

Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design and Data Collection
2.3. Data Analyses
3. Results
3.1. Species Richness of the Seedling Bank
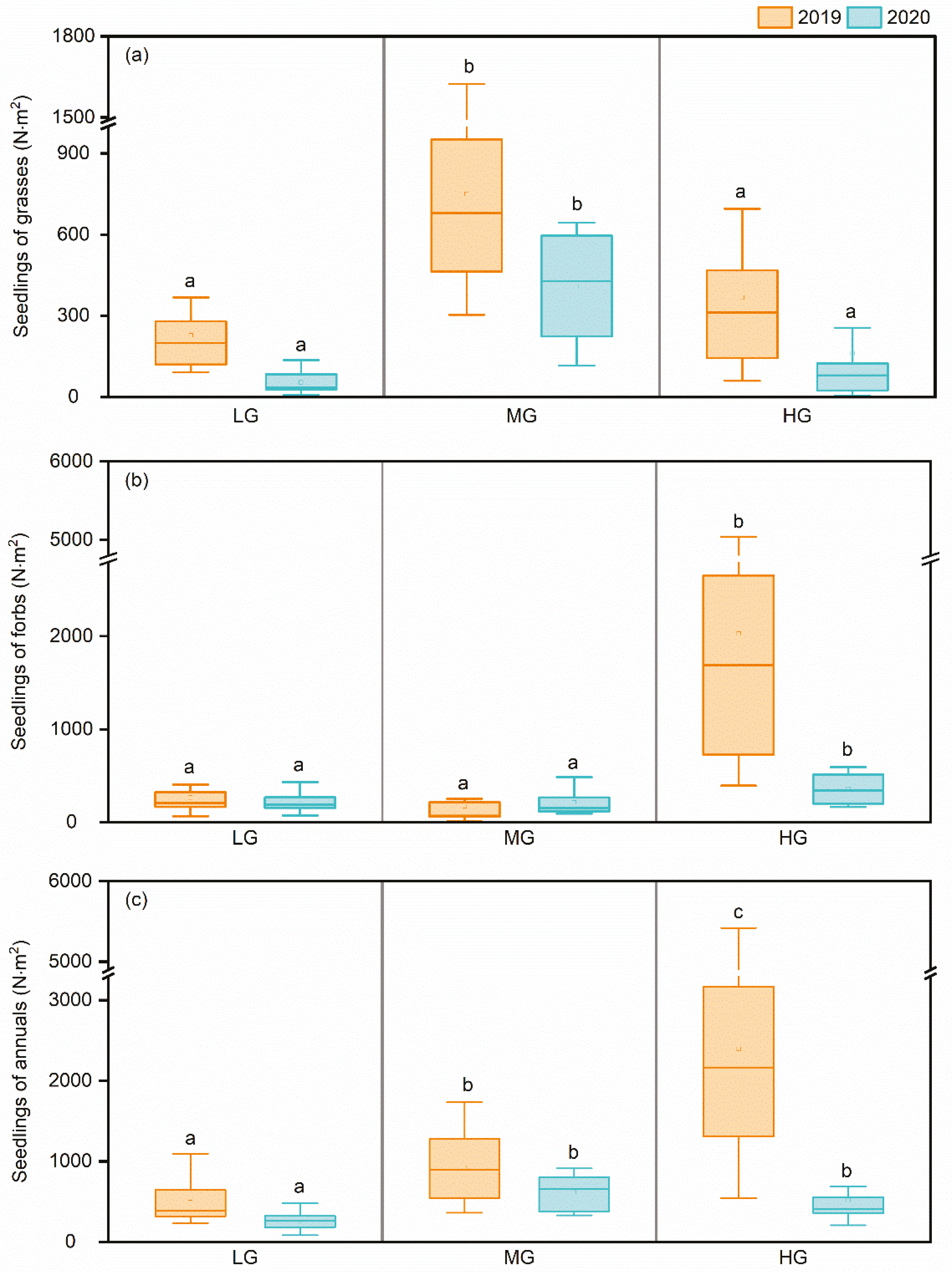
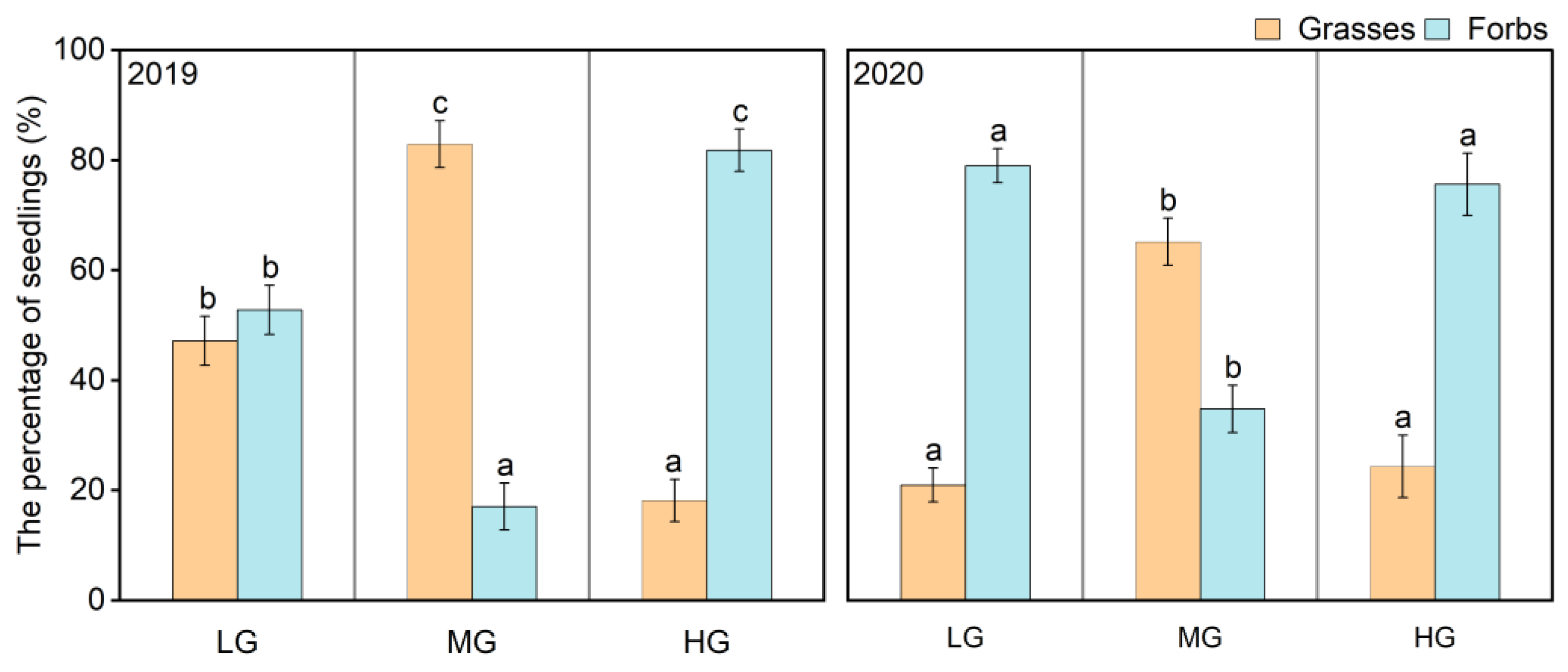
3.2. Abundance and Functional Composition of the Seedling Bank
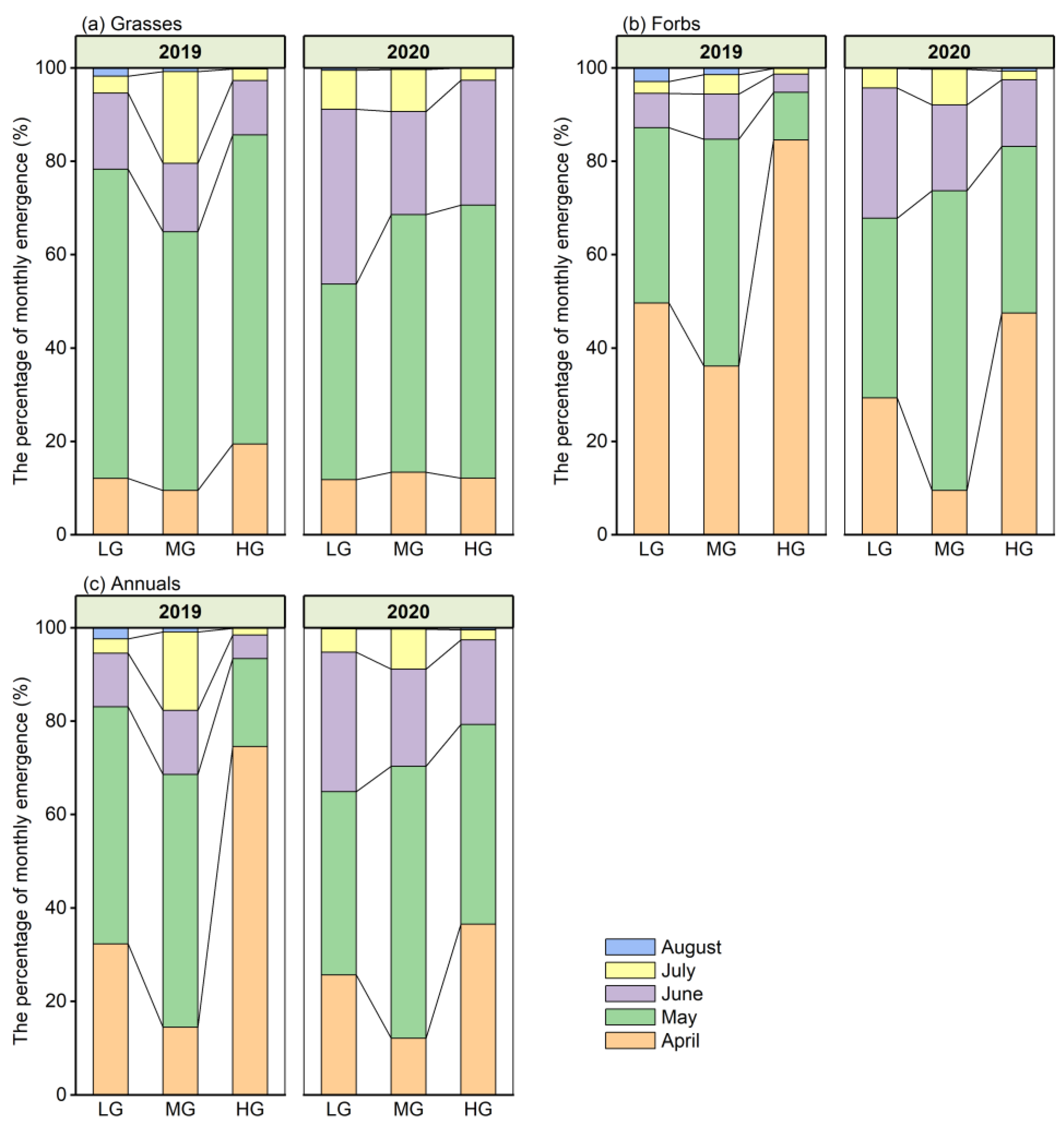
3.3. Seedling Emergence Dynamics
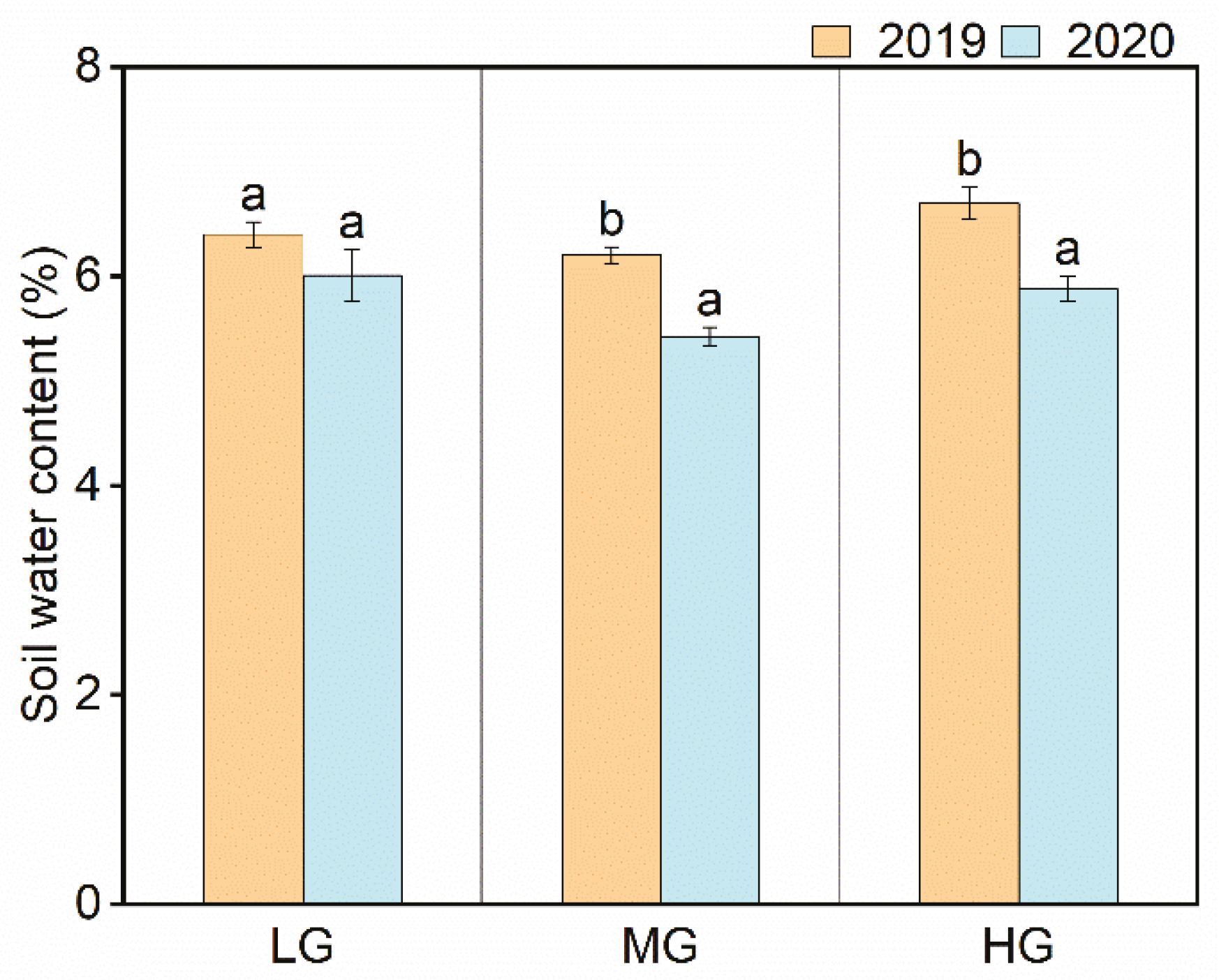
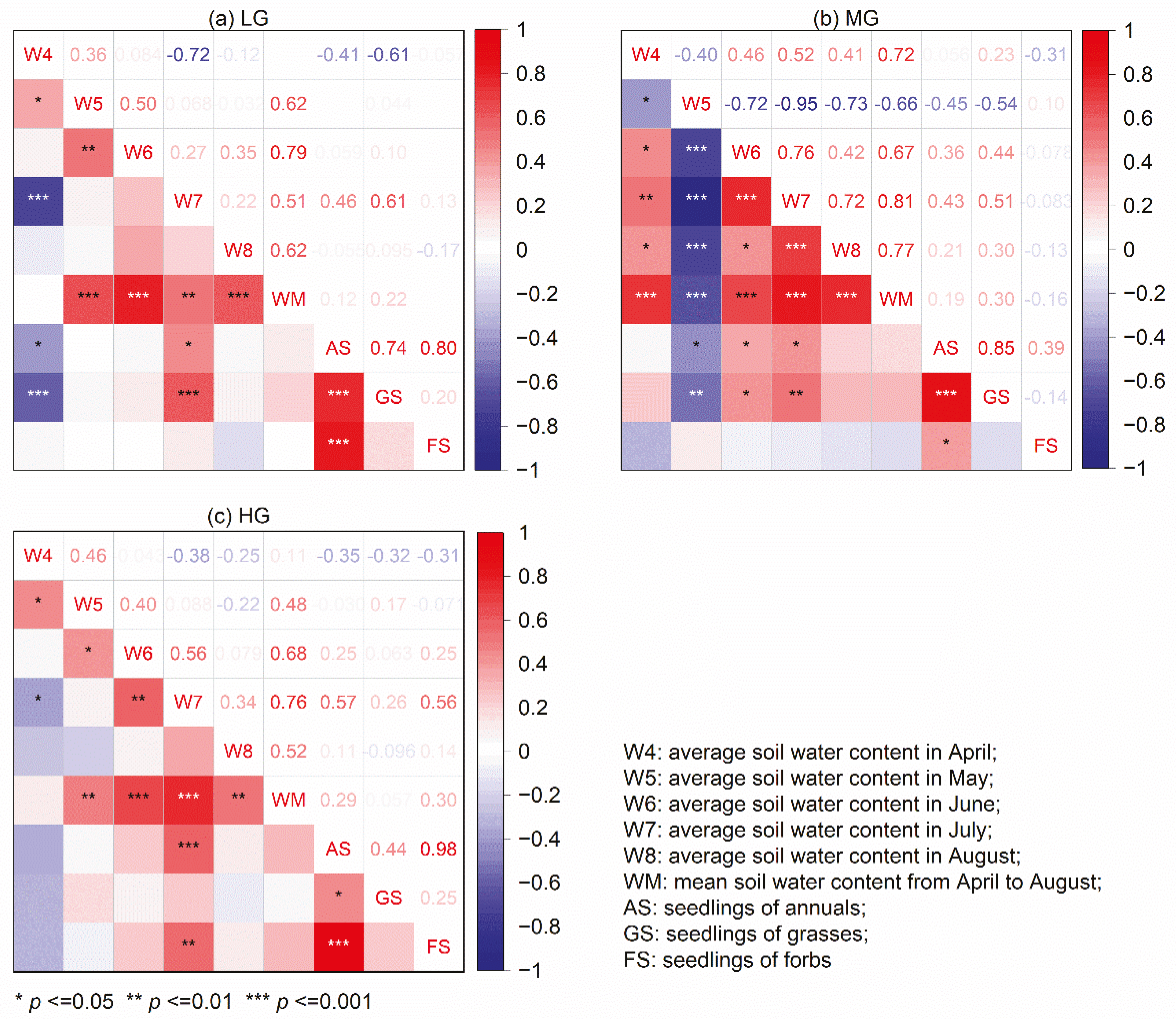
3.4. Correlation between Seedling Emergence and Soil Water Content
4. Discussion
4.1. Effects of Grazing on Species Richness of Seedlings in Wet and Dry Years
4.2. Effects of Grazing on Seedling Abundance of the Seedling Bank in Wet and Dry Years
4.3. Grazing Effects on Emergence Dynamics in Wet and Dry Years
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, H.; Jin, B.C.; Luo, K.; Pei, J.Y.; Zhang, X.L.; Zhang, Y.H.; Tang, J.Q.; Yang, Q.; Sun, G.J. Vegetation response to goats grazing intensity in semiarid hilly grassland of the Loess Plateau. Sustainability 2021, 13, 3569. [Google Scholar] [CrossRef]
- Pfeiffer, M.; Langan, L.; Linstadter, A.; Martens, C.; Gaillard, C.; Ruppert, J.C.; Higgins, S.I.; Mudongo, E.I.; Scheiter, S. Grazing and aridity reduce perennial grass abundance in semi-arid rangelands-Insights from a trait-based dynamic vegetation model. Ecol. Model. 2019, 395, 11–22. [Google Scholar] [CrossRef]
- Zhao, H.L.; Zhang, T.H.; Zhao, X.Y.; Zhou, R.L. Effects of sheep grazing and precipitation patterns on sandy grassland vegetation in Inner Mongolia, China. Environ. Sci. Develop. 2011, 4, 68–75. [Google Scholar]
- Zainelabdeen, Y.M.; Yan, R.; Xin, X.; Yan, Y.; Ahmed, A.I.; Hou, L.; Zhang, Y. The Impact of Grazing on the Grass Composition in Temperate Grassland. Agronomy 2020, 10, 1230. [Google Scholar] [CrossRef]
- Bat-Oyun, T.; Shinoda, M.; Cheng, Y.X.; Purevdorj, Y. Effects of grazing and precipitation variability on vegetation dynamics in a Mongolian dry steppe. J. Plant Ecol. 2016, 9, 508–519. [Google Scholar] [CrossRef]
- Wan, H.; Bai, Y.; Hooper, D.U.; Schönbach, P.; Gierus, M.; Schiborra, A.; Taube, F. Selective grazing and seasonal precipitation play key roles in shaping plant community structure of semi-arid grasslands. Landsc. Ecol. 2015, 30, 1767–1782. [Google Scholar] [CrossRef]
- Huang, X.T.; Luo, G.P.; Chen, C.B.; Peng, J.; Zhang, C.J.; Zhou, H.K.; Yao, B.Q.; Ma, Z.; Xi, X.Y. How precipitation and grazing influence the ecological functions of drought-prone grasslands on the northern slopes of the Tianshan Mountains, China? J. Arid Land 2021, 13, 88–97. [Google Scholar] [CrossRef]
- Ren, H.; Schönbach, P.; Wan, H.; Gierus, M.; Taubeet, F. Effects of grazing intensity and environmental factors on species composition and diversity in typical steppe of Inner Mongolia, China. PLoS ONE 2012, 7, e52180. [Google Scholar] [CrossRef] [PubMed]
- Esaete, J.; Muwanika, V.B.; Musiba, R.; Mawa, C.; Tabuti, J.R.S. Understory seedling bank in forest areas with a differing period of recovery in Mabira Central Forest Reserve, South Central Uganda. Environ. Manag. 2022, 1–11. [Google Scholar] [CrossRef]
- Gonzalez, M.; Deconchat, M.; Balent, G.; Cabanettes, A. Diversity of woody plant seedling banks under closed canopy in fragmented coppice forests. Ann. Forest Sci. 2008, 65, 511. [Google Scholar] [CrossRef]
- Albrecht, M.A.; McCarthy, B.C. Seedling establishment shapes the distribution of shade-adapted forest herbs across a topographical moisture gradient. J. Ecol. 2009, 97, 1037–1049. [Google Scholar] [CrossRef]
- Hu, A.; Zhang, J.; Chen, X.J.; Millner, J.P.; Chang, S.H.; Bowatte, S.; Hou, F.J. The composition, richness, and evenness of seedlings from the soil seed bank of a semi-arid steppe in northern China are affected by long-term stocking rates of sheep and rainfall variation. Range. J. 2019, 41, 23–32. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chu, L.; Daryanto, S.; Wang, L.X.; Lin, J.X.; Ala, M. The impact of grazing on seedling patterns in degraded sparse-elm grassland. Land Degrad. Dev. 2018, 29, 2330–2337. [Google Scholar] [CrossRef]
- Janisova, M.; Skodova, I.; Hegedusova, K.; Kochjarova, J. Seed bank and seedling recruitment of endangered Tephroseris longifolia subsp moravica (Asteraceae). Folia Geobot. 2016, 51, 343–360. [Google Scholar] [CrossRef]
- Grewel, B.J.; Gillard, M.B.; Futrell, C.J.; Castillo, J.M. Seedling Emergence from Seed Banks in Ludwigia hexapetala-Invaded Wetlands: Implications for Restoration. Plants 2019, 8, 451. [Google Scholar] [CrossRef] [PubMed]
- Oesterheld, M.; Sala, O.E. Effects of Grazing on Seedling Establishment-the Role of Seed and Safe-Site Availability. J. Veg. Sci. 1990, 1, 353–358. [Google Scholar] [CrossRef]
- Fay, P.A.; Schultz, M.J. Germination, survival, and growth of grass and forb seedlings: Effects of soil moisture variability. Acta Oecol. 2009, 35, 679–684. [Google Scholar] [CrossRef]
- Osem, Y.; Konsens, I.; Perevolotsky, A.; Kigel, J. Soil seed bank and seedling emergence of Sarcopoterium spinosum as affected by grazing in a patchy semiarid shrubland. Israel J. Plant Sci. 2007, 55, 35–43. [Google Scholar] [CrossRef]
- Eichberg, C.; Donath, T.W. Sheep trampling on surface-lying seeds improves seedling recruitment in open sand ecosystems. Restor. Ecol. 2018, 26, 211–219. [Google Scholar] [CrossRef]
- Gallacher, D.J.; Hill, J.P. Effects of camel grazing on density and species diversity of seedling emergence in the Dubai (UAE) inland desert. J. Arid Environ. 2008, 72, 853–860. [Google Scholar] [CrossRef]
- Zhu, Y.J.; Yang, X.J.; Baskin, C.C.; Baskin, J.M.; Dong, M.; Huang, Z.Y. Effects of amount and frequency of precipitation and sand burial on seed germination, seedling emergence and survival of the dune grass Leymus secalinus in semiarid China. Plant Soil 2014, 374, 399–409. [Google Scholar] [CrossRef]
- Classen, A.T.; Norby, R.J.; Campany, C.E.; Sides, K.E.; Weltzin, J.F. Climate change alters seedling emergence and establishment in an old-field ecosystem. PLoS ONE 2010, 5, e13476. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Davison, E.A. Emergence and survival of herbaceous seedlings in temperate grassy woodlands: Recruitment limitations and regeneration niche. Austral Ecol. 2004, 29, 320–331. [Google Scholar] [CrossRef]
- Wang, D.B.; Wang, X.Y.; Wu, Y.; Lin, H.L. Grazing buffers the effect of climate change on the species diversity of seedlings in an alpine meadow on the Tibetan Plateau. Ecol. Evol. 2019, 9, 1119–1126. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Li, F.R. Characteristics of the soil seed bank and the seedling bank in fenced sandy meadow. Acta Bot. Boreali-Occident. Sin. 2003, 23, 1725–1730. [Google Scholar]
- Yue, X.F.; Zhang, T.H.; Zhao, X.Y.; Liu, X.P.; Ma, Y.H. Effects of rainfall patterns on annual plants in Horqin Sandy Land, Inner Mongolia of China. J. Arid Land. 2016, 8, 389–398. [Google Scholar] [CrossRef]
- Liu, X.P.; He, Y.H.; Sun, S.S.; Zhang, T.H.; Luo, Y.Q.; Zhang, L.M.; Wang, M.M.; Cheng, L.; Hu, H.J.; Xu, Y.Z. Restoration of sand-stabilizing vegetation reduces deep percolation of precipitation in semi-arid sandy lands, northern China. Catena 2022, 208, 105728. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhao, H.L.; Zhao, X.Y.; Zhang, T.H.; Li, Y.L.; Cui, J.Y. Effects of grazing and livestock exclusion on soil physical and chemical properties in desertified sandy grassland, Inner Mongolia, northern China. Environ. Earth Sci. 2011, 63, 771–783. [Google Scholar] [CrossRef]
- Ma, Y.H.; Zhang, T.H.; Liu, X.P. Effect of intensity of small rainfall simulation in spring on annuals in Horqin Sandy Land, China. Environ. Earth Sci. 2015, 74, 727–735. [Google Scholar] [CrossRef]
- Yao, S.; Zhang, T.; Zhao, C. Multiple-time-scale Analysis on Precipitation in Naiman Banner of Horqin Sandy Land in 1970–2010. J. Desert Res. 2014, 34, 42–549. [Google Scholar] [CrossRef]
- He, Y.H.; Zhao, H.L.; Liu, X.P.; Zhang, T.H. Seed germination and population dynamic of two annual plants in sand land restoration process. J. Desert Res. 2010, 30, 1331–1335. [Google Scholar]
- Zhao, H.L.; Okuro, T.; Zhou, R.L.; Li, Y.L.; Zuo, X.A.; Huang, G. Effects of grazing and climate change on species diversity in sandy grassland, Inner Mongolia, China. Sci. Cold Arid Reg. 2009, 1, 30–38. [Google Scholar]
- Osem, Y.; Perevolotsky, A.; Kigel, J. Similarity between seed bank and vegetation in a semi-arid annual plant community: The role of productivity and grazing. J. Veg. Sci. 2006, 17, 29–36. [Google Scholar] [CrossRef]
- Li, X.H.; Li, X.L.; Jiang, D.; Liu, Z.; Yu, Q.H. Annual plants in arid and semi-arid desert regions. Front. Biol. 2008, 3, 259–264. [Google Scholar] [CrossRef]
- Harrison, S.; Inouye, B.D.; Safford, H.D. Ecological heterogeneity in the effects of grazing and fire on grassland diversity. Conserv. Biol. 2003, 17, 837–845. [Google Scholar] [CrossRef]
- Hopfensperger, K.N. A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 2007, 116, 1438–1448. [Google Scholar] [CrossRef]
- Tessema, Z.K.; de Boer, W.F.; Baars, R.M.T.; Prins, H.H.T. Influence of grazing on soil seed banks determines the restoration potential of aboveground vegetation in a semi-arid Savanna of Ethiopia. Biotropica 2012, 44, 211–219. [Google Scholar] [CrossRef]
- Zhao, W.Z.; Liu, Z.M.; Chang, X.L. Influence of grazing intensity on seed bank of a sandy grassland in Horqin Steppe of China. Ann. Arid Zone. 2001, 40, 397–404. [Google Scholar]
- Jahantab, E.; Yazdanshenas, H.; Saray, A.A.; Matinkhah, S.H.; Khazaei, M. Seed burial depth, seedling emergence, and height as affected by animal trampling in marl soils. Plant Ecol. 2022, 223, 493–506. [Google Scholar] [CrossRef]
- Liu, Z.M.; Hua, L.X.; Ping, L.R.; Luo, Y.M.; Wang, H.M.; Jiang, D.M. A comparative study on diaspore shape of 70 species found in the sandy land of Horqin. Acta Prataculturae Sci. 2003, 12, 55–61. [Google Scholar]
- Ramawat, K.G. Desert Plants: Biology and Biotechnology; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Willms, W.D.; Quinton, D.A. Grazing effects on germinable seeds on the fescue prairie. J. Range Manag. 1995, 48, 423–430. [Google Scholar] [CrossRef]
- Gutterman, Y. Seed Germination in Desert Plants; Springer: Berlin/Heidelberg, Germany, 1993; p. 253. [Google Scholar]
- Tobe, K.; Zhang, L.; Omasa, K. Seed germination and seedling emergence of three annuals growing on desert sand dunes in China. Ann. Bot. 2005, 95, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.P.; He, Y.H.; Zhao, X.Y.; Zhang, T.H.; Li, Y.L.; Yun, J.Y. The response of soil water and deep percolation under Caragana microphylla to rainfall in the Horqin Sand Land, northern China. Catena 2016, 139, 82–91. [Google Scholar] [CrossRef]
- Tadey, M.; Souto, C.P. Unexpectedly, intense livestock grazing in arid rangelands strengthens the seedling vigor of consumed plants. Agron. Sustain. Dev. 2016, 36, 63. [Google Scholar] [CrossRef]
- Schuster, M.Z.; Pelissari, A.; de Vioraes, A.; Harrison, S.K.; Sulc, R.M.; Lustosa, S.B.C.; Anghinoni, I.; Carvalho, P.C.F. Grazing intensities affect weed seedling emergence and the seed bank in an integrated crop-livestock system. Agric. Ecosyst. Environ. 2016, 232, 232–239. [Google Scholar] [CrossRef]
- Kiss, R.; Deak, B.; Tothmeresz, B.; Miglecz, T. Establishment gaps in species-poor grasslands: Artificial biodiversity hotspots to support the colonization of target species. Restor. Ecol. 2021, 29, e13135. [Google Scholar] [CrossRef] [Green Version]
- Garwood, N.C. Seed germination in a seasonal tropical forest in Panama: A community study. Ecol. Monogr. 1983, 53, 159–181. [Google Scholar] [CrossRef]
- Miller, T.E. Effects of emergence time on survival and growth in an early old-field plant community. Oecologia 1987, 72, 272–278. [Google Scholar] [CrossRef]
- Noe, G.B.; Zedler, J.B. Spatio-temporal variation of salt marsh seedling establishment in relation to the abiotic and biotic environment. J. Veg. Sci. 2001, 12, 61–74. [Google Scholar] [CrossRef]
- Li, X.H. Biology of Annual Plants in Arid and Semi-Arid Desert Regions of China; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Nunes, F.P.; Dayrell, R.L.; Silveira, F.A.; Negreiros, D.; Santana, D.G.d.; Carvalho, F.J.; Garcia, Q.S.; Fernandes, G.W. Seed Germination Ecology in Rupestrian Grasslands. In Ecology and Conservation of Mountaintop Grasslands in Brazil; Fernandes, G., Ed.; Springer: Cham, Switzerland, 2016; pp. 207–225. [Google Scholar]
- Houle, G. Environmental filters and seedling recruitment on a coastal dune in subarctic Quebec (Canada). Can. J. Bot. 1996, 74, 1507–1513. [Google Scholar] [CrossRef]
- Castillo-Díaz, D.; Chen, H.Y.; Harrison, R.D.; Wen, B.; Goodale, U.M. Seedling emergence and environmental filters determine Ficus recruitment in a subtropical landscape. Forest Ecol. Manag. 2021, 497, 119536. [Google Scholar] [CrossRef]
- Venable, D.L.; Lawlor, L. Delayed germination and dispersal in desert annuals: Escape in space and time. Oecologia 1980, 46, 272–282. [Google Scholar] [CrossRef] [PubMed]
- López, R.P.; Hayes, M.; Maidana-Tuco, Y.; Naoki, K. Summer annual plants exhibit germination responses to an aridity gradient consistent with both delayed germination and phenotypic plasticity. Plant Ecol. 2020, 221, 1233–1242. [Google Scholar] [CrossRef]
- Li, X.; Li, X.L.; Jiang, D.M.; Liu, Z.M. Germination strategies and patterns of annual species in the temperate semiarid region of China. Arid Land Res. Manag. 2006, 20, 195–207. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Precipitation (mm) | Temperature (°C) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| April | May | June | July | August | Total | April | May | June | July | August | Average | |
| 2019 | 6.2 | 72.0 | 59.8 | 61.6 | 141.4 | 341.0 | 9.8 | 18.3 | 21.4 | 24.5 | 21.2 | 19.0 |
| 2020 | 3.0 | 92.6 | 43.6 | 30.6 | 59.4 | 229.2 | 8.7 | 16.5 | 22.3 | 24.6 | 22.4 | 18.9 |
| Factors | Seedlings of Grasses | Seedlings of Forbs | Seedlings of All Annuals | |||
|---|---|---|---|---|---|---|
| F | Sig. | F | Sig. | F | Sig. | |
| Grazing | 24.283 | 0.000 *** | 17.353 | 0.000 *** | 21.666 | 0.000 *** |
| Year | 20.744 | 0.000 *** | 28.160 | 0.000 *** | 16.129 | 0.000 *** |
| Grazing × Year | 0.950 | 0.391 | 12.611 | 0.000 *** | 15.680 | 0.000 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Liu, X. Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland. Agronomy 2022, 12, 2131. https://doi.org/10.3390/agronomy12092131
He Y, Liu X. Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland. Agronomy. 2022; 12(9):2131. https://doi.org/10.3390/agronomy12092131
Chicago/Turabian StyleHe, Yuhui, and Xinping Liu. 2022. "Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland" Agronomy 12, no. 9: 2131. https://doi.org/10.3390/agronomy12092131
APA StyleHe, Y., & Liu, X. (2022). Grazing Effects on Seedling Emergence in Contrasting Wet and Dry Year in a Semiarid Grassland. Agronomy, 12(9), 2131. https://doi.org/10.3390/agronomy12092131