Abstract
The short-term effects of soil amendments on the structure, diversity and function of a nematode community of Miscanthus × giganteus was investigated. Crop was cultivated on marginal, nutrient-poor land amended with biochar in single and double doses (BD1 and BD2), biogas digestate (D), sewage sludge (SS), and hemicellulose waste (HW). Sampling was done after planting, in the middle and end of vegetation; morphology-based approach was used. 28 nematode taxa were identified, including 5 bacterivores genera, 4 fungivores genera, 5 herbivores genera (11 species), 2 omnivores genera, 5 predators genera. The general linear models, correspondence analysis and clustering were applied for evaluation. The total abundance of nematode taxa Filenchus, Dorylaimus, Cephalobus, Panagrolaimus, Aphelenchus, and Ditylenchus was depended on the sampling time and amendments. The incorporation of amendments affected nematode food web and resulted in suppression of plant-parasitic nematodes (PPNs). It was revealed that community structure was more mature for SS, less stable for D and had inconclusive effects for BD1, BD2, and HW. Using amendments ensured pest control benefits which is important given concern that PPNs can inflict crop damage during increased cultivation of M × g. Further research is needed to examine amendments which can minimise PPNs without reducing populations of nitrogen-fixing bacterivores and fungivores.
1. Introduction
The soil organic amendments produced from biomass or living organisms typically include compost, biochar, wood chips, charcoal, animal manure, straw, husk, geotextiles, sewage sludge, digestate, and hemicellulose waste from paper production [1,2,3,4,5,6,7]. These compounds are rich in plant nutrients and contain high levels of organic matter, macro- and microelements that improve soil fertility by enhancing microclimatic conditions and providing microbial growth substrates [8,9,10,11,12,13,14,15,16,17]. The incorporation of soil amendments can be beneficial in the production of Miscanthus × giganteus J.M. Greef & Deuter ex Hodkinson & Renvoize (Angiospermae: Poaceae) (M × g) in poor soils by increasing soil fertility and maximising crop yield [1]. M × g has a relatively high biomass yield and cellulose content compared with other lignocellulosic bioenergy crops [18], which makes it one of the most promising feedstocks for the production of biofuels [19] and bioproducts: fibres, insulation materials, and papers [20]. The cultivation of M × g on the slightly contaminated or marginal lands has environmental and economic benefits: in addition to the remediation potential [21,22], the plant demonstrates high efficiency in carbon sequestration [23].
Sewage sludge is a widely used soil amendment of M × g cultivation proposed to improve the biomass production on agricultural, marginal and slightly contaminated by trace elements land [2,3,4]. The application of this amendment ensures high crop yields and biomass quality that can be used as raw material for energy and bio-products and improves soil health.
There is a close connection between biochar and bioenergy production: this amendment can be produced from the residues of bioenergy crops, generated during biochar production, and biochar can be applied to fields where energy crops including M × g are grown [5,6]. Due to the ability of biochar to restore and remediate deteriorated soils [7], it is widely touted as a valuable land management tool with great potential to mitigate climate change [8]. Its application can improve soil quality in a variety of ways, including increasing soil water holding capacity and crop yield, raising pH, and increasing crop yield [8]. Biochar amendment affects soil structure and function [9], mainly by altering nutrient cycling [10] and influencing soil microbial communities [11].
During the production of energy crops on marginal land, another substance positively influencing soil fertility and biomass is the digestate from agricultural biogas plants [12]. It can be used directly or processed into the amendment, the latter approach is more sustainable and beneficial to the environment [13].
The use of paper mill sludge to improve soil quality and increase crop yield has been reported [14,15,16], but caution should be exercised when using it as an amendment. The main concern of using the sludge is related to its content of industrially-derived contaminants of trace elements. As a result, the incorporation of this amendment may lead to new (secondary) contamination of the soil by contaminants presented in the sludge [17].
While using soil amendments to increase M × g production on marginal land, it is necessary to investigate the regulation of soil quality, which can be done by studying the soil biota. Nematodes are a dominant component of the soil biota and serve as primary, secondary, and tertiary consumers in soil food webs, thus reflecting their complexity. These organisms, which live in soil microecosystems, are sensitive to environmental changes and can be proposed as valuable bioindicators for soil health assessment [24,25]. The abundance and diversity of soil nematode communities can be used to holistically assess biotic and functional soil conditions due to the use of organic amendments [26].
Subterranean nematodes are represented by two groups: plant-parasitic nematodes (PPNs) or herbivores that parasitise and damage plants and free-living nematodes (FLNs) [27] providing the various ecosystem services, e.g., nutrient cycling [28,29], microbial redistribution in the soil profile [30], and insect control [31]. FLNs are classified as fungivores, bacterivores, predators, and omnivores that can be beneficial for soil fertility and ecosystem function [27,32].
Numerous studies have investigated the effects of organic amendments and fertilisers on soil nematodes [33,34,35,36,37]. Organic amendments were proposed for the management of PPN populations [38,39]. Composted sewage sludge and fly ash from coal combustion had the potential to control populations of root-knot [40] and cyst nematodes [41,42]. Some studies on the effects of biosolids on nematode communities illustrated existed disparity in results. Nematode abundance and diversity were significantly affected by five different sewage sludge treatments in five soil types [43]. The most evident response to sewage sludge application was seen in bacterivores, with a negative correlation between the sewage sludge dose and nematode communities.
The effects of biochar on soil nematodes are less well studied [44], and results illustrated different responses of PPNs to the treatment. Biochar incorporation reduced the abundance of the stubby-root nematode Trichodorus sp. [45] in maize crops and the migratory endoparasite nematode Pratylenchus penetrans Cobb in carrot crops [46]. Five-year monitoring of the impact of biochar and manure added to low fertile yellow cinnamon soil showed significant decreasing of PPNs levels. Moreover, the strong dominance of bacterivores was caused by increasing the soil productivity, which increased wheat yield [47]. Wanjiku Kamau et al. [45] observed a significant reduction in bacterivorous nematodes during maize vegetation in biochar-enriched soils. Microbial and bacterivore alterations could explain PPNs suppression mechanisms that occurred in biochar-enriched soils [48]. The addition of biochar to rice potting soils reduced the crop’s susceptibility to the root-knot nematode Meloidogyne graminicola Golden & Birchfield [49]. In some cases [50,51] biochar did not impact to the potato cyst nematode Globodera sp., while neutral and negative effects were determined for the root-knot nematode (Meloidogyne sp.) [44,51,52]. Several studies [53,54] explored the effect of biogas digestate (D) and hemicellulose waste from paper production (HW) on soil nematodes. It was shown that the combined treatment of soil with HW and urea reduced the population of soybean nematodes under greenhouse conditions [53]. The addition of D showed the potential to limit infestations of Meloidogyne incognita Kofoid & White and Pratylenchus penetrans [54].
In our previous study [4], the impact of soil amendments and agricultural practices on soil nematode community under M × g cultivation was investigated, while the post-military Pb-contaminated land was amended with biosolid from the wastewater treatment plant in combination with tillage treatment. The results showed that three-year crop growth affected the soil nematode community, which was highest in two exploited cases: tillage and tillage + biosolids. In the control case without agronomic practices, the nematode community diversity was enriched by increased populations of herbivores, omnivores and predators.
We are not aware of any systematic study that comprehensively addresses the assemblage and diversity of the soil nematode community during the cultivation of M × g on marginal land amended with various organic soil amendments. The main goal of the current study was to evaluate the short-term (one vegetation season) effect of soil amendments of different origins on the composition and diversity of the nematode community under M × g growth on marginal land. The following organic amendments were used: biochar in doses of 5 and 10% (BD1 and BD2), biogas digestate (D), sewage sludge (SS), and hemicellulose waste (HW).
2. Materials and Methods
2.1. Experimental Field
The soil for investigation was taken from M × g plantation established in 2021 on marginal land in Nové Spořice, a suburb of Chomutov city, Czech Republic (50°27′38” N, 13°23′07” E).
The experimental field was a randomised complete block with four replicates for each soil amendment. Each plot was 2.5 m × 2 m and contained 30 M × g rhizomes planted at a depth of 10–12 cm, with 1 m passageways between plots. The properties of the soil amendments used in the experiment are listed in Table 1 and Table 2. The amendments were added to the soil on 26 April 2021, and the rhizomes were planted on 27 April 2021. Each rhizome was planted at the pointed place in the soil pre-treated with a targeted amendment. The planting material was M × g obtained from the Miscanthus d.o.o. deposit field (Popovača, Sisak-Moslovina County, Croatia; https://miscanthus.hr (accessed on 1 July 2022)). The rhizomes were three years old and had an average weight of 20 ± 2 g. At the end of the vegetation season, on 30 October 2021, the biomass was harvested.

Table 1.
Characteristics of the soil amendments utilized during establishing of M × g plantation on the marginal land.
Table 2.
Chemical content of the soil amendments.
2.2. Soil Characteristics
Prior to establishment of M × g plantation, the research soil was collected for the measurement of agrochemical parameters. Soil sampling was carried out from one 5 m × 5 m testing square using the standard approach DSTU 4287:2004 [55], and five samples were collected at a depth of 0–0.3 m by quartering and mixing. The agrochemical parameters of the soil are given in Table 3, and the characteristics of the soil with targeted amendments are presented in Table S1. According to the standards [56,57], the soil reacts strongly acidic after salt extraction to become medium acidic after water extraction; soil K content is high, Mg content is good, Ca content is satisfactory, P content is low, and soil organic matter content is classified as medium-high. According to [58], the soil type is Cambisol; the soil-forming substrate is a mainly acidic metamorphic rock. The average altitude of the land in the area concerned is 360 m above sea level. The average annual rainfall is quite low (the foothills of the rain shadow), reaching a value of 525 mm, and the average rainfall per growing season is 300 mm. The average annual air temperature is 7 °C.
Table 3.
The agrochemical parameters of the research soil.
2.3. Nematode Analysis
2.3.1. Nematode Sampling
A soil sampling for nematodes identification was done three times during the first growing season, i.e.: on 21 May 2021–immediately after planting M × g (Pi), on 20 July 2021–in the middle of vegetation (Pf1), and on 9 October 2021–at the end of vegetation (Pf2). The composite soil sample (500 g) was derived from five sampling spots of each replication. The soil was taken directly from the root zone of M × g at a depth of 35 cm. Upon arrival to the laboratory, the composite soil samples were thoroughly mixed and 100 cm3 of each was used for nematode identification.
2.3.2. Nematode Isolation and Identification
The rapid centrifugation–flotation method [62] was used to extract nematodes from the soil samples. The soil sample (100 cm3) was placed in a 600 mL beaker and sufficient volume of water was added to bring the total volume to 500 mL. The sample was stirred and allowed the soil to settle to the bottom of the beaker. Thereafter the sediment suspension was decanted and placed in a 200 mL tube to centrifuge 3 min at 2000× g (RCF). After that, the supernatant was poured off and discarded. The pellet was resuspended in about 80 mL sucrose solution (specific gravity, 1.18, 38.5% by weight or 484 g per 1000 mL of water). Tubes were centrifuged again 2 min at 1800× g (RCF). The supernatant containing the nematodes was gently poured over a 25 μm sieve. Nematodes were transferred from the sieve into glass containers.
The isolated nematodes were killed and fixed, then passed through a graded series of glycerol–ethanol solutions and stored in anhydrous glycerol on slides [63]. The isolated nematodes were heat-killed and fixed with 5% formalin. Fixed nematodes were transferred to a fixation dish with added S1 solution (20 mL of 96% ethanol, 1 mL glycerine, 79 mL distilled water). The dishes were placed in a desiccator with a small layer of 96% ethanol and put in the incubator at 40 °C. After 16–24 h, the S1 was sucked away under the dissecting microscope using a micropipette, and the S2 solution (93 mL of 96% ethanol and 7 mL glycerine) was added. The fixation dish was put back in the incubator. After 2 h, two drops of dehydrated glycerine were added. Once the alcohol evaporated after 24 h, the fixation dish was placed in the desiccator, containing silica gel to allow the last water traces to vaporize. 100 individuals initially detected per sample were identified as species (PPNs) and genus using the Zeiss Jena A-Scope microscope using the illustrated key by Brzeski [64] and Andrássy [65]: Bacterivore, Fungivore, Omnivore, and Predator.
2.3.3. Nematode Fauna Analysis Based on Life Traits and Feeding Habitats
The analysis of the nematode community was carried out using the total number of nematodes (abundance) and the weighted proportion of individuals detected in the soil sample. To calculate biodiversity indices, soil nematodes were assigned to 5 trophic groups, following Yeates et al. [32].
Each nematode taxon was classified into one of five coloniser-persisters (c-p) classes based on life history traits and food preferences. “Coloniser” nematodes at the lower end of the c-p scale were considered enrichment opportunists and therefore indicated resource availability; “persister” nematodes were considered at the upper end of the scale and indicated system stability and food web complexity.
The maturity index (MI) for FLNs and the plant parasite index (PPI) for herbivores were calculated based on the ratio of a given genus to the total number of nematodes and their assigned c-p values according to Bongers [66]. Several indices of the structure and complexity of the food web in the soil were calculated according to Ferris et al. [27]. In this context, the following parameters were determined: the channel index (CI) based on fast-growing, bacterivores and fungivores characterised by a c-p value of 1 or 2; the basal index (BI) based on bacterivores and fungivores with a c-p value of 2; the enrichment index (EI), which was considered an indicator of conditions favouring fast-growing bacterivores and the structure index (SI), which correlated with the maturity of an ecosystem that combines nematode genera with c-p values of 3 to 5. In addition, the relative abundance of nematode functional guilds (trophic and c-p groups) was calculated.
The SI–a weighted mean of the proportion of sensitive predatory and omnivorous nematodes–is an indicator of the complexity of the soil food web due to soil disturbance. The CI is the ratio of fungivores and bacterivores and indicates the prevalence of organic matter decomposition mediated by fungi. The BI is based on the abundance of general opportunist nematodes and indicates basal, disturbed conditions in the soil food web. The EI is based on the abundance of enrichment opportunist nematodes and indicates the rapid, bacterially mediated decomposition of organic matter [27]. The scatter plot of SI against the EI values, divided into four quadrants, has been suggested to diagnose the soil food web into four potential states: structured, maturing, degraded, and disturbed [27]. The ecological indices were calculated using an online tool NINJA: Nematode Indicator Joint Analysis [67].
2.4. Statistical Analysis
The Multivariate general linear models (MGLM) were proposed to test the significance of the impact of the applied agronomic practices on the nematode community using the Statistics v. 12.0 PL software package from TIBCO Software (Inc. Statistics, Palo Alto, CA, USA, 2014). Two multivariate GLMs were run with the part of the sampling time (three levels), soil amendment type (five levels), and sampling time × amendments; treatment (interaction) as categorical predictors. A MGLM with the set of nematode taxa and another MGLM with the set of community indices as the multivariate dependent variables were conducted, testing differences between individual treatments with a planned comparison.
Detrend correspondence analysis (DCA) was applied to determine whether species response to environmental gradients was predominantly monotonic or predominantly unimodal. The value was estimated from the length of the first major gradient of variation in the community data. If the length of the gradient exceeds two standard deviations, partial or constrained correspondence analysis (CCA) should be chosen as the ordinal approach. Otherwise, partial or constrained redundancy analysis (RDA) is most appropriate for the evaluation [68].
3. Results
An average of 984 nematodes per 100 g of dry soil were detected in all treatments right after planting M × g, this value was 1472 at the end of vegetation. 28 nematode taxa were identified, including 5 bacterivorous genera, 4 fungivorous genera, 5 herbivorous genera (11 species), 2 omnivorous genera, and 5 predatory genera (Table S2). The abundance of different nematode taxa was tested as a function of sampling time and amendment type (Table 4). These two effects explained up to 90% of the variation in the nematode population (Table 5). The abundance of Filenchus and Dorylaimus was higher in Pf1 than in Pf2, while the abundance of Cephalobus, Panagrolaimus, Aphelenchus, Ditylenchus, Filenchus, and Dorylaimus was lower in Pf1 than in Pf2.
Table 4.
Multivariate GLM results examining the effect of amendment type and sampling time on nematode assemblage composition.
Table 5.
Response of given nematode taxa abundance to the effect of the sampling time from multivariate GLMs.
The application of BD1 and BD2 reduced the abundance of 10 and 13 taxa, respectively. An increase in abundance was observed for only one taxon, Enchodelus sp. for both treatments BD1 and BD2. The application of D decreased the abundance of 12 taxa, while 4 taxa increased. SS decreased the abundance of 17 taxa, while an increase was observed only for Coomanus sp. The greatest effect of soil amendment on nematode abundance was observed for HW: a decrease in abundance was observed for 19 taxa and an increase for 4 taxa.
EIs were calculated to measure the susceptibility of the nematode community to environmental change by selecting appropriate principal components (PC1–PC4) with eigenvalues greater than 1 (Table 6). The first 4 components captured 89.6% of the variation in the indices. PC1 explained a maximum of 48.1% of the data variation, correlated positively with maturity, SI, the proportion of herbivores, predators, omnivores, and migratory endoparasites and negatively with the ratio of PPI and MI, ectoparasites, BI, enrichment footprint, and the proportions of fungivores and bacterivores. Thus, the variation in nematodes in PC1 could be determined by the maturity of the nematode community. PC2 captured 25.8% of the community variation, it correlated negatively with CI and positively with EI, total biomass, footprints of composite, structure, and omnivores. Nematode variation in PC2 could be determined as the biomass of the nematode community increased as omnivores increased. PC4 explained 6.8% of the variation and correlated positively with predator footprint. This component determined a change in the structure of trophic groups, which was associated with a variation in the importance of herbivores, bacterivores and predators.
Table 6.
PCA of variation in the synecological characteristics of nematode communities.
PC1–PC4 scores were affected individually by sampling time during the growing season and the amendments type (Table S3, Figure S1). The effects of the growing season and the amendments treatments were significant for PC1–PC3 (Table S4). Nematode community structure did not differ in sampling time Pf1 and Pf2, while in Pi a statistically significant increase in PC2 and PC3 was observed. The addition of BD1 and BD2 increased PC2 compared to the control; the incorporation of D decreased PC1 and increased PC2 and PC3. The application of SS increased PC1-PC3, and PC2 increased with the addition of HW.
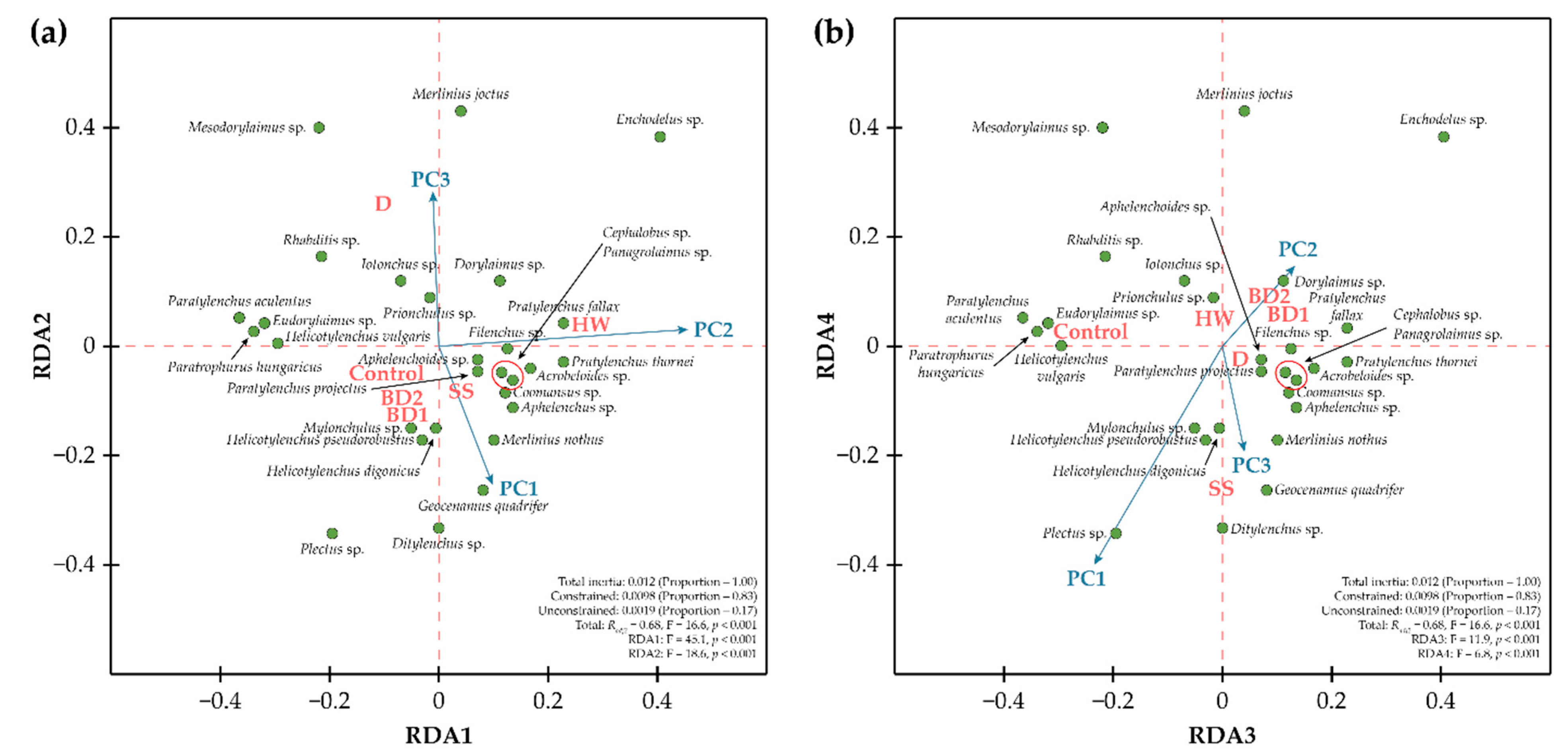
The length of the largest gradient identified by the detrended correspondence analysis was less than 2 (1.68), and the partial redundancy analysis (pRDA) was applied as an ordination procedure to examine trends in the variability of community composition for each treatment by soil amendments (Figure 1). The pRDA was statistically significant and explained a large portion of the community variances (Radj2 = 0.68, F = 16.6, p < 0.001). The total inertia was 0.012 and explained 83% of constrained variables represented by amendments’ treatment categories. The pRDA was useful in elucidating trends in the variability of the nematode community structure among amendments type. The shifts in nematode community composition could be explained by four statistically significant axes, RDA1–RDA4. The RDA1 reflected the variability of the nematode community caused by the influence of HW, and this axis correlated with PC2. The change in nematode community composition caused by HW was due to changes in the number of taxa, such as Paratylenchus projectus Jenkins, Pratylenchus fallax Seinhorst, Aphelenchoides sp., Coomansus sp., and Mylonchulus sp. The RDA2 axis showed the variability in the nematode community composition caused by the effect of D amendments; this axis showed a positive correlation with PC3 and a negative correlation with PC1. D amendments caused the variations in nematode community composition due to changes in the number of taxa such as Merlinius joctus Thorne. The RDA3 axis showed differences in the nematode community under the effects of treatments compared to the control. The RDA4 axis showed the effect on the nematode community composition caused by SS enrichment. The addition of SS was associated with an increase in PC1 and PC3 scores, while the addition of HW had the opposite effect.
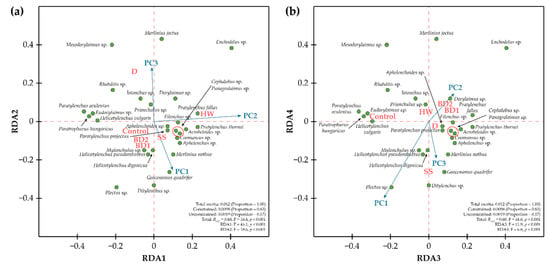
Figure 1.
Partial RDA of the nematode community with biological stimulators as constrained variables (red colour) and fitting of the synecological PC into the RDA axes (blue arrows). Green points indicate species positions. (a) RDA1-RDA2; (b) RDA3-RDA4.
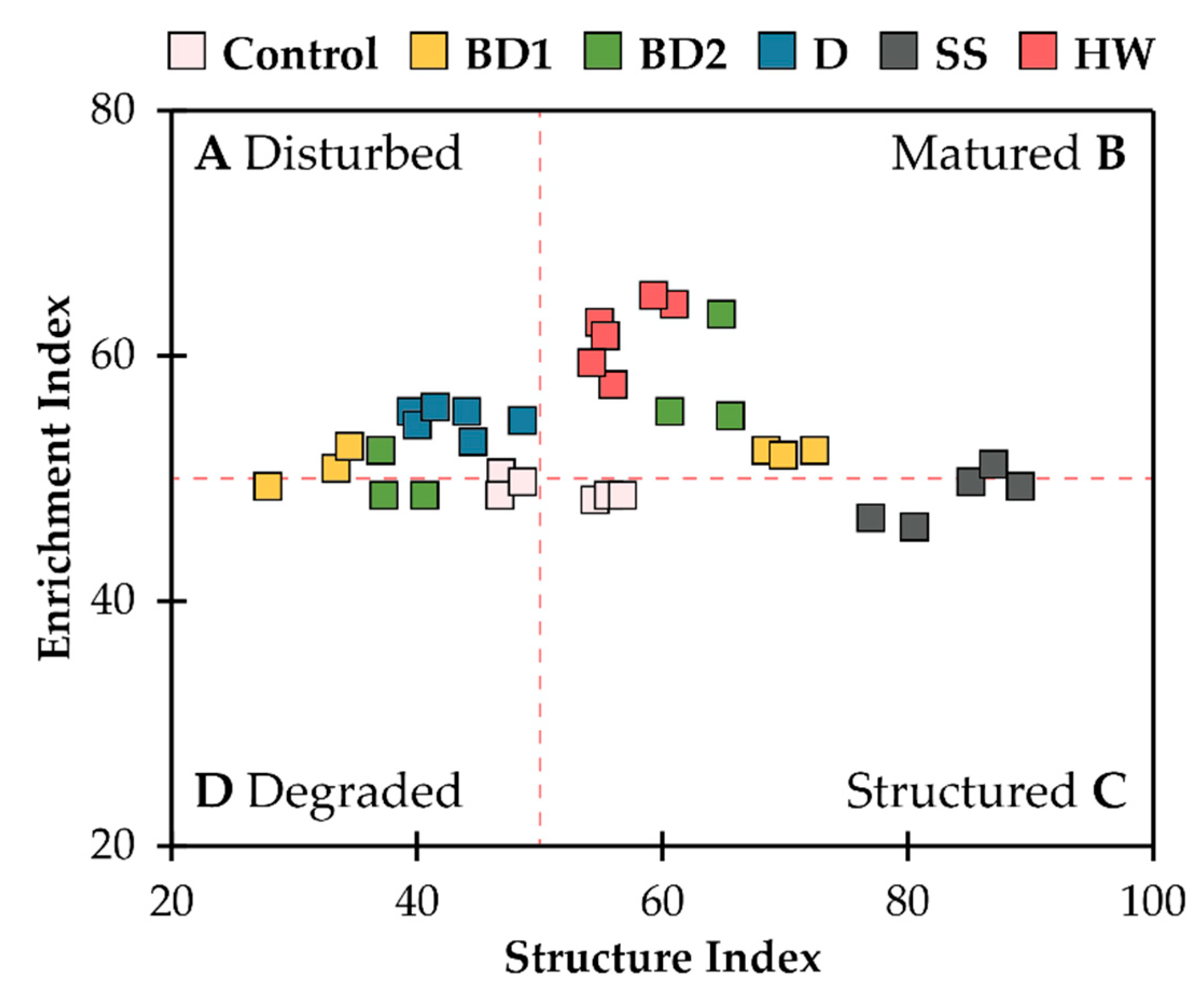
Using a scatter plot between SI and EI, the food web of soil nematodes into four possible states after the application of soil amendments was classified, namely: disturbed/enriched, matured/enriched, structured/stable, and degraded/depleted. A nematode community characterised by low EI and high SI was likely to be found in healthy soil (Figure 2). The addition of SS was reflected in a more stable and mature nematode community, and HW treatment resulted in a more mature state of the nematode food web. D treatment resulted in the nematode food web transformation towards a disturbed and degraded state. The effects of BD1 and BD2 treatments were more controversial, and changes in nematode composition at all trophic levels were observed in both the matured and degraded states.
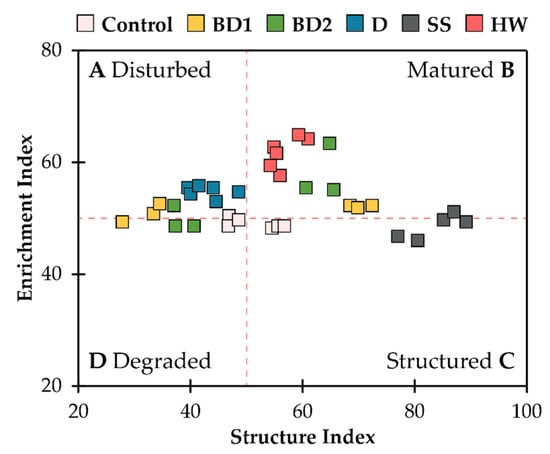
Figure 2.
Nematode faunal analysis indicating the food web status (A–D) in different amendments type: BD1, BD2, D, SS, and HW.
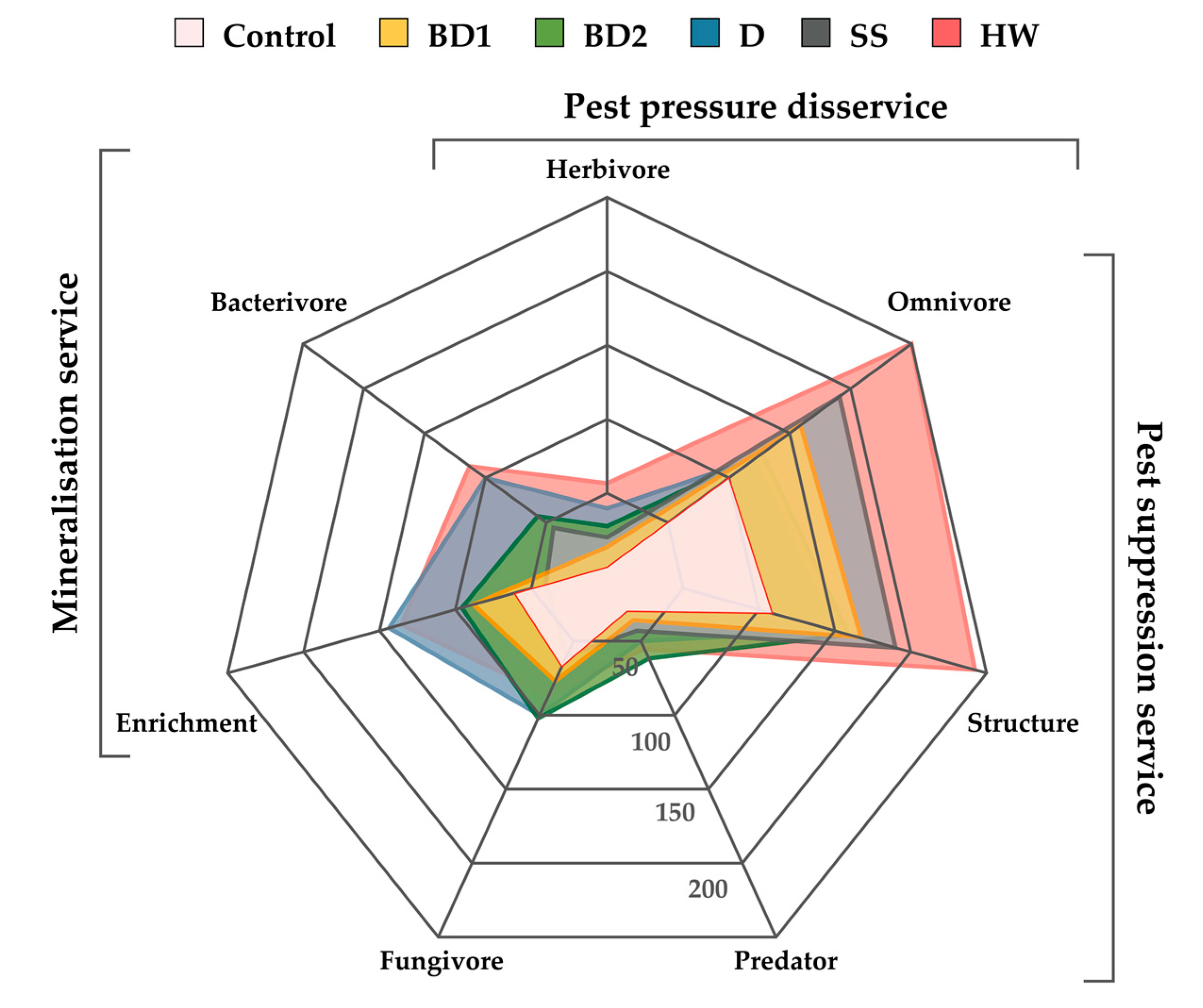
A radar chart was used to assess the ecosystem services of soil nematodes after the application of amendments, which allowed to show the trend of functional metabolic footprints of soil nematodes under the influence of different amendments (Figure 3). The greatest increase in nematode community biomass and/or activity occurred in HW treatment, with slightly smaller effects in SS, BD1, and BD2 treatments. D treatment had the least stimulatory effect on the ecosystem services of the nematode community. The application of amendments primarily activated the ecosystem service for disease suppression and pest control. It should be mentioned that the effect of amendments on M × g yield was not statistically significant (Table S3).
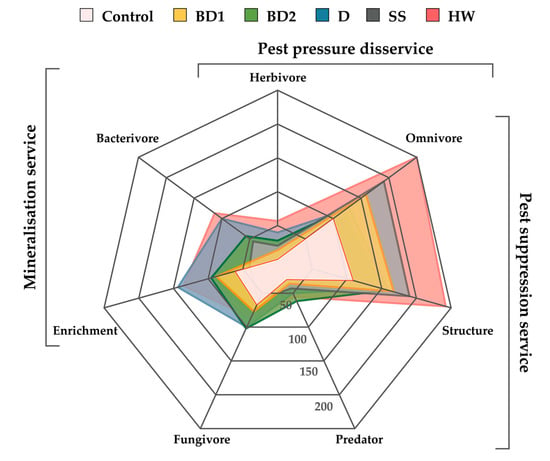
Figure 3.
Radar chart of the metabolic footprint of each nematode trophic group in the different treatments and delivered ecosystem services.
4. Discussion
This study investigated the effect of different organic amendments added to the marginal soil on the nematode community during three stages of the first M × g. vegetation season. The effect appeared at the level of individual taxa and the community as a whole. Using PCA, the most important patterns affecting the variability of the nematode community were revealed, i.e.: the variation in maturity, as determined by a set of quantified indices, and the ecological processes. Out of four amendments, the application of SS caused increasing in community maturity and change in selected ecological traits, whereas D had the opposite effect. The improvement in the soil nematode community caused by SS amendment indicated a trend toward a more stable and sustained soil ecosystem in marginal soil. Furthermore, since nematodes are suitable indicators for soil monitoring [4,69], it can be concluded that the application of SS resulted in healthier soil.
The ecological succession of nematodes is usually orderly and predictable unless set back by environmental disturbance such as cultivation, pollution or nutrient enrichment (incorporation of amendments and fertilisers) [4]. The impact of amendments on the nematode community and soil food web has been documented in several papers [47,70,71,72]. Following the addition of amendments, nematodes often undergo a successional transition with a high abundance of colonisers dominating disturbed or enriched habitats characterised by rapid reproduction [73]. In the current study, testing the short-term effects of soil amendments, maturity increase was associated with increasing nematode biomass with concomitant decreasing in total community abundance, which became more abundant for the fungal-feeder nematodes in the late decomposition stage. This funding is partly consistent with the suggestion that the impact of amendments on soil nematode abundance varies across application years: the short-term presence of organic amendments in the soil affects nematode function [74], whereas long-term application reduces total abundance and diversity [75]. Studies have shown that the long-term application of SS has no adverse effects on nematode abundance or diversity [70,76]. Nematode community abundance and structure tended to change with higher pollutant levels in SS [77]. However, total nematode abundance is a poor assessment tool for monitoring soil contamination and has to be modified by a variety of other factors [78].
The soil pollution [79] and agricultural practices such as tillage and fertilisation [4] disrupt the soil ecosystem and stimulate bacterial and fungal activities to which the nematode community responds by reducing MI [80]. Secondary succession leads to a reduction in trophic conditions and an increase in MI [66]. However, MI is only a simple indication of environmental disturbance. For a better understanding of the extent of disturbance and the dynamics of the restoration process of the soil ecosystem, it is necessary to integrate the functional guild and taxonomic data [81]. The use of PCA for a range of indices measuring the extent of ecosystem functions allows a comprehensive assessment of changes in the nematode community and functional diversity.
The enrichment of the soil ecosystem with nutrients leads to a shift in community dominance towards opportunistic coloniser species and returns the community to the earlier stages of successional dynamics [82]. MI and PPI are two indices that respond in opposite directions when soil nutrient status increases [83]. The larger the MI, the more the mature the soil ecosystem, and the lower is the degree of impact (PPI). Our results showed that MI and PPI indices changed coherently. This trait illustrated that the nematode community was influenced by enrichment of the soil with nutrients during application of amendments. Additionally, influence of changes in the physical properties of the environment, especially the air and water regimes of the soil, could not be excluded. However, this assumption requires further investigation.
For nematode communities living in the disturbed conditions, the ratio PPI/MI has a value of 1.25; for less disturbed communities it varies between 0.81 and 0.99 [84]. In the current study the ratio PPI/MI ranged between 1.18–1.26 for all experimental variants except SS, and only SS treatment had a range of 0.86 to 1.00. This indicated that the application of SS promoted the acceleration of succession dynamics of nematode community and stabilised the nutrient dynamics and the community itself. The results showed that fluctuations in community maturity were accompanied by changes in trophic structure. The proportions of PPNs, predators, and omnivores increased with maturity, while the proportion of fungivores and bacterivores decreased. The observed increasing in abundance of PPNs could be due to an increase in migratory endoparasites, which occurred simultaneously with a decrease in the proportion of ectoparasites.
When the community is more mature, the value of SI is higher, and values of BI and EI are lower. The growth of SI indicates the increasing complexity of the food web and the redundancy of the nematode community structure [85,86,87]. When soil is amended by SS, food resources are usually not limited and bacterial reproduction can be rapid and continuous. In such systems, nematode activity supports the active growth of bacterial populations and their distribution over the substrate [88]. In some cases, the application of SS increases nematode population densities even in highly contaminated soils [89,90], as some functional groups and taxa respond primarily to resource availability and have a higher tolerance to environmental stress.
PC2 captured common features of all amendments, stimulating the nematode community biomass, and this trend was strongest for HW treatment. The application of all amendments was characterised by enhancing the ecosystem service for disease and pest suppression. Under the influence of HW, the main trend in nematode community change was maturation due to the community’s ability to respond faster to environmental enrichment with nutrients [91].
In general, the nature of BDs’ effect was well described by PC2 and appeared to be similar to the effect of HW treatment. The influence of BDs on the community was not unidirectional: a trend was encountered of increasing the trophic structure of the community and its decreasing, illustrating the complex and multidirectional mechanisms of biochar influence on soil biota. Biochar affects soil acidity, which regulates the availability of nutrients to living organisms, ultimately leading to negative impacts on soil organisms [92]. The observed positive effect of biochar on crop yields may be due to improvement of water retention in the soil [93], which is more pronounced in nutrient-poor and acidic soils [94], as in our case the research soil had a pH equal to 4.7.
Biochar has a physical and chemical impact on the soil that initiates the dynamics of biological processes [95,96,97]. The microbial community changes with the incorporation of biochar [98,99], which affects the intensity of mineralisation of soil organic matter [100,101]. However, the medium-term field mesosome study provides evidence [102] that amendment by biochar does not affect the basic physicochemical and microbial soil properties, however, can change the nematode community: despite biochar does not affect total nematode abundance, taxa richness and total plant biomass, a taxonomic and functional shift in the nematode community is observed, influenced by biochar application rate (30 vs. 5 t ha−1). The low application rate promotes predatory and PPNs, while the high application rate favours bacterivores and fungivores. These published results [102] are partly consistent with the current study’s observation.
All tested amendments caused the suppression of PPNs, and following several studies [48,103,104,105], this fact demonstrated that amendments produced from waste and plant residues can control PPN populations. However, the incorporation of organic amendments into the soil does not necessarily reduce the number of PPNs [106,107,108], and even sometimes can increase it [109,110]. Chemical and biological mechanisms are responsible for the suppression of fungivores by soil amendments, such as the release of nematotoxic compounds originally presented in the incorporated materials, their generation during degradation in the soil, or the development of microbial antagonists of PPNs.
Our study showed that the application of different soil amendments on marginal land was likely to reduce the number of PPNs and provided pest control benefits. This is important funding in terms of growing concern that if the cultivation of energy crops such as M × g expands, the harboured PPNs may be able to cause crop damage [111]. The results ensured that the mechanisms affecting the soil biota under soil amendments and M × g cultivation are complex, multidirectional, and required long-term multi-year observation.
We did not observe any significant effect of amendments on M × g yield after the first vegetation (Table S5) which was not surprising, as M × g biomass is usually too low and economically invisible in the first 2–3 years of production; the intensity of biomass growth commonly accelerates in the subsequent years [18]. Simultaneously, during the first years, the plant can produce a considerable mass of underground organs: roots and short, vigorous rhizomes that can quickly reinforce the topsoil.
5. Conclusions
The impacts of different soil amendments, i.e., biochar in two doses (BD1 and BD2), biogas digestate (D), soil sludge (SS), and hemicellulose waste (HW) on the soil nematode community were studied during the first vegetation of M × g plantation on marginal, nutrient-poor land. While background soil nutrient concentrations were not determined, it was found that the soil nematode community associated with the cultivation of crop was sensitive to the type of applied organic amendments. The analysis of the nematode community in depth suggested that organic matter varies in the process of M × g cultivation and amendments had different influences on the soil nematode food web. It was revealed that out of four amendments tested, SS favoured a more stable maturity status of nematode community compared to the application of D, providing a snapshot of soil health. At the same time, impact to the nematode community on soil enrichment by BDs and HW remained inconclusive, and further multiyear monitoring of M × g cultivation in targeted amended soils is required.
To the best of our knowledge, this is the first report on how organic amendments incorporated into marginal soil affected the nematode community associated with M × g cultivation and the suppression activity of PPNs. Organic amendments can be considered as a potential tool for effective regulation of PPNs during long-term M × g cultivation. Further analysis of predatory/omnivorous and entomopathogenic nematodes would be very useful to understand whether organic amendments promote biological control organisms to maintain populations of PPNs and other members of the soil biota.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy12092063/s1, Table S1: Characteristics of amended soils; Table S2: Nematode community abundance and taxonomic diversity under amendments application and M × g growth; Table S3: Multivariate GLM results examining the effect of amendment type and sampling time on PC1–PC4 scores; Table S4: Response of given nematode taxa abundance to amendment type and sampling time from multivariate GLMs; Figure S1: Response of PC scores to the effects of amendments at different sampling time: Pi, Pf1, and Pf2; amendment type: BD1, BD2, D, SS, and HW. (a) PC1; (b) PC2; (c) PC3; Table S5: Parameters of M × g harvest value depending on the soil treatment.
Author Contributions
Conceptualization, T.S. and V.P.; methodology, T.S., A.S. and S.U.; software, O.Z. and A.M.; validation, A.S., D.K., R.A.N. and S.U.; formal analysis, T.S., A.S., V.P. and D.K.; investigation, A.S., D.K., A.M. and R.A.N.; resources, A.S. and V.P.; data curation, A.S., O.Z. and A.M.; writing—original draft preparation, T.S., V.P., A.S. and A.M.; writing—review and editing, T.S., V.P., A.S. and A.M.; visualization, O.Z. and A.M.; supervision, T.S. and V.P.; project administration, V.P.; funding acquisition, A.S. and V.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Czech-German CORNET project “MiscanValue”, Reg. No.CZ.01.1.02/0.0/0.0/19_263/0018837, co-financed by European Union from the European Regional Development Fund through the Operational Programmer Research, Development, and Education.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Anna Wesołowska, Nematology Department, National Horticulture Institute Skierniewice, Poland for the assistance during the nematode identification.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ansari, R.A.; Sumbul, A.; Rizvi, R.; Mahmood, I. Organic Soil Amendments: Potential Tool for Soil and Plant Health Management. In Plant Health Under Biotic Stress: Volume 1: Organic Strategies; Ansari, R.A., Mahmood, I., Eds.; Springer: Singapore, 2019; pp. 1–35. ISBN 9789811360435. [Google Scholar]
- Antonkiewicz, J.; Kołodziej, B.; Bielińska, E.J.; Popławska, A. The Possibility of Using Sewage Sludge for Energy Crop Cultivation Exemplified by Reed Canary Grass and Giant Miscanthus. Soil Sci. Annu. 2019, 70, 21–33. [Google Scholar] [CrossRef]
- Dubis, B.; Jankowski, K.J.; Załuski, D.; Sokólski, M. The Effect of Sewage Sludge Fertilization on the Biomass Yield of Giant Miscanthus and the Energy Balance of the Production Process. Energy 2020, 206, 118189. [Google Scholar] [CrossRef]
- Alasmary, Z.; Hettiarachchi, G.M.; Roozeboom, K.L.; Davis, L.C.; Erickson, L.E.; Pidlisnyuk, V.; Stefanovska, T.; Trögl, J. Phytostabilization of a Contaminated Military Site Using Miscanthus and Soil Amendments. J. Environ. Qual. 2021, 50, 1220–1232. [Google Scholar] [CrossRef] [PubMed]
- McCormack, S.A.; Ostle, N.; Bardgett, R.D.; Hopkins, D.W.; Vanbergen, A.J. Biochar in Bioenergy Cropping Systems: Impacts on Soil Faunal Communities and Linked Ecosystem Processes. GCB Bioenergy 2013, 5, 81–95. [Google Scholar] [CrossRef]
- Pidlisnyuk, V.; Newton, R.A.; Mamirova, A. Miscanthus Biochar Value Chain-A Review. J. Environ. Manag. 2021, 290, 112611. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bian, Q.; Jiang, Y.; Zhu, L.; Chen, Y.; Liang, Y.; Sun, B. Organic Amendments Drive Shifts in Microbial Community Structure and Keystone Taxa Which Increase C Mineralization across Aggregate Size Classes. Soil Biol. Biochem. 2021, 153, 108062. [Google Scholar] [CrossRef]
- Tan, Z.; Lin, C.S.K.; Ji, X.; Rainey, T.J. Returning Biochar to Fields: A Review. Appl. Soil Ecol. 2017, 116, 1–11. [Google Scholar] [CrossRef]
- Conte, P.; Bertani, R.; Sgarbossa, P.; Bambina, P.; Schmidt, H.-P.; Raga, R.; Lo Papa, G.; Chillura Martino, D.F.; Lo Meo, P. Recent Developments in Understanding Biochar’s Physical–Chemistry. Agronomy 2021, 11, 615. [Google Scholar] [CrossRef]
- Kavitha, B.; Reddy, P.V.L.; Kim, B.; Lee, S.S.; Pandey, S.K.; Kim, K.-H. Benefits and Limitations of Biochar Amendment in Agricultural Soils: A Review. J. Environ. Manag. 2018, 227, 146–154. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Wong, J.T.F.; Hashimoto, Y.; Huang, L.; Rinklebe, J.; Chang, S.X.; Bolan, N.; Wang, H.; Ok, Y.S. Response of Microbial Communities to Biochar-Amended Soils: A Critical Review. Biochar 2019, 1, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Krzyżaniak, M.; Stolarski, M.J.; Warmiński, K. Life Cycle Assessment of Giant Miscanthus: Production on Marginal Soil with Various Fertilisation Treatments. Energies 2020, 13, 1931. [Google Scholar] [CrossRef]
- Winkler, B.; Mangold, A.; von Cossel, M.; Clifton-Brown, J.; Pogrzeba, M.; Lewandowski, I.; Iqbal, Y.; Kiesel, A. Implementing Miscanthus into Farming Systems: A Review of Agronomic Practices, Capital and Labour Demand. Renew. Sustain. Energy Rev. 2020, 132, 110053. [Google Scholar] [CrossRef]
- Ziadi, N.; Gagnon, B.; Nyiraneza, J. Crop Yield and Soil Fertility as Affected by Papermill Biosolids and Liming By-Products. Can. J. Soil Sci. 2013, 93, 319–328. [Google Scholar] [CrossRef]
- Mozaffari, M.; Hays, H.C. Effect of a Newly Developed Pelleted Papermill Biosolids on Crop and Soil. J. Agric. Chem. Environ. 2019, 9, 1. [Google Scholar] [CrossRef]
- Dalzell, C.G. Purpose-Grown Biomass Crops: Efficient Production and Real-World Verification. Master’s Thesis, Saint Mary’s University, Halifax, NS, Canada, 2021. [Google Scholar]
- Kowalska, A.; Grobelak, A.; Almås, Å.R.; Singh, B.R. Effect of Biowastes on Soil Remediation, Plant Productivity and Soil Organic Carbon Sequestration: A Review. Energies 2020, 13, 5813. [Google Scholar] [CrossRef]
- Kvak, V.; Stefanovska, T.; Pidlisnyuk, V.; Alasmary, Z.; Kharytonov, M. The Long-Term Assessment of Miscanthus × gigantheus Cultivation in the Forest-Steppe Zone of Ukraine. INMATEH-Agric. Eng. 2018, 54, 113–120. [Google Scholar]
- Wang, C.; Kong, Y.; Hu, R.; Zhou, G. Miscanthus: A Fast-Growing Crop for Environmental Remediation and Biofuel Production. GCB Bioenergy 2021, 13, 58–69. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Scordia, D.; Testa, G.; Monti, A.; Alexopoulou, E.; Christou, M. 1-The Importance of Perennial Grasses as a Feedstock for Bioenergy and Bioproducts. In Perennial Grasses for Bioenergy and Bioproducts; Alexopoulou, E., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1–33. ISBN 978-0-12-812900-5. [Google Scholar]
- Pidlisnyuk, V.; Shapoval, P.; Zgorelec, Ž.; Stefanovska, T.; Zhukov, O. Multiyear Phytoremediation and Dynamic of Foliar Metal(Loid)s Concentration during Application of Miscanthus × giganteus Greef et Deu to Polluted Soil from Bakar, Croatia. Environ. Sci. Pollut. Res. 2020, 27, 31446–31457. [Google Scholar] [CrossRef]
- Pidlisnyuk, V.; Herts, A.; Khomenchuk, V.; Mamirova, A.; Kononchuk, O.; Ust’ak, S. Dynamic of Morphological and Physiological Parameters and Variation of Soil Characteristics during Miscanthus × giganteus Cultivation in the Diesel-Contaminated Land. Agronomy 2021, 11, 798. [Google Scholar] [CrossRef]
- Al Souki, K.S.; Burdová, H.; Trubač, J.; Štojdl, J.; Kuráň, P.; Kříženecká, S.; Machová, I.; Kubát, K.; Popelka, J.; Malinská, H.A.; et al. Enhanced Carbon Sequestration in Marginal Land Upon Shift towards Perennial C4 Miscanthus × giganteus: A Case Study in North-Western Czechia. Agronomy 2021, 11, 293. [Google Scholar] [CrossRef]
- Neher, D.A. Role of Nematodes in Soil Health and Their Use as Indicators. J. Nematol. 2001, 33, 161–168. [Google Scholar] [PubMed]
- Liu, X.; Zhang, D.; Li, H.; Qi, X.; Gao, Y.; Zhang, Y.; Han, Y.; Jiang, Y.; Li, H. Soil Nematode Community and Crop Productivity in Response to 5-Year Biochar and Manure Addition to Yellow Cinnamon Soil. BMC Ecol. 2020, 20, 39. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Bongers, T. Nematode Indicators of Organic Enrichment. J. Nematol. 2006, 38, 3–12. [Google Scholar] [PubMed]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A Framework for Soil Food Web Diagnostics: Extension of the Nematode Faunal Analysis Concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Holajjer, P.; Kamra, A.; Singh, P. Influence of Nematode-Bacterial Interactions on N and P Mineralisation in Soil and on Decomposition of Crop Residues during Aerobic Composting. Appl. Ecol. Environ. Res. 2016, 14, 283–299. [Google Scholar] [CrossRef]
- Trap, J.; Bonkowski, M.; Plassard, C.; Villenave, C.; Blanchart, E. Ecological Importance of Soil Bacterivores for Ecosystem Functions. Plant Soil 2016, 398, 1–24. [Google Scholar] [CrossRef]
- Jiang, Y.; Qian, H.; Wang, X.; Chen, L.; Liu, M.; Li, H.; Sun, B. Nematodes and Microbial Community Affect the Sizes and Turnover Rates of Organic Carbon Pools in Soil Aggregates. Soil Biol. Biochem. 2018, 119, 22–31. [Google Scholar] [CrossRef]
- Jagodič, A.; Trdan, S.; Laznik, Ž. Entomopathogenic Nematodes: Can We Use the Current Knowledge on Belowground Multitrophic Interactions in Future Plant Protection Programmes?–Review. Plant Prot. Sci. 2019, 55, 243–254. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding Habits in Soil Nematode Families and Genera—An Outline for Soil Ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- McSorley, R. Overview of Organic Amendments for Management of Plant-Parasitic Nematodes, with Case Studies from Florida. J. Nematol. 2011, 43, 69–81. [Google Scholar]
- Herren, G.L.; Habraken, J.; Waeyenberge, L.; Haegeman, A.; Viaene, N.; Cougnon, M.; Reheul, D.; Steel, H.; Bert, W. Effects of Synthetic Fertilizer and Farm Compost on Soil Nematode Community in Long-Term Crop Rotation Plots: A Morphological and Metabarcoding Approach. PLoS ONE 2020, 15, e0230153. [Google Scholar] [CrossRef]
- Lazarova, S.; Coyne, D.; Rodríguez, M.G.; Peteira, B.; Ciancio, A. Functional Diversity of Soil Nematodes in Relation to the Impact of Agriculture—A Review. Diversity 2021, 13, 64. [Google Scholar] [CrossRef]
- Nguyen, S.V.; Nguyen, P.T.K.; Araki, M.; Perry, R.N.; Ba Tran, L.; Minh Chau, K.; Min, Y.Y.; Toyota, K. Effects of Cropping Systems and Soil Amendments on Nematode Community and Its Relationship with Soil Physicochemical Properties in a Paddy Rice Field in the Vietnamese Mekong Delta. Appl. Soil Ecol. 2020, 156, 103683. [Google Scholar] [CrossRef]
- Pulavarty, A.; Egan, A.; Karpinska, A.; Horgan, K.; Kakouli-Duarte, T. Plant Parasitic Nematodes: A Review on Their Behaviour, Host Interaction, Management Approaches and Their Occurrence in Two Sites in the Republic of Ireland. Plants 2021, 10, 2352. [Google Scholar] [CrossRef]
- Zasada, I.A.; Halbrendt, J.M.; Kokalis-Burelle, N.; LaMondia, J.; McKenry, M.V.; Noling, J.W. Managing Nematodes without Methyl Bromide. Annu. Rev. Phytopathol. 2010, 48, 311–328. [Google Scholar] [CrossRef]
- Rosskopf, E.; Di Gioia, F.; Hong, J.C.; Pisani, C.; Kokalis-Burelle, N. Organic Amendments for Pathogen and Nematode Control. Annu. Rev. Phytopathol. 2020, 58, 277–311. [Google Scholar] [CrossRef]
- Mennan, S.; Melakeberhan, H. Effects of Biosolid Amendment on Populations of Meloidogyne Hapla and Soils with Different Textures and pHs. Bioresour. Technol. 2010, 101, 7158–7164. [Google Scholar] [CrossRef]
- Zasada, I.; Rogers, S.; Sardanelli, S. Application of Alkaline-Stabilised Biosolids for Meloidogyne Incognita Suppression in Microplots. Nematology 2007, 9, 123–129. [Google Scholar] [CrossRef]
- Zasada, I.A.; Avendano, F.; Li, Y.C.; Logan, T.; Melakeberhan, H.; Koenning, S.R.; Tylka, G.L. Potential of an Alkaline-Stabilized Biosolid to Manage Nematodes: Case Studies on Soybean Cyst and Root-Knot Nematodes. Plant Dis. 2008, 92, 4–13. [Google Scholar] [CrossRef]
- Yeates, G.W.; Speir, T.W.; Taylor, M.D.; Clucas, L.; Schaik, A.P. Nematode Responses to Biosolids Incorporation in Five Soil Types. Biol. Fertil. Soils 2006, 42, 550–555. [Google Scholar] [CrossRef]
- Zhang, X.-K.; Li, Q.; Liang, W.-J.; Zhang, M.; Bao, X.-L.; Xie, Z.-B. Soil Nematode Response to Biochar Addition in a Chinese Wheat Field. Pedosphere 2013, 23, 98–103. [Google Scholar] [CrossRef]
- Wanjiku Kamau, J.; Biber-Freudenberger, L.; Lamers, J.P.A.; Stellmacher, T.; Borgemeister, C. Soil Fertility and Biodiversity on Organic and Conventional Smallholder Farms in Kenya. Appl. Soil Ecol. 2019, 134, 85–97. [Google Scholar] [CrossRef]
- George, C.; Kohler, J.; Rillig, M.C. Biochars Reduce Infection Rates of the Root-Lesion Nematode Pratylenchus penetrans and Associated Biomass Loss in Carrot. Soil Biol. Biochem. 2016, 95, 11–18. [Google Scholar] [CrossRef]
- Liu, T.; Chen, X.; Hu, F.; Ran, W.; Shen, Q.; Li, H.; Whalen, J.K. Carbon-Rich Organic Fertilizers to Increase Soil Biodiversity: Evidence from a Meta-Analysis of Nematode Communities. Agric. Ecosyst. Environ. 2016, 232, 199–207. [Google Scholar] [CrossRef]
- Oka, Y. Mechanisms of Nematode Suppression by Organic Soil Amendments—A Review. Appl. Soil Ecol. 2010, 44, 101–115. [Google Scholar] [CrossRef]
- Huang, W.; Ji, H.; Gheysen, G.; Debode, J.; Kyndt, T. Biochar-Amended Potting Medium Reduces the Susceptibility of Rice to Root-Knot Nematode Infections. BMC Plant Biol. 2015, 15, 267. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, N.; Viaene, N.; Vandecasteele, B.; D’Hose, T.; Debode, J.; Cremelie, P.; De Tender, C.; Moens, M. Traditional and New Soil Amendments Reduce Survival and Reproduction of Potato Cyst Nematodes, except for Biochar. Appl. Soil Ecol. 2016, 107, 191–204. [Google Scholar] [CrossRef]
- Harel, Y.M.; Kolton, M.; Elad, Y.; Rav-David, D.; Cytryn, E.; Borenshtein, M.; Shulchani, R.; Graber, E.R. Biochar Impact on Plant Development and Disease Resistance in Pot Trials. IOBCWPRS Bull 2012, 78, 141–147. [Google Scholar]
- Marra, R.; Vinale, F.; Cesarano, G.; Lombardi, N.; d’Errico, G.; Crasto, A.; Mazzei, P.; Piccolo, A.; Incerti, G.; Woo, S.L.; et al. Biochars from Olive Mill Waste Have Contrasting Effects on Plants, Fungi and Phytoparasitic Nematodes. PLoS ONE 2018, 13, e0198728. [Google Scholar] [CrossRef]
- Huebner, R.A.; Rodriguez-Kabana, R.; Patterson, R.M. Hemicellulosic Waste and Urea for Control of Plant Parasitic Nematodes: Effect on Soil Enzyme Activities. Nematropica 1983, 13, 37–54. [Google Scholar]
- Min, Y.Y.; Toyota, K.; Sato, E.; Takada, A. Effects of Anaerobically Digested Slurry on Meloidogyne incognita and Pratylenchus penetrans in Tomato and Radish Production. Appl. Environ. Soil Sci. 2011, 2011, e528712. [Google Scholar] [CrossRef]
- DSTU 4287:2004. Soil Quality. Sampling. DP “UkrNDNC”: Kyiv, Ukraine, 2005; p. 9.
- DSTU:ISO:10390:2001. Soil Quality. Determination of pH. DP “UkrNDNC”: Kyiv, Ukraine, 2002; p. 10.
- 275/1998 Sb. Release of the Ministry of Agriculture Related the Agrochemical Testing of the Soils and Evaluation the Land Properties of the Forest Land (with Amendments 335/2017 Sb). Ministry of Agriculture: Nové Město, Czech Republic, 2017.
- Němeček, J.; Mühlhanselová, M.; Macků, J.; Vokoun, J.; Vavříček, D.; Novák, P. Czech Taxonomic Soil Classification System, 2nd ed.; Czech University of Life Sciences in Prague: Prague, Czech Republic, 2011; ISBN 978-80-213-2155-7. [Google Scholar]
- DSTU:7632-2014; Soils. Method for Determination of Organic Matter. DP “UkrNDNC”: Kyiv, Ukraine, 2014; p. 15.
- DSTU:4115:2002; Soils. Determination of Mobile Phosphorus and Potassium Compounds by the Modified Chirikov’s Method. DP “UkrNDNC”: Kyiv, Ukraine, 2003; p. 12.
- DSTU:7861:2015; Soil Quality. Determination of Exchanges Calcium, Magnesium, Sodium and Potassium in Soil according to Shollenberger in NSC ISSAR Named after Sokolovsky Modification. DP “UkrNDNC”: Kyiv, Ukraine, 2016; p. 12.
- Szczygieł, A. Application of the Centrifugal Method for Extraction of Nematodes from Soil. J. Prog. Agric. Sci. 1971, 12, 169–179. [Google Scholar]
- Seinhorst, J.W. Killing Nematodes for Taxonomic Study with Hot f.a. 4:1. Nematologica 1966, 12, 178–178a. [Google Scholar] [CrossRef]
- Brzeski, M.W. Nematodes of Tylenchina in Poland and Temperate Europe; Muzeum i Instytutu Zoologii, Polska Akademia Nauk (MiIZ PAN): Warsaw, Poland, 1998; ISBN 83-85192-84-0. [Google Scholar]
- Andrássy, I. Free-Living Nematodes of Hungary (Nematoda Errantia). In Pedozoologica Hungarica; Csuzdi, C., Mahunka, S., Eds.; Hungarian Natural History Museum and Systematic Zoology Research Group of the Hungarian Academy of Sciences: Budapest, Hungary, 2007; Volume II, ISBN 963. [Google Scholar]
- Bongers, T. The Maturity Index: An Ecological Measure of Environmental Disturbance Based on Nematode Species Composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G.M. NINJA: An Automated Calculation System for Nematode-Based Biological Monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Prentice, I.C. A Theory of Gradient Analysis. In Advances in Ecological Research; Begon, M., Fitter, A.H., Ford, E.D., Macfadyen, A., Eds.; Academic Press: Cambridge, MA, USA, 1988; Volume 18, pp. 271–317. [Google Scholar]
- Stefanovska, T.; Skwiercz, A.; Zouhar, M.; Pidlisnyuk, V.; Zhukov, O. Plant-Feeding Nematodes Associated with Miscanthus × giganteus and Their Use as Potential Indicators of the Plantations’ State. Int. J. Environ. Sci. Technol. 2021, 18, 57–72. [Google Scholar] [CrossRef]
- Tabarant, P.; Villenave, C.; Risede, J.-M.; Roger-Estrade, J.; Thuries, L.; Dorel, M. Effects of Four Organic Amendments on Banana Parasitic Nematodes and Soil Nematode Communities. Appl. Soil Ecol. 2011, 49, 59–67. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhou, H.; Chen, L.; Yuan, Y.; Fang, H.; Luan, L.; Chen, Y.; Wang, X.; Liu, M.; Li, H.; et al. Nematodes and Microorganisms Interactively Stimulate Soil Organic Carbon Turnover in the Macroaggregates. Front. Microbiol. 2018, 9, 2803. [Google Scholar] [CrossRef]
- Liang, W.; Lou, Y.; Li, Q.; Zhong, S.; Zhang, X.; Wang, J. Nematode Faunal Response to Long-Term Application of Nitrogen Fertilizer and Organic Manure in Northeast China. Soil Biol. Biochem. 2009, 41, 883–890. [Google Scholar] [CrossRef]
- Bouwman, L.A.; Zwart, K.B. The Ecology of Bacterivorous protozoans and Nematodes in Arable Soil. Agric. Ecosyst. Environ. 1994, 51, 145–160. [Google Scholar] [CrossRef]
- Pan, F.; Han, X.; Li, N.; Yan, J.; Xu, Y. Effect of Organic Amendment Amount on Soil Nematode Community Structure and Metabolic Footprints in Soybean Phase of a Soybean-Maize Rotation on Mollisols. Pedosphere 2020, 30, 544–554. [Google Scholar] [CrossRef]
- Li, J.; Wang, D.; Fan, W.; He, R.; Yao, Y.; Sun, L.; Zhao, X.; Wu, J. Comparative Effects of Different Organic Materials on Nematode Community in Continuous Soybean Monoculture Soil. Appl. Soil Ecol. 2018, 125, 12–17. [Google Scholar] [CrossRef]
- Nieminen, J.K.; Räisänen, M. Effects of Sewage Sludge Addition to Norway Spruce Seedlings on Nitrogen Availability and Soil Fauna in Clear-Cut Areas. Environ. Pollut. 2013, 178, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Creamer, R.E.; Rimmer, D.L.; Black, H.I.J. Do Elevated Soil Concentrations of Metals Affect the Diversity and Activity of Soil Invertebrates in the Long-Term? Soil Use Manag. 2008, 24, 37–46. [Google Scholar] [CrossRef]
- Kapusta, P.; Szarek-Łukaszewska, G.; Stefanowicz, A.M. Direct and Indirect Effects of Metal Contamination on Soil Biota in a Zn–Pb Post-Mining and Smelting Area (S Poland). Environ. Pollut. 2011, 159, 1516–1522. [Google Scholar] [CrossRef]
- Chauvin, C.; Trambolho, M.; Hedde, M.; Makowski, D.; Cérémonie, H.; Jimenez, A.; Villenave, C. Soil Nematodes as Indicators of Heavy Metal Pollution: A Meta-Analysis. Open J. Soil Sci. 2020, 10, 579–601. [Google Scholar] [CrossRef]
- Gupta, D.; Bhandari, S.; Bhusal, D.R. Variation of Nematode Indices under Contrasting Pest Management Practices in a Tomato Growing Agro-Ecosystem. Heliyon 2019, 5, e02621. [Google Scholar] [CrossRef]
- Ortiz, V.; Phelan, S.; Mullins, E. A Temporal Assessment of Nematode Community Structure and Diversity in the Rhizosphere of Cisgenic phytophthora Infestans-Resistant Potatoes. BMC Ecol. 2016, 16, 55. [Google Scholar] [CrossRef]
- Odum, E.P. Trends Expected in Stressed Ecosystems. BioScience 1985, 35, 419–422. [Google Scholar] [CrossRef]
- Bongers, T.; van der Meulen, H.; Korthals, G. Inverse Relationship between the Nematode Maturity Index and Plant Parasite Index under Enriched Nutrient Conditions. Appl. Soil Ecol. 1997, 6, 195–199. [Google Scholar] [CrossRef]
- Čermák, V.; Gaar, V.; Háněl, L.; Široká, K. Composition and Vertical Distribution of Free Living and Plant Parasitic Nematodes in Hop Gardens in the Czech Republic. Helminthologia 2011, 48, 124–136. [Google Scholar] [CrossRef]
- Peralta, G.; Schon, N.L.; Dickie, I.A.; St. John, M.G.; Orwin, K.H.; Yeates, G.W.; Peltzer, D.A. Contrasting Responses of Soil Nematode Communities to Native and Non-Native Woody Plant Expansion. Oecologia 2019, 190, 891–899. [Google Scholar] [CrossRef]
- Ney, L.; Franklin, D.; Mahmud, K.; Cabrera, M.; Hancock, D.; Habteselassie, M.; Newcomer, Q.; Dahal, S.; Subedi, A. Sensitivity of Nematode Community Analysis to Agricultural Management Practices and Inoculation with Local Effective Microorganisms in the Southeastern United States. Soil Syst. 2019, 3, 41. [Google Scholar] [CrossRef]
- Wang, Y.M.; Guan, P.T.; Chen, J.W.; Li, Z.X.; Yang, Y.R.; Wang, P. A Comparison of Soil Nematode Community Structure and Environmental Factors along Fen-Bush-Forest Succession in a Peatland, Northeastern China. Glob. Ecol. Conserv. 2021, 28, e01679. [Google Scholar] [CrossRef]
- Abrams, B.I.; Mitchell, M.J. Role of Nematode-Bacterial Interactions in Heterotrophic Systems with Emphasis on Sewage Sludge Decomposition. Oikos 1980, 35, 404–410. [Google Scholar] [CrossRef]
- Tejada, M. Application of Different Organic Wastes in a Soil Polluted by Cadmium: Effects on Soil Biological Properties. Geoderma 2009, 153, 254–268. [Google Scholar] [CrossRef]
- Martinez, J.G.; dos Santos, G.; Derycke, S.; Moens, T. Effects of Cadmium on the Fitness of, and Interactions between, Two Bacterivorous Nematode Species. Appl. Soil Ecol. 2012, 56, 10–18. [Google Scholar] [CrossRef]
- Ferris, H. Form and Function: Metabolic Footprints of Nematodes in the Soil Food Web. Eur. J. Soil Biol. 2010, 46, 97–104. [Google Scholar] [CrossRef]
- Brtnicky, M.; Datta, R.; Holatko, J.; Bielska, L.; Gusiatin, Z.M.; Kucerik, J.; Hammerschmiedt, T.; Danish, S.; Radziemska, M.; Mravcova, L.; et al. A Critical Review of the Possible Adverse Effects of Biochar in the Soil Environment. Sci. Total Environ. 2021, 796, 148756. [Google Scholar] [CrossRef]
- Jeffery, S.; Verheijen, F.G.A.; van der Velde, M.; Bastos, A.C. A Quantitative Review of the Effects of Biochar Application to Soils on Crop Productivity Using Meta-Analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Jeffery, S.; Abalos, D.; Prodana, M.; Bastos, A.C.; van Groenigen, J.W.; Hungate, B.A.; Verheijen, F. Biochar Boosts Tropical but Not Temperate Crop Yields. Environ. Res. Lett. 2017, 12, 053001. [Google Scholar] [CrossRef]
- Anderson, C.R.; Condron, L.M.; Clough, T.J.; Fiers, M.; Stewart, A.; Hill, R.A.; Sherlock, R.R. Biochar Induced Soil Microbial Community Change: Implications for Biogeochemical Cycling of Carbon, Nitrogen and Phosphorus. Pedobiologia 2011, 54, 309–320. [Google Scholar] [CrossRef]
- Fox, A.; Kwapinski, W.; Griffiths, B.S.; Schmalenberger, A. The Role of Sulfur- and Phosphorus-Mobilizing Bacteria in Biochar-Induced Growth Promotion of Lolium perenne. FEMS Microbiol. Ecol. 2014, 90, 78–91. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar Effects on Soil Biota–A Review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Watzinger, A.; Feichtmair, S.; Kitzler, B.; Zehetner, F.; Kloss, S.; Wimmer, B.; Zechmeister-Boltenstern, S.; Soja, G. Soil Microbial Communities Responded to Biochar Application in Temperate Soils and Slowly Metabolized 13C-Labelled Biochar as Revealed by 13C PLFA Analyses: Results from a Short-Term Incubation and Pot Experiment. Eur. J. Soil Sci. 2014, 65, 40–51. [Google Scholar] [CrossRef]
- Azeem, M.; Hale, L.; Montgomery, J.; Crowley, D.; McGiffen, M.E., Jr. Biochar and Compost Effects on Soil Microbial Communities and Nitrogen Induced Respiration in Turfgrass Soils. PLoS ONE 2020, 15, e0242209. [Google Scholar] [CrossRef]
- Bruun, S.; EL-Zehery, T. Biochar Effect on the Mineralization of Soil Organic Matter. Pesqui. Agropecuária Bras. 2012, 47, 665–671. [Google Scholar] [CrossRef]
- Tsai, C.-C.; Chang, Y.-F. Kinetics of C Mineralization of Biochars in Three Excessive Compost-Fertilized Soils: Effects of Feedstocks and Soil Properties. Agronomy 2020, 10, 1749. [Google Scholar] [CrossRef]
- Domene, X.; Mattana, S.; Sánchez-Moreno, S. Biochar Addition Rate Determines Contrasting Shifts in Soil Nematode Trophic Groups in Outdoor Mesocosms: An Appraisal of Underlying Mechanisms. Appl. Soil Ecol. 2021, 158, 103788. [Google Scholar] [CrossRef]
- Cole, E.J.; Barker, A.V.; Zandvakili, O.R.; Sadeghpour, A.; Xing, B.; Hashemi, M.; Allan-Perkins, E.; Jung, G. Soil Nutrient and Nematode Community Changes in Response to Hardwood Charcoal Application. Commun. Soil Sci. Plant Anal. 2021, 52, 917–925. [Google Scholar] [CrossRef]
- Akhtar, M.; Malik, A. Roles of Organic Soil Amendments and Soil Organisms in the Biological Control of Plant-Parasitic Nematodes: A Review. Bioresour. Technol. 2000, 74, 35–47. [Google Scholar] [CrossRef]
- Litterick, A.M.; Harrier, L.; Wallace, P.; Watson, C.A.; Wood, M. The Role of Uncomposted Materials, Composts, Manures, and Compost Extracts in Reducing Pest and Disease Incidence and Severity in Sustainable Temperate Agricultural and Horticultural Crop Production—A Review. Crit. Rev. Plant Sci. 2004, 23, 453–479. [Google Scholar] [CrossRef]
- Jaffee, B.A.; Ferris, H.; Scow, K.M. Nematode-Trapping Fungi in Organic and Conventional Cropping Systems. Phytopathology 1998, 88, 344–350. [Google Scholar] [CrossRef]
- Porazinska, D.L.; Duncan, L.W.; McSorley, R.; Graham, J.H. Nematode Communities as Indicators of Status and Processes of a Soil Ecosystem Influenced by Agricultural Management Practices. Appl. Soil Ecol. 1999, 13, 69–86. [Google Scholar] [CrossRef]
- McSorley, R.; Gallaher, R. Cultural Practices Improve Crop Tolerance to Nematodes. Nematropica 1995, 25, 53–60. [Google Scholar]
- Kimpinski, J.; Gallant, C.E.; Henry, R.; Macleod, J.A.; Sanderson, J.B.; Sturz, A.V. Effect of Compost and Manure Soil Amendments on Nematodes and on Yields of Potato and Barley: A 7-Year Study. J. Nematol. 2003, 35, 289–293. [Google Scholar]
- Bélair, G.; Tremblay, N. The Influence of Chitin-Urea Amendments Applied to an Organic Soil on a Meloidogyne hapla Population and on the Growth of Greenhouse Tomato. Phytoprotection 1995, 76, 75–80. [Google Scholar] [CrossRef]
- Stefanovska, T.; Skwiercz, A.; Flis, Ł.; Pidlisnyuk, V.; Zouhar, M. First Record of the Ectoparasitic Nematode Amplimerlinius macrurus (Nematoda: Tylenchida) on the Perennial Grass Miscanthus × giganteus (Angiosperms: Poaceae) in Ukraine. J. Nematol. 2021, 53, e2021-24. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).