15, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Liu, X.; Guo, C.; Gu, J.; Xiao, K. Identification and characterization of microRNAs from wheat (Triticum aestivum L.) under phosphorus deprivation. Biotechnology 2013, 22, 113–123. [Google Scholar] [CrossRef]
- Li, L.; Eichten, S.R.; Shimizu, R.; Petsch, K.; Yeh, C.T.; Wu, W.; Muehlbauer, G.J. Genome-wide discovery and characterization of maize long non-coding RNAs. Genome Biol. 2014, 15, R40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Li, S.; Hu, L.; Zhang, C. Genome-wide analysis of long non-coding RNAs (lncRNAs) in two contrasting rapeseed (Brassica napus L.) genotypes subjected to drought stress and re-watering. BMC Plant Biol. 2020, 20, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Yu, X.; Lei, N.; Cheng, Z.; Zhao, P.; He, Y.; Peng, M. Genome-wide identification and functional prediction of cold and/or drought-responsive lncRNAs in cassava. Sci. Rep. 2017, 7, 45981. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.J.; Jung, H.; Jeong, D.H.; Ha, S.H.; Choi, Y.D.; Kim, J.K. Transcriptome profiling of drought responsive noncoding RNAs and their target genes in rice. BMC Genom. 2016, 17, 563. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Han, Z.; Guo, Q.; Liu, Y.; Zheng, Y.; Wu, F.; Jin, W. Identification of maize long non-coding RNAs responsive to drought stress. PLoS ONE 2014, 9, e98958. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Xie, S.; Liu, Y.; Yi, F.; Yu, J. Genome-wide annotation of genes and noncoding RNAs of foxtail millet in response to simulated drought stress by deep sequencing. Plant Mol. Biol. 2013, 83, 459–473. [Google Scholar] [CrossRef]
- Zhang, C.; Tang, G.; Peng, X.; Sun, F.; Liu, S.; Xi, Y. Long non-coding RNAs of switchgrass (Panicum virgatum L.) in multiple dehydration stresses. BMC Plant Biol. 2018, 18, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Q.; Wu, F.; Yan, Z.; Li, J.; Ma, T.; Zhang, Y.; Zhang, J. Differential co-expression networks of long non-coding RNAs and mRNAs in Cleistogenes songorica under water stress and during recovery. BMC Plant Biol. 2019, 19, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidong, Q.I.; Hongping, C.; Zuozhen, Y.; Biaolin, H.U.; Xiangdong, L.; Bing, A.; Fantao, Z. Systematic characterization of long non-coding RNAs and their responses to drought stress in Dongxiang wild rice. Rice Sci. 2020, 27, 21–31. [Google Scholar] [CrossRef]
- Jha, U.C.; Nayyar, H.; Jha, R.; Khurshid, M.; Zhou, M.; Mantri, N.; Siddique, K.H. Long non-coding RNAs: Emerging players regulating plant abiotic stress response and adaptation. BMC Plant Biol. 2020, 20, 466. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Lu, P.; Xu, Y.; Li, Z.; Yu, S.; Liu, J.; Cao, P. PLncDB V2. 0: A comprehensive encyclopedia of plant long noncoding RNAs. Nucleic Acids Res. 2021, 49, D1489–D1495. [Google Scholar] [CrossRef] [PubMed]
- Gallart, A.P.; Pulido, A.H.; De Lagrán, I.A.M.; Sanseverino, W.; Cigliano, R.A. GREENC: A Wiki-based database of plant lncRNAs. Nucleic Acids Res. 2016, 44, D1161. [Google Scholar]
- Zhou, B.; Zhao, H.; Yu, J.; Guo, C.; Dou, X.; Song, F.; Wang, J. EVLncRNAs: A manually curated database for long non-coding RNAs validated by low-throughput experiments. Nucleic Acids Res. 2018, 46, D100–D105. [Google Scholar] [CrossRef] [Green Version]
- Quek, X.C.; Thomson, D.W.; Maag, J.L.; Bartonicek, N.; Signal, B.; Clark, M.B.; Dinger, M.E. lncRNAdb v2.0: Expanding the reference database for functional long noncoding RNAs. Nucleic Acids Res. 2015, 43, D168–D173. [Google Scholar] [CrossRef]
- Xuan, H.; Zhang, L.; Liu, X.; Han, G.; Li, J.; Li, X.; Zhang, S. PLNlncRbase: A resource for experimentally identified lncRNAs in plants. Gene 2015, 573, 328–332. [Google Scholar] [CrossRef]
- Szcześniak, M.W.; Bryzghalov, O.; Ciomborowska-Basheer, J.; Makałowska, I. CANTATAdb 2.0: Expanding the collection of plant long noncoding RNAs. Methods Mol. Biol. 2019, 1933, 415–429. [Google Scholar]
- Sweeney, B.A.; Tagmazian, A.A.; Ribas, C.E.; Finn, R.D.; Bateman, A.; Petrov, A.I. Exploring Non-Coding RNAs in RNAcentral. Curr. Protoc. Bioinform. 2020, 71, e104. [Google Scholar] [CrossRef] [PubMed]
- Satyavathi, C.T.; Ambawat, S.; Khandelwal, V.; Srivastava, R.K. Pearl Millet: A Climate-Resilient Nutricereal for Mitigating Hidden Hunger and Provide Nutritional Security. Front. Plant Sci. 2021, 12, 1828. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, E.; White, J.; Seney, S.; Hekmat, S.; McDowell, T.; Sumarah, M.; Reid, G. A Novel Millet-Based Probiotic Fermented Food for the Developing World. Nutrients 2017, 22, 529. [Google Scholar] [CrossRef] [Green Version]
- Sloan, A.E. Positive eating and problem treating: Nutraceuticals and cereal-based foods in the 21st century. Cereal Foods World 1999, 44, 746–750. [Google Scholar]
- Jaiswal, S.; Antala, T.J.; Mandavia, M.K.; Chopra, M.; Jasrotia, R.S.; Tomar, R.S.; Kheni, J.; Angadi, U.B.; Iquebal, M.A.; Golakia, B.A.; et al. Transcriptomic signature of drought response in pearl millet (Pennisetum glaucum (L.) and development of web-genomic resources. Sci. Rep. 2018, 8, 3382. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Sanz-Jimenez, P.; Zhu, X.T.; Feng, J.W.; Shao, L.; Song, J.M.; Chen, L.L. Analysis of rice transcriptome reveals the LncRNA/CircRNA regulation in tissue development. Rice 2021, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Basak, J.; Nithin, C. Targeting non-coding RNAs in plants with the CRISPR-Cas technology is a challenge yet worth accepting. Front. Plant Sci. 2015, 6, 1001. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC, a Quality Control Tool for High throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 12 May 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, X.J.; Butler, G.; Storms, R.; Tsang, A. OrfPredictor: Predicting protein-coding regions in EST-derived sequences. Nucleic Acids Res. 2005, 33 (Suppl. S2), W677–W680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shumayla, S.S.; Taneja, M.; Tyagi, S.; Singh, K.; Upadhyay, S.K. Survey of High Throughput RNA-Seq Data Reveals Potential Roles for lncRNAs during Development and Stress Response in Bread Wheat. Front. Plant Sci. 2017, 8, 1019. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2013, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. HMMER web server: 2015 update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef]
- Tarazona, S.; Furió-Tarí, P.; Turra, D.; Pietro, A.D.; Nueda, M.J.; Ferrer, A.; Conesa, A. Data quality aware analysis of differential expression in RNA-seq with NOISeq R/Bioc package. Nucleic Acids Res. 2015, 43, e140. [Google Scholar]
- Szcześniak, M.W.; Rosikiewicz, W.; Makałowska, I. CANTATAdb: A collection of plant long non-coding RNAs. Plant Cell Physiol. 2015, 57, e8. [Google Scholar] [CrossRef] [Green Version]
- Griffiths-Jones, S. miRBase: The microRNA sequence database. MicroRNA Protoc. 2006, 342, 129–138. [Google Scholar]
- Hofacker, I.L. Vienna RNA secondary structure server. Nucleic Acids Res. 2003, 31, 3429–3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, L.; Zhu, B.; Zhu, H.; Luo, Y.; Wang, Q.; Zuo, J. Integrative analysis of long non-coding RNA acting as ceRNAs involved in chilling injury in tomato fruit. Gene 2018, 667, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Shuai, P.; Liang, D.; Tang, S.; Zhang, Z.; Ye, C.Y.; Su, Y. Genome-wide identification and functional prediction of novel and drought-responsive lincRNAs in Populus trichocarpa. J. Exp. Bot. 2014, 65, 4975–4983. [Google Scholar] [CrossRef] [PubMed]
- Pauli, A.; Valen, E.; Lin, M.F.; Garber, M.; Vastenhouw, N.L.; Levin, J.Z. Systematic identification of long noncoding RNAs expressed during zebrafish embryogenesis. Genome Res. 2012, 22, 577–591. [Google Scholar] [CrossRef] [Green Version]
- Ghatak, A.; Chaturvedi, P.; Nagler, M.; Roustan, V.; Lyon, D.; Bachmann, G.; Weckwerth, W. Comprehensive tissue-specific proteome analysis of drought stress responses in Pennisetum glaucum (L.) R. Br. (Pearl millet). J. Proteom. 2016, 143, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R.K.; Megha, S.; Basu, U.; Rahman, M.H.; Kav, N.N. Genome wide identification and functional prediction of long non-coding RNAs responsive to Sclerotinia sclerotiorum infection in Brassica napus. PLoS ONE 2016, 11, e0158784. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, W.; Yang, Y.; Li, X.; Chen, T.; Liu, T.; Ma, N.; Yang, X.; Liu, R.; Zhang, B. Genome-wide analysis of tomato long non-coding RNAs and identification as endogenous target mimic for microRNA in response to TYLCV infection. Sci. Rep. 2015, 5, 16946. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wang, C.; Bao, H.; Chen, H.; Wang, Y. Genome-wide identification and characterization of novel lncRNAs in Populus under nitrogen deficiency. Mol. Genet. Genom. 2016, 291, 1663–1680. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, X.; Wang, C.; Xu, Z.; Wang, Y.; Liu, X.; Ji, W. Long non-coding genes implicated in response to stripe rust pathogen stress in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2013, 40, 6245–6253. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cho, W.K.; Byun, H.S.; Chavan, V.; Kil, E.J.; Lee, S.; Hong, S.W. Genome-wide identification of long non-coding RNAs in tomato plants irradiated by neutrons followed by infection with Tomato yellow leaf curl virus. PeerJ 2019, 7, 6286–7013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Sahu, S.; Rao, A.R.; Pandey, J.; Gaikwad, K.; Ghoshal, S.; Mohapatra, T. Genome-wide identification and characterization of lncRNAs and miRNAs in cluster bean (Cyamopsis tetragonoloba). Gene 2018, 667, 112–121. [Google Scholar] [CrossRef]
- Ding, J.; Lu, Q.; Ouyang, Y.; Mao, H.; Zhang, P.; Yao, J.; Zhang, Q. A long noncoding RNA regulates photoperiod-sensitive male sterility, an essential component of hybrid rice. Proc. Natl. Acad. Sci. USA 2012, 109, 2654–2659. [Google Scholar] [CrossRef] [Green Version]
- Jabnoune, M.; Secco, D.; Lecampion, C.; Robaglia, C.; Shu, Q.; Poirier, Y. A rice cis-natural antisense RNA acts as a translational enhancer for its cognate mRNA and contributes to phosphate homeostasis and plant fitness. Plant Cell 2013, 25, 4166–4182. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, D.; Zhang, D.; Yin, D.; Zhao, Y.; Ji, C.; Zhu, L. A novel antisense long noncoding RNA, TWISTED LEAF, maintains leaf blade flattening by regulating its associated sense R2R3-MYB gene in rice. New Phytol. 2018, 218, 774–788. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Luo, X.; Sun, F.; Hu, J.; Zha, X.; Su, W.; Yang, J. Overexpressing lncRNA LAIR increases grain yield and regulates neighbouring gene cluster expression in rice. Nat. Commun. 2018, 9, 3516. [Google Scholar] [CrossRef]
- Röhrig, H.; Schmidt, J.; Miklashevichs, E.; Schell, J.; John, M. Soybean ENOD40 encodes two peptides that bind to sucrose synthase. Proc. Natl. Acad. Sci. USA 2002, 99, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Campalans, A.; Kondorosi, A.; Crespi, M. Enod40, a short open reading frame–containing mRNA, induces cytoplasmic localization of a nuclear RNA binding protein in Medicago truncatula. Plant Cell 2004, 16, 1047–1059. [Google Scholar] [CrossRef] [Green Version]
- Gultyaev, A.P.; Roussis, A. Identification of conserved secondary structures and expansion segments in enod40 RNAs reveals new enod40 homologues in plants. Nucleic Acids Res. 2007, 35, 3144–3152. [Google Scholar] [CrossRef] [PubMed]
- Held, M.A.; Penning, B.; Brandt, A.S.; Kessans, S.A.; Yong, W.; Scofield, S.R.; Carpita, N.C. Small-interfering RNAs from natural antisense transcripts derived from a cellulose synthase gene modulate cell wall biosynthesis in barley. Proc. Natl. Acad. Sci. USA 2008, 105, 20534–20539. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.; Liu, X.; Sun, F.; Cao, J.; Huo, N.; Wuda, B.; Yao, Y. Wheat miR9678 affects seed germination by generating phased siRNAs and modulating abscisic acid/gibberellin signaling. Plant Cell 2018, 30, 796–814. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, T.; He, W.; Shen, X.; Zhao, Q.; Bai, J.; You, M. Genome-wide identification and characterization of putative lncRNAs in the diamondback moth, Plutella xylostella (L.). Genomics 2017, 110, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional gene regulation by long noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef] [Green Version]
- Gelaw, T.A.; Sanan-Mishra, N. Non-Coding RNAs in Response to Drought Stress. Int. J. Mol. Sci. 2021, 22, 12519. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zuo, X.; Deng, H.; Liu, X.; Liu, L.; Ji, A. Roles of long noncoding RNAs in brain development, functional diversification and neurodegenerative diseases. Brain Res. Bull. 2013, 97, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Tattini, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytol. 2004, 163, 547–561. [Google Scholar] [CrossRef]
- Kubra, G.; Khan, M.; Munir, F.; Gul, A.; Shah, T.; Hussain, A.; Amir, R. Expression characterization of flavonoid biosynthetic pathway genes and transcription factors in peanut under water deficit conditions. Front. Plant Sci. 2021, 12, 1140. [Google Scholar] [CrossRef]
- Geng, D.; Shen, X.; Xie, Y.; Yang, Y.; Bian, R.; Gao, Y.; Guan, Q. Regulation of phenylpropanoid biosynthesis by MdMYB88 and MdMYB124 contributes to pathogen and drought resistance in apple. Hortic. Res. 2020, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Zhao, J.; Wang, Z.; Cheng, K.; Zhang, P.; Tian, G.; Wang, Y. Transcriptome and metabolite analysis reveal the drought tolerance of foxtail millet significantly correlated with phenylpropanoids-related pathways during germination process under PEG stress. BMC Plant Biol. 2020, 20, 274. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Wu, T.; Pei, T.; Wang, Z.; Yang, H.; Jiang, J.; Xu, X. Overexpression of SlGATA17 promotes drought tolerance in transgenic tomato plants by enhancing activation of the phenylpropanoid biosynthetic pathway. Front. Plant Sci. 2021, 12, 634888. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Raw Reads | Unique Mapped Reads | Multiple Mapped Reads | Overall Aligned Rate |
|---|---|---|---|---|
| Leaf-Control | 3,406,324 | 346,223 (68.58%) | 24,342 (4.82%) | 73.46% |
| Leaf-Treated | 3,103,247 | 1,011,944 (67.86%) | 84,531 (5.67%) | 73.53% |
| Root-Control | 2,272,632 | 762,761 (65.64%) | 48,489 (4.17%) | 69.81% |
| Root-Treated | 3,360,164 | 775,850 (52.66%) | 298,994 (20.29%) | 72.95% |
| TOTAL | 12,142,367 | 2,896,778 (62.54%) | 456,356 (9.85%) | 72.39% |
| Sample | Transcripts | Multi Exon | Multi Exon/Transcripts |
|---|---|---|---|
| Leaf-Control | 4872 | 2159 | 44.31% |
| Leaf-Treated | 12,076 | 6855 | 56.76% |
| Root-Control | 13,506 | 7507 | 55.58% |
| Root-Treated | 9524 | 4988 | 52.37% |
| TOTAL | 39,978 | 21,509 | 53.80% |
| Query ID | Seq ID | Q Coverage | P-Identity | Length | QStart | QEnd | SeqStart | SeqEend | E-Value |
|---|---|---|---|---|---|---|---|---|---|
| TCONS_00024950 | Pgl_GLEAN_10035860 | 99 | 87.793 | 213 | 1 | 212 | 540 | 751 | 1.02 × 10−65 |
| TCONS_00024950 | Pgl_GLEAN_10005667 | 84 | 91.16 | 181 | 1 | 181 | 711 | 891 | 3.67 × 10−65 |
| TCONS_00024950 | Pgl_GLEAN_10005566 | 84 | 91.11 | 180 | 2 | 181 | 688 | 867 | 1.32 × 10−64 |
| TCONS_00024950 | Pgl_GLEAN_10016655 | 84 | 90.608 | 181 | 1 | 181 | 273 | 453 | 1.71 × 10−63 |
| TCONS_00024950 | Pgl_GLEAN_10032756 | 84 | 88.333 | 180 | 2 | 181 | 805 | 984 | 2.88 × 10−56 |
| TCONS_00024950 | Pgl_GLEAN_10030411 | 84 | 87.845 | 181 | 1 | 181 | 981 | 1161 | 3.72 × 10−55 |
| TCONS_00024950 | Pgl_GLEAN_10008530 | 83 | 88.202 | 178 | 2 | 179 | 529 | 706 | 3.72 × 10−55 |
| TCONS_00024950 | Pgl_GLEAN_10036781 | 83 | 87.079 | 178 | 2 | 179 | 55 | 232 | 8.06 × 10−52 |
| Sequence ID | Cantana DB Seq ID | Length | QStart | QEnd | SStart | SEnd | Organism |
|---|---|---|---|---|---|---|---|
| TCONS_00000829 | CNT2084344 | 236 | 1 | 232 | 524 | 758 | Oryza nivara |
| TCONS_00000829 | CNT2084343 | 236 | 1 | 232 | 524 | 758 | Oryza nivara |
| TCONS_00012492 | CNT20135897 | 207 | 7 | 213 | 14 | 220 | Oryza rufipogon |
| TCONS_00012492 | CNT2081201 | 207 | 7 | 213 | 1936 | 2142 | Oryza nivara |
| TCONS_00012492 | CNT20187813 | 207 | 7 | 213 | 1837 | 2043 | Oryza sativa |
| TCONS_00012492 | CNT20187804 | 207 | 7 | 213 | 434 | 228 | Oryza sativa |
| TCONS_00012492 | CNT2020493 | 195 | 7 | 201 | 188 | 6 | Brachypodium distachyon |
| TCONS_00009216 | CNT2099321 | 220 | 7 | 224 | 229 | 11 | Setaria italica |
| TCONS_00013098 | CNT2099766 | 408 | 13 | 416 | 40 | 446 | Setaria italica |
| Mirna | Lncrna | Length | Mismatch | MiRNA Start | MiRNA End | LncRNA Start | LncRNA End |
|---|---|---|---|---|---|---|---|
| dps-mir-2526 | TCONS_00024337 | 27 | 3 | 72 | 98 | 395 | 371 |
| dps-mir-2526 | TCONS_00024337 | 27 | 3 | 49 | 75 | 395 | 371 |
| dps-mir-2526 | TCONS_00024337 | 27 | 3 | 26 | 52 | 395 | 371 |
| dps-mir-2526 | TCONS_00024337 | 27 | 3 | 3 | 29 | 395 | 371 |
| oni-mir-10840-1 | TCONS_00026046 | 29 | 1 | 27 | 53 | 192 | 219 |
| oni-mir-10840-1 | TCONS_00026046 | 25 | 0 | 1 | 23 | 218 | 195 |
| oni-mir-10840-2 | TCONS_00026046 | 26 | 0 | 30 | 53 | 195 | 219 |
| oni-mir-10840-2 | TCONS_00026046 | 28 | 1 | 1 | 26 | 218 | 192 |
| MiRNA_Acc. | Target_Acc. | UPE$ | MiRNA Start | MiRNA End | mRNA Start | mRNA End | Aligned mRNA Sequence | Inhibition |
|---|---|---|---|---|---|---|---|---|
| dps-miR-2526-3p | JT845468.1 | −1 | 1 | 20 | 121 | 140 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | JT845783.1 | −1 | 1 | 20 | 308 | 327 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | KR908724.1 | −1 | 1 | 20 | 770 | 789 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | CD725410.1 | −1 | 1 | 20 | 187 | 206 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | JT844189.1 | −1 | 1 | 20 | 150 | 169 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | CD725449.1 | −1 | 1 | 20 | 20 | 40 | GCCGUCGGCGUGUU-GCGGUG | Cleavage |
| dps-miR-2526-3p | CD725265.1 | −1 | 1 | 20 | 346 | 365 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | JZ681148.1 | −1 | 1 | 20 | 359 | 378 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | JZ681188.1 | −1 | 1 | 20 | 375 | 394 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | EB410974.1 | −1 | 1 | 20 | 379 | 398 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | JZ681109.1 | −1 | 1 | 20 | 102 | 121 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| dps-miR-2526-3p | CD724914.1 | −1 | 1 | 20 | 47 | 66 | GCCGUCGGCGUGUUGCGGUG | Cleavage |
| oni-miR-10840 | CD725654.1 | −1 | 1 | 22 | 199 | 220 | AAUGAGCUCAUUUUGAGAACUU | Cleavage |
| oni-miR-10840 | HQ214676.1 | −1 | 1 | 22 | 1582 | 1603 | AAUGAGCUCAUUUUGAGAACUU | Cleavage |
| oni-miR-10840 | CD725654.1 | −1 | 1 | 22 | 199 | 220 | AAUGAGCUCAUUUUGAGAACUU | Cleavage |
| oni-miR-10840 | HQ214676.1 | −1 | 1 | 22 | 1582 | 1603 | AAUGAGCUCAUUUUGAGAACUU | Cleavage |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, B.; Kumar, A.; Jaiswal, S.; Iquebal, M.A.; Angadi, U.B.; Tomar, R.S.; Rai, A.; Kumar, D. Genome-Wide Identification of Long Non-Coding RNAs in Pearl Millet (Pennisetum glaucum (L.)) Genotype Subjected to Drought Stress. Agronomy 2022, 12, 1976. https://doi.org/10.3390/agronomy12081976
Kumar B, Kumar A, Jaiswal S, Iquebal MA, Angadi UB, Tomar RS, Rai A, Kumar D. Genome-Wide Identification of Long Non-Coding RNAs in Pearl Millet (Pennisetum glaucum (L.)) Genotype Subjected to Drought Stress. Agronomy. 2022; 12(8):1976. https://doi.org/10.3390/agronomy12081976
Chicago/Turabian StyleKumar, Baibhav, Animesh Kumar, Sarika Jaiswal, Mir Asif Iquebal, Ulavappa B. Angadi, Rukam S. Tomar, Anil Rai, and Dinesh Kumar. 2022. "Genome-Wide Identification of Long Non-Coding RNAs in Pearl Millet (Pennisetum glaucum (L.)) Genotype Subjected to Drought Stress" Agronomy 12, no. 8: 1976. https://doi.org/10.3390/agronomy12081976
APA StyleKumar, B., Kumar, A., Jaiswal, S., Iquebal, M. A., Angadi, U. B., Tomar, R. S., Rai, A., & Kumar, D. (2022). Genome-Wide Identification of Long Non-Coding RNAs in Pearl Millet (Pennisetum glaucum (L.)) Genotype Subjected to Drought Stress. Agronomy, 12(8), 1976. https://doi.org/10.3390/agronomy12081976