Abstract
Saponarin content in barley sprouts may vary greatly with environmental conditions, such as climate, leading to difficulty in uniformly producing saponarin-rich barley sprouts in situ farmlands throughout the year. This research was an early attempt to identify the optimal conditions of various climatic factors, such as temperature, light, and humidity according to seasonal change, for maximizing the saponarin content of sprouted barley through the two-year field experiment. As a result, the growth index, as leaf length relative to growth period, of barley sprouts varied greatly with sowing time, and they tended to decrease with an increase in the ambient temperature, such as average daily temperature. In contrast, higher saponarin contents were observed in the sprouts collected in March, April, September, and October than those collected from May to August. We also found significantly positive correlations of saponarin content with daily temperature range and average light period, indicating that they could be decisive climatic factors for the production of barley sprouts with a higher saponarin content. Interestingly, the polynomial relationship between saponarin yield and leaf length showed the highest yield with 2.18 mg plant−1 at 15.9 cm in length, suggesting a best cutting time for the production of saponarin-rich barely sprouts based on the leaf length. Overall, the decisive climatic factors according to seasonal change for saponarin biosynthesis may be considered to be daily temperature differences and light hours.
1. Introduction
Globally, there has been growing interest from the food and medicinal industries in the past few decades in commercializing natural substrates, i.e., secondary metabolites, derived from cereals, vegetables, and fruits, because of their nutritional value against chronic diseases [1,2,3], along with the desire to prolong life. In this context, currently, crop sprouts, often named miracle food, have become popular to grow to produce food supplements possessing high levels of minerals, proteins, and vitamins, as well as health-supporting substances, including glucosinolates, phenolics, and Se-containing compounds [4,5]. With an uptrend in the consumption of healthy foods, sprout vegetables have increased enough to occupy over 30% of the total vegetable market in the United States, Europe, and Australia [6,7].
In general, various metabolic pathways are activated during the early stages of crop growth, thereby generating beneficial substances in sprouts. Firstly, during germination, the metabolic enzymes in seeds activate or generate biochemical alterations, such as the conversion of fats to free fatty acids, the degradation of polysaccharides, and the reduction of proteins to oligopeptides and free amino acids [5,6,7,8]. Hereafter, various nutritional and functional compounds in sprout substrates are generated a few days after germination, namely during sprouting, which can be considered as an excellent source for healthy foods, including cereals, legumes, vegetables, and oilseeds [9]. Currently, barley sprouts (Hordeum vulgare L. Poaceae family) have been widely used as health substances and dietary supplements because of their biological and pharmacological components, including flavonoids, saponarin, γ-aminobutyric acid (GABA), policosanols, and lutonarin [10,11]. Primarily, such compounds are renowned for their potential health effects in humans, including in treating diabetes and hypertension, and in antioxidant and antitumor capacities [12,13]. Based on these benefits, several studies are focusing on the product development and quality improvement of sprout crops, especially in South Korea [11,14].
Saponarin of the diglycoside flavone structure (apigenin-6-C-glucosyl-7-O-glucoside) is the significant phenolic compound (approximately 72%) in barley sprouts [15,16]. Although several studies documented that the saponarin effect on human health exhibits significant benefits in the prevention of lifestyle-related diseases, such as inflammation, hypocholesterolemia, and carcinogenesis [10,11,16,17], there are no reports of experimental data comparing the difference in the secondary metabolite in barley sprouts under various growing conditions. Given the widespread fact that the contents, profiles, and biosynthesis of health-beneficial compounds in crops vary with cropping system, growth conditions (e.g., availability of water and nutrients), climatic factors (e.g., temperature and radiation), and postharvest handling conditions [14,18], it is likely that saponarin content in sprouted barley would show similar trends. As such, the field environmental condition is expected to be a major contributor for saponarin production. Through our earlier study using a hydroponic system, we found that saponarin content in barley sprouts was dependent on water stress, but not on nutrient levels [19]. Meanwhile, there have been no studies about the influence of climatic conditions on saponarin production in barley sprouts, especially in situ arable lands, so far.
In our continued screening of bioactive phenolics in diverse crops, we found that the saponarin contents of 60 commercial foods using barley sprouts exhibited remarkable differences in the range of 0–1000 mg 100 g−1. This seems to indicate that the optimal growth conditions (i.e., climate) and cultivation methods of barley plants are critical to obtaining high phenolic contents and strong biological activities concerning the development of functional foods. Thus, such external factors may influence saponarin production significantly, which would lead to difficulty in uniformly producing saponarin-rich barley sprouts. The current work was designed to investigate critical climatic factors in field conditions affecting saponarin production in barley sprouts by seasonal sowing times. Hence, this field trial would document for the first time the optimal conditions for saponarin synthesis depending on climate factors and sowing times.
2. Materials and Methods
2.1. Plant Material and Cultivation
Seeds of barley (Hordeum vulgare L.) cultivar ‘Keunalbori 1’, which contains higher saponarin content [20], were obtained from the Rural Development Administration (RDA), Republic of Korea, for this study. This field work was performed for two years at the experimental farmland at Gyeongsang National University, Jinju, Korea (35°08′58.1″ N 128°05′52.3″ E). The field (approximately 1920 m2) was installed with a total of three plots (10 m × 60 m) at intervals of one meter. The upland had been managed with no fertilization during the two-year field trials, as it was proved through earlier research that the production of sprouted barley containing high saponarin was not related to the nutrient condition [19]. Primary tillage was conducted a week before the first sowing in March, and the upland was plowed immediately after harvest every month in order to turn over the sprout residues and control weeds. The seeds were scattered into the upland for sowing as 250 kg ha−1, and their sprouts were harvested every month as follows: March to October in 2019 (first year), and March, May, September, and October in 2020 (second year). The reasons for excluding the sprout cultivation in April and June to August in the second-year experiment are as follows: (1) difficulty in land management due to heavy rainfall, (2) low production of saponarin as a result of the first-year experiment over this season, and (3) multiple cropping, which is commonly conducted at this season to increase crop production and income as well as improve overall soil health. In addition, information on the climatic factors over the experimental duration was collected from the website of the Korea Meteorological Administration, and the climate data (Table S1) are in the Supplementary Materials, which includes average daily temperature (ADT), accumulated temperature (ACT), daily temperature range (DTR), average light period (ALP), average daily photons (ADP), accumulated photons (ACP), relative humidity (RH), and rainfall.
2.2. Soil and Plant Sampling
Surface soil (0–20 cm) of three plots was collected twice from barley grown upland at the beginning of each year’s field experiment to analyze the soil properties. Three subsamples were collected from each plot, and they were mixed properly prior to soil analysis. For the collection of barley sprouts, a study of primary leaves grown only erect under high plant density was conducted on the basis of their lengths (approximately 10, 15, and 18 cm) from leaf sheath to apex. For plant sampling, the sprout length was selected based on the 15 cm length recommended by the RDA [19]. In each plot, the sprouts were randomly sub-sampled from a specific area of 50 cm × 50 cm, which was quintuplicated. After the removal of dead and yellow leaves, the leaf samples were gently washed with deionized water prior to plant analysis. The growth period (GP) of each sowing time’s sample was recorded, and the growth index (GI) was calculated as follows: GI = leaf length/GP. The samples were immediately freeze-dried at −50 °C for five days using a vacuum freeze dryer (HyperCOOL HC3110, Hanil Scientific Inc., Gimpo, Korea), before being pulverized to 100 mesh and then stored at −20 °C until saponarin analysis.
2.3. Analytical
The collected soil was air-dried and sieved (<2 mm) prior to analysis of the properties. The pH and electrical conductivity (EC) of the soil were measured in a suspension of soil–distilled water (1:5 = w/v) using a pH meter (Orion Star 2-Star, Thermo Scientific, Waltham, MA, USA) and an EC meter (Orion 3-Star, Thermo Scientific, Waltham, MA, USA). Soil organic matter (OM) was determined by the Tyurin method [21], and total nitrogen (T-N) was analyzed by the Kjeldahl method [22]. Lancaster’s method [23] was used to determine plant available phosphorous (Av. P2O5). After extraction with 1 N NH4OAc at pH 7.0 [23], cation exchange capacity (CEC) was determined using the Kjeldahl method (K-355, BÜCHI, Flawil, Switzerland), and exchangeable potassium (K), calcium (Ca), and magnesium (Mg) were determined using ICP–OES (OPTIMA 5300DV, PerkinElmer, Waltham, MA, USA).
The fresh weight (FW) and dried weight (DW) of each sample were recorded. Saponarin content in barley sprouts was determined according to the method of Seo et al. (2014). Here, 1 g of the homogenized sample was added to 20 mL of 50% ethanol (v/v) and incubated at 35 °C for one day. After centrifugation at 7800 rpm for 5 min, the supernatant was filtered through a 0.2 μm polytetrafluoroethylene (PTFE) syringe filter (6784–1302, Whatman, Maidstone, UK). The saponarin content in this extract was measured using ultra-high-performance liquid chromatography (UHPLC) with an UV detector (Dionex Ultimate 3000, Thermo Scientific, Waltham, MA, USA). Separation was achieved using a Zorbax Eclipse XDB-C18 column (4.6 mm × 150 mm, 5 μm) at 35 °C. The mobile phase was programmed as 0.1% trifluoro acetic acid (TFA) in water (solvent A) and acetonitrile (solvent B) with a flow rate of 0.5 mL min−1 as follows: 0–3 min, 3% B; 3–10 min, 15% B; 10–13 min, 30% B; 13–15 min, 50% B; 15–16 min, 90% B; 16–18 min, 90% B; 18–20 min, 100 B, and then maintained at 100% B for 5 min. The injection volume was 10 µL, and the detection wavelength was 325 nm.
The saponarin yield was estimated by calculating the total content in each barley sprout as follows: saponarin content (mg 100 g−1) × DW (g plant−1).
2.4. Statistical Analysis
Values of soil and plant parameters were analyzed as the mean (±standard deviations) of triple measurements. The overall effects of sowing time, leaf length, and interaction with plant FW were assessed using two-way ANOVA. To compare the differences in plant parameters, such as GP, GI, and saponarin content among different cultivation times, a one-way ANOVA with Tukey’s honestly significant difference (HSD) test at the 0.05 probability level was conducted (n = 3). Pearson correlation analysis was used to measure how saponarin content is related to FW and environmental factors. Principal component analysis (PCA) was performed to investigate patterns of variation in the dataset, focusing on the degree of relationship between saponarin content and climatic factors. All statistical analyses were conducted using XLStat-Base software (Addinsoft, New York, NY, USA) in an Excel program.
3. Results
3.1. Property of Field Soil
The physicochemical properties of farmland soil are shown in Table 1. The levels of most soil parameters were within the appropriate soil nutrient range for barley cultivation based on the Korean RDA recommendation (http://soil.rda.go.kr; accessed on 1 March 2019), but the Mg solubility exhibited a slightly low value (<2.0 cmolc kg−1).

Table 1.
Initial soil properties of this experiment in 2019 and 2020 year.
3.2. Growth of Barley Sprouts
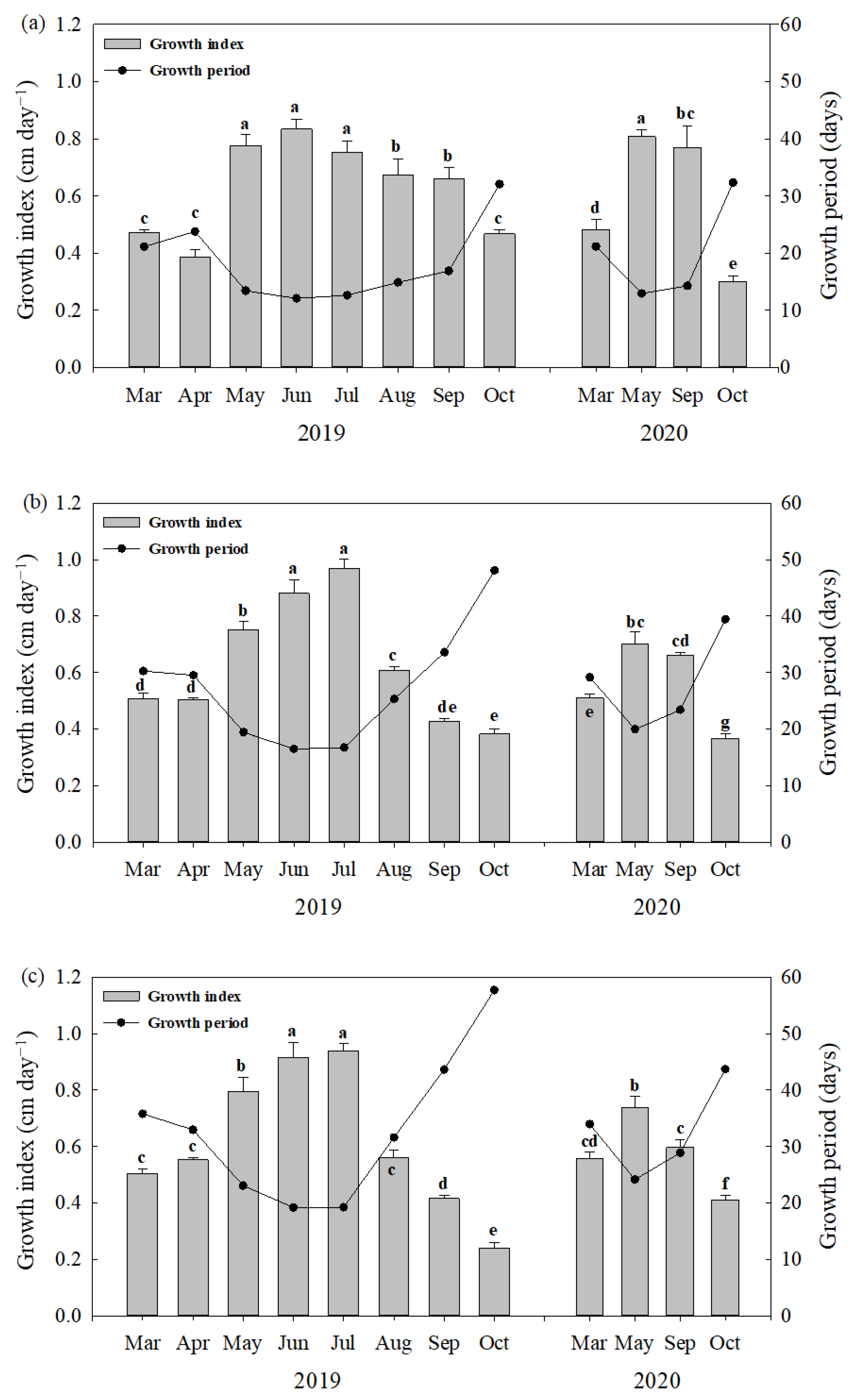
The average FWs of barley sprouts sampled in 2019 were 0.094, 0.154, and 0.201 g for 10, 15, and 18 cm lengths, respectively (Table 2). The FWs of sprouts among different leaf lengths were significantly different (p < 0.05), but there were no significant differences in the FWs according to the sowing time (p > 0.05) (Table 2). Such tendencies in the FWs and GPs of sprouts were observed in 2020. On the other hand, the GPs of sprouts until harvesting differed greatly depending on the sowing time (Figure 1). In the first year, the GP in October was 48 days, while the GPs between May and July were less than 20 days. After that, the GP tended to increase from August to October. A similar tendency in GPs depending on the sowing time was observed in the second year.
Table 2.
Fresh weights (FWs, g) of 10, 15, and 18 cm barley sprouts grown in different months and years.
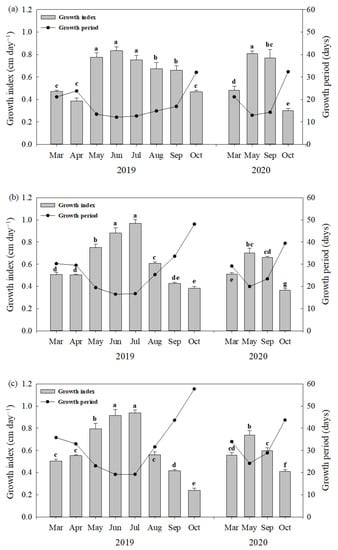
Figure 1.
Growth period (GP) and growth index (GI) estimated as a ratio of leaf length to GP of 10 cm (a), 15 cm and (b), and 18 cm; (c) lengths of barley sprouts according to the sowing time in 2019 and 2020. Data of bar graph represent mean ± standard deviation (n = 3) and same letters indicate no significant difference in the GI values across the two-year experiment (Tukey’s HSD, p < 0.05).
The estimated growth index of barley sprouts varied with the sowing time, but its change pattern was similar for all lengths of the sprouts sampled (Figure 1). For 15 cm barley sprouts sampled in 2019 (Figure 1b), the highest value of GI was found in July (0.96 cm day−1), followed by June (0.88 cm day−1), May (0.75 cm day−1), August (0.60 cm day−1), and so on. Meanwhile, the lowest GI was observed in October (0.36 cm day−1). In 2020, the highest GI value was found in May (0.70 cm day−1), followed by September (0.66 cm day−1), March (0.51 cm day−1), and October (0.36 cm day−1).
3.3. Determination of Saponarin Content in Barley Sprouts
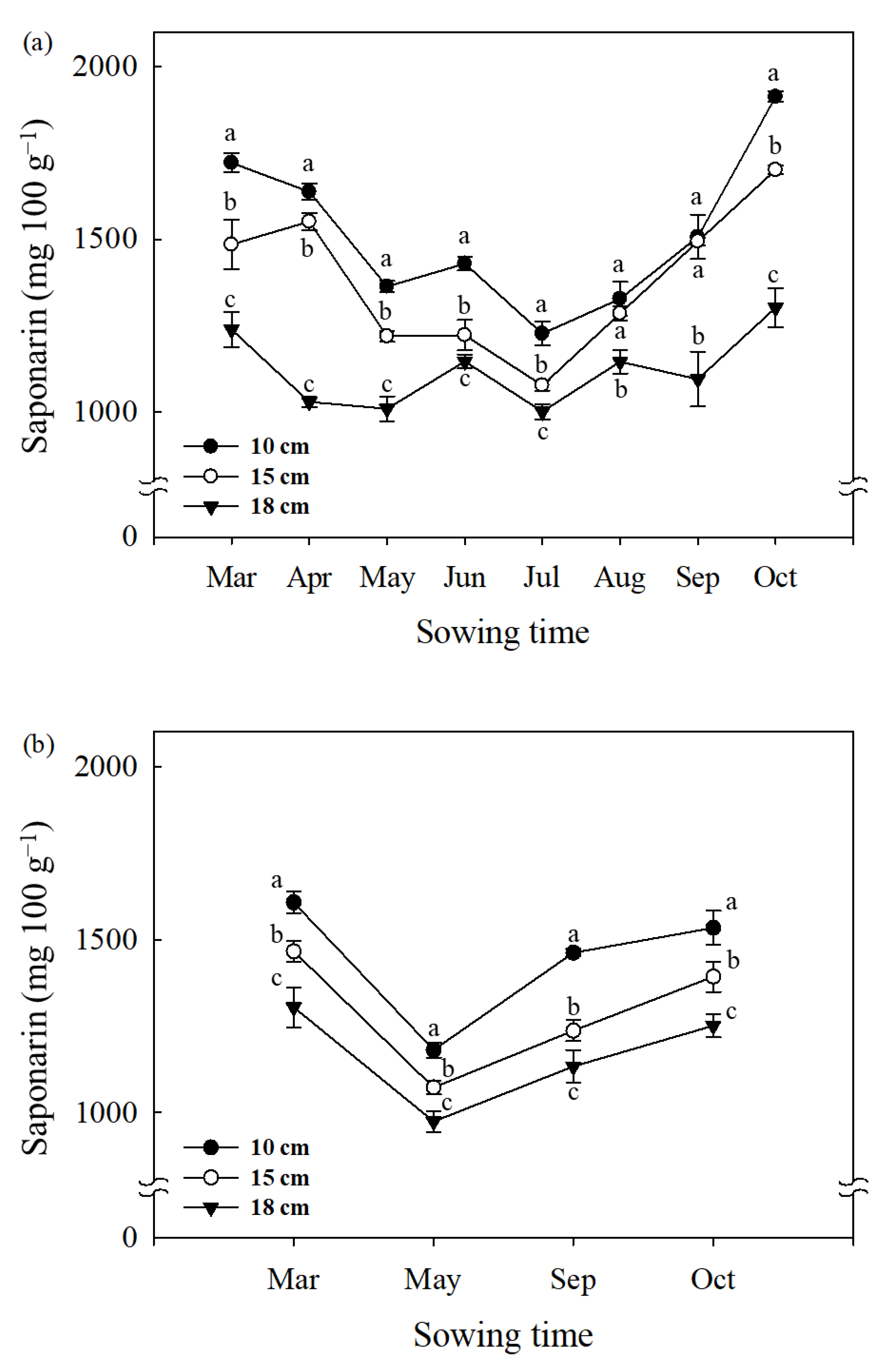
The saponarin content of barley sprouts varied significantly with the sowing time (Figure 2). The saponarin content presented a trend of decrease from March to July and increased later from August to October (Figure 2a). The highest saponarin contents were found in sprout samples grown in October (1913 mg 100 g−1 for 10 cm, 1701 mg 100 g−1 for 15 cm, and 1302 mg 100 g−1 for 18 cm), whereas the lowest contents were observed in July (Figure 2a). In the second year of the experiment, the change in saponarin content of barley sprouts depending on the sowing time appears to be similar. The highest values were found in barley sprouts (10, 15, and 18 cm) sown in March (1608, 1466, 1304 mg 100 g−1, respectively), followed by October (1534, 1393, 1251 mg 100 g−1, respectively), September (1462, 1236, 1133 mg 100 g−1, respectively), and May (1181, 1072, 974 mg 100 g−1, respectively).
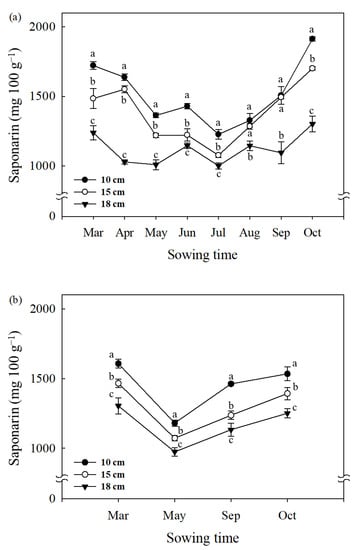
Figure 2.
Change in saponarin content of barley sprouts (10, 15, and 18 cm length) grown at different sowing times in 2019 (a) and 2020 (b). Data represent mean ± standard deviation (n = 3) and same letters in each sowing month indicate no significant difference in the saponarin content (Tukey’s HSD, p < 0.05).
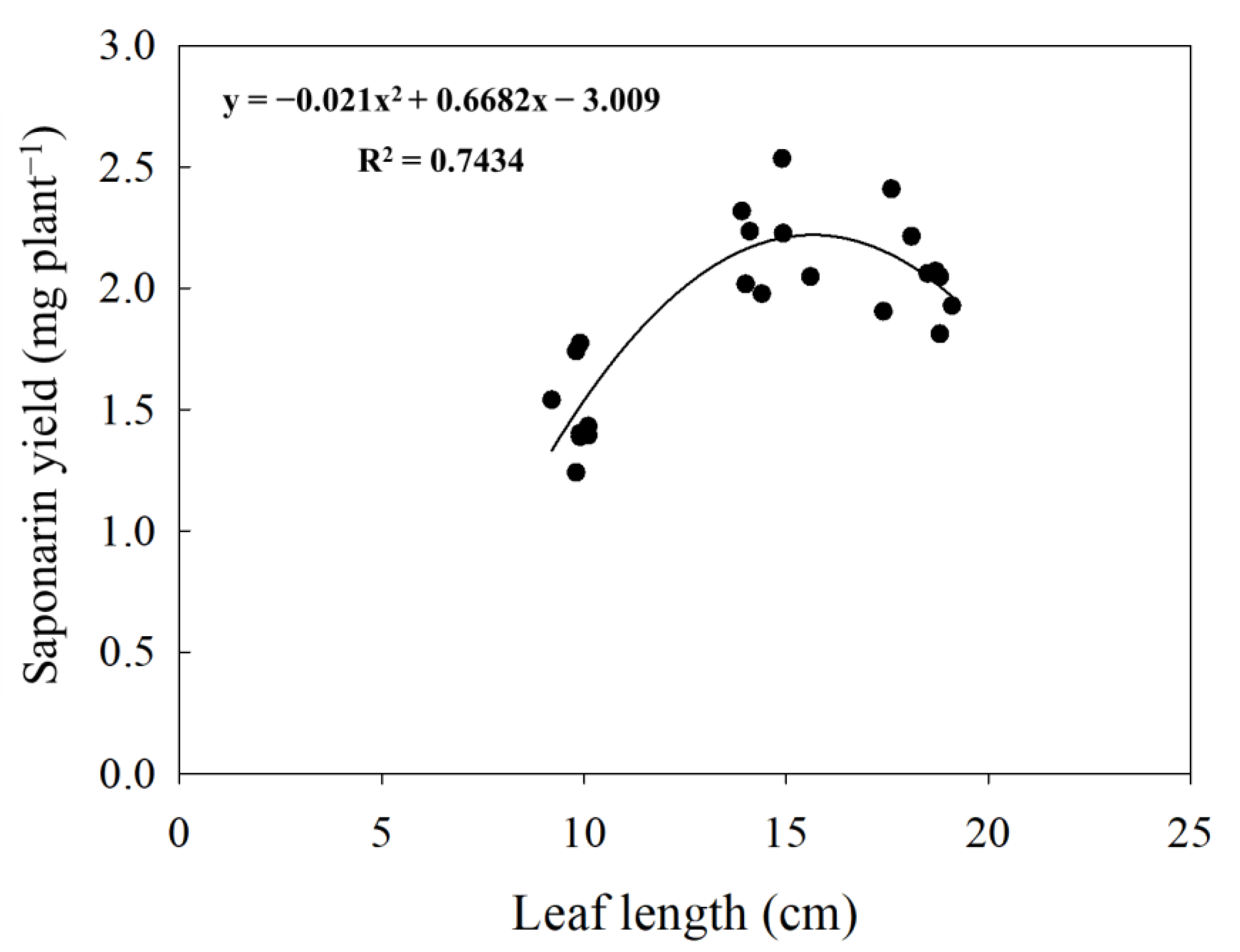
In general, it is clear that the saponarin content of barley sprouts tended to decrease with the increase in primary leaf length sampled in all two years (10 cm > 15 cm > 18 cm). On the other hand, the total saponarin yield per plant showed a different tendency depending on the leaf length (Table 3). The most significant yield of saponarin was found in the leaf samples of 15 cm length (2.2 mg plant−1), followed by 18 cm (2.0 mg plant−1), and 10 cm (1.4 mg plant−1). We also found a significant polynomial relationship between the yield of saponarin (mg DW plant−1) and the length of leaves (r2 = 0.85, p < 0.001) (Figure 3).
Table 3.
Total saponarin yield in barley sprouts through different leaf lengths (10, 15, and 18 cm).
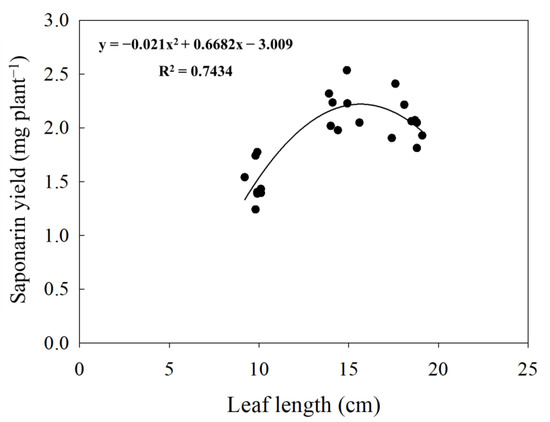
Figure 3.
Polynomial relationship between sprout length and saponarin yield (n = 36). Black dots indicate the saponarin yield of barley sprouts cultivated with different sowing times in 2019 and 2020. The peak value of saponarin yield in this study was discovered from the sprouts of 15.9 cm length.
3.4. Relationship between Saponarin Contents and Environmental Parameters
Table 4 shows significant correlations of saponarin contents with diverse parameters of plant growth and climatic factors. The saponarin contents were positively correlated with DTR and ALP (r = 0.535 and 0.446, respectively) and negatively correlated with FW, leaf length, GI, ADT, ACT, RH, and rainfall (r = 0.700, 0.704, 0.395, 0.435, 0.597, 0.461, and 0.553, respectively).
Table 4.
Pearson’s correlation coefficient of saponarin content with FW, leaf length, GI, GP, and environmental factors.
3.5. Analysis of PCA through Sprout Growth
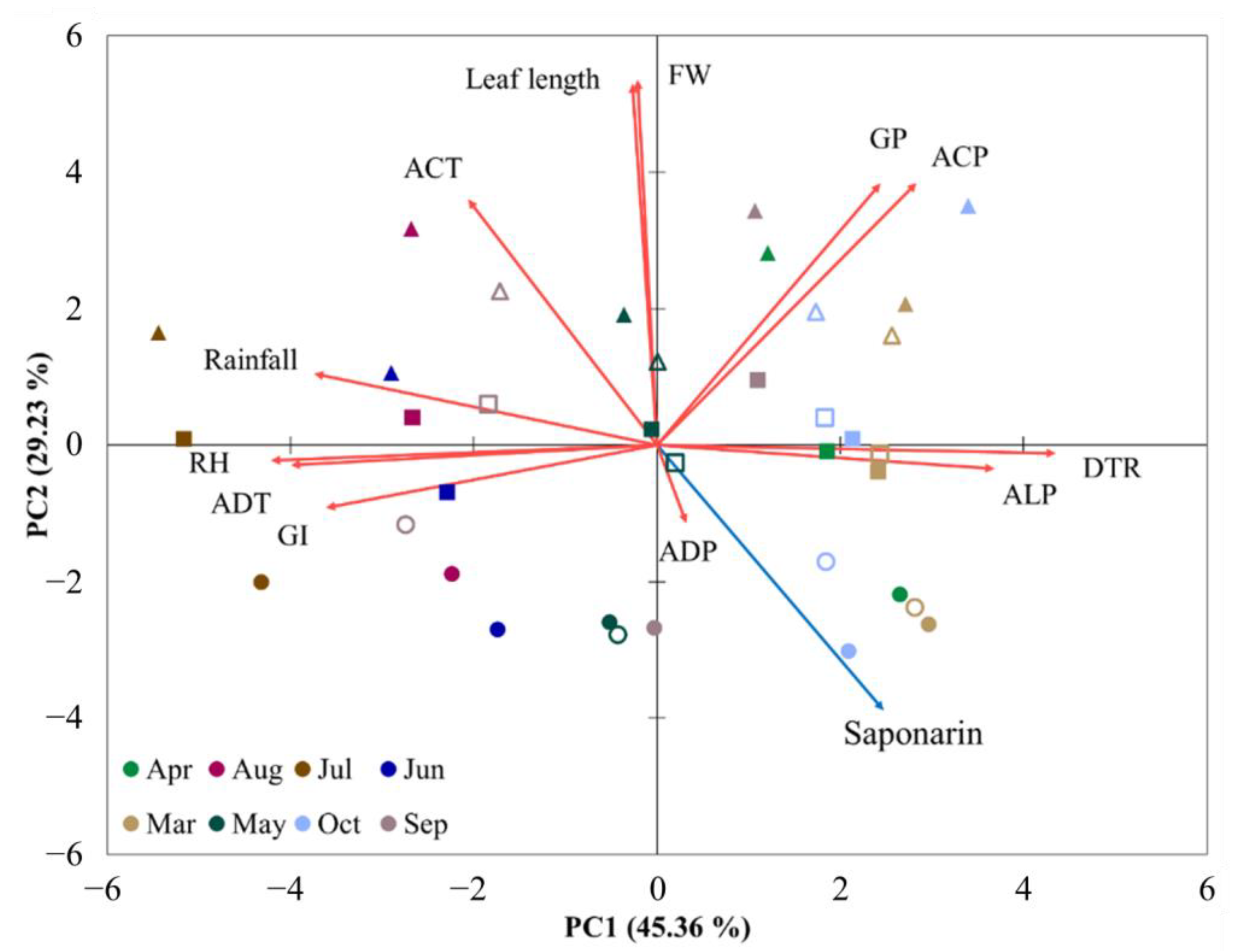
Principal component analysis of all parameters, including FW, leaf length, and environmental factors, provided a clear picture of the effects of sowing time and sprout length (Figure 4). As illustrated in the PC1 axis (45.4% variance), there was a clear separation of samples with the sowing times, due primarily to environmental factors, such as DTR, ALP, ADT, RH, and rainfall. Moreover, the samples were separated by sprout length and FW on the PC2 axis (29.2% variance); in particular, the 10 cm sprouts showed complete separation from samples of sprouts of different lengths, predominantly reflecting the difference in the saponarin content. These multivariate analyses provide essential information on the saponarin patterns in barley sprouts, and their contents may be considered excellent factors of the daily temperature difference and daylight hours between May and October.
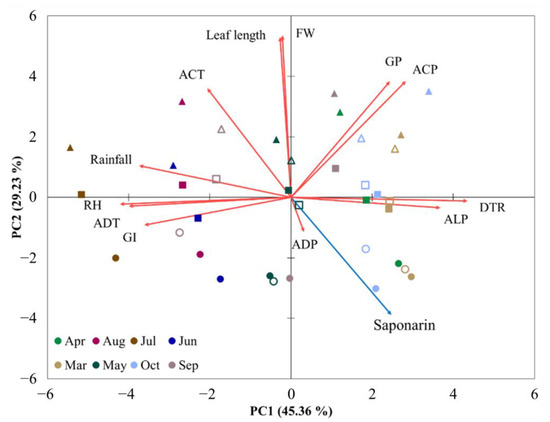
Figure 4.
Principal component analysis (PCA) of the plant parameters with environmental factors. Scores for first two principal components of the sowing times (from March to November). Sowing times are indicated by different colors. Different shapes of symbols indicate the following: circle, 10 cm of leaf length; square, 15 cm; triangle, 18 cm; closed symbols, data in 2019; open symbols, data in 2020. Loading plot of the plant parameters of saponarin (blue arrow) and FW, leaf length, GI, GP, and climatic factors (red arrow). Abbreviations—FW, fresh weight; GI, growth index; GP, growth period; ADT, average daily temperature; DTR, daily temperature range; ACT, accumulated temperature; ALP, average light period; ADP, average daily photons; ACP, accumulated photons; RH, relative humidity.
4. Discussion
The commercial production of sprout vegetables is usually carried out in controlled environments, such as greenhouses and high tunnels. Additionally, crop sprouts have widely been self-produced even at home due to the advantage of growing readily and quickly in limited environmental conditions, such as absences in light and growing medium [5]. In South Korea, a large proportion of commercial production of barley sprouts is performed mainly in outdoor fields rather than inside greenhouses where environmental conditions can be controlled. Thus, many researchers currently have a significant interest in identifying the optimal environmental conditions during germination and early seedling stages to improve the quality and yield of the sprouts. The primary results of this study have proven that the growth of barley sprouts can be primarily correlated with the variations in climatic factors through seasonal change.
Above all, the GI value of this study showed the degree of productivity of barley sprouts according to the sowing times. It is likely that the larger the GI, the more sprouts can be produced through multiple cropping within temporally and spatially limited field conditions. In the present study, the GI values in Jun–July (summer) were approximately twice those in Mar–Apr (spring) and Aug–Oct (autumn) (Figure 1). Such a seasonal effect on GI is thought to influence the total biomass production of barley sprouts at each sowing time, which can be elucidated by the different correlations with various climatic factors. The GI had positive correlations with ADT and RH and a negative correlation with DTR (Table S2 or Table S3), indicating that the differences in temperature, light hours, and humidity along with seasonal changes are the main factors influencing the sprout productivity.
On the other hand, the saponarin content in the sprouted barley exhibited the opposite trend to growth factors, including FW and leaf length, in response to changes in seasonal environmental conditions. This finding showed that the patterns of this compound significantly differed by sowing time, which was highest in October and lowest in July (Figure 2). Namely, the above results indicate that the favorable climatic conditions for sprout growth at high temperatures do not positively affect the production of saponarin. Similarly, many studies have reported that the variations in primary and secondary metabolites (phenolics, flavonoids, and sugars) were considerably different according to the growth seasons of crop sources [24,25,26]; specifically, these substances showed high contents in the spring and autumn compared with summer. It has also been well demonstrated that the optimal temperature of the flavonoid biosynthesis mechanism is between 16 and 25 °C, and flavonoid derivatives show low contents at temperatures exceeding 25 °C [25,27]. Cheng et al. [25] reported that the flavonoid contents of Ginkgo biloba L. were 30% higher in October–November than August–September, when the ADT decreased by 10 °C according to the season change. Consistently, such seasonal decrease in ADT appeared to increase the saponarin content of barley sprouts in this study. Furthermore, the DTRs in October and March when the sprouts contained the highest saponarin content were approximately 14.3–14.8 °C. With the positive correlation concerning the increased rate of DTR patterns (Table 4), daily temperature differences can be pointed out as one of the major climatic factors that promote saponarin biosynthesis.
Several researchers have investigated the effects of different light wavelengths on saponarin contents [14,28]; however, there are few studies about the effects of light intensity. Light has an essential role in plant metabolism by inducing or regulating plant secondary metabolites [29]. Thus, variation in light intensity with the seasonal change may influence saponarin generation in barley sprouts. As reported by previous research data [17,25], some critical substances in flavonoid generation using chalcone synthase (CHS), which is the first enzyme in the flavonoid biosynthesis pathway to synthesize chalcone from p-coumaryl-CoA and malonyl-CoA, and chalcone isomerase (CHI), which synthesizes flavanone from chalcone, were affected by change in light/dark conditions according to season. In the present study, the saponarin contents showed a positive correlation with ALP, but not with ADP and ACP (Table 4). This phenomenon in our field results showed significant differences in comparison with previous results of other indoor studies. Generally, the light intensity in chamber experiments is limited to 380 μmol m−2 s−1; therefore, the saponarin content of barley sprouts is approximately 20 to 800 mg 100 g−1 [17,28,30]. On the other hand, the light intensity of our field areas reached 1711 μmol m−2 s−1, resulting in high production of saponarin in barley sprouts of up to 1900 mg 100 g−1 (data not shown). Barley and other C-3 plants saturate the photosynthetic system when photons are provided at approximately 400 μmol m−2 s−1 for 16 h during the daily growth period [31]. Overall, considering that the light intensity range (571–1711 μmol m−2 s−1) during this field experiment was enough to provide an appropriate light intensity value for barley photosynthetic activity, it seems that temperature has a greater effect than light intensity in terms of saponarin production.
In addition, overall, young leaves of barley have high saponarin contents, and their contents gradually decrease as growth progresses. Inverse relationships of saponarin with FW and leaf length, as well as GI (Figure 4, Table S3), indicate that this functional compound tends to rely greatly on primary leaf elongation, that is, the younger sprouts are, the higher the contents. However, when considering maximizing the total production of saponarin-enriched barley sprouts, a specific growth length was found to recommend the most appropriate harvesting time, which was supported by the polynomial regression test (Figure 3), as follows: a 2.18 mg plant−1 at 15.9 cm in length. Similarly, it was reported that the policosanol contents in barley sprouts were the highest patterns on the 10th and 13th days after sprouting and then decreased on the 20th day [20], and there was an increase in soluble protein content in the boosting stage compared with the tillering and jointing stages [32]. Brauch et al. [30] also demonstrated that the saponarin contents of barley sprouts were lower in the second and third leaves than in the first leaf.
The major summaries of the present research are as follows: (i) The FW of barley sprouts in each leaf length had no significant difference by sowing time, while GP varied; (ii) the GI result showed an inverse relationship with saponarin content; (iii) the saponarin content increased when DTR and ALP (related to temperature and light intensity, respectively) were high, especially in October and March. The annual value of saponarin content in field-grown barley sprouts was 1428 mg 100 g−1. According to the sowing time, the maximum value (1701 mg 100 g−1 for 15 cm) and the minimum value (1207 mg 100 g−1 for 15 cm) were found in October and July, respectively. The environmental conditions to maximize saponarin content during the cultivation period were as follows: average temperature (8.8 °C), daily temperature range (14 °C), accumulated temperature (328 °C), daylight period (7.0 h), daily average photons (52 mol m−2), accumulated photons (2020 mol m−2), relative humidity (67%), and rainfall (25.6 mm), respectively. The yield of barley sprouts cultivated in the field might vary with sowing time due to the difference in growth periods that mediates the possibility of multiple cropping. The saponarin contents in the sprouts appeared to be clearly affected by climatic factors, such as daily temperature differences and daylight hours, although they decreased with increasing rates of leaf length. We demonstrated for the first time the maximum saponarin yield of this species, displaying approximately 16 cm of sprout length. Our data may provide several choices for the use of barley sprouts in agricultural practices of sowing and harvest times. In addition, our work reveals that the optimal conditions for high saponarin yield may be considered an effective natural source for the development of functional foods in barley sprouts. Further studies are needed to comprehensively evaluate the effects of a wide range of environmental factors, including climate factors, for production of high-quality barley sprouts.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy12092056/s1, Table S1: Climatic information in an upland field during two-year experimental duration; Table S2: Correlation matrix of FW, leaf length, GI, GP, and climatic factors; Table S3: Loading values of the plant parameters of saponarin, FW, leaf length, GI, GP, and climatic factors.
Author Contributions
Conceptualization, Y.-E.Y., W.D.S., J.H.L., M.S.C., K.-A.L., Y.-N.K. and Y.B.L.; Methodology, Y.-E.Y., V.K., W.D.S. and Y.B.L.; Data curation, Y.-E.Y. and Y.-N.K.; Formal analysis, Y.-E.Y., H.C. and V.K.; Validation, H.C.; Writing—original draft, Y.-E.Y.; Writing—review & editing, J.H.L., M.S.C., K.-A.L. and Y.-N.K.; Investigation, Y.-N.K.; Funding acquisition, Y.B.L.; Project administration, Y.B.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ014212032022)” Rural Development Administration, Republic of Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Daferera, D.J.; Ziogas, B.N.; Polissiou, M.G. GC-MS Analysis of Essential Oils from Some Greek Aromatic Plants and Their Fungitoxicity on Penicillium digitatum. J. Agric. Food Chem. 2000, 48, 2576–2581. [Google Scholar] [CrossRef]
- Smith, J.V.; Luo, Y. Studies on Molecular Mechanisms of Ginkgo biloba Extract. Appl. Microbiol. Biotechnol. 2004, 64, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Shu, Z.; Hussain Sh, A.; Shahen, M.; Wang, H.; Alagawany, M.; Abd El-Hac, M.E.; Ali Kalhor, S.; Rashid, M.; Ali Shar, P. Pharmacological Uses of Ginkgo biloba Extracts for Cardiovascular Disease and Coronary Heart Diseases. Int. J. Pharmacol. 2018, 15, 1–9. [Google Scholar] [CrossRef]
- Aborus, N.E.; Čanadanović-Brunet, J.; Ćetković, G.; Šaponjac, V.T.; Vulić, J.; Ilić, N. Powdered Barley Sprouts: Composition, Functionality and Polyphenol Digestibility. Int. J. Food Sci. Technol. 2017, 52, 231–238. [Google Scholar] [CrossRef]
- Di Gioia, F.; Renna, M.; Santamaria, P. Sprouts, Micogreens and “Baby Leaf” Vegetables. In Minimally Processed Refrigerated Fruits and Vegetables; Yildiz, F., Wiley, R.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 723–745. ISBN 978-1-4939-7016-2. [Google Scholar]
- Meyerowitz, S. Sprouts, the Miracle Food: The Complete Guide to Sprouting, 8th ed.; Sproutman Publication: Great Barrington, MA, USA, 2010. [Google Scholar]
- Lee, W.Y.; Emmy, H.K.I.; Abbe, M.M.J.; Amin, I. Antioxidant Capacity and Phenolic Content of Selected Commercially Available Cruciferous Vegetables. Malays. J. Nutr. 2007, 13, 71–80. [Google Scholar]
- Abdallah, M.M.F. Seed Sprouts, a Pharaoh’S Heritage to Improve Food Quality. Arab Univ. J. Agric. Sci. 2008, 16, 469–478. [Google Scholar] [CrossRef]
- Ebert, A. Sprouts, Microgreens, and Edible Flowers: The Potential for High Value Specialty Produce in Asia. In Proceedings of the SEAVEG 2012: High Value Vegetables in Southeast Asia: Production, Supply and Demand, Chiang Mai, Thailand, 24–26 January 2012; pp. 216–227. [Google Scholar]
- Kamiyama, M.; Shibamoto, T. Flavonoids with Potent Antioxidant Activity Found in Young Green Barley Leaves. J. Agric. Food Chem. 2012, 60, 6260–6267. [Google Scholar] [CrossRef]
- Song, J.S.; Lee, M.J.; Ra, J.E.; Lee, K.S.; Eom, S.; Ham, H.M.; Kim, H.Y.; Kim, S.B.; Lim, J. Growth and Bioactive Phytochemicals in Barley (Hordeum vulgare L.) Sprouts Affected by Atmospheric Pressure Plasma during Seed Germination. J. Phys. D Appl. Phys. 2020, 53, 314002. [Google Scholar] [CrossRef]
- Byun, A.R.; Chun, H.; Lee, J.; Lee, S.W.; Lee, H.S.; Shim, K.W. Effects of a Dietary Supplement with Barley Sprout Extract on Blood Cholesterol Metabolism. Evidence-based Complement. Altern. Med. 2015, 2015, 473056. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, S.Y.; Kim, B.; Seo, W.D.; Jia, Y.; Wu, C.; Jun, H.J.; Lee, S.J. Barley Sprout Extract Containing Policosanols and Polyphenols Regulate AMPK, SREBP2 and ACAT2 Activity and Cholesterol and Glucose Metabolism In Vitro and In Vivo. Food Res. Int. 2015, 72, 174–183. [Google Scholar] [CrossRef]
- Kim, J.S.; Jeong, E.; Jo, S.M.; Park, J.; Kim, J.Y. Comparative Study of the Effects of Light Controlled Germination Conditions on Saponarin Content in Barley Sprouts and Lipid Accumulation Suppression in HepG2 Hepatocyte and 3T3-L1 Adipocyte Cells Using Barley Sprout Extracts. Molecules 2020, 25, 5349. [Google Scholar] [CrossRef] [PubMed]
- Frangne, N.; Eggmann, T.; Koblischke, C.; Weissenböck, G.; Martinoia, E.; Klein, M. Flavone Glucoside Uptake into Barley Mesophyll and Arabidopsis Cell Culture Vacuoles. Energization Occurs by H+-Antiport and ATP-binding Cassette-type Mechanisms. Plant Physiol. 2002, 128, 726–733. [Google Scholar] [CrossRef]
- Seo, K.H.; Park, M.J.; Ra, J.E.; Han, S.I.; Nam, M.H.; Kim, J.H.; Lee, J.H.; Seo, W.D. Saponarin from Barley Sprouts Inhibits NF-κB and MAPK on LPS-induced RAW 264.7 Cells. Food Funct. 2014, 5, 3005–3013. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Woo, S.-Y.; Ra, J.-E.; Lee, K.-S.; Seo, W.D.; Lee, J.H. Saponarin Content and Biosynthesis-related Gene Expression in Young Barley (Hordeum vulgare L.) seedlings. J. Plant Biotechnol. 2019, 46, 247–254. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Bassal, A.; Leonardi, C.; Giuffrida, F.; Colla, G. Vegetable Quality as Affected by Genetic Agronomic and Environmental Factors. J. Food Agric. Environ. 2012, 10, 680–688. [Google Scholar]
- Yoon, Y.-E. Establishment of Optimal Growing Condition for Production of Saponarin-Enriched Barley Sprouts (Hordeum vulgare L.). Ph.D. Thesis, Gyeongsang National University, Jinju, Korea, 2022. [Google Scholar]
- Seo, W.D.; Yuk, H.J.; Curtis-Long, M.J.; Jang, K.C.; Lee, J.H.; Han, S.I.; Kang, H.W.; Nam, M.H.; Lee, S.J. Effect of the Growth Stage and Cultivar on Policosanol Profiles of Barley Sprouts and Their Adenosine 5′-monophosphate-activated Protein Kinase Activation. J. Agric. Food Chem. 2013, 61, 1117–1123. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis: Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Summer, M.E., Eds.; Soil Science Society of America-American Society of Agronomy: Madison, WI, USA, 1996; Chapter 34; pp. 961–1010. [Google Scholar] [CrossRef]
- Kjeldahl, J.A. New Method for the Determination of Nitrogen in Organic Matter. Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef]
- NIAST. Method of Soil and Plant Analysis; National Institute of Agriculture Science and Technology: Suwon, Korea, 2000. [Google Scholar]
- Cao, Y.; Fang, S.; Fu, X.; Shang, X.; Yang, W. Seasonal Variation in Phenolic Compounds and Antioxidant Activity in Leaves of Cyclocarya paliurus (Batal.) Iljinskaja. Forests 2019, 10, 624. [Google Scholar] [CrossRef]
- Cheng, S.; Xu, F.; Li, L.; Cheng, H.; Zhang, W. Seasonal Pattern of Flavonoid Content and Related Enzyme Activities in Leaves of Ginkgo biloba L. Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 98–106. [Google Scholar] [CrossRef]
- Ribeiro, D.A.; Camilo, C.J.; de Fátima Alves Nonato, C.; Rodrigues, F.F.G.; Menezes, I.R.A.; Ribeiro-Filho, J.; Xiao, J.; de Almeida Souza, M.M.; da Costa, J.G.M. Influence of Seasonal Variation on Phenolic Content and In Vitro Antioxidant Activity of Secondatia floribunda A. DC. (Apocynaceae). Food Chem. 2020, 315, 126277. [Google Scholar] [CrossRef]
- Xu, F.; Deng, G.; Cheng, S.; Zhang, W.; Huang, X.; Li, L.; Cheng, H.; Rong, X.; Li, J. Molecular Cloning, Characterization and Expression of the Phenylalanine Ammonia-lyase Gene from Juglans regia. Molecules 2012, 17, 7810–7823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, N.J.; Kim, J.Y.; Lee, Y.; Shin, S.H.; Song, J.S.; Shin, S.C.; Kim, B.T. Variations of Saponarin Content in Young Barley Leaves Illuminated with Different Light-Emitting Diodes (LEDs). J. Crop Sci. Biotechnol. 2019, 22, 317–322. [Google Scholar] [CrossRef]
- Yang, J.; Wang, P.; Wu, W.; Zhao, Y.; Idehen, E.; Sang, S. Steroidal Saponins in Oat Bran. J. Agric. Food Chem. 2016, 64, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Brauch, D.; Porzel, A.; Schumann, E.; Pillen, K.; Mock, H.P. Changes in Isovitexin-O-glycosylation during the Development of Young Barley Plants. Phytochemistry 2018, 148, 11–20. [Google Scholar] [CrossRef]
- Sager, J.C.; McFarlane, J.C. Radiation. In Plant Growth Chamber Handbook. In Growth Chamber Handbook; North Central Regional Research Publication No. 340; Iowa State University: Ames, IA, USA, 1997; pp. 1–29. [Google Scholar]
- Deng, L.N.; Feng, G.N.; Gao, Y.; Shen, Y.X.; Li, H.S.; Gu, Y.; Luan, H.Y. Phytochemical Constituents and Antioxidant Enzyme Activity Profiles of Different Barley (Hordeum vulgare L.) Cultivars at Different Developmental Stages. Agronomy 2020, 10, 37. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).