
Optimizing Indoor Hemp Cultivation Efficiency through Differential Day–Night Temperature Treatment


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Measurement of Plant Growth Parameters
2.3. Measurement of Leaf Gas Exchange
2.4. Analysis of the Total Phenolic Compound Content and DPPH Scavenging Activity
2.5. Analysis of the Cannabinoid Content
2.6. Calculation of the Total Biomass Yield and the Total Cannabinoid Yield
2.7. Statistical Analysis
3. Results
3.1. Morphogenesis and Growth Parameters of Hemp
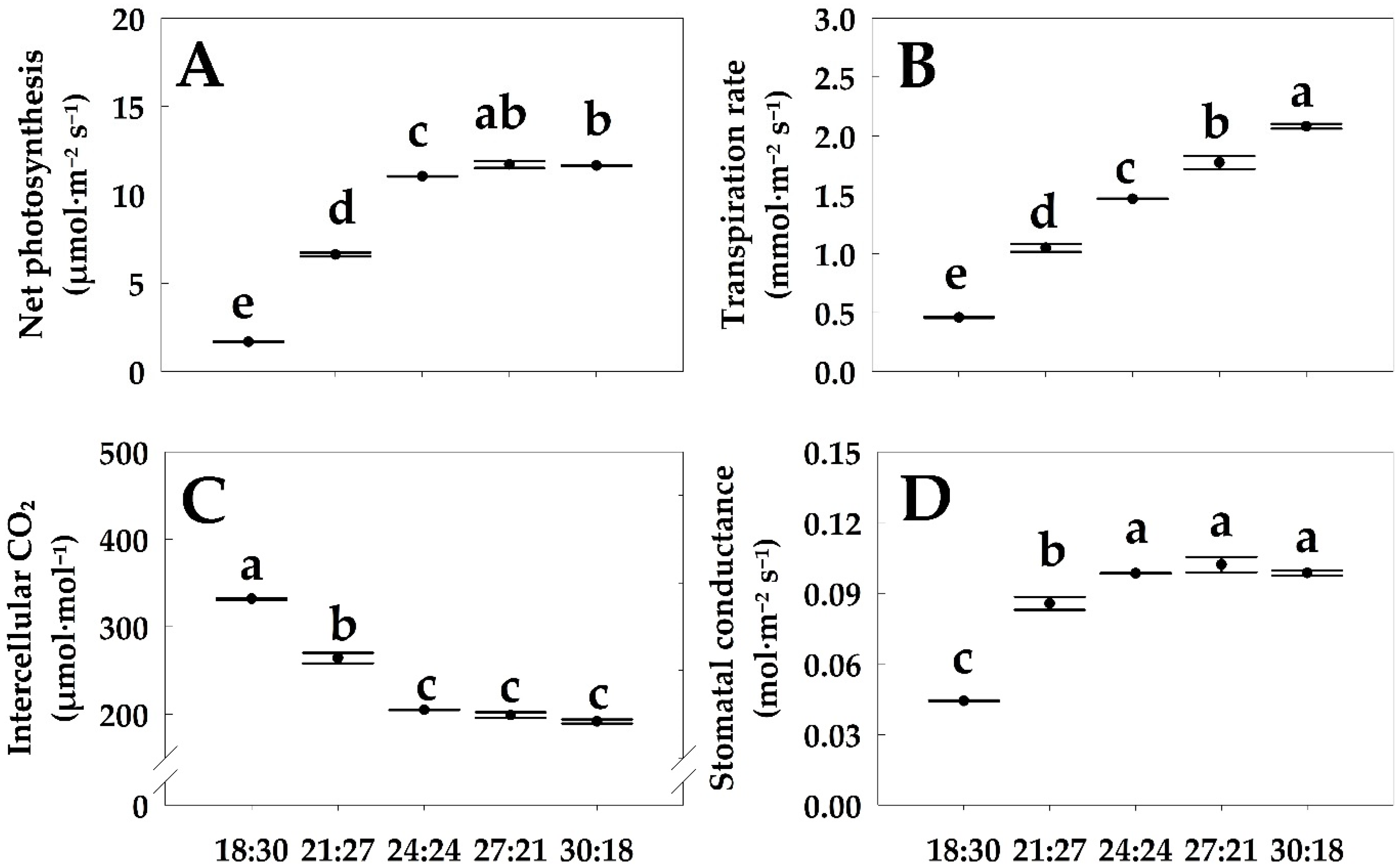
3.2. Photosynthesis-Related Parameters
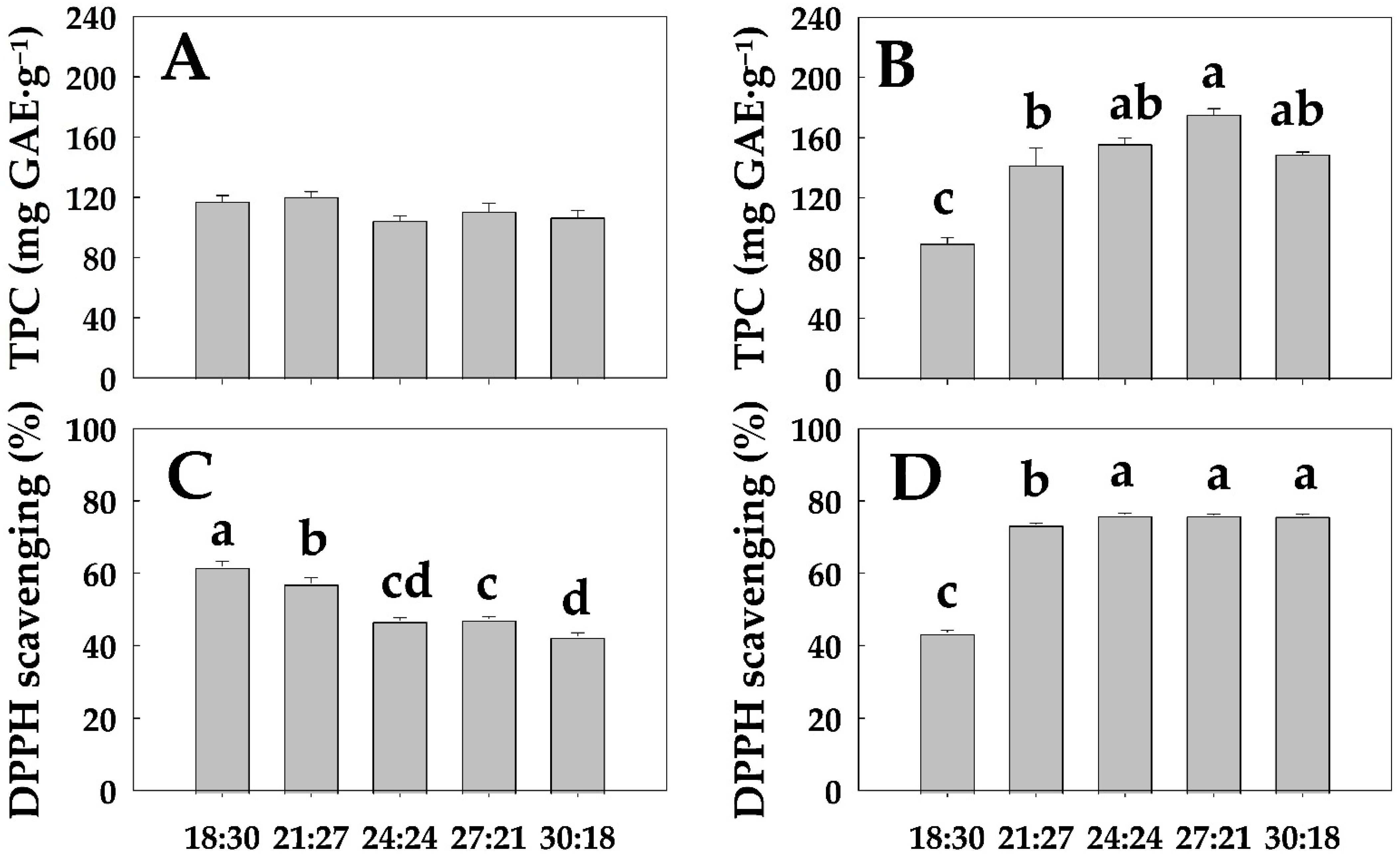
3.3. TPC Content and DPPH Scavenging Activity
3.4. Quantitative Analysis of Cannabinoids
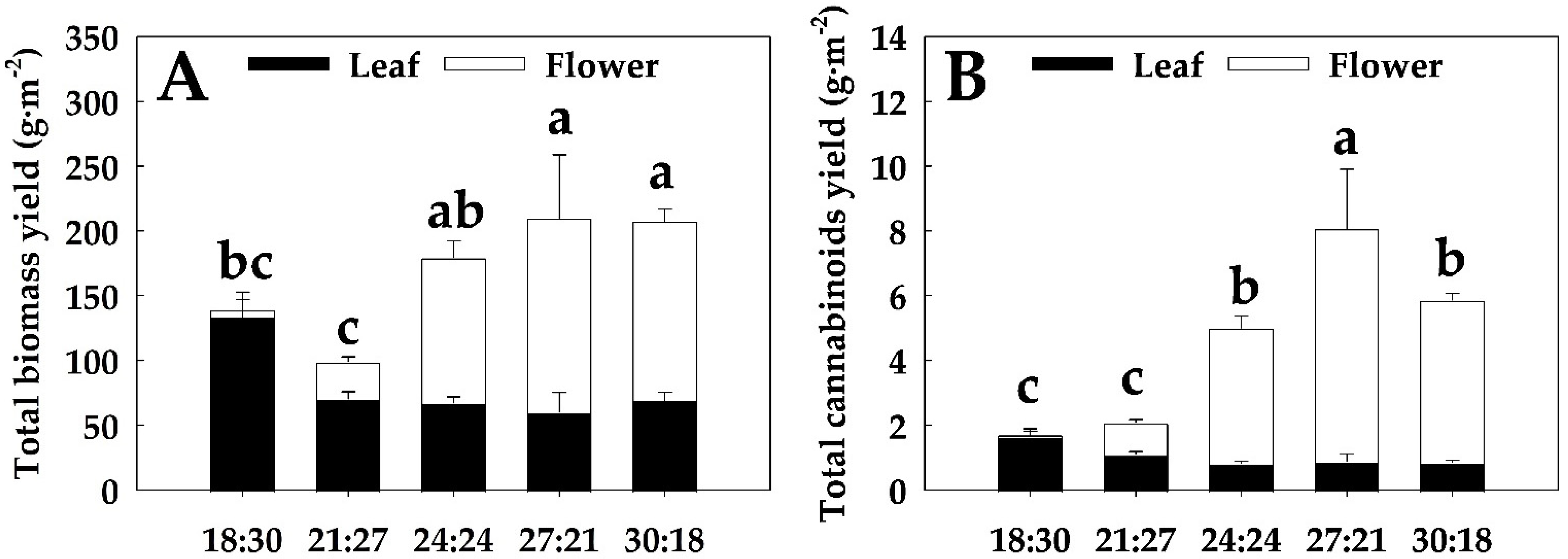
3.5. Total Biomass Yield and Total Cannabinoid Yield
4. Discussion
4.1. DIF Treatment Significantly Affects the Growth and Flowering of Hemp
4.2. Physiological Changes in Hemp Based on Its Photosynthetic Capacity in Relation to Temperature
4.3. Optimal DIF Conditions for Indoor Hemp Cultivation and the Practical Utilization Potential of Domestic Hemp
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrovska, B.B. Historical Review of Medicinal Plants’ Usage. Pharmacogn. Rev. 2012, 6, 1–5. [Google Scholar] [CrossRef]
- Small, E.; Cronquist, A. A Practical and Natural Taxonomy for Cannabis. Taxon 1976, 25, 405–435. [Google Scholar] [CrossRef]
- Johnson, R. Hemp as an Agricultural Commodity; Congressional Research Service: Washington, DC, USA, 2013.
- Jama, K.H. Medical Marijuana for Treatment of Chronic Pain and Other Medical and Psychiatric Problems: A Clinical Review. JAMA 2015, 313, 2474–2483. [Google Scholar] [CrossRef]
- Bahji, A.; Stephenson, C. International Perspectives on the Implications of Cannabis Legalization: A Systematic Review & Thematic Analysis. Int. J. Environ. Res. Public Health 2019, 16, 3095. [Google Scholar] [CrossRef]
- Palmieri, B.; Laurino, C.; Vadalà, M. Short-Term Efficacy of CBD-Enriched Hemp Oil in Girls with Dysautonomic Syndrome after Human Papillomavirus Vaccination. Isr. Med. Assoc. J. 2017, 19, 79–84. [Google Scholar]
- Schluttenhofer, C.; Yuan, L. Challenges Towards Revitalizing Hemp: A Multifaceted Crop. Trends Plant Sci. 2017, 22, 917–929. [Google Scholar] [CrossRef]
- Acosta, A.; Almirall, J. Differentiation between Hemp-Type and Marijuana-Type Cannabis Using the Fast Blue BB Colorimetric Test. Forensic. Chem. 2021, 26, 100376. [Google Scholar] [CrossRef]
- Simiyu, D.C.; Jang, J.H.; Lee, O.R. Understanding Cannabis sativa L.: Current Status of Propagation, Use, Legalization, and Haploid-Inducer-Mediated Genetic Engineering. Plants 2022, 11, 1236. [Google Scholar] [CrossRef]
- Monthony, A.S.; Page, S.R.; Hesami, M.; Jones, A.M.P. The Past, Present and Future of Cannabis sativa Tissue Culture. Plants 2021, 10, 185. [Google Scholar] [CrossRef]
- Kozai, T.; Niu, G.; Takagaki, M. Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Engler, N.; Krarti, M. Review of Energy Efficiency in Controlled Environment Agriculture. Renew. Sustain. Energy Rev. 2021, 141, 110786. [Google Scholar] [CrossRef]
- van Delden, S.H.; SharathKumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Kaiser, E.; Klamer, R.S.; Klerkx, L.; Kootstra, G.; et al. Current Status and Future Challenges in Implementing and Upscaling Vertical Farming Systems. Nat. Food 2021, 2, 944–956. [Google Scholar] [CrossRef]
- Moosavi-Nezhad, M.; Salehi, R.; Aliniaeifard, S.; Winans, K.S.; Nabavi-Pelesaraei, A. An Analysis of Energy Use and Economic and Environmental Impacts in Conventional Tunnel and LED-Equipped Vertical Systems in Healing and Acclimatization of Grafted Watermelon Seedlings. J. Clean Prod. 2022, 361, 132069. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global Food Demand and the Sustainable Intensification of Agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef]
- Folta, K.M.; Weber, E.A. Breeding New Varieties for Controlled Environments. Plant Biol. 2019, 21, 6–12. [Google Scholar] [CrossRef]
- Rademacher, W. Chemical Regulators of Gibberellin Status and Their Application in Plant Production. Annu. Plant Rev. Gibberellins 2016, 49, 359–404. [Google Scholar] [CrossRef]
- Ghosh, R.; Barbacci, A.; Leblanc-Fournier, N. Mechanostimulation: A Promising Alternative for Sustainable Agriculture Practices. J. Exp. Bot. 2021, 72, 2877–2888. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Khan, I.A.; Elsohly, M.A. Photosynthetic Response of Cannabis sativa L. to Variations in Photosynthetic Photon Flux Densities, Temperature and CO2 Conditions. Physiol. Mol. Biol. Plants 2008, 14, 299–306. [Google Scholar] [CrossRef]
- Gunn, S.; Farrar, J.F. Effects of a 4 °C Increase in Temperature on Partitioning of Leaf Area and Dry Mass, Root Respiration and Carbohydrates. Funct. Ecol. 1999, 13, 12–20. [Google Scholar] [CrossRef]
- Der Werf, H.M.G.V.; Brouwer, K.; Wijlhuizen, M.; Withagen, J.C.M. The Effect of Temperature on Leaf Appearance and Canopy Establishment in Fibre Hemp (Cannabis sativa L.). Ann. Appl. Biol. 1995, 126, 551–561. [Google Scholar] [CrossRef]
- Jin, D.; Jin, S.; Chen, J. Cannabis Indoor Growing Conditions, Management Practices, and Post-harvest Treatment: A Review. Am. J. Plant Sci. 2019, 10, 925–946. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Dusek, D.; Seigler, D.S.; Haney, A.W. Photosynthesis and Cannabinoid Content of Temperate and Tropical Populations of Cannabis sativa. Biochem. Syst. Ecol. 1975, 3, 15–18. [Google Scholar] [CrossRef]
- Song, J.; Yang, J.; Jeong, B.R. Difference between Day and Night Temperature (DIF) and Light Intensity Affect Growth and Photosynthetic Characteristics of Panax Ginseng Meyer Sprouts. Plants 2023, 12, 2820. [Google Scholar] [CrossRef] [PubMed]
- Moe, R.; Fjeld, T.; Mortensen, L.M. Stem Elongation and Keeping Quality in Poinsettia (Euphorbia pulcherrima Willd.) as Affected by Temperature and Supplementary Lighting. Sci. Hortic. 1992, 50, 127–136. [Google Scholar] [CrossRef]
- Shimizu, H. Effect of Day and Night Temperature Alternations on Plant Morphogenesis. Environ. Control Biol. 2007, 45, 259–265. [Google Scholar] [CrossRef]
- Grimstad, S.O.; Frimanslund, E. Effect of Different Day and Night Temperature Regimes on Greenhouse Cucumber Young Plant Production, Flower Bud Formation and Early Yield. Sci. Hortic. 1993, 53, 191–204. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Braca, A.; Fico, G.; Morelli, I.; De Simone, F.; Tomè, F.; De Tommasi, N. Antioxidant and Free Radical Scavenging Activity of Flavonol Glycosides from Different Aconitum species. J. Ethnopharmacol. 2003, 86, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Hahm, S.; Lee, B.; Bok, G.; Kim, S.; Park, J. Diniconazole Promotes the Yield of Female Hemp (Cannabis sativa) Inflorescence and Cannabinoids in a Vertical Farming System. Agronomy 2023, 13, 1497. [Google Scholar] [CrossRef]
- Ohtaka, K.; Yoshida, A.; Kakei, Y.; Fukui, K.; Kojima, M.; Takebayashi, Y.; Yano, K.; Imanishi, S.; Sakakibara, H. Difference between Day and Night Temperatures Affects Stem Elongation in Tomato (Solanum Lycopersicum) Seedlings via Regulation of Gibberellin and Auxin Synthesis. Front. Plant Sci. 2020, 11, 577235. [Google Scholar] [CrossRef]
- Kim, Y.H.; Yang, H.C.; Bae, Y.H.; Hyeon, S.J.; Hwang, S.J.; Kim, D.H.; Jang, D.C. Preventing Overgrowth of Cucumber and Tomato Seedlings Using Difference between Day and Night Temperature in a Plant Factory with Artificial Lighting. Plants 2023, 12, 3164. [Google Scholar] [CrossRef]
- Grindal, G.; Ernstsen, A.; Reid, J.B.; Junttila, O.; Lindgård, B.; Moe, R. Endogenous Gibberellin A1 Levels Control Thermoperiodic Stem Elongation in Pisum sativum. Physiol. Plant 1998, 102, 523–531. [Google Scholar] [CrossRef]
- Islam, N.; Wold, A.-B.; Torre, S.; Gislerød, H.R. Effects of Temperature DIF and Drop on the Growth, Quality, Total Phenolic Content and Antioxidant Activity of Herbs. J. Multidiscip. Eng. Sci. Stud. 2016, 2, 266–271. [Google Scholar]
- Myster, J.; Moe, R. Effect of Diurnal Temperature Alternations on Plant Morphology in Some Greenhouse Crops—A Mini Review. Sci. Hortic. 1995, 62, 205–215. [Google Scholar] [CrossRef]
- Lucidos, J.G.; Ryu, K.B.; Younis, A.; Kim, C.K.; Hwang, Y.J.; Son, B.G.; Lim, K.B. Different Day and Night Temperature Responses in Lilium hansonii in Relation to Growth and Flower Development. Hortic. Environ. Biotechnol. 2013, 54, 405–411. [Google Scholar] [CrossRef]
- Thingnaes, E.; Torre, S.; Ernstsen, A.; Moe, R. Day and Night Temperature Responses in Arabidopsis: Effects on Gibberellin and Auxin Content, Cell Size, Morphology and Flowering Time. Ann. Bot. 2003, 92, 601–612. [Google Scholar] [CrossRef]
- Blázquez, M.A.; Ahn, J.H.; Weigel, D. A Thermosensory Pathway Controlling Flowering Time in Arabidopsis Thaliana. Nat. Genet. 2003, 33, 168–171. [Google Scholar] [CrossRef]
- Ye, S.; Shao, Q.; Xu, M.; Li, S.; Wu, M.; Tan, X.; Su, L. Effects of Light Quality on Morphology, Enzyme Activities, and Bioactive Compound Contents in Anoectochilus Roxburghii. Front. Plant Sci. 2017, 8, 266635. [Google Scholar] [CrossRef]
- Alexander, J.D.; Donnelly, J.R.; Shane, J.B. Photosynthetic and Transpirational Responses of Red Spruce Understory Trees to Light and Temperature. Tree Physiol. 1995, 15, 393–398. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Khan, I.A.; ElSohly, M.A. Temperature Response of Photosynthesis in Different Drug and Fiber Varieties of Cannabis sativa L. Physiol. Mol. Biol. Plants 2011, 17, 297–303. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Cerling, T.E. Atmospheric CO2 and the Ratio of Intercellular to Ambient CO2 Concentrations in Plants. Tree Physiol. 1995, 15, 105–111. [Google Scholar] [CrossRef]
- Kong, L.; Wen, Y.; Jiao, X.; Liu, X.; Xu, Z. Interactive Regulation of Light Quality and Temperature on Cherry Tomato Growth and Photosynthesis. Environ. Exp. Bot. 2021, 182, 104326. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic Compounds in Plants and Agri-Industrial by-Products: Antioxidant Activity, Occurrence, and Potential Uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Tian, S.; Nakamura, K.; Kayahara, H. Analysis of Phenolic Compounds in White Rice, Brown Rice, and Germinated Brown Rice. J. Agric. Food Chem. 2004, 52, 4808–4813. [Google Scholar] [CrossRef]
- Ho, K.Y.; Tsai, C.C.; Chen, C.P.; Huang, J.S.; Lin, C.C. Screening of Brazilian Plant Extracts for Antioxidant Activity by the Use of DPPH Free Radical Method. Phytother. Res. 2001, 15, 127–130. [Google Scholar] [CrossRef]
- Jeong, S.W.; Kim, G.S.; Lee, W.S.; Kim, Y.H.; Kang, N.J.; Jin, J.S.; Lee, G.M.; Kim, S.T.; Abd El-Aty, A.M.; Shim, J.H.; et al. The Effects of Different Night-Time Temperatures and Cultivation Durations on the Polyphenolic Contents of Lettuce: Application of Principal Component Analysis. J. Adv. Res. 2015, 6, 493–499. [Google Scholar] [CrossRef]
- Izzo, L.; Castaldo, L.; Narváez, A.; Graziani, G.; Gaspari, A.; Rodríguez-Carrasco, Y.; Ritieni, A. Analysis of Phenolic Compounds in Commercial Cannabis sativa L. Inflorescences Using UHPLC-Q-Orbitrap HRMS. Molecules 2020, 25, 631. [Google Scholar] [CrossRef] [PubMed]
- Hanuš, L.O.; Meyer, S.M.; Muñoz, E.; Taglialatela-Scafati, O.; Appendino, G. Phytocannabinoids: A Unified Critical Inventory. Nat. Prod. Rep. 2016, 33, 1357–1392. [Google Scholar] [CrossRef]
- Radwan, M.M.; Chandra, S.; Gul, S.; Elsohly, M.A. Cannabinoids, Phenolics, Terpenes and Alkaloids of Cannabis. Molecules 2021, 26, 2774. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.K.; Rahman, M.H.; Cheong, E.J.; Lim, J.D.; Lim, Y.S. Cannabinoids Accumulation in Hemp (Cannabis sativa L.) Plants under Led Light Spectra and Their Discrete Role as a Stress Marker. Biology 2021, 10, 710. [Google Scholar] [CrossRef]
- Pisanti, S.; Malfitano, A.M.; Ciaglia, E.; Lamberti, A.; Ranieri, R.; Cuomo, G.; Abate, M.; Faggiana, G.; Proto, M.C.; Fiore, D.; et al. Cannabidiol: State of the Art and New Challenges for Therapeutic Applications. Pharmacol. Ther. 2017, 175, 133–150. [Google Scholar] [CrossRef]
- Ladin, D.A.; Soliman, E.; Griffin, L.T.; Van Dross, R. Preclinical and Clinical Assessment of Cannabinoids as Anti-cancer Agents. Front. Pharmacol. 2016, 7, 361. [Google Scholar] [CrossRef] [PubMed]
- Karler, R.; Cely, W.; Turkanis, S.A. The Anticonvulsant Activity of Cannabidiol and Cannabinol. Life Sci. 1973, 13, 1527–1531. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.E.; Elsohly, M.A. Biological Activity of Cannabichromene, Its Homologs and Isomers. J. Clin. Pharmacol. 1981, 21, 283S–291S. [Google Scholar] [CrossRef] [PubMed]
- U.S. Congress. S.2667-Hemp Farming Act of 2018. S.2667—115th Congress (2017–2018). Available online: https://www.congress.gov/bill/115th-congress/senate-bill/2667/text#:~:text=2667%20%2d%20115th%20congress%20(2017%2d,congress.gov%20%7c%20library%20of%20congress (accessed on 12 September 2023).
- Visković, J.; Zheljazkov, V.D.; Sikora, V.; Noller, J.; Latković, D.; Ocamb, C.M.; Koren, A. Industrial Hemp (Cannabis sativa L.) Agronomy and Utilization: A Review. Agronomy 2023, 13, 931. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Stage | Temperature (DT:NT z, °C) | Humidity (%) | CO2 Concentration (μmol·mol−1) | PPFD y (μmol·m−2·s−1) | Photoperiod (DT:NT, h) | Cultivation Period (week) | Electrical Conductivity (dS·m−1), pH |
|---|---|---|---|---|---|---|---|
| Rooting | 24 | 80 | 400 | 100 | 20:4 | 3 | 1.0, 5.8 |
| Vegetative | 24 | 60 | 400 | 200 | 20:4 | 1 | 1.5, 5.8 |
| Reproductive | 18:30 21:27 24:24 27:21 30:18 | 40 | 400 | 400 ~ 700 | 12:12 | 7 | 2.0, 5.8 ~ 2.5, 5.8 |
| DIF Treatment | Shoot Width (cm) | Shoot Height (cm) | Leaf Number | Leaf Area (cm2) | Flower Number | SPAD Value |
|---|---|---|---|---|---|---|
| 18:30 z | 46.55 b y | 55.98 bc | 715.9 a | 4440 a | 100.7 b | 56.73 |
| 21:27 | 56.85 a | 77.85 a | 548.0 ab | 3519 a | 209.3 a | 56.70 |
| 24:24 | 45.00 bc | 58.49 b | 202.0 b | 1568 b | 88.3 b | 65.23 |
| 27:21 | 36.95 cd | 53.69 bc | 161.3 b | 1039 b | 70.7 b | 64.70 |
| 30:18 | 34.31 d | 47.17 c | 150.0 b | 822 b | 56.0 b | 52.53 |
| Significance x | *** | *** | *** | *** | *** | n.s |
| DIF Treatment | Fresh Weight (g) | Dry Weight (g) | ||||||
|---|---|---|---|---|---|---|---|---|
| Stem | Leaf | Flower | Total Shoot | Stem | Leaf | Flower | Total Shoot | |
| 18:30 z | 56.49 a y (38.2) x | 87.39 a (59.2) | 3.85 c (2.6) | 147.73 ab (100) | 11.50 ab (32.7) | 22.76 a (64.7) | 0.93 d (2.6) | 35.18 ab (100) |
| 21:27 | 70.41 a (41.6) | 66.17 a (39.1) | 32.65 b (19.3) | 169.22 a (100) | 16.07 a (38.5) | 18.69 a (44.8) | 6.99 c (16.7) | 41.76 a (100) |
| 24:24 | 30.46 b (21.0) | 35.02 b (24.2) | 79.50 a (54.8) | 144.98 ab (100) | 8.36 bc (22.6) | 10.38 b (28.1) | 18.23 a (49.3) | 36.97 a (100) |
| 27:21 | 23.53 b (19.5) | 24.57 b (20.4) | 72.29 a (60.0) | 120.39 ab (100) | 6.45 c (21.0) | 7.08 b (23.1) | 17.17 ab (55.9) | 30.69 ab (100) |
| 30:18 | 19.88 b (19.4) | 24.71 b (24.1) | 58.03 a (56.5) | 102.62 b (100) | 4.91 c (20.1) | 6.60 b (27.0) | 12.95 b (52.9) | 24.46 b (100) |
| Significance w | *** | *** | *** | *** | *** | *** | *** | *** |
| DIF Treatment | CBDA y (mg·g−1) | CBCA (mg·g−1) | THCA (mg·g−1) | CBGA (mg·g−1) | CBL (mg·g−1) | CBT (mg·g−1) | CBC (mg·g−1) | CBG (mg·g−1) | CBD (mg·g−1) | THC (mg·g−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| 18:30 z | 9.7 c x | 0.96 | 0.643 b | 0.206 bc | 0.215 a | 0.224 b | 0.198 b | 0.0526 b | 0.092 c | 0.0828 |
| 21:27 | 12.9 a | 0.86 | 0.699 a | 0.247 a | 0.212 a | 0.344 a | 0.195 b | 0.0691 a | 0.079 c | 0.0899 |
| 24:24 | 9.9 c | 0.82 | 0.584 c | 0.175 d | 0.195 b | 0.348 a | 0.208 a | 0.0562 b | 0.083 c | 0.0869 |
| 27:21 | 11.7 b | 1.03 | 0.652 b | 0.218 b | 0.215 a | 0.368 a | 0.199 b | 0.0660 a | 0.116 b | 0.0900 |
| 30:18 | 9.4 c | 0.91 | 0.598 c | 0.192 cd | 0.216 a | 0.393 a | 0.200 b | 0.0670 a | 0.151 a | 0.0893 |
| Significance w | *** | n.s. | *** | *** | ** | *** | *** | *** | *** | n.s. |
| DIF Treatment | CBDA y (mg·g−1) | CBCA (mg·g−1) | THCA (mg·g−1) | CBGA (mg·g−1) | CBL (mg·g−1) | CBT (mg·g−1) | CBC (mg·g−1) | CBG (mg·g−1) | CBD (mg·g−1) | THC (mg·g−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| 18:30 z | 11.6 e x | 0.89 d | 0.76 d | 0.32 e | 0.230 e | 0.341 ab | 0.224 e | 0.040 d | 0.075 e | 0.077 d |
| 21:27 | 29.2 d | 1.89 c | 1.33 c | 1.26 a | 0.370 d | 0.326 b | 0.240 d | 0.137 c | 0.096 d | 0.080 d |
| 24:24 | 31.3 b | 1.81 c | 1.50 b | 1.05 b | 0.398 c | 0.332 b | 0.252 c | 0.185 b | 0.141 c | 0.087 c |
| 27:21 | 40.3 a | 2.60 a | 2.12 a | 0.99 c | 0.519 a | 0.390 a | 0.302 a | 0.293 a | 0.231 a | 0.110 a |
| 30:18 | 30.3 c | 2.13 b | 1.52 b | 0.91 d | 0.429 b | 0.360 ab | 0.258 b | 0.196 b | 0.214 b | 0.095 b |
| Significance w | *** | *** | *** | *** | *** | ** | *** | *** | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bok, G.; Hahm, S.; Shin, J.; Park, J. Optimizing Indoor Hemp Cultivation Efficiency through Differential Day–Night Temperature Treatment. Agronomy 2023, 13, 2636. https://doi.org/10.3390/agronomy13102636
Bok G, Hahm S, Shin J, Park J. Optimizing Indoor Hemp Cultivation Efficiency through Differential Day–Night Temperature Treatment. Agronomy. 2023; 13(10):2636. https://doi.org/10.3390/agronomy13102636
Chicago/Turabian StyleBok, Gwonjeong, Seungyong Hahm, Juhyung Shin, and Jongseok Park. 2023. "Optimizing Indoor Hemp Cultivation Efficiency through Differential Day–Night Temperature Treatment" Agronomy 13, no. 10: 2636. https://doi.org/10.3390/agronomy13102636
APA StyleBok, G., Hahm, S., Shin, J., & Park, J. (2023). Optimizing Indoor Hemp Cultivation Efficiency through Differential Day–Night Temperature Treatment. Agronomy, 13(10), 2636. https://doi.org/10.3390/agronomy13102636