1. Introduction
Wheat (
Triticum aestivum L.) is hexaploid (2 n = 6 x = 42) (Bhatta, 2017) and one of the most important foods in human nutrition all over the World [
1]. It is also the staple diet for the majority of the global population, containing carbohydrates, protein, minerals, B-group vitamins, dietary fiber, etc. [
2]. Wheat can be grown in different climatic zones ranging from the Caspian Sea to China [
3]. The presumed center of wheat origin and diversity is situated in the Fertile Crescent [
4], which includes part of present-day Turkey [
5]. Thus, the diversity of wheat in Turkey has a global role in providing important genetic resources for wheat improvement. In recent years, there has been a rapid increase in the number of varieties released in Turkey, and there are now 201 officially listed bread wheat varieties [
6]. Turkey, particularly Southeast Anatolia, is known as the primary gene center of wheat diversification as well as the area of first wheat domestication around 10,000 BP [
7,
8,
9,
10,
11]. Turkish wheat variations have received a great deal of attention since the beginning of the 20th century. Exploration and collection missions were mounted, and the collected germplasm was evaluated in different countries [
8,
12,
13].
World annual wheat production was realized at 777 million tons in 2021, but world wheat demand is expected to be 1 billion tons by the year 2024 [
14]. Wheat annual average production has entered a period of stagnation and declining in recent years due to biotic and abiotic stressors posing serious threats on wheat cultivation [
15]. In order to use gene resources effectively in wheat breeding to produce tolerant genotypes, it is important to know the characteristics of existing gene resources and to carry out targeted breeding studies. Germplasm characterization is considered a prerequisite for breeding activities as it facilitates the introduction of novel genetic variations to the breeders that can be used for marker-assisted breeding [
16].
Conventional breeding methods are largely employed to improve plant resistance against these stress factors. The process of cultivating plants has resulted in a narrowing of the genetic base in several crop types, including wheat [
17]. Such a narrow genetic base poses a serious threat to sustainable production, which is very important for the rapidly increasing global population and the changing climate conditions due to global warming in recent years [
18].
Genetic diversity is the basis for the long-lasting success of breeding programs, and modeling genetic diversity with various methods can reveal possible adaptations to different regions. The study of genetic variation also allows the identification of genetic characters associated with important breeding targets [
19]. Different genetic materials are used in wheat breeding programs; they are used to expand the existing genetic basis [
20]. Evaluation of variation in germplasm is an important step in breeding programs because it can help select varieties and lines with higher diversity and better performance under certain conditions [
21,
22]. Molecular markers are widely used to reveal wheat genetic diversity [
18,
20]. Molecular markers have become important tools for breeding studies and allow breeding studies to be concluded in a shorter time. Molecular markers are independent of environmental conditions, providing a direct estimation of genetic variation at the DNA level. Thanks to scientific studies, molecular markers with different properties have been developed over time [
23,
24]. Various methods have used molecular markers, including amplified fragment length polymorphisms (AFLPs) [
25], random amplified polymorphic DNA (RAPD) [
26], start codon targeted markers (SCoT) [
27], single nucleotide polymorphism (SNP) [
28], inter simple-sequence repeat (ISSR) [
29], simple-sequence repeats (SSR) [
30], expressed sequence tag (EST) [
31], next generation sequencing (NGS) [
20], diversity Arrays Technology (DArT) [
32] and single nucleotide polymorphism (SNP) [
33], all to assess the genetic diversity and associations among several
Triticum species.
Retrotransposons (RTNs) are an important source of plant genetic variation, and in many plants with large genomes, over 50% of the nuclear DNA is made by retrotransposons [
34]. The inter-primer binding site (iPBS) method has been verified to be an influential DNA fingerprinting technique with no need for previous knowledge of a sequence. iPBS-retrotransposons are the only retrotransposon-based marker system that has allowed polymorphism visualization throughout the plant kingdom. iPBS-retrotransposon DNA markers have been successfuly utilized in assessing genetic diversity [
35]. The iPBS-retrotransposon marker has been demonstrated by many studies, and there are studies on genetic diversity for these markers [
16,
24]. Previously, different molecular markers have been used for the molecular characterization of wheat germplasm [
16]. However, there are limited studies on the characterization of wheat populations using iPBS-retrotransposons. Therefore, the present study aimed to genetically characterize and determine the population structure of the Turkish bread wheat genotypes for the evaluation of genetic diversity. In order for breeding studies to be sustainable, it is necessary to identify, define, and use genetic resources. We believe that the findings from the study will help with the use, development, and conservation of local varieties that can adapt to changing environmental conditions.
4. Discussion
The determination of genetic variation and population structure in plants is important for the future and progress of plant breeding programs [
50]. A rational study has been documented for the characterization of the bread wheat genotype and its wild relatives using several types of molecular markers [
51]. Molecular markers such as iPBS play a critical role in the selection of targeted parents, independent of external ecological factors, to reveal the genetic relationships among plant species. Along with the role of retrotransposons in the diversification of genetic material, it is stated that retrotransposon activation is one of the key factors involved in host adaptation to ecological differences [
52]. The iPBS markers used in our study helped identify wheat (
Triticum aestivum L.) accessions at the molecular level. The obtained data provided important information about the genetic relationships among wheat accessions. The information obtained by the iPBS marker system indicates that it can be usefully used for diversity studies and genetic analysis of wheat accessions. Similarly, Nadeem [
16] used the iPBS marker system for the molecular characterization of bread wheat landraces and cultivars. In addition, Demirel, [
53] used iPBS-retrotransposon markers for the molecular characterization of emmer and durum wheat.
Total and polymorphic bands reported in this study were higher than Nadeem [
16] and Demirel [
53] using iPBS markers, as well as Kumar et al. [
54] using ISSR markers, Alshehri et al. [
55] using SCoT and ISSR primers, and Çifçi and Yağdi [
56] using RAPD markers. The PIC value is an important piece of information in scoring the efficacy of polymorphic loci and demonstrating the discriminative power of a primer [
57]. In our study, the PIC varied between 0.048 (iPBS-2087) and 0.279 (iPBS 2380 and iPBS 2381) (mean 0.175). In a similar study of wheat in which iPBS markers were used, PIC values were reported between 0.12 and 0.25 (mean 0.19) [
53]. The results differ from those of [
58], who found PIC values between 0.306 and 0.913 (mean 0.660) in their study with iPBS-retrotransposons in wheat. In addition, PIC values ranged from 0.702 to 0.11, and the mean PIC value was 0.42 via Nadeem [
16], who used the iPBS marker for the molecular characterization of 74 bread wheat accessions, including a total of 54 landraces and 20 commercial cultivars. This suggests that the difference in results between studies is probably due to the different genotypes and the use of fewer markers by other researchers.
The mean number of effective alleles (ne), genetic diversity of Nei (h), and Shannon’s information index (I) value of the wheat varieties were calculated as 1.157, 0.95, and 0.144, respectively. In a similar study conducted in wheat, the mean ne, h, and I values were 1.950, 0.489, and 0.502, respectively [
58]. These values were higher than in our study. A higher number of effective alleles is an indication of higher genetic variation and is therefore always desirable. In a study conducted by Kumar et al. [
54] and Nadeem [
16], higher gene diversity was observed for the primers producing higher alleles. In addition, Shannon’s information index is one of the important criteria that distinguishes genetic variation within a population and makes variation more meaningful. Shannon’s mean information index observed in the present report was lower than Nadeem’s [
16] and Demirel’s [
53]. The presence of lower values for various diversity indices in this study might be due to differences in germplasm. The Nei’s genetic distance revealed G1 (Aksel 200) and G63 (Karasu 90) as genetically distinct accessions. Therefore, Aksel and Karasu accessions can be used for future bread wheat breeding. Arystanbekkyzy et al. [
58] stated that genetically distinct accessions can be helpful to start breeding activities for favorable traits.
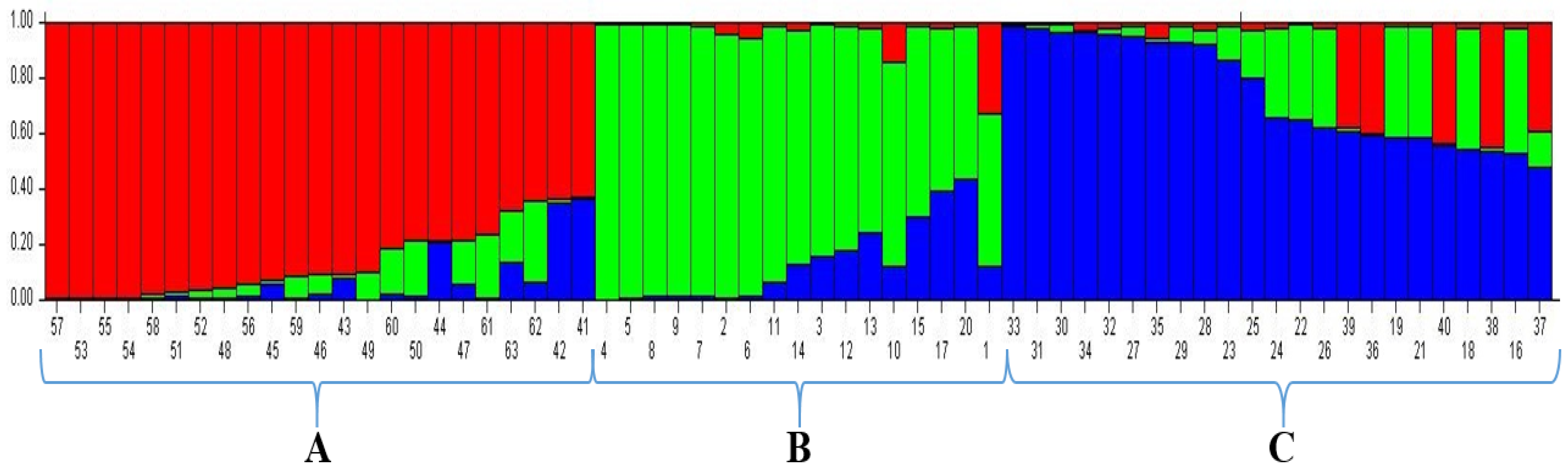
The UPGMA dendrogram placed the 63 wheat genotypes into three clusters. Clusters I, II, and III included 14 (22.25%), 14 (22.25%), and 35 (55.5%) genotypes, respectively. The UPGMA analysis showed admixture of accessions because accessions from various provinces were grouped under the same sub-groups. This result was similar to Nadeem [
16]. Uncovering genetic variations occurring between populations of plant species and the information obtained are valuable for breeding and conservation of genetic resources [
59]. The population structure within each wheat genotype or cultivar can be used to make crossbreeding studies more efficient. There was concordance between the population structure identified in this study and distance-based clustering from principal coordinate analysis (PCoA). In our study, it can be thought that intraspecies crossings may produce stronger hybrids due to greater genetic distances. During this study, winter and alternative bread wheat genotypes were used as plant material. Therefore, calculated diversity indices among genotypes showed the presence of fewer genetic variations. Our result is in contrast with Nadeem [
16] and Demirel [
53]. This can be due to the use of different genetic materials. In addition, Keser et al. [
6] evaluated 22 registered winter bread wheat varieties developed for rainfed conditions under dryland conditions and presented the pedigree information of the varieties, of which 17 genotypes were common with our study. Based on pedigree information of varieties, cv. Sonmez-2001 was originated from cv. Bezostaya-1. In our study, cv Sonmez-2001 fell into a different PCoA group than cv. Bezostaya-1. There could be two explanations for this phenomenon. One could be due to the limitations of iPBS markers. There could be similar mobility or a similar pattern of retrotransposon elements in these cultivars. Therefore, iPBS markers could not sufficiently differentiate between these two genotypes or identify genetic differences as these genotypes are in the genic region of the genome. A second explanation would be that iPBS markers work well. These two genotypes could be different genotypically. As it was seen in the pedigree of cv. Sonmez-2001, we see the contribution of other four genotypes (Tevere/3/Kremena/Lovrin-29/4/Katya-1) into the Sonmez-2001 genotypic background. Indeed, cv Sonmez-2001 has been genetically separated from Bezostaya in every generation.
During this study, bread wheat cultivars were used as plant material. Therefore, calculated diversity indices among cultivars showed the presence of higher genetic variations among cultivars. PCoA analysis was also performed to reveal genetic relationships among wheat inclusions, and in the first three axes, PCoA analysis explained 60.10% of the total variation. As a result of the PCoA analysis, it is stated that the data obtained from this matrix is generally safe if the axes explain 25% or more of the total variation [
60]. The PCoA analysis supported the clustering of the model-based structure algorithm and separated the Turkish bread wheat germplasm into three populations.
The AMOVA results revealed that the maximum genetic variations in Turkish bread wheat germplasm are present within the population. The result from the molecular analysis of variance (99%) revealed high variation among wheat accessions. Many factors, such as pollination patterns of species, selection, gene flow between locations, and variation in genotypes, affect the variation [
41]. Thus, it is stated that Turkish bread wheat germplasm has a great level of genetic variation within the population that can be helpful for the breeding of this crop in the future. The findings revealed that wheat genotypes were divided into three groups according to genetic structure analysis. The results of the AMOVA were supported by previous studies as they also revealed higher genetic variations within populations [
16,
61].
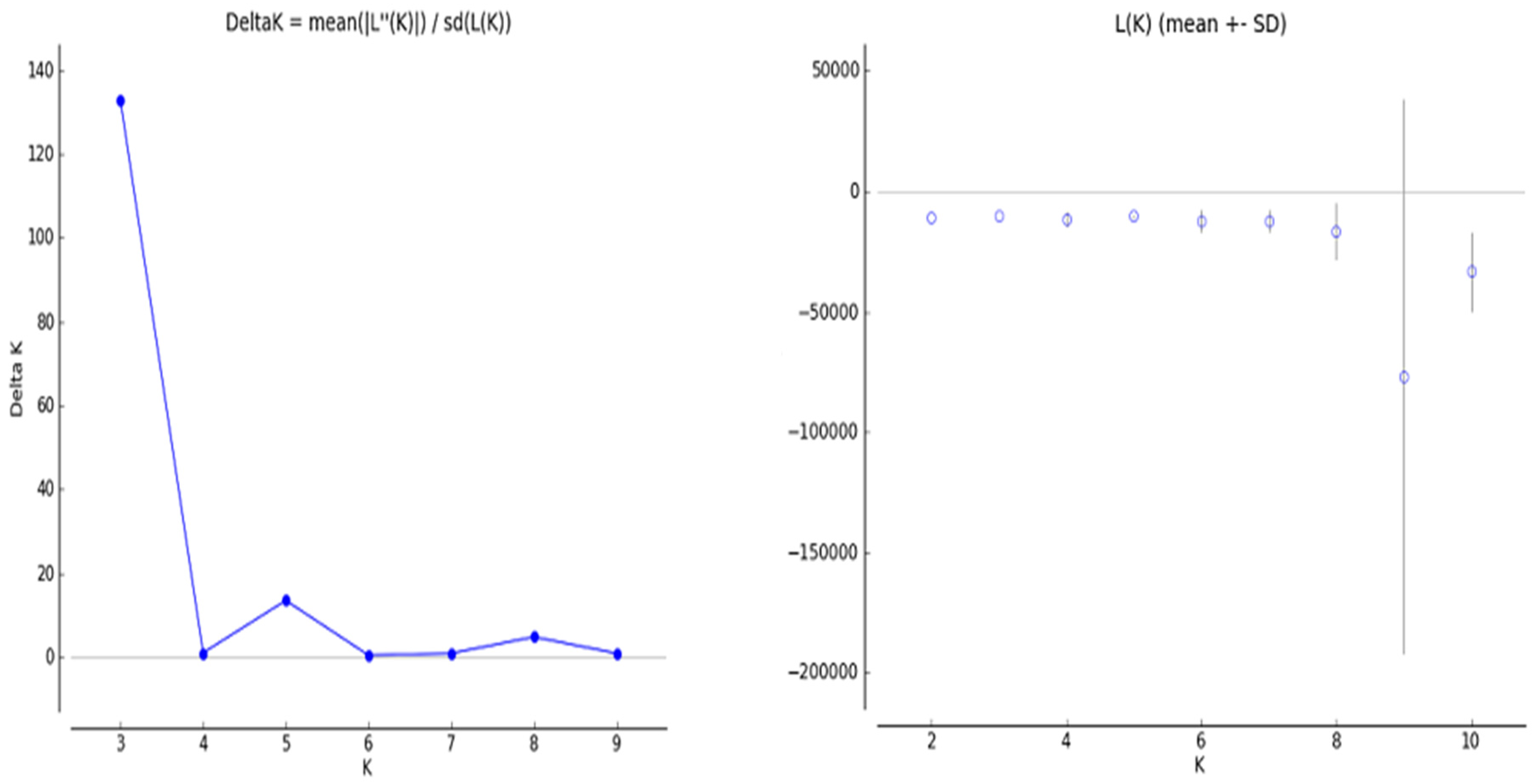
The structure algorithm has been proven more trustworthy and informative compared to other clustering algorithms [
62]. Recently, it has become important to understand population structure, which allows for the selection of various parents for use in breeding programs and the mapping of marker-trait relationships. Analysis of population structure is an important tool for estimating the levels of similarity between individuals and subpopulations, or, in other words, for understanding population structure. The model-based structure algorithm grouped 63 bread wheat accessions into three populations on the basis of their collection points. Population A was found to be larger than population B and C by accounting for 23.80% (15 genotypes) of its accessions. Population B accounted for a total of 13 accessions (20.63%), and population C was found to be the lowest. The other two populations accounted for a total of 11 accessions (17.46%), as well as mixed groups with 38.09% and a total of 24 accessions (membership probability <0.8). Cömertpay et al. [
35] showed that in the STRUCTURE analysis, the number of groups (K) varied from 1 to 10. The estimated ∆K value was 302.29 for the 81 rice accessions, which represented the three subpopulations. The population structure analysis revealed that all of the varieties used in this study were derived from three subpopulations, or three gene pools. Throughout the evolutionary process, varieties accumulate several living mutations that form the basis of genetic diversity. Moreover, forces such as recombination, random drift, and natural selection shape the genetic makeup of populations. The results show that mixed groups have the highest amount, an indication that genotypes have crossing with each other over the years and how narrow range of the genetic pool.





 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}