
Assessment of Yield Stability of Bambara Groundnut (Vigna subterranea (L.) Verdc.) Using Genotype and Genotype–Environment Interaction Biplot Analysis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
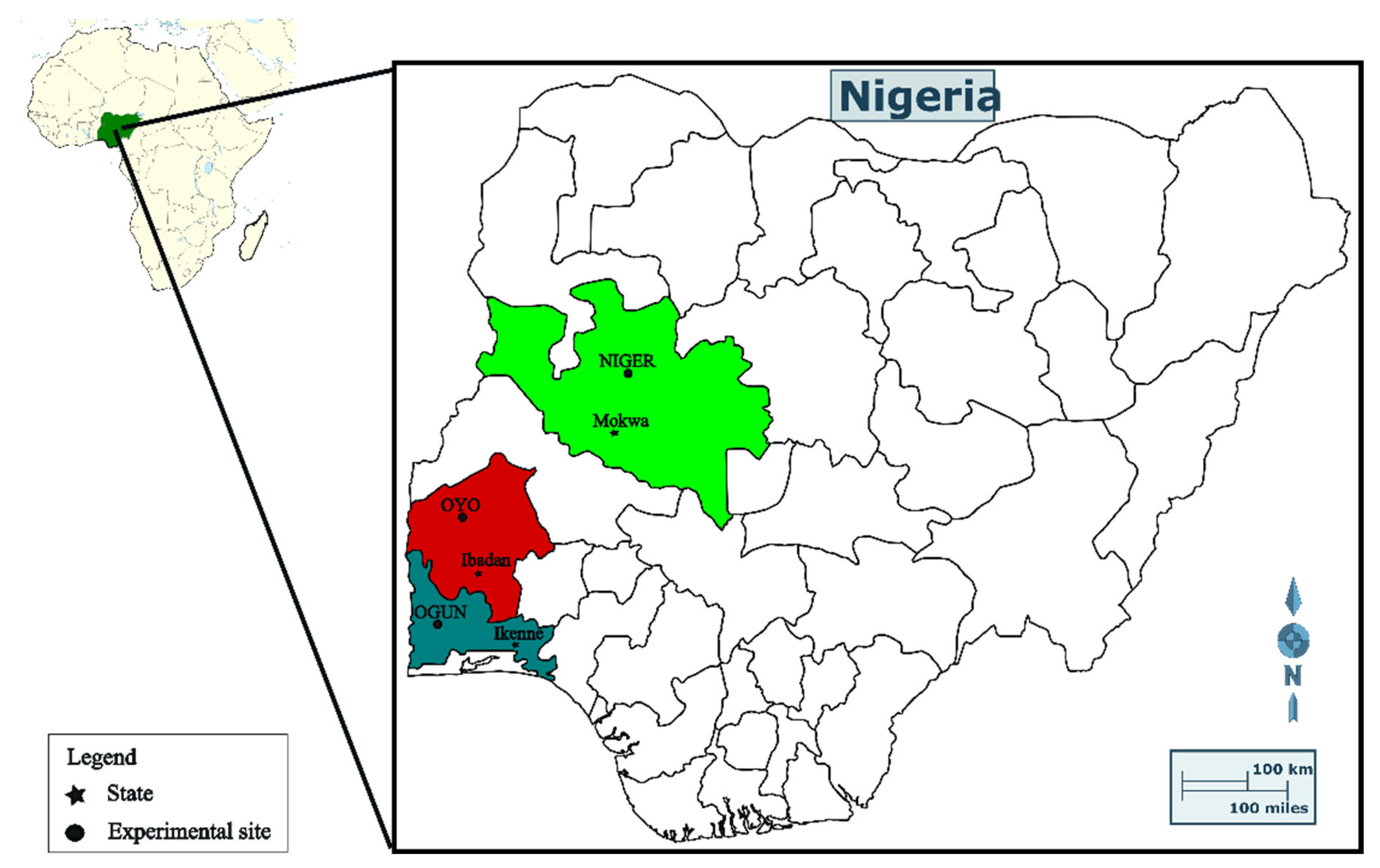
2.2. Study Site Description
2.3. Soil Sampling and Analysis
2.4. Field Trials and Yield Data
2.5. Statistical Analysis
3. Results and Discussion
3.1. Soil Analysis
3.2. Pooled Analysis of Variance
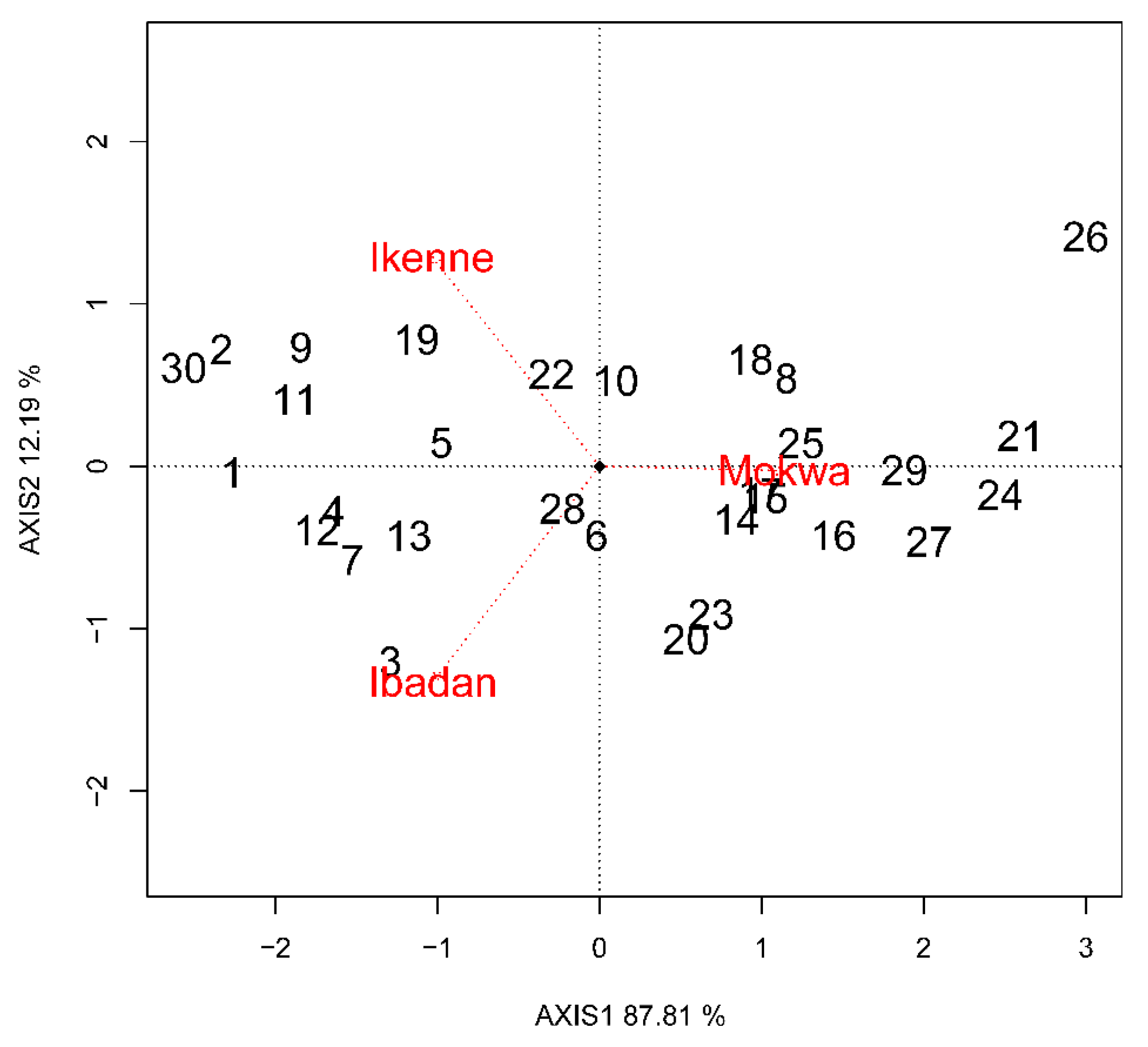
3.3. GGE Biplot Analysis
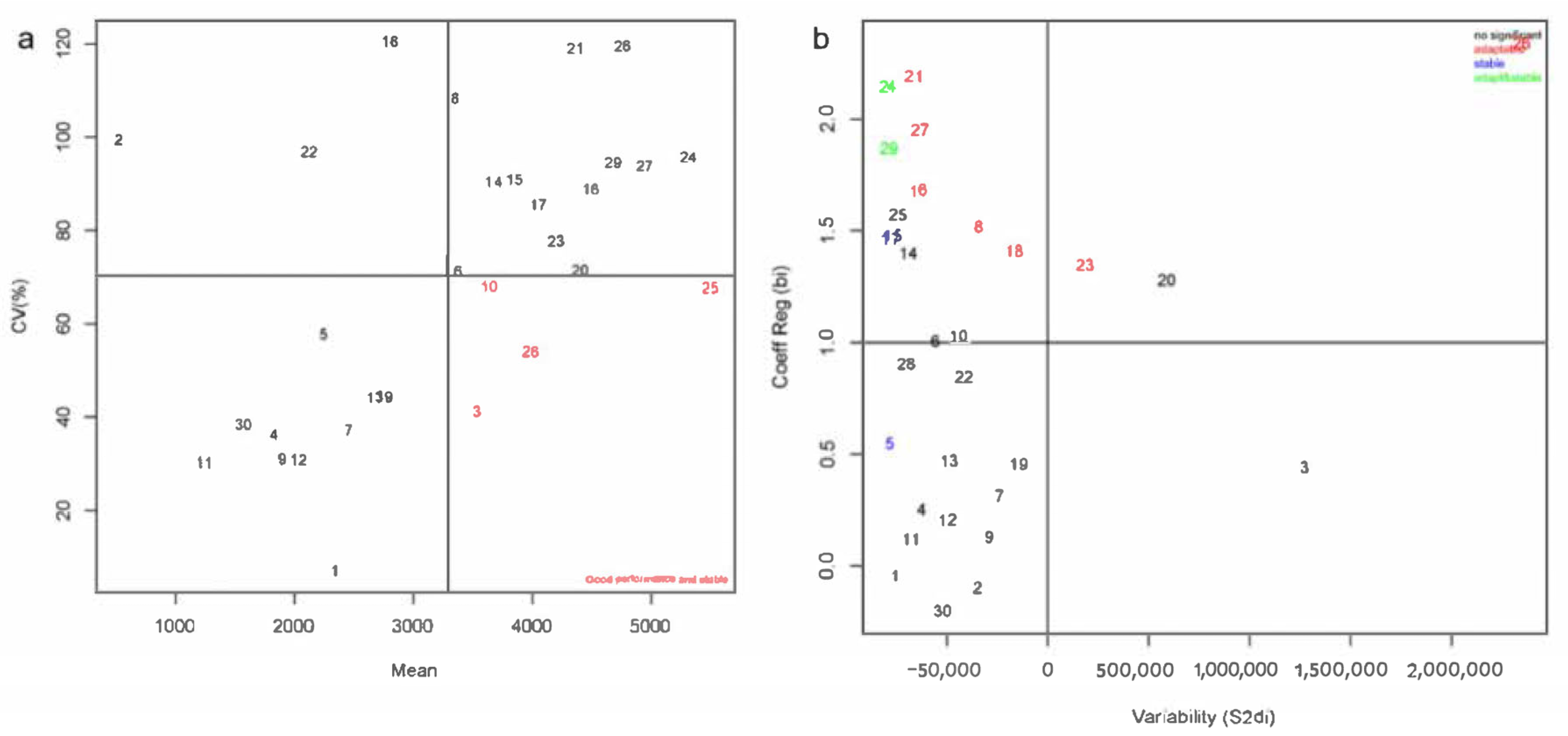
3.4. Stability Analysis
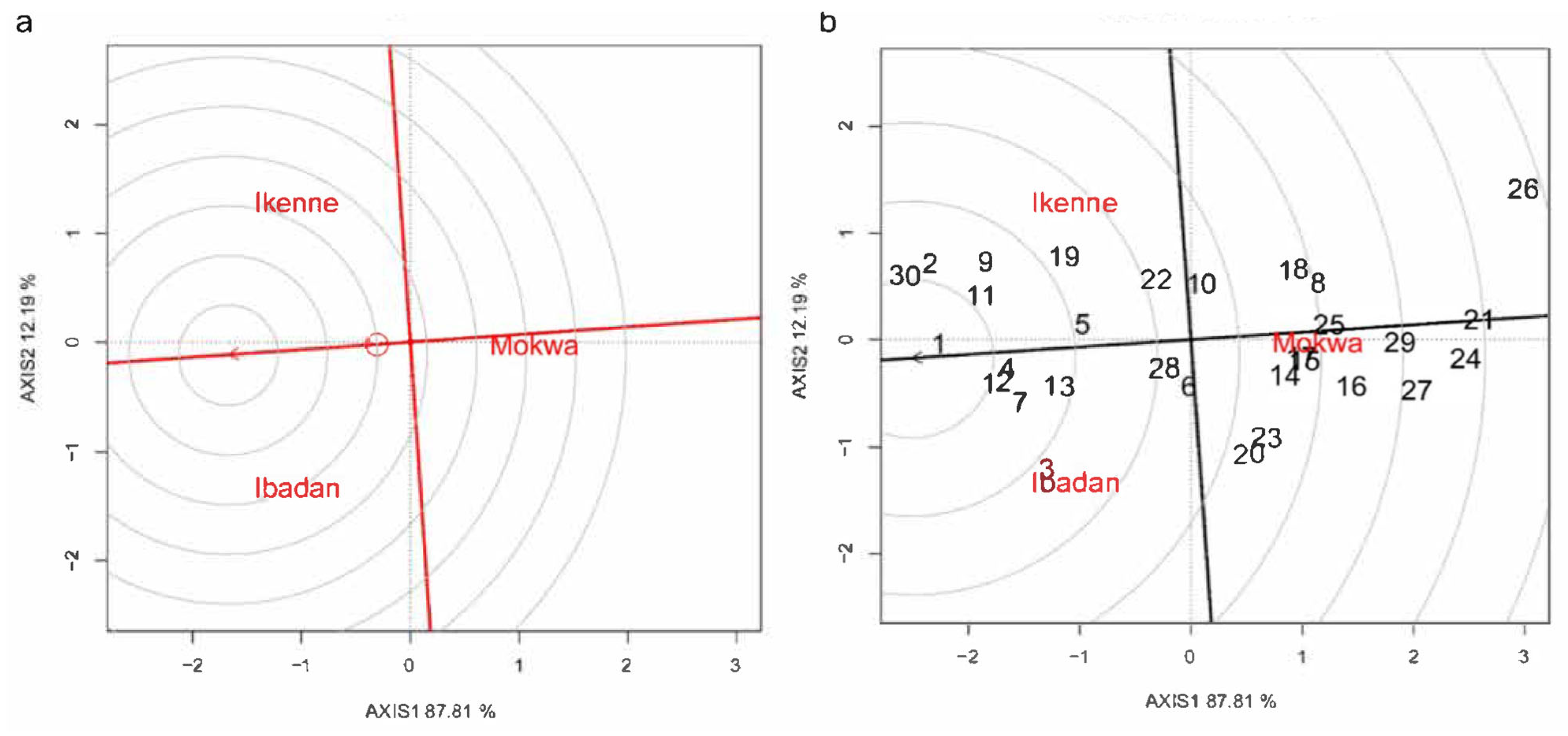
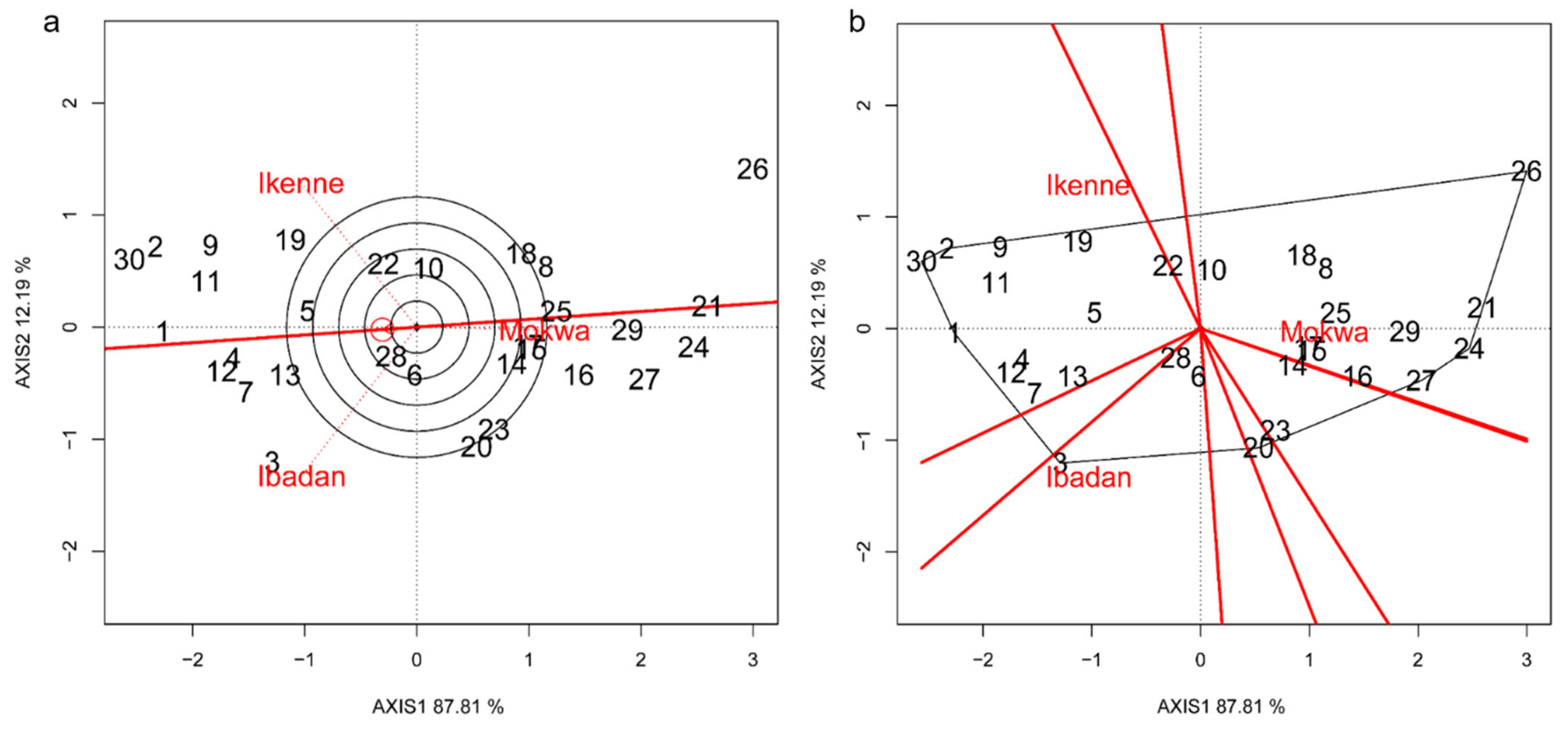
3.5. Environment and Genotype Ranking Analysis and Relationship among Environments
3.6. Discriminating and Representativeness and “Which-Won-Where” Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. World Food and Agriculture—Statistical Yearbook; FAO: Rome, Italy, 2020; p. 366. [Google Scholar]
- Atoyebi, O.J.; Osilesi, O.; Adebawo, O.; Abberton, M. Evaluation of nutrient parameters of selected African accessions of Bambara groundnut (Vigna subterranea (L.) Verdc.). Am. J. Food Nutr. 2017, 5, 83–89. [Google Scholar] [CrossRef]
- Harris, T.; Jideani, V.; Le Roes-Hill, M. Flavonoids and tannin composition of Bambara groundnut (Vigna subterranea) of Mpumalanga, South Africa. Heliyon 2018, 4, e00833. [Google Scholar] [CrossRef] [PubMed]
- Halimi, R.A.; Barkla, B.J.; Mayes, S.; King, G.J. The potential of the underutilized pulse Bambara groundnut (Vigna subterranea (L.) Verdc.) for nutritional food security. J. Food Compos. Anal. 2019, 77, 47–59. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Oyatomi, O.; Abberton, M.; Babalola, O.O. Variations of Nutrient and Antinutrient Components of Bambara Groundnut (Vigna subterranea (L.) Verdc.) Seeds. J. Food Qual. 2022, 2022, 2772362. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Oyatomi, O.; Babalola, O.O.; Abberton, M. Breeding potentials of Bambara groundnut for food and nutrition security in the face of climate change. Front. Plant Sci. 2022, 12, 3186. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Oyatomi, O.; Babalola, O.O.; Abberton, M. Genetic Diversity and Environmental Influence on Growth and Yield Parameters of Bambara Groundnut. Front. Plant Sci. 2021, 12, 796352. [Google Scholar] [CrossRef]
- Pasquet, R.S.; Schwedes, S.; Gepts, P. Isozyme diversity in Bambara groundnut. Crop Sci. 1999, 39, 1228–1236. [Google Scholar] [CrossRef]
- Aliyu, S.; Massawe, F.; Mayes, S. Genetic diversity and population structure of Bambara groundnut (Vigna subterranea (L.) Verdc.): Synopsis of the past two decades of analysis and implications for crop improvement programmes. Genet. Resour. Crop Evol. 2016, 63, 925–943. [Google Scholar] [CrossRef]
- Puozaa, D.K.; Jaiswal, S.K.; Dakora, F.D. African origin of Bradyrhizobium populations nodulating Bambara groundnut (Vigna subterranea L. Verdc) in Ghanaian and South African soils. PLoS ONE 2017, 12, e0184943. [Google Scholar] [CrossRef]
- Babalola, O.O.; Olanrewaju, O.S.; Dias, T.; Ajilogba, C.F.; Kutu, F.R.; Cruz, C. Biological Nitrogen Fixation: The Role of Underutilized Leguminous Plants. In Microorganisms for Green Revolution: Volume 1: Microbes for Sustainable Crop Production; Panpatte, D.G., Jhala, Y.K., Vyas, R.V., Shelat, H.N., Eds.; Springer: Singapore, 2017; pp. 431–443. [Google Scholar] [CrossRef]
- Majola, N.G.; Gerrano, A.S.; Shimelis, H. Bambara groundnut (Vigna subterranea [L.] Verdc.) production, utilisation and genetic improvement in Sub-Saharan Africa. Agronomy 2021, 11, 1345. [Google Scholar] [CrossRef]
- Adeleke, O.R.; Adiamo, O.Q.; Fawale, O.S. Nutritional, physicochemical, and functional properties of protein concentrate and isolate of newly-developed Bambara groundnut (Vigna subterrenea L.) cultivars. Food Sci. Nutr. 2018, 6, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Adeleke, O.; Adiamo, O.Q.; Fawale, O.S.; Olamiti, G. Effect of soaking and boiling on anti-nutritional factors, oligosaccharide contents and protein digestibility of newly developed bambara groundnut cultivars. Turk. J. Agric.-Food Sci. Technol. 2017, 5, 1006–1014. [Google Scholar] [CrossRef]
- Oyeyinka, A.T.; Pillay, K.; Siwela, M. Consumer awareness and acceptability of Bambara groundnut as a protein source for use in complementary foods in rural KwaZulu-Natal. S. Afr. J. Clin. Nutr. 2017, 30, 87–92. [Google Scholar] [CrossRef]
- Khan, M.M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Mamun, A. Genetic Variability, Heritability, and Clustering Pattern Exploration of Bambara Groundnut (Vigna subterranea L. Verdc) Accessions for the Perfection of Yield and Yield-Related Traits. BioMed Res. Int. 2020, 2020, 2195797. [Google Scholar] [CrossRef] [PubMed]
- Mayes, S.; Ho, W.K.; Chai, H.H.; Gao, X.; Kundy, A.C.; Mateva, K.I.; Zahrulakmal, M.; Hahiree, M.K.I.M.; Kendabie, P.; Licea, L.C.S.; et al. Bambara groundnut: An exemplar underutilised legume for resilience under climate change. Planta 2019, 250, 803–820. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Al-Mamun, M. Bambara Groundnut (Vigna subterranea L. Verdc): A Crop for the New Millennium, Its Genetic Diversity, and Improvements to Mitigate Future Food and Nutritional Challenges. Sustainability 2021, 13, 5530. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Plant growth-promoting rhizobacteria for orphan legume production: Focus on yield and disease resistance in Bambara groundnut. Front. Sustain. Food Syst. 2022, 6, 922156. [Google Scholar] [CrossRef]
- Bendu, N.J.M.; Mansaray, N.A.; Bah, N.A.M.; Momoh, N.E.J.J. Effect of Planting Method and Spacing on the Growth Parameters of Three Bambara Groundnut Cultivars in Two Agro-Climatic Zones of Sierra Leone. World J. Biol. Pharm. Health Sci. 2023, 13, 111–121. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Oyatomi, O.; Babalola, O.O.; Abberton, M. GGE biplot analysis of genotype × environment interaction and yield stability in Bambara groundnut. Agronomy 2021, 11, 1839. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Miah, G.; Hussin, G.; Ramli, A. Genotype × Environment interaction and stability analyses of yield and yield components of established and mutant rice genotypes tested in multiple locations in Malaysia. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2017, 67, 590–606. [Google Scholar] [CrossRef]
- Frutos, E.; Galindo, M.P.; Leiva, V. An interactive biplot implementation in R for modeling genotype-by-environment interaction. Stoch. Environ. Res. Risk Assess. 2014, 28, 1629–1641. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 47, 643–653. [Google Scholar] [CrossRef]
- Angelini, J.; Faviere, G.S.; Bortolotto, E.B.; Arroyo, L.; Valentini, G.H.; Domingo Lucio Cervigni, G. Biplot pattern interaction analysis and statistical test for crossover and non-crossover genotype-by-environment interaction in peach. Sci. Hortic. 2019, 252, 298–309. [Google Scholar] [CrossRef]
- Mndolwa, E.; Msolla, S.; Porch, T.; Miklas, P. GGE biplot analysis of yield stability for Andean dry bean accessions grown under different abiotic stress regimes in Tanzania. Afr. Crop Sci. J. 2019, 27, 413–425. [Google Scholar] [CrossRef]
- Dalló, S.C.; Zdziarski, A.D.; Woyann, L.G.; Milioli, A.S.; Zanella, R.; Conte, J.; Benin, G. Across year and year-by-year GGE biplot analysis to evaluate Soybean performance and stability in multi-environment trials. Euphytica 2019, 215, 113. [Google Scholar] [CrossRef]
- Woyann, L.G.; Zdziarski, A.D.; Baretta, D.; Meira, D.; Dallacorte, L.V.; Benin, G. Selection of high-yielding, adapted and stable wheat lines in preliminary trials. Crop Breed. Appl. Biotechnol. 2019, 19, 412–419. [Google Scholar] [CrossRef]
- Saeidnia, F.; Taherian, M.; Nazeri, S.M. Graphical analysis of multi-environmental trials for wheat grain yield based on GGE-biplot analysis under diverse sowing dates. BMC Plant Biol. 2023, 23, 198. [Google Scholar] [CrossRef]
- Singh, C.; Gupta, A.; Gupta, V.; Kumar, P.; Sendhil, R.; Tyagi, B.S.; Singh, G.; Chatrath, R.; Singh, G.P. Genotype × environment interaction analysis of multi-environment wheat trials in India using AMMI and GGE biplot models. Crop Breed. Appl. Biotechnol. 2019, 19, 309–318. [Google Scholar] [CrossRef]
- Bhartiya, A.; Aditya, J.P.; Singh, K.; Pushpendra, P.; Purwar, J.P.; Agarwal, A. AMMI and GGE biplot analysis of multi environment yield trial of Soybean in North Western Himalayan state Uttarakhand of India. Legume Res. Int. J. 2016, 40, 306–312. [Google Scholar] [CrossRef]
- Rezende, W.S.; Beyene, Y.; Mugo, S.; Ndou, E.; Gowda, M.; Sserumaga, J.P.; Asea, G.; Ngolinda, I.; Jumbo, M.; Oikeh, S.O.; et al. Performance and yield stability of maize hybrids in stress-prone environments in eastern Africa. Crop J. 2020, 8, 107–118. [Google Scholar] [CrossRef]
- Enyew, M.; Feyissa, T.; Geleta, M.; Tesfaye, K.; Hammenhag, C.; Carlsson, A.S. Genotype by environment interaction, correlation, AMMI, GGE biplot and cluster analysis for grain yield and other agronomic traits in sorghum (Sorghum bicolor L. Moench). PLoS ONE 2021, 16, e0258211. [Google Scholar] [CrossRef] [PubMed]
- Ngailo, S.; Shimelis, H.; Sibiya, J.; Mtunda, K.; Mashilo, J. Genotype-by-environment interaction of newly-developed sweet potato genotypes for storage root yield, yield-related traits and resistance to sweet potato virus disease. Heliyon 2019, 5, e01448. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Jnanesha, A.C.; Kumar, V.M.; Lal, R.K. GGE biplot vs. AMMI analysis of genotype-by-environment data on essential oil yield in lemongrass [Cymbopogon flexuosus (nees ex. Steud) wats.] grown in semi-arid tropical regions of southern India under different agro-climatic conditions. Biochem. Syst. Ecol. 2022, 103, 104439. [Google Scholar] [CrossRef]
- Fekadu, W.; Mekbib, F.; Lakew, B.; Haussmann, B.I.G. Genotype × environment interaction and yield stability in barley (Hordeum vulgare L.) genotypes in the central highland of Ethiopia. J. Crop Sci. Biotechnol. 2023, 26, 119–133. [Google Scholar] [CrossRef]
- Gerrano, A.S.; Jansen van Rensburg, W.S.; Mathew, I.; Shayanowako, A.I.; Bairu, M.W.; Venter, S.L.; Swart, W.; Mofokeng, A.; Mellem, J.; Labuschagne, M. Genotype and genotype × environment interaction effects on the grain yield performance of cowpea genotypes in dryland farming system in South Africa. Euphytica 2020, 216, 80. [Google Scholar] [CrossRef]
- Senguttuvel, P.; Sravanraju, N.; Jaldhani, V.; Divya, B.; Beulah, P.; Nagaraju, P.; Manasa, Y.; Prasad, A.H.; Brajendra, P.; Gireesh, C. Evaluation of genotype by environment interaction and adaptability in lowland irrigated rice hybrids for grain yield under high temperature. Sci. Rep. 2021, 11, 15825. [Google Scholar] [CrossRef] [PubMed]
- Rife, T.W.; Poland, J.A. Field book: An open-source application for field data collection on android. Crop Sci. 2014, 54, 1624–1627. [Google Scholar] [CrossRef]
- Eberhart, S.A.; Russell, W. Stability parameters for comparing varieties 1. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Pacheco, A.; Vargas, M.; Alvarado, G.; Rodríguez, F.; Crossa, J.; Burgueño, J. GEA-R (Genotype × Environment Analysis with R for Windows) Version 4.0; CIMMYT: Mexico City, Mexico, 2015. [Google Scholar]
- Yan, W. Singular-value partitioning in biplot analysis of multienvironment trial data. Agron. J. 2002, 94, 990–996. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists and Agronomists; CRC: Boca Raton, FL, USA, 2003; pp. 1–263. [Google Scholar] [CrossRef]
- Riaz, M.; Farooq, J.; Ahmed, S.; Amin, M.; Chattha, W.S.; Ayoub, M.; Kainth, R.A. Stability analysis of different cotton genotypes under normal and water-deficit conditions. J. Integr. Agric. 2019, 18, 1257–1265. [Google Scholar] [CrossRef]
- Popović, V.; Ljubičić, N.; Kostić, M.; Radulović, M.; Blagojević, D.; Ugrenović, V.; Popović, D.; Ivošević, B. Genotype× environment interaction for wheat yield traits suitable for selection in different seed priming conditions. Plants 2020, 9, 1804. [Google Scholar] [CrossRef]
- Tena, E.; Goshu, F.; Mohamad, H.; Tesfa, M.; Tesfaye, D.; Seife, A. Genotype × environment interaction by AMMI and GGE-biplot analysis for sugar yield in three crop cycles of sugarcane (Saccharum officinirum L.) clones in Ethiopia. Cogent Food Agric. 2019, 5, 1651925. [Google Scholar] [CrossRef]
- Sharma, S.P.; Leskovar, D.I.; Crosby, K.M.; Ibrahim, A. GGE biplot analysis of genotype-by-environment interactions for melon fruit yield and quality traits. Hortic. Sci. 2020, 55, 533–542. [Google Scholar] [CrossRef]
- Aruna, C.; Rakshit, S.; Shrotria, P.K.; Pahuja, S.K.; Jain, S.K.; Siva Kumar, S.; Modi, N.D.; Deshmukh, D.T.; Kapoor, R.; Patil, J.V. Assessing genotype-by-environment interactions and trait associations in forage Sorghum using GGE biplot analysis. J. Agric. Sci. 2016, 154, 73–86. [Google Scholar] [CrossRef]
- Moradi, Y.; Khadivi, A.; Salehi-Arjmand, H. Morphological and pomological characterizations of cornelian cherry (Cornus mas L.) to select the superior accessions. Sci. Hortic. 2019, 249, 208–218. [Google Scholar] [CrossRef]
- Dewi, N.; Nugroho, K.; Terryana, R.T.; Lestari, P. Evaluation of SSR and important agronomical characters of promising mutant lines of Soybean. Biodiversitas J. Biol. Divers. 2020, 21, 299–310. [Google Scholar] [CrossRef]
- Aremu, C.O.; Ojuederie, O.B.; Ayo-Vaughan, F.; Dahunsi, O.; Adekiya, A.O.; Olayanju, A.; Adebiyi, O.T.; Sunday, I.; Inegbedion, H.; Asaleye, A.J.; et al. Morphometric analysis and characterization of the nutritional quality in African yam bean accessions. Plant Physiol. Rep. 2019, 24, 446–459. [Google Scholar] [CrossRef]
- Qiao, L.; Wang, X.; Smith, P.; Fan, J.; Lu, Y.; Emmett, B.; Li, R.; Dorling, S.; Chen, H.; Liu, S. Soil quality both increases crop production and improves resilience to climate change. Nat. Clim. Chang. 2022, 12, 574–580. [Google Scholar] [CrossRef]
- Soane, B.; Van Ouwerkerk, C. Implications of soil compaction in crop production for the quality of the environment. Soil Tillage Res. 1995, 35, 5–22. [Google Scholar] [CrossRef]
- Sun, W.; Canadell, J.G.; Yu, L.; Yu, L.; Zhang, W.; Smith, P.; Fischer, T.; Huang, Y. Climate drives global soil carbon sequestration and crop yield changes under conservation agriculture. Glob. Chang. Biol. 2020, 26, 3325–3335. [Google Scholar] [CrossRef] [PubMed]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Oldfield, E.E.; Bradford, M.A.; Wood, S.A. Global meta-analysis of the relationship between soil organic matter and crop yields. Soil 2019, 5, 15–32. [Google Scholar] [CrossRef]
- Bedane, H.R.; Beketie, K.T.; Fantahun, E.E.; Feyisa, G.L.; Anose, F.A. The impact of rainfall variability and crop production on vertisols in the central highlands of Ethiopia. Environ. Syst. Res. 2022, 11, 26. [Google Scholar] [CrossRef]
- Helliwell, J.R.; Sturrock, C.J.; Miller, A.J.; Whalley, W.R.; Mooney, S.J. The role of plant species and soil condition in the structural development of the rhizosphere. Plant Cell Environ. 2019, 42, 1974–1986. [Google Scholar] [CrossRef] [PubMed]
- Chibarabada, T.P.; Modi, A.T.; Mabhaudhi, T. Adaptation and Productivity of Selected Grain Legumes in Contrasting Environments of Kwazulu-Natal, South Africa. Int. J. Plant Prod. 2018, 12, 169–180. [Google Scholar] [CrossRef]
- Hudson, A.I.; Odell, S.G.; Dubreuil, P.; Tixier, M.-H.; Praud, S.; Runcie, D.E.; Ross-Ibarra, J. Analysis of genotype-by-environment interactions in a maize mapping population. G3 Genes Genomes Genet. 2022, 12, jkac013. [Google Scholar] [CrossRef]
- Azrai, M.; Efendi, R.; Muliadi, A.; Aqil, M.; Suwarti; Zainuddin, B.; Syam, A.; Junaedi; Syah, U.T.; Dermail, A.; et al. Genotype by Environment Interaction on Tropical Maize Hybrids Under Normal Irrigation and Waterlogging Conditions. Front. Sustain. Food Syst. 2022, 6, 913211. [Google Scholar] [CrossRef]
- Esan, V.I.; Oke, G.O.; Ogunbode, T.O.; Obisesan, I.A. AMMI and GGE Biplot Analyses of Bambara Groundnut [Vigna subterranea (L.) Verdc.] for Agronomic Performances Under Three Environmental Conditions. Front. Plant Sci. 2023, 13, 997429. [Google Scholar] [CrossRef]
- Ullah, H.; Khalil, I.H.; Durrishahwar; Iltafullah; Khalil, I.A.; Fayaz, M.; Yan, J.; Ali, F. Selecting high yielding and stable mungbean [Vigna radiata (L.) Wilczek] genotypes using GGE biplot techniques. Can. J. Plant Sci. 2012, 92, 951–960. [Google Scholar] [CrossRef]
- Aktaş, H. Tracing highly adapted stable yielding bread Wheat (Triticum aestivum L.) genotypes for greatly variable South-Eastern Turkey. Appl. Ecol. Environ. Res. 2016, 14, 159–176. [Google Scholar] [CrossRef]
- Zhang, P.-P.; Song, H.; Ke, X.-W.; Jin, X.-J.; Yin, L.-H.; Liu, Y.; Qu, Y.; Su, W.; Feng, N.-J.; Zheng, D.-F.; et al. GGE biplot analysis of yield stability and test location representativeness in proso millet (Panicum miliaceum L.) genotypes. J. Integr. Agric. 2016, 15, 1218–1227. [Google Scholar] [CrossRef]
- Viera, W.; Brito, B.; Zambrano, E.; Ron, L.; Merino, J.; Campaña, D.; Álvarez, H. Genotype × Environment interaction in the yield and fruit quality of passion fruit germplasm grown in the Ecuadorian Littoral. Int. J. Fruit Sci. 2020, 20, S1829–S1844. [Google Scholar] [CrossRef]
- Khan, M.M.H.; Rafii, M.Y.; Ramlee, S.I.; Jusoh, M.; Al Mamun, M. AMMI and GGE biplot analysis for yield performance and stability assessment of selected Bambara groundnut (Vigna subterranea L. Verdc.) genotypes under the multi-environmental trials (METs). Sci. Rep. 2021, 11, 22791. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Nehe, A.; Akin, B.; Sanal, T.; Evlice, A.K.; Ünsal, R.; Dinçer, N.; Demir, L.; Geren, H.; Sevim, I.; Orhan, Ş.; et al. Genotype × environment interaction and genetic gain for grain yield and grain quality traits in Turkish spring wheat released between 1964 and 2010. PLoS ONE 2019, 14, e0219432. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Accessions | S/N | Accessions |
|---|---|---|---|
| 1 | TVSu-2188 | 16 | TVSu-2209 |
| 2 | TVSu-2190 | 17 | TVSu-2207 |
| 3 | TVSu-2193 | 18 | TVSu-2204 |
| 4 | TVSu-2194 | 19 | TVSu-2206 |
| 5 | TVSu-2199 | 20 | TVSu-2223 |
| 6 | TVSu-2200 | 21 | TVSu-2226 |
| 7 | TVSu-2201 | 22 | TVSu-2235 |
| 8 | TVSu-2184 | 23 | TVSu-2236 |
| 9 | TVSu-2202 | 24 | TVSu-2240 |
| 10 | TVSu-2181 | 25 | TVSu-2241 |
| 11 | TVSu-2285 | 26 | TVSu-2244 |
| 12 | TVSu-2284 | 27 | TVSu-2249 |
| 13 | TVSu-2256 | 28 | TVSu-2254 |
| 14 | TVSu-2221 | 29 | TVSu-2283 |
| 15 | TVSu-2218 | 30 | TVSu-2214 |
| Locations | pH (1:1) | bray P (mg/kg) | % OC | % N | % SAND | % CLAY | % SILT | Ca (cmol/kg) | Mg (cmol/kg) | K (cmol/kg) | Na (cmol/kg) | ppm Zn | ppm Cu | ppm Mn | ppm Fe |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mokwa | 5.51 | 1.73 | 0.05 | 0.01 | 83.00 | 10.67 | 6.33 | 0.91 | 0.27 | 0.02 | 0.02 | 2.49 | 0.43 | 41.24 | 1303.16 |
| Ibadan | 6.30 | 13.90 | 0.20 | 0.10 | 80.67 | 13.67 | 5.67 | 2.69 | 0.80 | 0.54 | 0.10 | 4.35 | 0.34 | 128.29 | 83.34 |
| Ikenne | 4.91 | 22.46 | 0.30 | 0.12 | 76.33 | 20.00 | 3.67 | 1.51 | 0.40 | 0.24 | 0.08 | 1.20 | 2.05 | 116.71 | 88.29 |
| Source of Variations | Df | Sum Sq. | Mean Sq. | F value | P r (>F) |
|---|---|---|---|---|---|
| Rep | 2 | 56,702,056 | 28,351,028 | 10.1142 | 6.912 × 10−5 *** |
| Env | 2 | 13,354,504 | 506,677,252 | 180.757 | <2.2 × 10−16 *** |
| Accns | 29 | 427,984,896 | 14,758,100 | 5.2649 | 1.043 × 10−12 *** |
| Env:Accns | 58 | 590,078,489 | 10,173,767 | 3.6295 | 2.547 × 10−11 *** |
| Residuals | 178 | 498,949,269 | 2,803,086 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linus, R.A.; Olanrewaju, O.S.; Oyatomi, O.; Idehen, E.O.; Abberton, M. Assessment of Yield Stability of Bambara Groundnut (Vigna subterranea (L.) Verdc.) Using Genotype and Genotype–Environment Interaction Biplot Analysis. Agronomy 2023, 13, 2558. https://doi.org/10.3390/agronomy13102558
Linus RA, Olanrewaju OS, Oyatomi O, Idehen EO, Abberton M. Assessment of Yield Stability of Bambara Groundnut (Vigna subterranea (L.) Verdc.) Using Genotype and Genotype–Environment Interaction Biplot Analysis. Agronomy. 2023; 13(10):2558. https://doi.org/10.3390/agronomy13102558
Chicago/Turabian StyleLinus, Rita Adaeze, Oluwaseyi Samuel Olanrewaju, Olaniyi Oyatomi, Emmanuel Ohiosinmuan Idehen, and Michael Abberton. 2023. "Assessment of Yield Stability of Bambara Groundnut (Vigna subterranea (L.) Verdc.) Using Genotype and Genotype–Environment Interaction Biplot Analysis" Agronomy 13, no. 10: 2558. https://doi.org/10.3390/agronomy13102558