
The Molecular Composition of Humic Acids in Permafrost Peats in the European Arctic as Paleorecord of the Environmental Conditions of the Holocene




Abstract
:1. Introduction
2. Field Sampling
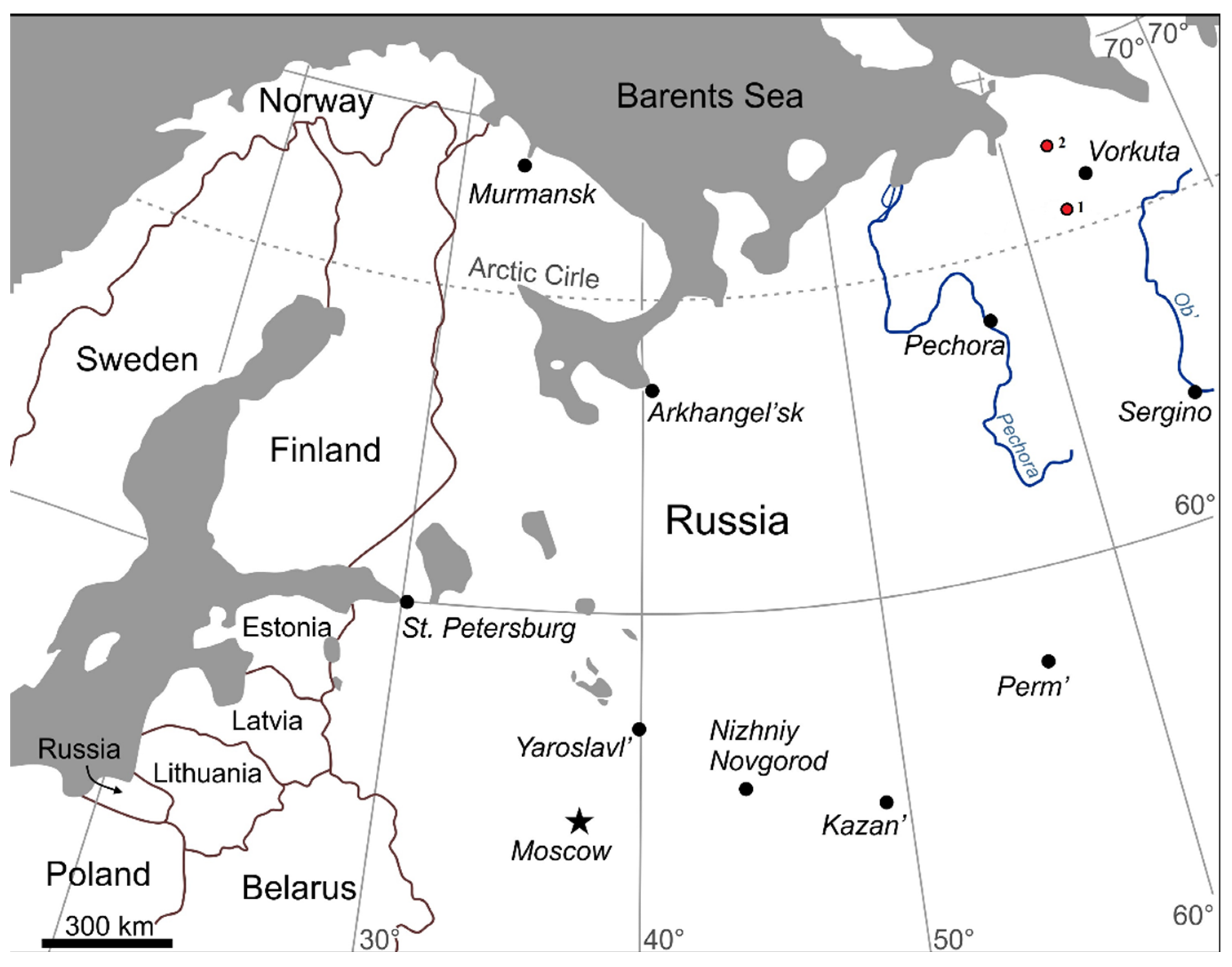
2.1. Geographical Setting
2.2. Sampling Sites
3. Methods
3.1. Soil Analysis
3.2. Radiocarbon Dating
3.3. Botanical Composition and Degree of Peat Decomposition
3.4. Extraction of HAs
3.5. UV-Vis Spectroscopic Analyses of HAs
3.6. Elemental Analyses of HAs
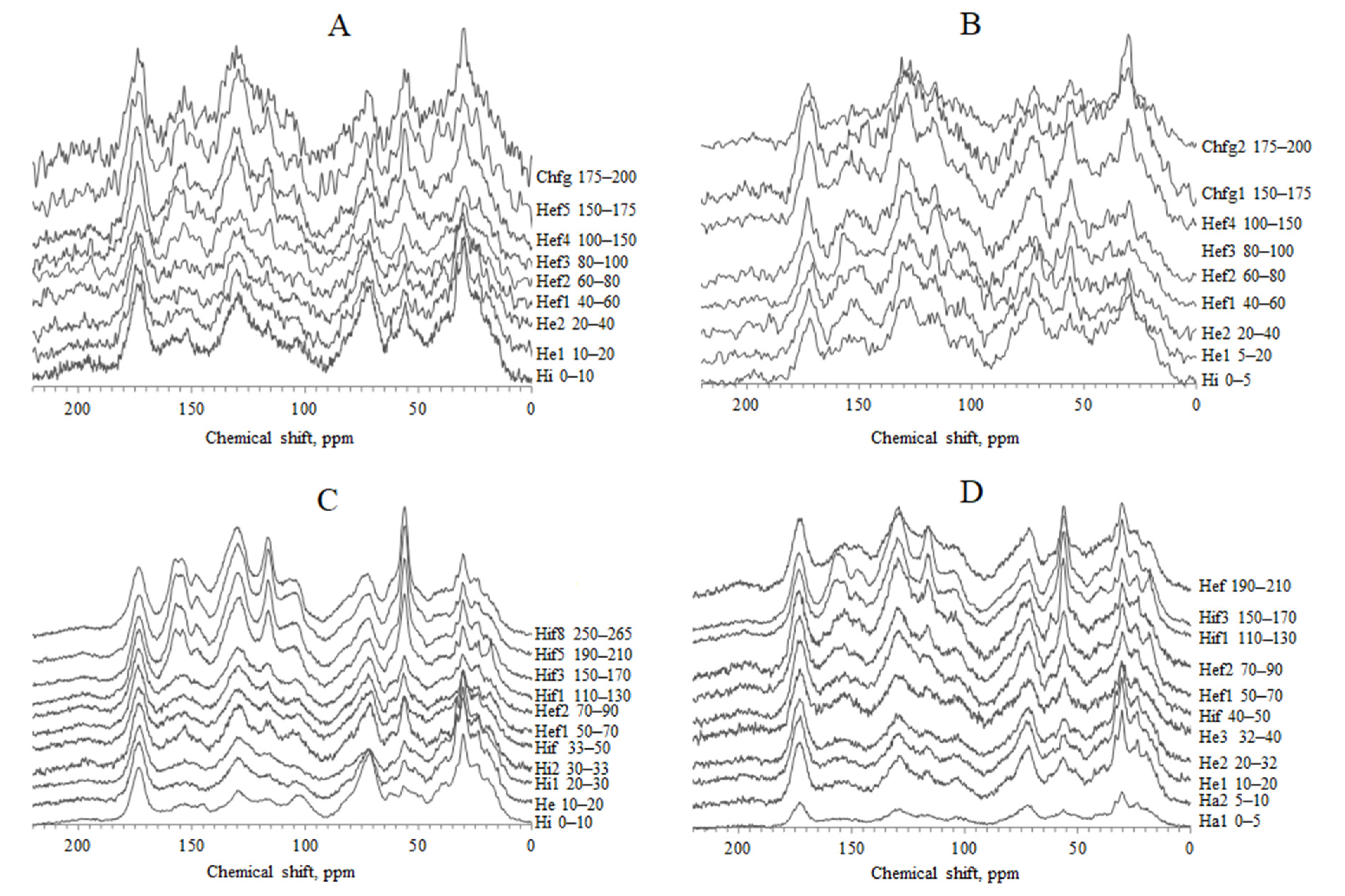
3.7. The 13C NMR Measurements of HAs
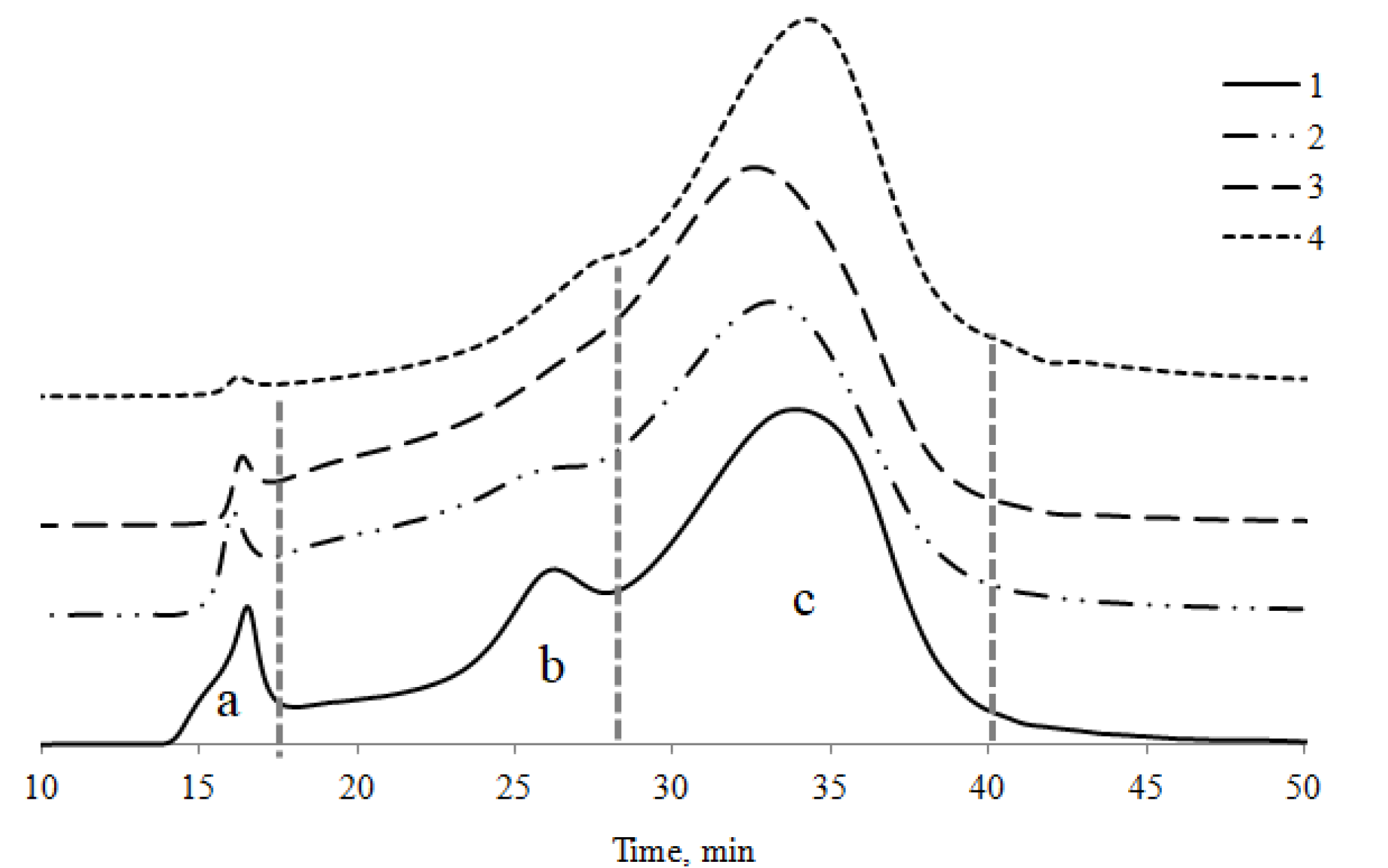
3.8. Size-Exclusion Chromatography Analyses of HAs
3.9. Statistical Analyses
4. Results and Discussion
4.1. Basic Properties of Peat Soils
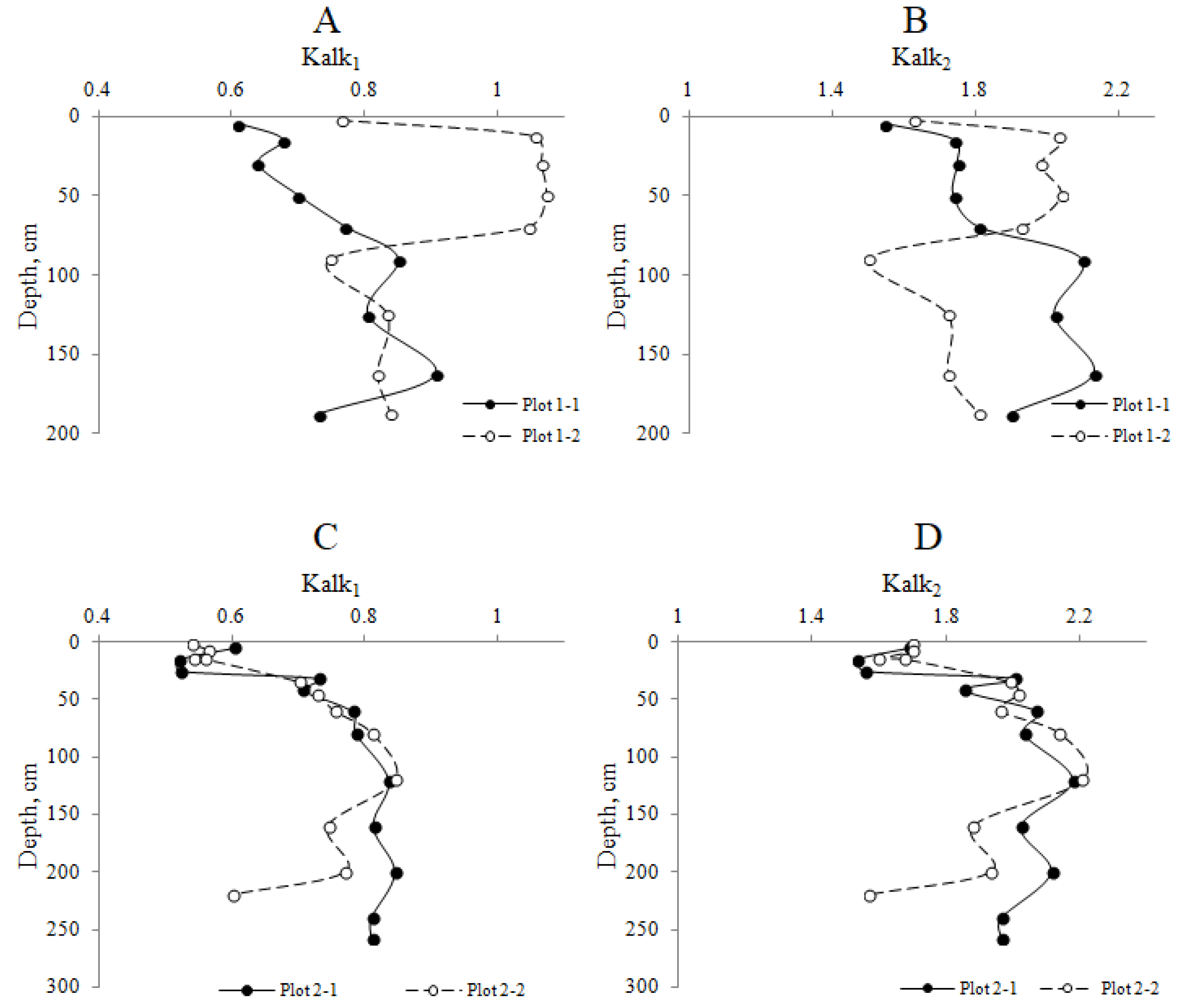
4.2. Elemental Composition and Optical Properties of HAs
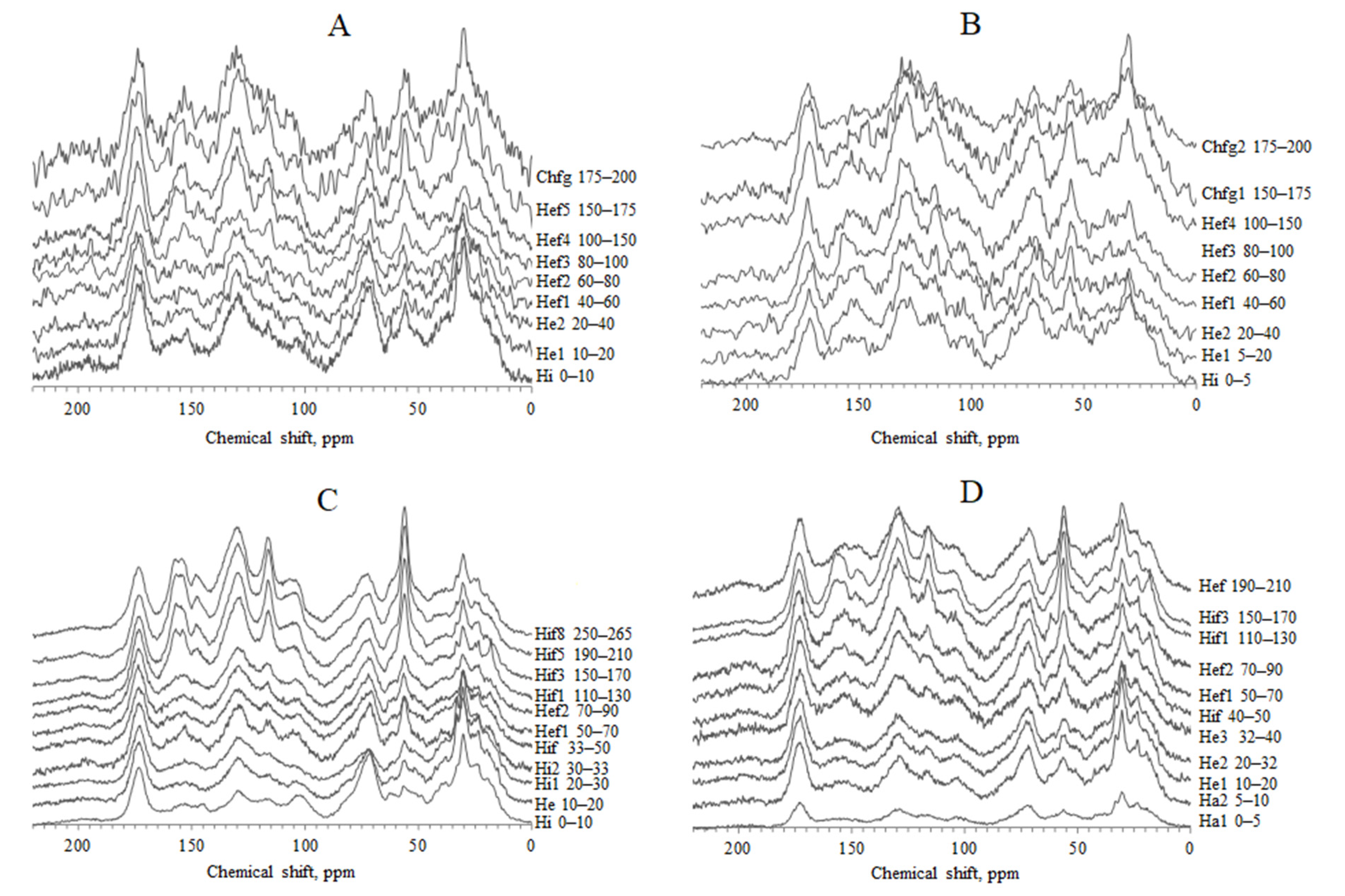
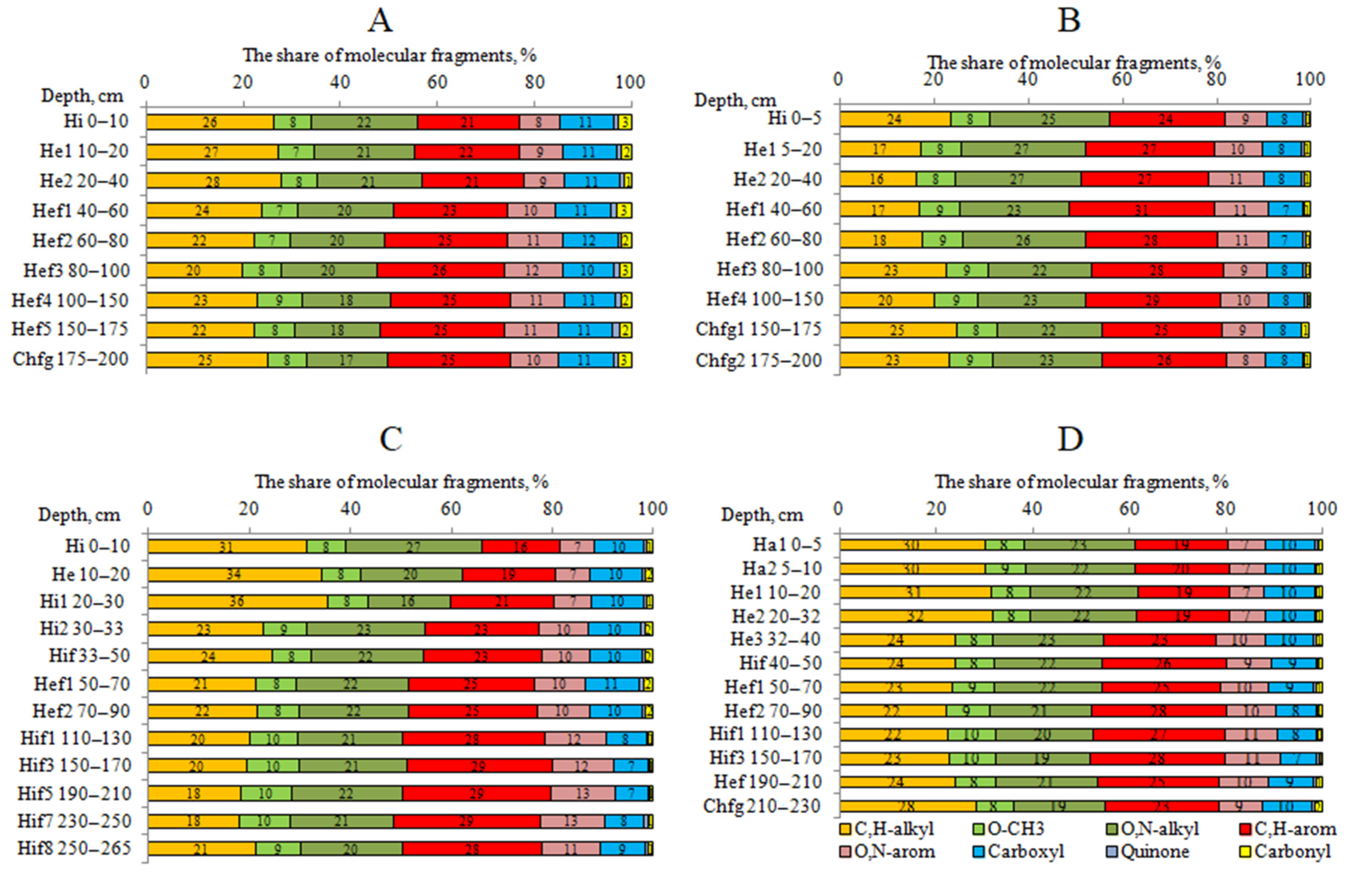
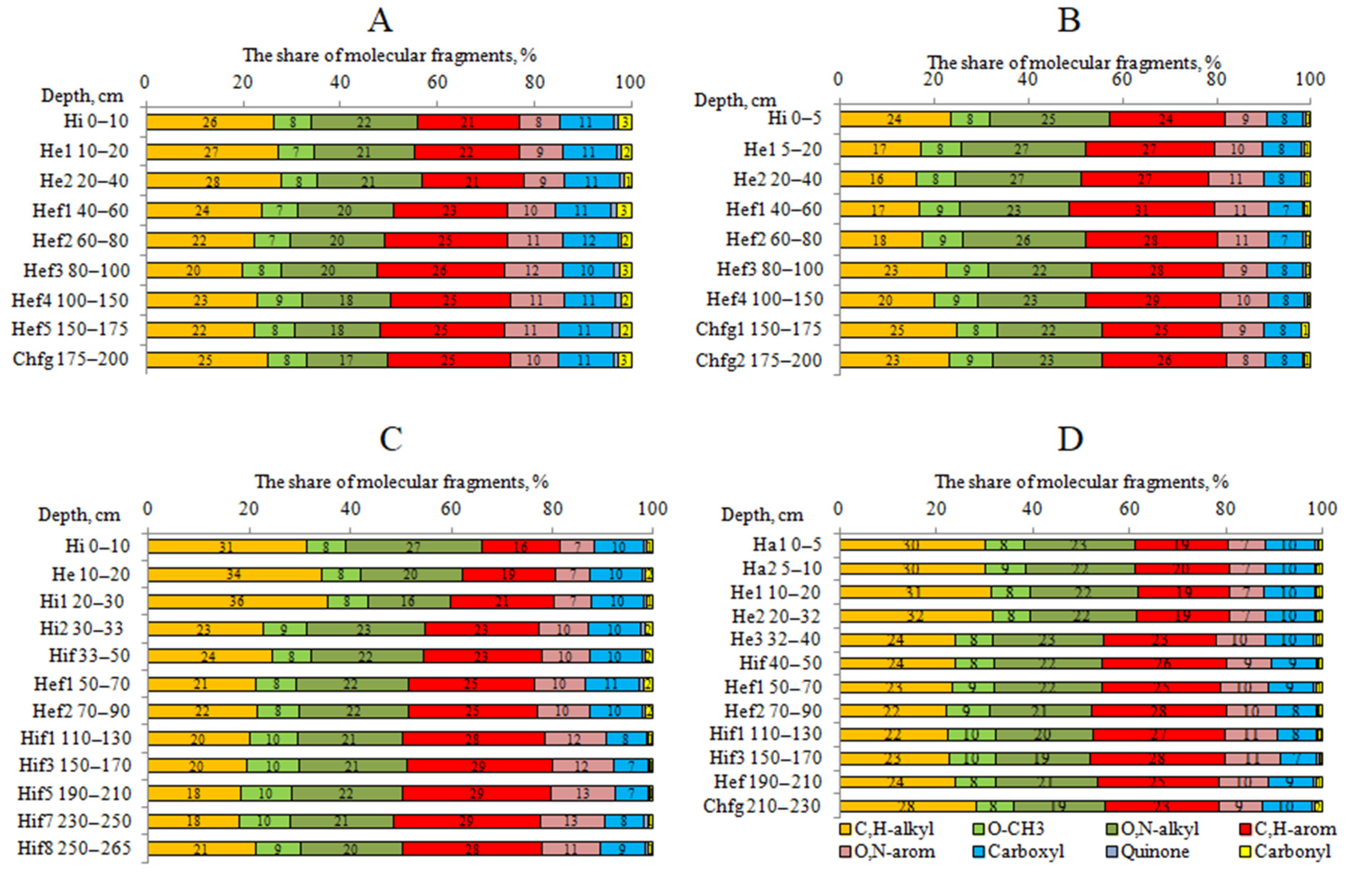
4.3. The 13C NMR Data of HAs
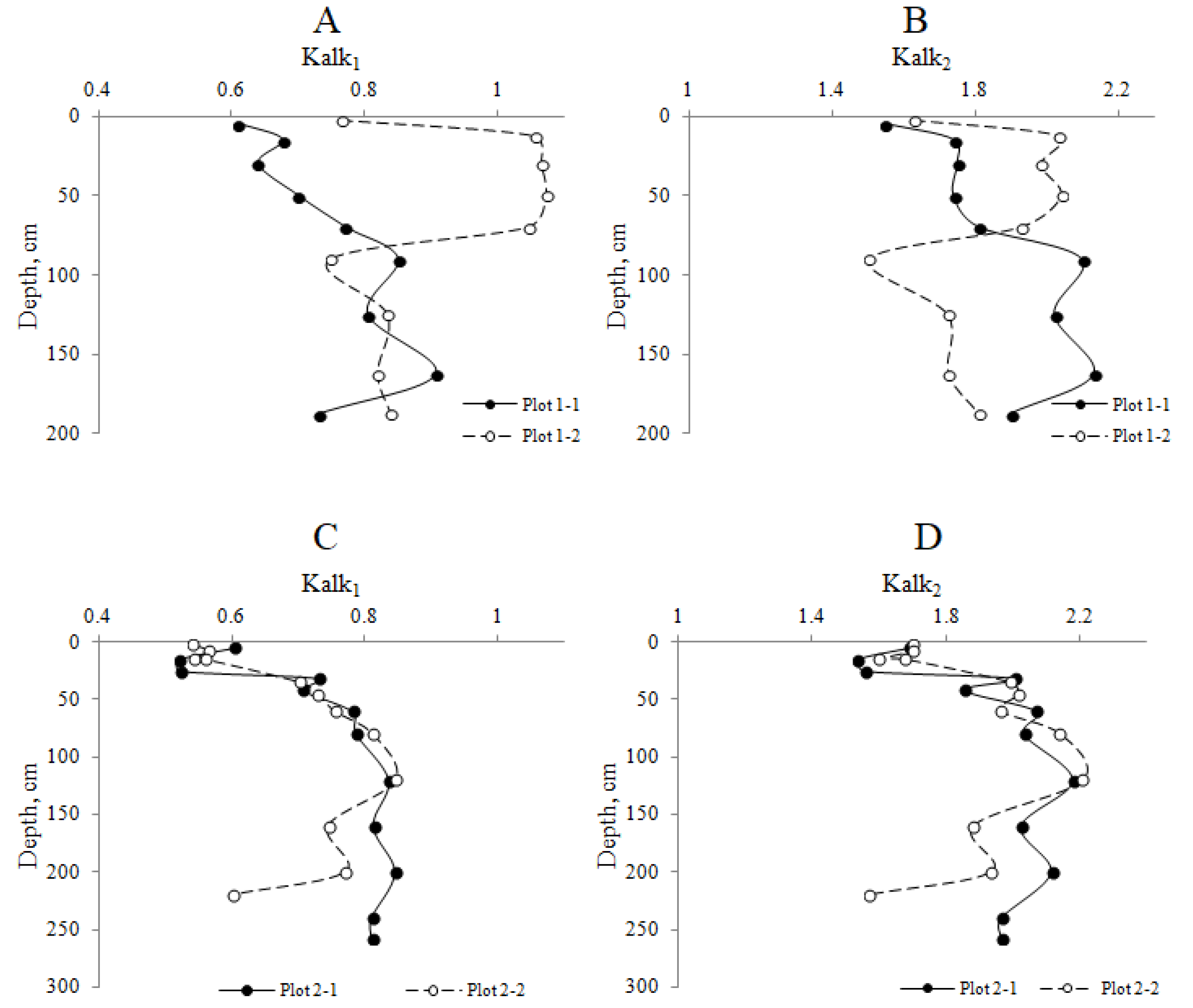
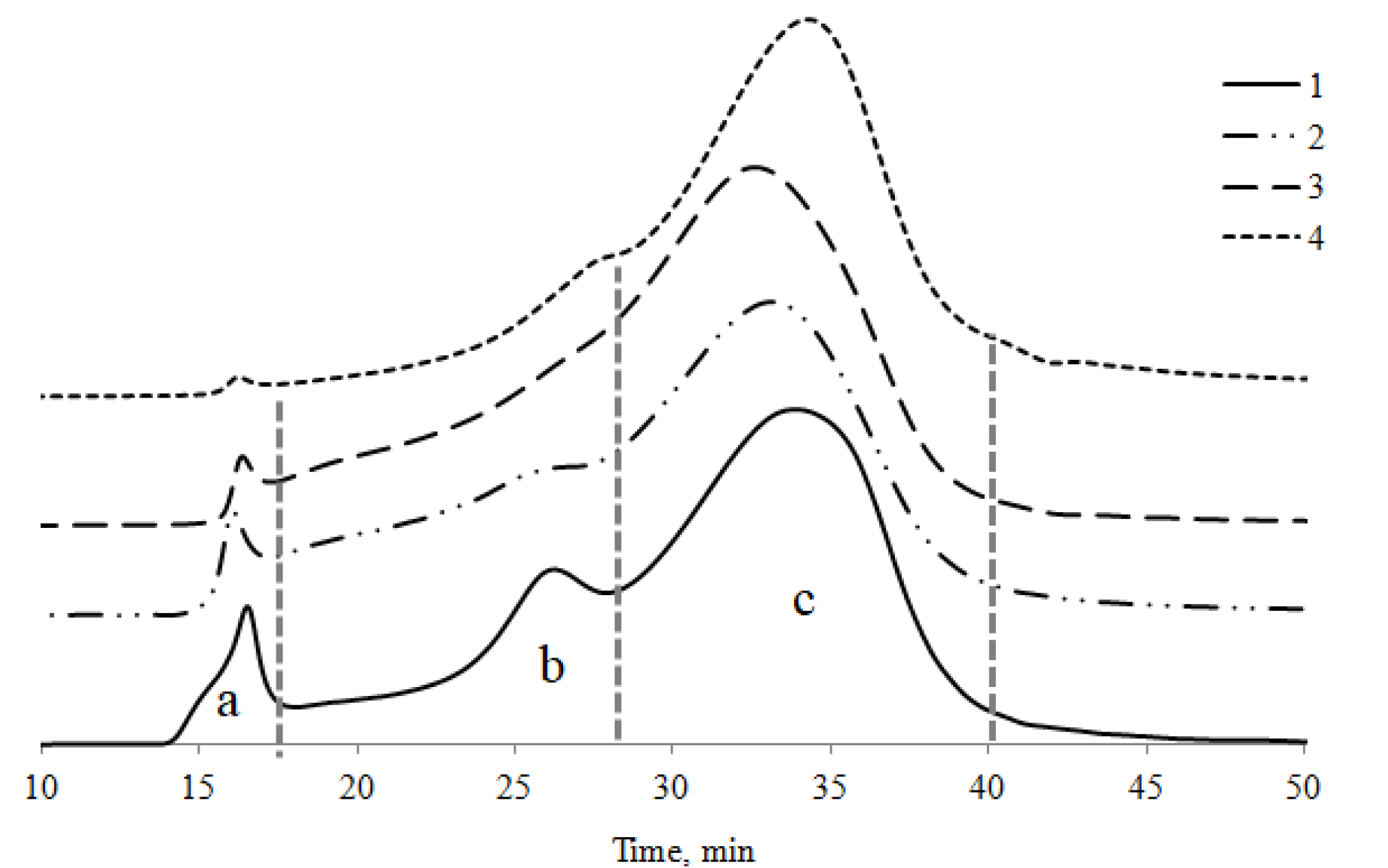
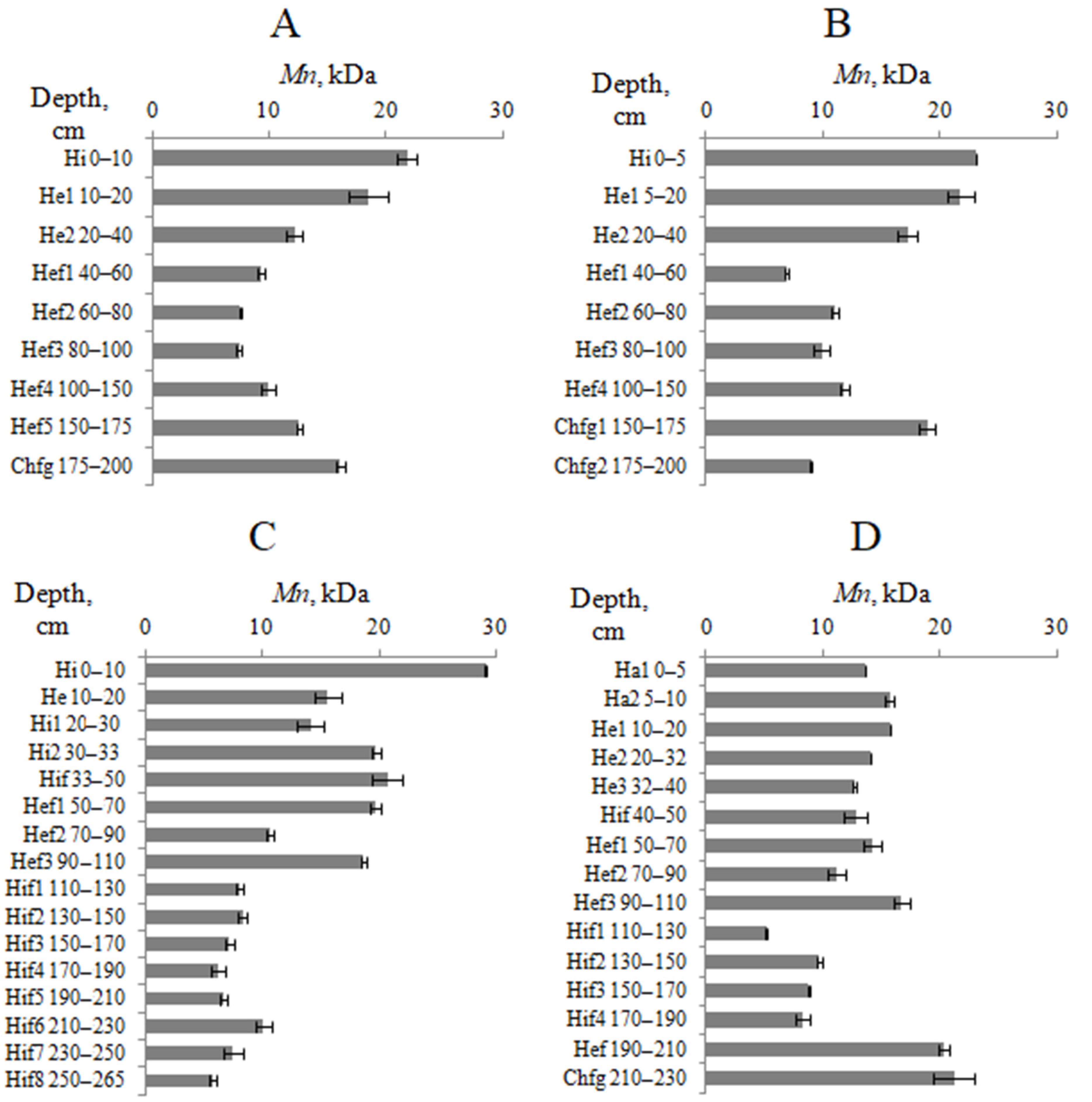
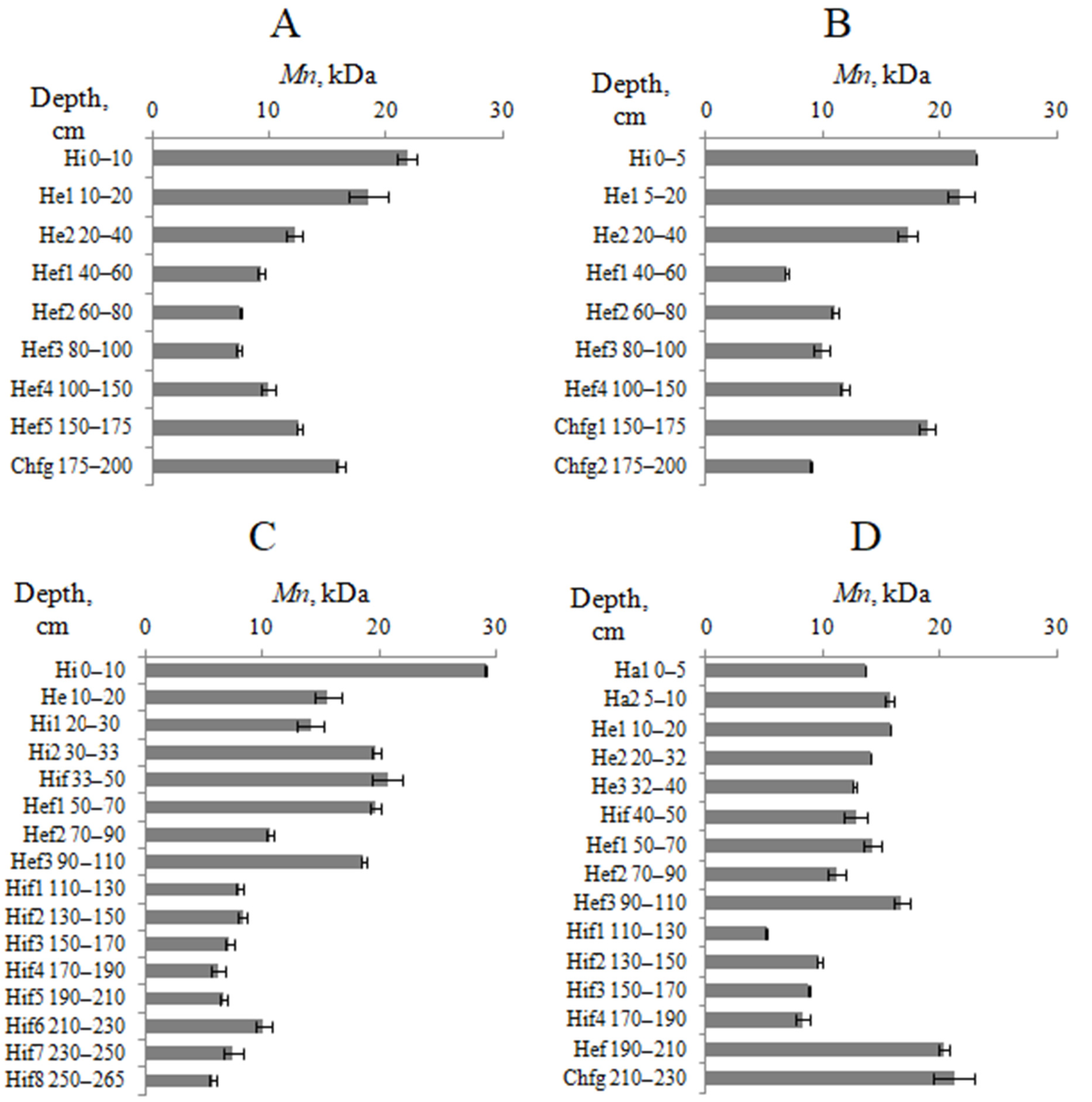
4.4. Molecular Weight Distribution of HAs
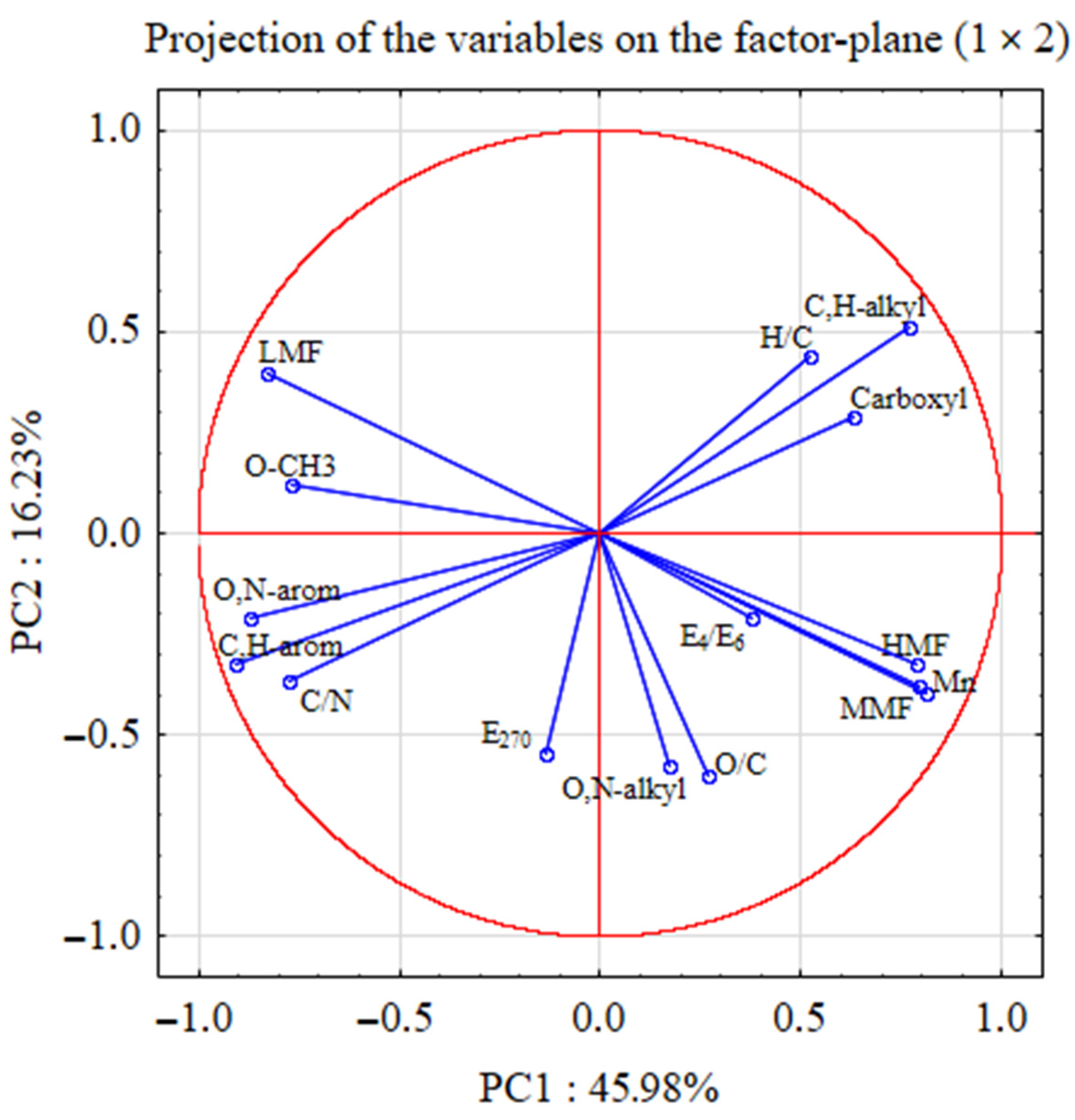
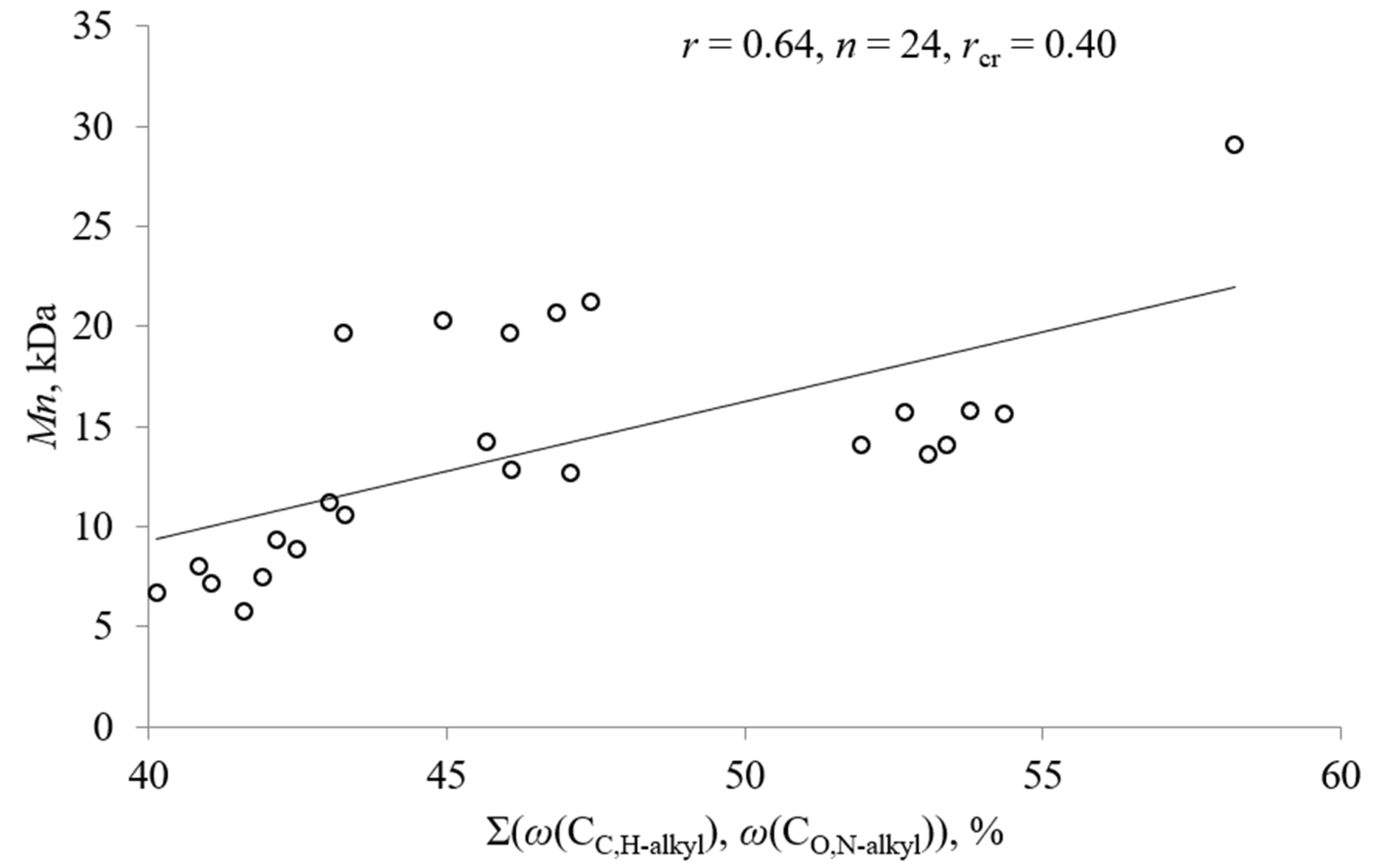
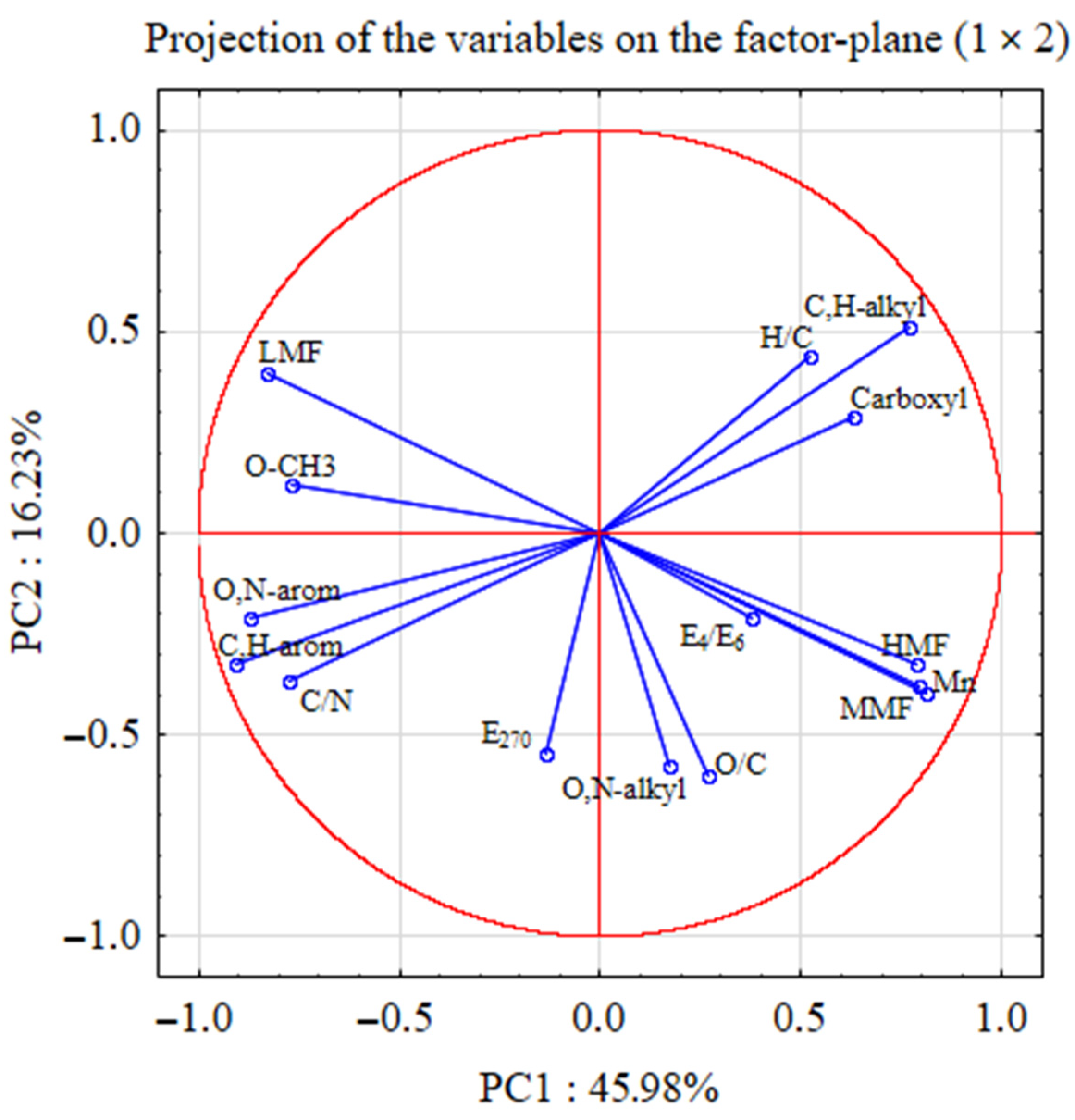
5. Statistical Analyses
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christensen, T.R.; Jonasson, S.; Callaghan, T.V.; Havstrom, M. On the potential CO2 release from tundra soils in a changing climate. Appl. Soil Ecol. 1999, 11, 127–134. [Google Scholar] [CrossRef]
- Zimov, S.A.; Schuur, E.A.G.; Chapin, F.S. Permafrost and the global carbon budget. Science 2006, 312, 1612–1613. [Google Scholar] [CrossRef]
- Schuur, E.A.; Bockheim, G.J.; Canadell, J.G.; Euskirchen, E.; Field, C.B.; Goryachkin, S.V.; Hagemann, S.; Kuhry, P.; Lafleur, P.M.; Lee, H.; et al. Vulnerability of permafrost carbon to climate change: Implications for the global carbon cycle. BioScience 2008, 58, 701–714. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Mcguire, A.D.; Grosse, G.; Harden, J.W.; Hayes, D.J.; Hugelius, G.; Koven, C.D.; Kuhry, P.; Lawrence, D.M.; Natali, S.M.; et al. Climate change and the permafrost carbon feedback. Nature 2015, 520, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Klavinš, M.; Purmališ, O.; Rodinov, V. Peat humic acid properties and factors influencing their variability in a temperate bog ecosystem. Est. J. Ecol. 2013, 62, 35–52. [Google Scholar] [CrossRef]
- Olk, D.C.; Bloom, P.R.; Perdue, E.M.; McKnight, D.M.; Chen, Y.; Farenhorst, A.; Senesi, N.; Chin, Y.-P.; Schmitt-Kopplin, P.; Hertkorn, N.; et al. Environmental and agricultural relevance of humic fractions extracted by alkali from soils and natural waters. J. Environ. Qual. 2019, 48, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Wei, Y.; Huang, J.; Chen, X.; Cai, C. Soil organic carbon stock and fractional distribution across central-south China. Int. Soil Water Conserv. Res. 2021, 9, 620–630. [Google Scholar] [CrossRef]
- Gondar, D.; Lopez, R.; Fiol, S.; Antelo, J.M.; Arce, F. Characterization and acid-base properties of fulvic and humic acids isolated from two horizons of an ombrotrophic peat bog. Geoderma 2005, 126, 367–374. [Google Scholar] [CrossRef]
- Swindles, G.T.; Patterson, T.R.; Roe, H.M.; Galloway, J.M. Evaluating periodicities in peat-based climate proxy records. Quat. Sci. Rev. 2012, 41, 94–103. [Google Scholar] [CrossRef]
- Klavinš, M.; Purmališ, O. Properties and structure of raised bog peat humic acids. J. Mol. Struct. 2013, 1050, 103–113. [Google Scholar] [CrossRef]
- Perminova, I.V.; Hatfield, K. Remediation chemistry of humic substances: Theory and implications for technology. In The Use of Humic Substances to Remediate Polluted Environment: From Theory to Practice; Perminova, I.V., Hatfield, K., Hertkorn, N., Eds.; Science Series IV: Earth and Environmental Sciences; Springer: Dordrecht, The Netherlands, 2005; Volume 52, pp. 3–36. [Google Scholar] [CrossRef]
- García, A.C.; Souza, L.G.A.; Pereira, M.G.; Castro, R.N.; García-Mina, J.M.; Zonta, E.; Lisboa, F.J.G.; Berbara, R.L.L. Structure-property-function relationship in humic substances to explain the biological activity in plants. Sci. Rep. 2016, 6, 20798. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-K.; Hur, J. Using two-dimensional correlation size exclusion chromatography (2D-CoSEC) to explore the size-dependent heterogeneity of humic substances for copper binding. Environ. Pollut. 2017, 227, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Olaetxea, M.; De Hita, D.; Garcia, A.; Fuentes, M.; Baigorri, R.; Mora, V.; Garica, M.; Urrutia, O.; Erro, J.; Zamarreño, A.M.; et al. Hypothetical framework integrating the main mechanisms involved in the promoting action of rhizospheric humic substances on plant root- and shoot growth. Appl. Soil Ecol. 2018, 123, 521–537. [Google Scholar] [CrossRef]
- Tan, K.H. Humic Matter in Soil and the Environment, 1st ed.; Marcel Dekker: New York, NY, USA, 2003; 495p. [Google Scholar]
- Zaccone, C.; Miano, T.M.; Shotyk, W. Qualitative comparison between raw peat and related humic acids in an ombrotrophic bog profile. Org. Geochem. 2007, 38, 151–160. [Google Scholar] [CrossRef]
- D’Orazio, V.; Senesi, N. Spectroscopic properties of humic acids isolated from the rhizosphere and bulk soil compartments and fractionated by size-exclusion chromatography. Soil Biol. Biochem. 2009, 41, 1775–1781. [Google Scholar] [CrossRef]
- Lodygin, E.D.; Beznosikov, V.A. The molecular structure and elemental composition of humic substances from Albeluvisols. Chem. Ecol. 2010, 26, 87–95. [Google Scholar] [CrossRef]
- Asakawa, D.; Iimura, I.; Kiyota, T.; Yanagi, Y.; Fujitake, N. Molecular size fractionation of soil humic acids using preparative high performance size-exclusion chromatography. J. Chromatogr. A 2011, 1218, 6448–6453. [Google Scholar] [CrossRef]
- Kholodov, V.A.; Konstantinov, A.I.; Kudryavtsev, A.V.; Perminova, I.V. Structure of humic acids in zonal soils from 13C NMR data. Eurasian Soil Sci. 2011, 44, 976–983. [Google Scholar] [CrossRef]
- Sartakov, M.P.; Komissarov, I.D.; Shundrin, L.A. The peat humic acids electronic paramagnetism research for Ob-Irtysh flood plains. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1685–1692. [Google Scholar]
- Sartakov, M.P.; Novikov, A.A.; Chukhareva, N.V. Study of humic acids in various types and kinds of peats at Khantymansi autonomous area—Ygra by NMR 13C spectroscopy. Int. J. Pharm. Technol. 2016, 8, 14204–14213. [Google Scholar]
- Kaverin, D.A.; Pastukhov, A.V.; Lapteva, E.M.; Biasi, C.; Marushchak, M.; Martikainen, P. Morphology and properties of the soils of permafrost peatlands in the southeast of the Bol’shezemel’skaya tundra. Eurasian Soil Sci. 2016, 49, 498–511. [Google Scholar] [CrossRef]
- World reference base for soil resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2015; 203p. [Google Scholar]
- Hua, Q.; Barbetti, M.; Rakowski, A.Z. Atmospheric radiocarbon for the period 1950–2010. Radiocarbon 2013, 55, 2059–2072. [Google Scholar] [CrossRef]
- Reimer, P.J.; Bard, E.; Bayliss, A.; Beck, J.W.; Blackwell, P.G.; Bronk, R.C.; Buck, C.E.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 2013, 55, 1869–1887. [Google Scholar] [CrossRef]
- Shishlina, N.; Sevastyanov, V.; Zazovskaya, E.; van der Plicht, J. Reservoir effect of archaeological samples from steppe bronze age cultures in southern Russia. Radiocarbon 2014, 56, 767–778. [Google Scholar] [CrossRef]
- Nikiforova, L.D. Dynamics of Holocene landscape zones in the northeast of the European part of the USSR. In Development of the Nature of the Territory of the USSR in the Late Pleistocene and Holocene; Nauka: Moscow, Russia, 1982; pp. 154–179. (In Russian) [Google Scholar]
- GOST 10650-2013; Peat. Methods for the Determination of the Disintegration Degree. Standartinform: Moscow, Russia, 2014. (In Russian)
- Swift, R.S. Organic matter characterization. In Methods of Soil Analysis: Part 3 Chemical Methods, 5.3; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1018–1020. [Google Scholar] [CrossRef]
- Lodygin, E.D.; Beznosikov, V.A. The 13C NMR study of the molecular structure of humus acids from podzolic and bog-podzolic soils. Eurasian Soil Sci. 2003, 36, 967–975. [Google Scholar]
- Liang, B.C.; Gregorich, E.G.; Schnitzer, M.; Schulten, H.R. Characterization of water extracts of two manures and their absorption on soils. Soil Sci. Soc. Am. J. 1996, 60, 1758–1763. [Google Scholar] [CrossRef]
- Perminova, I.V.; Frimme, F.H.; Kudryavtsev, A.V.; Kulikova, N.A.; Abbt-Braun, G.; Hesse, S.; Petrosyan, V.S. Molecular weight characteristics of humic substances from different environments as determined by size exclusion chromatography and their statistical evaluation. Environ. Sci. Technol. 2003, 37, 2477–2485. [Google Scholar] [CrossRef]
- Szajdak, L.W.; Jezierski, A.; Wegner, K.; Meysner, T.; Szczepański, M. Influence of drainage on peat organic matter: Implications for development, stability and transformation. Molecules 2020, 25, 2587. [Google Scholar] [CrossRef]
- Routh, J.; Hugelius, G.; Kuhry, P.; Filley, T.; Tillman, P.K.; Becher, M.; Crill, P. Multi-proxy study of soil organic matter dynamics in permafrost peat deposits reveal vulnerability to climate change in the European Russian Arctic. Chem. Geol. 2014, 368, 104–117. [Google Scholar] [CrossRef]
- Vasilevich, R.S.; Beznosikov, V.A. Effect of climate changes in the holocene on the distribution of humic substances in the profile of forest-tundra peat mounds. Eurasian Soil Sci. 2017, 50, 1271–1282. [Google Scholar] [CrossRef]
- Orlov, D.S. Humic Substances of Soils and General Theory of Humification, 1st ed.; Taylor & Francis: London, UK, 1995; 325p. [Google Scholar] [CrossRef]
- Sire, J.; Klavins, M. Influence of the humification process on the properties of peat humic acids. Proc. Latv. Acad. Sci. Sect. B 2010, 64, 167–173. [Google Scholar] [CrossRef]
- Dergacheva, M.I.; Nekrasova, O.A.; Okoneshnikova, M.V.; Vasil’eva, D.I.; Gavrilov, D.A.; Ochur, K.O.; Ondar, E.E. Ratio of elements in humic acids as a source of information on the environment of soil formation. Contemp. Probl. Ecol. 2012, 5, 497–504. [Google Scholar] [CrossRef]
- Hugelius, G.; Routh, J.; Kuhry, P.; Crill, C. Mapping the degree of decomposition and thaw remobilization potential of soil organic matter in discontinuous permafrost terrain. J. Geophys. Res. 2012, 117, G02030. [Google Scholar] [CrossRef]
- Trubetskoi, O.A.; Trubetskaya, O.E. Three-dimensional fluorescence analysis of chernozem humic acids and their electrophoretic fractions. Eurasian Soil Sci. 2017, 50, 1018–1024. [Google Scholar] [CrossRef]
- Radmanović, S.; Nikolić, N.; Dordević, A. Humic acids optical properties of rendzina soils in diverse environmental conditions of Serbia. Arch. Tech. Sci. 2018, 18, 63–70. [Google Scholar] [CrossRef]
- Trubetskoi, O.A.; Trubetskaya, O.E. 13CNMR analysis of components of chernozem humic acids and their fractions with different molecular sizes and electrophoretic mobilities. Eurasian Soil Sci. 2011, 44, 281–285. [Google Scholar] [CrossRef]
- Ilina, S.M.; Drozdova, O.Y.; Lapitsky, S.A.; Alekhin, Y.V.; Demin, V.V.; Zavgorodnaya, Y.A.; Shirokova, L.S.; Viers, J.; Pokrovsky, O.S. Size fractionation and optical properties of dissolved organic matter in the continuum soil solution-bogriver and terminal lake of a boreal watershed. Org. Geochem. 2014, 66, 14–24. [Google Scholar] [CrossRef]
- Vinci, G.; Mazzei, P.; Drosos, M.; Zaccone, C.; Piccolo, A. Molecular characterization of ombrotrophic peats by humeomics. Chem. Biol. Technol. Agric. 2020, 7, 18. [Google Scholar] [CrossRef]
- Liu, H.; Zak, D.; Rezanezhad, F.; Lennartz, B. Soil degradation determines release of nitrous oxide and dissolved organic carbon from peatlands. Environ. Res. Lett. 2019, 14, 094009. [Google Scholar] [CrossRef]
- Andreicheva, L.; Marchenko-Vagapova, T.; Buravskaya, M.; Golubeva, Y. Neopleistocene and Holocene Natural Environment of European North-East of Russia, 1st ed.; GEOS: Moscow, Russia, 2015; 224p. (In Russian) [Google Scholar]
- Kovaleva, N.O.; Kovalev, I.V. Lignin phenols in soils as biomarkers of paleovegetation. Eurasian Soil Sci. 2015, 48, 946–958. [Google Scholar] [CrossRef]
- Lodygin, E.; Vasilevich, R. Environmental aspects of molecular composition of humic substances from soils of northeastern European Russia. Polish Polar Res. 2020, 41, 115–135. [Google Scholar] [CrossRef]
- Ricca, G.; Severini, F. Structural investigations of humic substances by IR-FT, 13C-NMR spectroscopy and comparison with a maleic oligomer of known structure. Geoderma 1993, 58, 233–244. [Google Scholar] [CrossRef]
- Vasilevich, R.S.; Lodygin, E.D.; Beznosikov, V.A. Molecular-mass distribution of tundra soils humic substances from the European Northeast of Russia. Biol. Commun. 2015, 4, 103–112. [Google Scholar] [CrossRef]
- Lodygin, E.; Vasilevich, R. Molecular-mass distribution of humic substances from Arctic soils according to size exclusion chromatography. Polish Polar Res. 2020, 41, 1–17. [Google Scholar] [CrossRef]
- Dmitrieva, E.D.; Leontyeva, M.M.; Siundiukova, K.V. Molecular-mass distribution of humic substances and hymatomelanic acids from different origin peats of the Tula region. Khimiya Rastitel’nogo Syr’ya 2017, 4, 187–194. [Google Scholar] [CrossRef]
- Piccolo, A.; Mirabella, A. Molecular weight distribution of peat humic substances extracted with different inorganic and organic solutions. Sci. Total Environ. 1987, 62, 39–46. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Horizon and Depth, cm | Peat Soils Characteristics | HA Characteristics | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 14C-Age (yr BP) | Climate Period | L2, mm/yr | Peat Type | Dominant Botanical Species | R3, % | TOC, % 4 | Ash, % | pH H2O | E2700.001% | E4650.001% | E4/E65 | ΔlogK 6 | Molar Ratios 7 | ||
| H/C | O/C | ||||||||||||||
| Plot 1-1 | |||||||||||||||
| Hi 0–10 | 1450 ± 70 | SA-2 | 0.04 | Raised | Polytrichum | 20–25 | 47.3 | 5.4 | 3.75 | 0.226 | 0.029 | 6.57 | 0.80 | 1.16 ± 0.12 8 | 0.54 ± 0.11 |
| He1 10–20 | n.d. 1 | n.d. | n.d | Raised | Subshrub | 35–40 | 48.8 | 5.9 | 3.70 | 0.246 | 0.033 | 6.67 | 0.80 | 1.10 ± 0.11 | 0.55 ± 0.11 |
| He2 20–40 | 3480 ± 70 | SB-2 | 0.08 | Fen | Carex, Eriophorum | 30–40 | 50.0 | 4.2 | 3.74 | 0.226 | 0.027 | 6.83 | 0.81 | 1.07 ± 0.11 | 0.53 ± 0.11 |
| Hef1⊥ 40–60 | 5230 ± 80 | AT-3 | 0.11 | Fen, frozen | Betula sp., wood | 30–35 | 34.5 | 6.4 | 4.34 | 0.243 | 0.029 | 6.62 | 0.81 | 1.06 ± 0.10 | 0.56 ± 0.10 |
| Hef2⊥ 60–80 | 5680 ± 80 | AT-3 | 0.43 | Fen, frozen | Menyanthes, Carex | 30–35 | 43.7 | 33.1 | 4.86 | 0.250 | 0.030 | 7.17 | 0.85 | 0.98 ± 0.10 | 0.58 ± 0.10 |
| Hef3⊥ 80–100 | 5710 ± 80 | AT-3 | 5.71 | Fen, frozen | Betula sp., wood, Carex | 35–40 | 29.0 | 16.6 | 4.94 | 0.210 | 0.031 | 6.15 | 0.80 | 0.92 ± 0.09 | 0.41 ± 0.09 |
| Hef4⊥ 100–150 | 6130 ± 80 | AT-2 | 0.92 | Fen, frozen | Betula sp., wood, Carex | 30–40 | 24.9 | 47.5 | 4.95 | 0.202 | 0.028 | 6.24 | 0.81 | 0.90 ± 0.09 | 0.38 ± 0.09 |
| Hef5⊥ 150–175 | n.d. | n.d. | n.d | Fen, frozen | Betula sp., wood, Carex | 35–40 | 24.3 | 54.7 | 5.37 | 0.229 | 0.033 | 5.96 | 0.77 | 0.94 ± 0.09 | 0.48 ± 0.10 |
| Chfg⊥ 175–200 | 7160 ± 80 | AT-1 | 0.52 | Fen, frozen | Betula sp., Picea, wood, grass | 50 | 6.2 | 86.3 | 5.33 | 0.234 | 0.033 | 6.32 | 0.81 | 0.98 ± 0.10 | 0.49 ± 0.10 |
| Plot 1-2 | |||||||||||||||
| Hi 0–5 | 1310 ± 70 | SA-2 | 0.04 | Raised | Polytrichum | 20–25 | 46.7 | 8.0 | 3.61 | 0.234 | 0.031 | 7.15 | 0.83 | 1.05 ± 0.10 | 0.52 ± 0.10 |
| He1 5–20 | n.d. | n.d. | n.d | Transitional | Subshrub, Eriophorum | 40–45 | 50.7 | 4.9 | 3.42 | 0.232 | 0.032 | 7.91 | 0.91 | 1.01 ± 0.10 | 0.59 ± 0.10 |
| He2 20–40 | 4720 ± 70 | SB-1 | 0.06 | Fen | Carex | 30–35 | 49.6 | 4.6 | 3.66 | 0.242 | 0.032 | 7.75 | 0.89 | 0.82 ± 0.08 | 0.60 ± 0.09 |
| Hef1⊥ 40–60 | 5270 ± 80 | AT-3 | 0.33 | Fen, frozen | Hypnaceous, Carex | 30–50 | 47.0 | 14.1 | 4.24 | 0.218 | 0.033 | 6.48 | 0.82 | 0.83 ± 0.08 | 0.66 ± 0.09 |
| Hef2⊥ 60–80 | n.d. | n.d. | n.d. | Fen, frozen | Subshrub, Carex | 35–40 | 23.1 | 54.3 | 5.05 | 0.237 | 0.035 | 7.47 | 0.87 | 0.93 ± 0.09 | 0.62 ± 0.09 |
| Hef3⊥ 80–100 | n.d. | n.d. | n.d. | Fen, frozen | Carex | 35 | 28.3 | 44.9 | 5.00 | 0.216 | 0.030 | 6.51 | 0.80 | 0.93 ± 0.09 | 0.38 ± 0.09 |
| Hef4⊥ 100–150 | n.d. | n.d. | n.d. | Fen, frozen | Betula sp., Picea, wood, Carex | 35–40 | 24.6 | 65.1 | 5.47 | 0.220 | 0.030 | 6.40 | 0.80 | 0.91 ± 0.09 | 0.39 ± 0.09 |
| Chfg1⊥ 150–175 | n.d. | n.d. | n.d. | Fen, frozen | Betula sp., Picea, wood, Carex | 40–45 | 11.7 | 76.9 | 5.32 | 0.225 | 0.034 | 6.33 | 0.77 | 0.92 ± 0.09 | 0.46 ± 0.09 |
| Chfg2⊥ 175–200 | n.d. | n.d. | n.d. | Fen, frozen | Equisetum, Betula sp., Picea, wood, Carex | >50 | 5.1 | 88.2 | 5.51 | 0.236 | 0.035 | 6.35 | 0.77 | 0.92 ± 0.09 | 0.48 ± 0.09 |
| Plot 2-1 | |||||||||||||||
| Hi 0–10 | n.d. | n.d. | n.d. | Raised | Hypnaceous | 20 | 46.0 | 2.6 | 3.72 | 0.159 | 0.023 | 6.72 | 0.83 | 1.19 ± 0.12 | 0.46 ± 0.12 |
| He 10–20 | 1820 ± 60 | SA-1 | 0.07 | Transitional | Hypnaceous | 35 | 48.8 | 8.7 | 3.84 | 0.208 | 0.027 | 6.93 | 0.84 | 1.04 ± 0.10 | 0.45 ± 0.11 |
| Hi1 20–30 | 3680 ± 70 | SB-2 | 0.04 | Transitional | Sphagnum | 20–25 | 50.1 | 9.2 | 3.84 | 0.239 | 0.030 | 6.75 | 0.82 | 0.99 ± 0.10 | 0.44 ± 0.10 |
| Hi2 30–33 | n.d. | n.d. | n.d. | Transitional | Sphagnum | 20 | 47.4 | 3.7 | 3.94 | 0.240 | 0.029 | 8.49 | 0.95 | 0.88 ± 0.09 | 0.48 ± 0.09 |
| Hif⊥ 33–50 | 4700 ± 80 | SB-1 | 0.15 | Transitional, frozen | Sphagnum | 25 | 40.6 | 9.4 | 4.60 | 0.251 | 0.033 | 7.00 | 0.84 | 0.89 ± 0.09 | 0.48 ± 0.09 |
| Hef1⊥ 50–70 | n.d. | n.d. | n.d. | Fen, frozen | Carex, Betula pubescens | 40 | 47.0 | 10.3 | 5.23 | 0.255 | 0.033 | 6.31 | 0.82 | 0.89 ± 0.09 | 0.47 ± 0.09 |
| Hef2⊥ 70–90 | 5620 ± 70 | AT-3 | 0.36 | Fen, frozen | Betula pubescens, Carex | 40 | 49.3 | 5.7 | 5.46 | 0.237 | 0.036 | 6.72 | 0.85 | 0.89 ± 0.09 | 0.46 ± 0.09 |
| Hef3⊥ 90–110 | n.d. | n.d. | n.d. | Fen, frozen | Betula pubescens, Carex, Menyanthes | 40 | 45.1 | 6.3 | 5.50 | 0.247 | 0.037 | 6.39 | 0.81 | 0.93 ± 0.09 | 0.48 ± 0.10 |
| Hif1⊥ 110–130 | 6590 ± 130 | AT-2 | 0.37 | Fen, frozen | Sphagnum | 25 | 48.5 | 6.3 | 5.61 | 0.233 | 0.033 | 6.82 | 0.88 | 0.90 ± 0.09 | 0.42 ± 0.09 |
| Hif2⊥ 130–150 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum, Hypnaceous | 25 | 50.6 | 4.1 | 5.63 | 0.209 | 0.029 | 6.53 | 0.91 | 0.95 ± 0.10 | 0.39 ± 0.10 |
| Hif3⊥ 150–170 | 6780 ± 130 | AT-2 | 2.55 | Fen, frozen | Sphagnum, Carex | 20 | 49.5 | 4.5 | 5.52 | 0.216 | 0.030 | 6.46 | 0.91 | 0.94 ± 0.09 | 0.40 ± 0.09 |
| Hif4⊥ 170–190 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum, grass | 20 | 50.0 | 3.9 | 5.51 | 0.200 | 0.027 | 6.24 | 0.89 | 0.93 ± 0.09 | 0.40 ± 0.09 |
| Hif5⊥ 190–210 | 7420 ± 120 | n.d. | 0.67 | Fen, frozen | Sphagnum, Carex | 20 | 49.1 | 4.2 | 5.51 | 0.216 | 0.028 | 6.54 | 0.91 | 0.93 ± 0.09 | 0.40 ± 0.09 |
| Hif6⊥ 210–230 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum | 20 | 49.6 | 4.5 | 5.54 | 0.217 | 0.030 | 6.40 | 0.89 | 0.92 ± 0.09 | 0.40 ± 0.09 |
| Hif7⊥ 230–250 | 7460 ± 130 | AT-1 | n.d. | Fen, frozen | Hypnaceous, Menyanthes | 25 | 52.5 | 4.0 | 5.73 | 0.216 | 0.030 | 6.52 | 0.90 | 0.96 ± 0.10 | 0.37 ± 0.10 |
| Hif8⊥ 250–265 | n.d. | n.d. | n.d. | Fen, frozen | Menyanthes, Carex | 25–30 | 42.6 | 22.3 | 5.31 | 0.258 | 0.041 | 6.55 | 0.84 | 0.87 ± 0.09 | 0.43 ± 0.09 |
| Cfg⊥ 265–285 | 7870 ± 90 | AT-1 | 0.74 | - | - | n.d. | 0.7 | 98.1 | 5.55 | 0.302 | 0.050 | 6.28 | 0.76 | 0.92 ± 0.09 | 0.43 ± 0.09 |
| Plot 2-2 | |||||||||||||||
| Ha1 0–5 | 2070 ± 70 | SA-1 | n.d. | Transitional | Hypnaceous, Menyanthes, | >50 | 47.6 | 8.4 | 3.70 | 0.213 | 0.029 | 7.95 | 0.90 | 1.01 ± 0.10 | 0.48 ± 0.10 |
| Ha2 5–10 | n.d. | n.d. | n.d. | Transitional | Hypnaceous | >50 | 47.5 | 7.7 | 3.59 | 0.216 | 0.029 | 8.33 | 0.93 | 0.99 ± 0.09 | 0.46 ± 0.10 |
| He1 10–20 | n.d. | n.d. | n.d. | Transitional | Shrub, Eriophorum | 40 | 48.3 | 7.2 | 3.63 | 0.203 | 0.028 | 8.14 | 0.92 | 1.06 ± 0.10 | 0.46 ± 0.11 |
| He2 20–32 | n.d. | n.d. | n.d. | Transitional | Shrub, Eriophorum | 35–40 | 48.1 | 7.2 | 3.68 | 0.214 | 0.029 | 7.27 | 0.88 | 0.99 ± 0.09 | 0.45 ± 0.10 |
| He3 32–40 | n.d. | n.d. | n.d. | Fen | Shrub, Eriophorum, Carex | 35–40 | 49.1 | 5.7 | 3.80 | 0.255 | 0.036 | 8.20 | 0.91 | 0.84 ± 0.08 | 0.50 ± 0.09 |
| Hif⊥ 40–50 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum, Carex | 30 | 51.4 | 4.7 | 4.32 | 0.246 | 0.037 | 6.24 | 0.79 | 0.82 ± 0.08 | 0.47 ± 0.09 |
| Hef1⊥ 50–70 | n.d. | n.d. | n.d. | Fen, frozen | Carex, Menyanthes | 35 | 49.0 | 7.2 | 5.26 | 0.230 | 0.034 | 6.82 | 0.83 | 0.86 ± 0.09 | 0.46 ± 0.09 |
| Hef2⊥ 70–90 | n.d. | n.d. | n.d. | Fen, frozen | Hypnaceous, Carex | 35 | 50.5 | 5.7 | 5.35 | 0.240 | 0.036 | 6.39 | 0.83 | 0.79 ± 0.08 | 0.46 ± 0.08 |
| Hef3⊥ 90–110 | n.d. | n.d. | n.d. | Fen, frozen | Carex, Pinus sylvestris, Betula nana | 35–40 | 48.8 | 12.4 | 5.40 | 0.255 | 0.039 | 6.38 | 0.78 | 0.76 ± 0.08 | 0.50 ± 0.08 |
| Hif1⊥ 110–130 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum, Carex | 20–25 | 46.8 | 5.4 | 5.36 | 0.227 | 0.030 | 6.85 | 0.90 | 0.88 ± 0.09 | 0.41 ± 0.09 |
| Hif2⊥ 130–150 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum, Carex, wood | 15–20 | 49.0 | 4.5 | 5.38 | 0.210 | 0.027 | 6.57 | 0.91 | 0.89 ± 0.09 | 0.39 ± 0.09 |
| Hif3⊥ 150–170 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum, grass | 20 | 49.4 | 3.9 | 5.39 | 0.210 | 0.026 | 6.55 | 0.91 | 0.95 ± 0.10 | 0.38 ± 0.10 |
| Hif4⊥ 170–190 | n.d. | n.d. | n.d. | Fen, frozen | Sphagnum, grass | 25 | 48.0 | 8.7 | 5.17 | 0.243 | 0.033 | 6.37 | 0.86 | 0.96 ± 0.10 | 0.40 ± 0.10 |
| Hef⊥190–210 | n.d. | n.d. | n.d. | Fen, frozen | Equisetum, Pinus sylvestris | 35–40 | 40.1 | 24.7 | 5.09 | 0.261 | 0.042 | 6.00 | 0.76 | 0.76 ± 0.08 | 0.49 ± 0.08 |
| Chfg⊥ 210–230 | n.d. | n.d. | n.d. | Fen, frozen | Carex | n.d. | 1.5 | 96.6 | 5.31 | 0.251 | 0.040 | 6.33 | 0.74 | 0.79 ± 0.08 | 0.49 ± 0.08 |
| Cfg⊥ 230–240 | n.d. | n.d. | n.d. | - | - | n.d. | 0.6 | 98.1 | 5.50 | n.d. | n.d. | n.d. | n.d. | 0.92 ± 0.09 | 0.38 ± 0.09 |
| Horizon/Depth, cm | HMF | MMF | LMF | Mw/Mn | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mr 1, kDa | SD2 | Molar Fraction | Mr, kDa | SD | Molar Fraction | Mr, kDa | SD | Molar Fraction | |||||
| x | SD | x | SD | x | SD | ||||||||
| Plot 1-1 | |||||||||||||
| Hi 0–10 | 418 | 3 | 3.65 | 0.17 | 20.2 | 0.3 | 28.2 | 0.26 | 1.19 | 0.03 | 68.1 | 0.4 | 13.7 |
| He1 10–20 | 408 | 4 | 3.0 | 0.3 | 19.7 | 0.6 | 27.2 | 0.4 | 1.19 | 0.03 | 69.8 | 0.7 | 15.0 |
| He2 20–40 | 403.6 | 1.0 | 1.82 | 0.14 | 17.6 | 0.5 | 23.07 | 0.16 | 1.10 | 0.03 | 75.1 | 0.3 | 20.3 |
| Hef1⊥ 40–60 | 389 | 5 | 1.21 | 0.06 | 16.32 | 0.03 | 22.77 | 0.18 | 1.18 | 0.03 | 76.02 | 0.24 | 21.8 |
| Hef2⊥ 60–80 | 391.1 | 2.9 | 0.83 | 0.03 | 15.52 | 0.04 | 21.39 | 0.08 | 1.26 | 0.03 | 77.78 | 0.08 | 23.2 |
| Hef3⊥ 80–100 | 367.9 | 0.3 | 0.96 | 0.04 | 16.04 | 0.24 | 19.20 | 0.11 | 1.05 | 0.03 | 79.88 | 0.15 | 24.4 |
| Hef4⊥ 100–150 | 370 | 4 | 1.44 | 0.18 | 18.0 | 0.4 | 20 | 7 | 1.18 | 0.04 | 79 | 7 | 21.1 |
| Hef5⊥ 150–175 | 372.5 | 0.9 | 1.69 | 0.06 | 18.73 | 0.14 | 28.58 | 0.28 | 1.31 | 0.03 | 69.7 | 0.3 | 15.5 |
| Chfg⊥ 175–200 | 376 | 8 | 2.28 | 0.05 | 20.2 | 0.7 | 32.9 | 0.9 | 1.45 | 0.14 | 64.8 | 1.0 | 12.9 |
| Plot 1-2 | |||||||||||||
| Hi 0–5 | 440 | 15 | 3.80 | 0.11 | 20.4 | 0.5 | 27.46 | 0.22 | 1.20 | 0.06 | 68.75 | 0.11 | 14.0 |
| He1 5–20 | 400 | 4 | 3.60 | 0.21 | 22.0 | 0.3 | 30.01 | 0.22 | 1.27 | 0.03 | 66.4 | 0.4 | 12.4 |
| He2 20–40 | 386 | 4 | 2.7 | 0.2 | 20.3 | 0.6 | 29.05 | 0.10 | 1.27 | 0.04 | 68.2 | 0.3 | 14.0 |
| Hef1⊥ 40–60 | 467.9 | 2.3 | 0.75 | 0.03 | 14.55 | 0.21 | 16.70 | 0.06 | 1.20 | 0.04 | 82.56 | 0.10 | 35.0 |
| Hef2⊥ 60–80 | 379.0 | 2.8 | 1.45 | 0.08 | 17.68 | 0.26 | 25.50 | 0.20 | 1.27 | 0.03 | 73.04 | 0.12 | 18.1 |
| Hef3⊥ 80–100 | 383.7 | 3 | 1.28 | 0.13 | 17.4 | 0.5 | 23.8 | 0.4 | 1.18 | 0.03 | 74.9 | 0.6 | 19.8 |
| Hef4⊥ 100–150 | 390.4 | 1.9 | 1.67 | 0.09 | 17.5 | 0.3 | 25.00 | 0.17 | 1.25 | 0.04 | 73.33 | 0.08 | 18.8 |
| Chfg1⊥ 150–175 | 390 | 5 | 2.90 | 0.18 | 20.3 | 0.8 | 32.6 | 0.4 | 1.57 | 0.09 | 64.51 | 0.20 | 12.7 |
| Chfg2⊥ 175–200 | 364 | 4 | 1.02 | 0.03 | 17.2 | 0.5 | 25.05 | 0.03 | 1.24 | 0.08 | 73.93 | 0.04 | 17.9 |
| Plot 2-1 | |||||||||||||
| Hi 0–10 | 333.7 | 0.3 | 6.17 | 0.10 | 24.9 | 0.9 | 30.45 | 0.20 | 1.41 | 0.03 | 63.4 | 0.3 | 8.3 |
| He 10–20 | 307.5 | 0.7 | 2.9 | 0.3 | 20.5 | 0.4 | 27.92 | 0.23 | 1.46 | 0.03 | 69.2 | 0.6 | 11.7 |
| Hi1 20–30 | 309 | 4 | 2.4 | 0.4 | 20.0 | 0.3 | 27.6 | 0.7 | 1.59 | 0.04 | 69.9 | 1.0 | 12.1 |
| Hi2 30–33 | 302.4 | 0.7 | 3.96 | 0.14 | 23.15 | 0.06 | 28.98 | 0.06 | 1.45 | 0.03 | 67.06 | 0.09 | 9.8 |
| Hif⊥ 33–50 | 373 | 4 | 3.61 | 0.29 | 21.7 | 0.4 | 28.33 | 0.03 | 1.62 | 0.03 | 68.07 | 0.28 | 12 |
| Hef1⊥ 50–70 | 370 | 5 | 3.4 | 0.3 | 21.62 | 0.10 | 27.62 | 0.28 | 1.64 | 0.03 | 68.98 | 0.17 | 12.3 |
| Hef2⊥ 70–90 | 302.9 | 1.4 | 1.93 | 0.07 | 18.8 | 0.4 | 19.97 | 0.06 | 1.31 | 0.03 | 78.10 | 0.12 | 16.3 |
| Hef3⊥ 90–110 | 296.4 | 2.1 | 3.72 | 0.05 | 22.36 | 0.11 | 29.81 | 0.03 | 1.49 | 0.03 | 66.47 | 0.03 | 9.8 |
| Hif1⊥ 110–130 | 304.9 | 2.9 | 1.42 | 0.08 | 17.04 | 0.28 | 16.26 | 0.07 | 1.11 | 0.04 | 82.31 | 0.15 | 21.3 |
| Hif2⊥ 130–150 | 307 | 5 | 1.43 | 0.13 | 16.51 | 0.16 | 18.19 | 0.03 | 1.10 | 0.03 | 80.38 | 0.03 | 24.7 |
| Hif3⊥ 150–170 | 312 | 3 | 1.16 | 0.13 | 16.34 | 0.08 | 16.80 | 0.03 | 1.05 | 0.03 | 82.04 | 0.12 | 22.5 |
| Hif4⊥ 170–190 | 309 | 7 | 0.96 | 0.15 | 15.34 | 0.29 | 15.48 | 0.17 | 1.01 | 0.03 | 84.1 | 0.4 | 31.7 |
| Hif5⊥ 190–210 | 305 | 3 | 1.06 | 0.06 | 15.86 | 0.15 | 16.6 | 0.4 | 1.01 | 0.03 | 82.4 | 0.4 | 22.9 |
| Hif6⊥ 210–230 | 309 | 8 | 1.83 | 0.12 | 17.41 | 0.16 | 20.7 | 0.5 | 1.09 | 0.05 | 77.5 | 0.6 | 17.7 |
| Hif7⊥ 230–250 | 312 | 5 | 1.23 | 0.20 | 15.42 | 0.18 | 17.6 | 0.3 | 1.11 | 0.03 | 81.2 | 0.5 | 22.3 |
| Hif8⊥ 250–265 | 297 | 4 | 0.89 | 0.08 | 15.89 | 0.26 | 13.83 | 0.08 | 1.16 | 0.03 | 85.28 | 0.16 | 24.2 |
| Plot 2-2 | |||||||||||||
| Ha1 0–5 | 306.5 | 0.7 | 2.47 | 0.04 | 19.96 | 0.09 | 25.59 | 0.09 | 1.30 | 0.03 | 71.94 | 0.07 | 13.1 |
| Ha2 5–10 | 306.9 | 0.3 | 2.89 | 0.09 | 20.2 | 0.4 | 28.8 | 0.4 | 1.46 | 0.03 | 68.3 | 0.5 | 11.6 |
| He1 10–20 | 303.9 | 0.3 | 2.93 | 0.03 | 20.53 | 0.05 | 28.54 | 0.14 | 1.45 | 0.03 | 68.53 | 0.14 | 11.4 |
| He2 20–32 | 294.9 | 0.3 | 2.55 | 0.03 | 20.16 | 0.03 | 27.43 | 0.08 | 1.49 | 0.04 | 70.02 | 0.08 | 11.7 |
| He3 32–40 | 278.7 | 0.4 | 2.08 | 0.06 | 20.03 | 0.03 | 28.86 | 0.18 | 1.62 | 0.03 | 69.05 | 0.24 | 10.7 |
| Hif⊥ 40–50 | 281 | 3 | 2.25 | 0.27 | 20.2 | 0.4 | 26.85 | 0.19 | 1.49 | 0.03 | 70.9 | 0.5 | 11.5 |
| Hef1⊥ 50–70 | 296 | 6 | 2.7 | 0.19 | 20.19 | 0.05 | 25.9 | 0.3 | 1.43 | 0.03 | 71.4 | 0.5 | 12.2 |
| Hef2⊥ 70–90 | 292 | 6 | 1.98 | 0.21 | 18.9 | 0.4 | 22.85 | 0.11 | 1.39 | 0.03 | 75.2 | 0.3 | 14.2 |
| Hef3⊥ 90–110 | 294.4 | 0.7 | 3.28 | 0.21 | 21.5 | 0.5 | 27.64 | 0.05 | 1.55 | 0.04 | 69.08 | 0.26 | 10.7 |
| Hif1⊥ 110–130 | 304 | 9 | 1.55 | 0.10 | 17.44 | 0.04 | 19.05 | 0.24 | 1.18 | 0.03 | 79.40 | 0.14 | 18.5 |
| Hif2⊥ 130–150 | 294.9 | 1.4 | 1.62 | 0.06 | 17.64 | 0.17 | 22.61 | 0.17 | 1.18 | 0.03 | 75.8 | 0.3 | 15.8 |
| Hif3⊥ 150–170 | 303.4 | 0.7 | 1.44 | 0.03 | 18.77 | 0.13 | 21.58 | 0.07 | 1.16 | 0.03 | 83.7 | 0.5 | 15.9 |
| Hif4⊥ 170–190 | 308 | 6 | 1.38 | 0.13 | 16.4 | 0.15 | 17.9 | 0.8 | 1.24 | 0.04 | 80.8 | 0.9 | 20.3 |
| Hef⊥ 190–210 | 294.9 | 1.4 | 4.19 | 0.11 | 22.2 | 0.3 | 30.72 | 0.10 | 1.69 | 0.04 | 65.09 | 0.20 | 9.2 |
| Chfg⊥ 210–230 | 271.4 | 2.6 | 4 | 0.5 | 23.9 | 0.6 | 37.65 | 0.26 | 1.99 | 0.03 | 58.3 | 0.8 | 7.1 |
| Principal Components | Eigenvalues | % of Total Variance | Cumulative Eigenvalues | Cumulative % of Variance |
|---|---|---|---|---|
| PC 1 | 6.90 | 45.98 | 6.90 | 45.98 |
| PC 2 | 2.43 | 16.23 | 9.33 | 62.21 |
| PC 3 | 1.86 | 12.40 | 11.19 | 74.61 |
| PC 4 | 1.39 | 9.28 | 12.58 | 83.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasilevich, R.; Lodygin, E.; Abakumov, E. The Molecular Composition of Humic Acids in Permafrost Peats in the European Arctic as Paleorecord of the Environmental Conditions of the Holocene. Agronomy 2022, 12, 2053. https://doi.org/10.3390/agronomy12092053
Vasilevich R, Lodygin E, Abakumov E. The Molecular Composition of Humic Acids in Permafrost Peats in the European Arctic as Paleorecord of the Environmental Conditions of the Holocene. Agronomy. 2022; 12(9):2053. https://doi.org/10.3390/agronomy12092053
Chicago/Turabian StyleVasilevich, Roman, Evgeny Lodygin, and Evgeny Abakumov. 2022. "The Molecular Composition of Humic Acids in Permafrost Peats in the European Arctic as Paleorecord of the Environmental Conditions of the Holocene" Agronomy 12, no. 9: 2053. https://doi.org/10.3390/agronomy12092053
APA StyleVasilevich, R., Lodygin, E., & Abakumov, E. (2022). The Molecular Composition of Humic Acids in Permafrost Peats in the European Arctic as Paleorecord of the Environmental Conditions of the Holocene. Agronomy, 12(9), 2053. https://doi.org/10.3390/agronomy12092053