1. Introduction
With the development of greenhouses in agriculture, plants can be cultivated by providing the required environmental conditions for better growth and sustainable food production [
1]. Furthermore, greenhouses allow plants to be grown in any season that would otherwise inhibit their growth. Greenhouse technologies have promoted science-based solutions for optimal plant production by adjusting internal climatic factors, which creates more economic value [
2].
In recent years, climate change and extreme weather events, such as increasingly frequent heat stress, have increased [
3,
4], and high-temperature stress on plants from the internal environments of greenhouses is expected to affect plant growth and development [
5]. During the summer in South China, the air temperature inside greenhouses reaches 40 °C or more. This temperature-increasing trend may seriously affect average plant growth [
6] because the air temperature increases the heat load on the plants. An extremely high temperature also increases plant water vapor demand, leading to higher leaf transpiration rates and stomatal closure, which results in higher leaf temperature and a decrease in net photosynthetic rate [
7]. Thus, plant growth is inhibited by several factors, such as leaf wilting or curling, which can affect plant yield and quality [
8,
9]. In addition, because of the above issue, it is essential to cool the air temperature inside greenhouses, which requires an appropriate method to cool the air temperature around the plant canopy. The current common cooling methods used in greenhouses include natural ventilation, shade nets, and wet curtain fans, but these methods have several drawbacks. In addition, natural ventilation is ineffective when the temperature difference between the indoor and outdoor environment is significantly higher, sunshades will affect normal photosynthesis in crops, and wet curtain fans are costly and difficult to maintain. Evaporative cooling can effectively reduce the increasing heat of plants and drought stresses [
10]. Micro-spray, with its evaporative cooling characteristics, is one of the most commonly used irrigation methods to improve the growing environment of crops under greenhouse conditions [
11,
12]. A micro-spray cooling system that applies water near the plant canopy can enhance evaporative cooling during periods of heat stress in plant growth [
13]. Previously, the micro-spray cooling system was investigated in many studies worldwide on different plants.
Micro-spray reduces air temperature and increases the relative humidity [
10], photosynthetic rate, and stomatal conductance [
14,
15] and increases the effective quantum yield of PSII plants [
6]. However, the effects of micro-spray on plant transpiration rates can vary. Previously, it was found that it reduced maize and jujube transpiration rates by 0.75–1.03 mmol m
−2s
−1 and 0.61–0.66 mmol m
−2s
−1, respectively [
16]. Still, one study showed that micro-spray increased globe artichoke transpiration rates by about 0.84 mmol m
−2s
−1 [
15].
Cucumbers (
Cucumis sativus L.) and tomatoes (
Solanum lycopersicum L.) are essential horticultural crops planted in greenhouses globally. In a greenhouse, fluctuations in air temperature can cause heat stress and induce leaf and stem wilting, which has a significant negative effect on average growth, quality, and photosynthesis in cucumbers (air temperature over 30 °C) and tomatoes (air temperature over 35 °C) [
17,
18]. The susceptibility of cucumbers and tomatoes to high temperatures varies according to genotype and growth stage [
19]. So, studying the physiological effects of micro-spray on different crops is essential in designing a proper and scientific micro-spray system.
In a micro-spray system, the amount of water sprayed can be small in the form of droplets, as the aim is not to wet the rooting zone but to increase the evaporative cooling around the crops. These droplets can potentially reduce the amount of irrigation but not enough to meet a plant’s irrigation needs [
20]. Therefore, micro-spray can be used as a supplementary irrigation method with drip irrigation for plants in greenhouses to avoid high-temperature stresses. Drip and micro-spray irrigation have recently been used in China to improve crop growth and regulate the micro-climate in greenhouses [
21,
22]. For this combined irrigation method, the operating time of micro-spray needs to be explored. Therefore, rapid feedback and accurate techniques, such as chlorophyll fluorescence technology, should be used to detect plant physiology responses to micro-spray under heat-stress conditions.
Chlorophyll fluorescence technology has been used to study the distribution of excitation energy inside the photosynthetic apparatus. It is used to explore the mechanism and regulation of photosynthesis as an indicator to assess plant growth responses [
23,
24]. It is also applied to monitor the physiological status of crops and discover the effects of biotic and abiotic stresses on crops [
25]. For abiotic stresses, such as heat stress, several studies have shown that chlorophyll fluorescence analysis can be used as a sensitive indicator of chloroplast membrane damage and changes in the function of the photosynthetic apparatus [
26]. Chlorophyll fluorescence parameters, such as the effective quantum yield of PSII (
ΦPSII), reflect the photosynthetic efficiency measured under natural light conditions and are excellent parameters for monitoring heat stresses [
27]. Studies have shown that photosynthetically active radiation (PAR) is the most critical factor influencing the
ΦPSII, and there is a negative and significant correlation between PAR and the
ΦPSII [
6]. The K-means clustering algorithm is considered one of the research community′s most influential and popular data mining algorithms. It has the remarkable features of being fast and easy to implement [
28,
29]. The K-means clustering algorithm has been applied in agriculture, where it is used to determine the change in moisture content corresponding to different temperature ranges [
30].
Therefore, the objectives of this study are as follows: (1) to investigate the effects of combined micro-spray and drip irrigation (MSDI) and drip irrigation (DI) on plant growth and fruiting; (2) to analyze the effects of MSDI and DI on daily changes in the net leaf photosynthetic rate (Pn), the transpiration rate (Tr), and stomatal conductance (Gs) at the leaf scale; (3) to study the effect of PAR on the effective quantum yield of PSII (ΦPSII) of crops using K-means cluster analysis; and (4) to explore the specific effect of micro-spray on the ΦPSII of crops under MSDI and DI.
2. Materials and Methods
2.1. Experimental Site
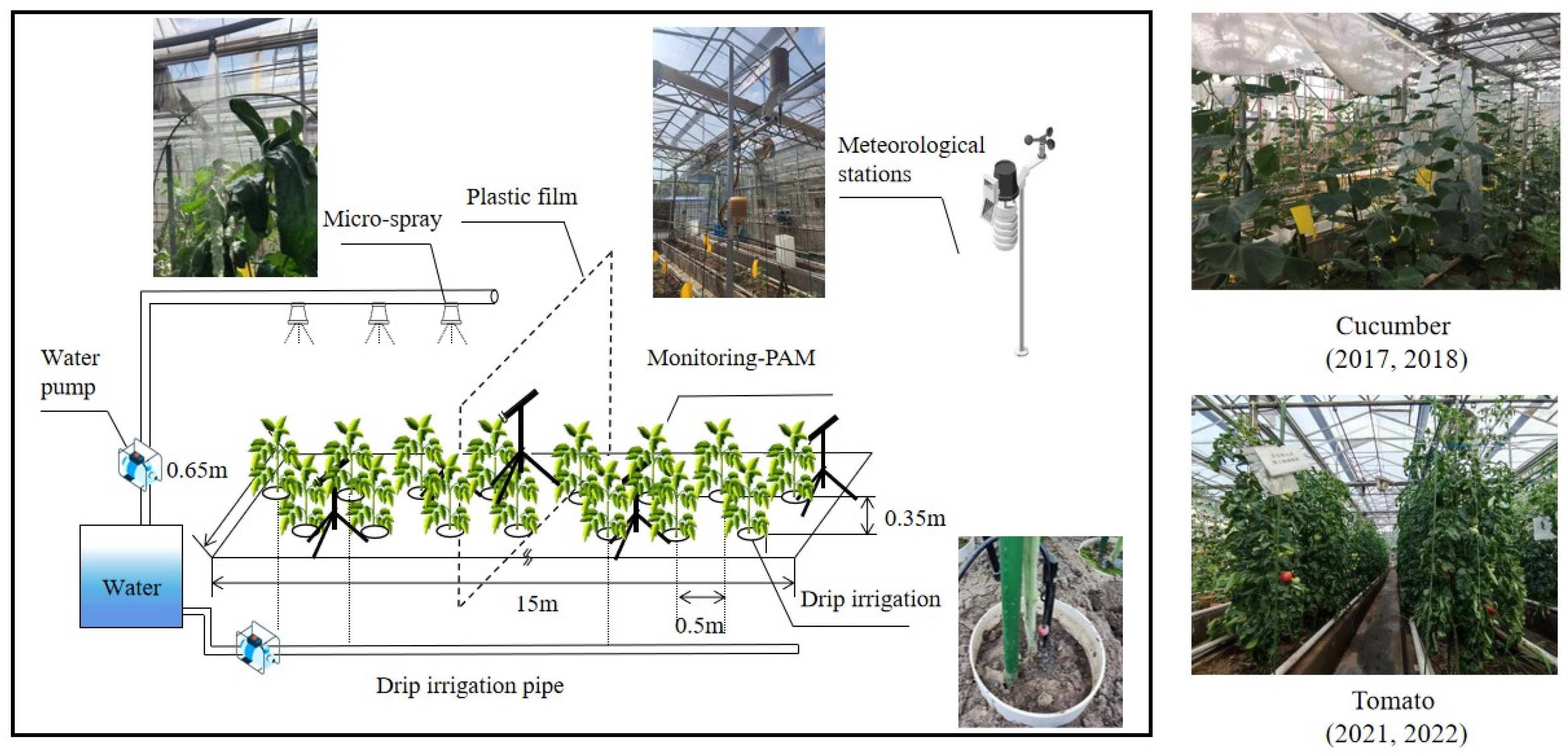
The experiment was conducted in a greenhouse in Zhenjiang, Jiangsu Province, China (32°20′ N, 119°45′ E), for two growing seasons of cucumber, in 2017 and 2018 (transplanted in April and harvested in July), and tomato, in 2021 and 2022 (transplanted in March and harvested in July). The greenhouse is 32 m in length, 20 m in width, and 3.8 m in height, with the longer side oriented east–west and covered with 4 mm float glass with a transmittance greater than 89%. The greenhouse has 5 soil troughs for growing plants (1500, 65, and 30 cm in length, width, and depth, respectively), with an aisle (0.85 m in width) between each soil trough. In the summer, the maximum air temperature outside the greenhouse exceeds 30 °C, and the air temperature inside reaches over 40 °C. The daily irrigation of plants in the four-year experimental treatments was based on the daily evaporation data measured with the evaporation pans (20 cm in diameter).
2.2. Experimental Design
The experiment was conducted using cucumber (Cucumis sativus L., variety Youliang 3–2) and tomato (Solanum lycopersicum L, variety Hezuo 908) in four growing seasons (two growing seasons each for cucumber (2017 and 2018) and tomato (2021 and 2022)) under two treatments: drip irrigation (DI) and micro-spray combined with drip irrigation (MSDI). We set up the experimental area to coincide with the cycle of local production activities so that the results of the trials could be applied to actual production activities. Also, the fertigation for cucumber and tomato plants under DI and MSDI was applied as per the local practice at four different growing stages.
Cucumber seeds were sown in transplant trays on 21 February 2017 and 25 February 2018, and then 32 seedlings were transplanted into soil troughs on 6 April 2017 and 11 April 2018, respectively. Moreover, tomato seeds were sown in transplant trays on 26 January 2021 and 13 January 2022. Then, 50 seedlings were transplanted into soil troughs on 20 March 2021 and 1 March 2022, respectively. Micro-spray was applied for cucumbers 30 days after transplanting (DAT) in 2017 and 25 DAT in 2018, and for tomatoes, 40 DAT in 2021 and 55 DAT in 2022. However, the plant densities were maintained at 3.3 plants/m2 for cucumber and 5.1 plants/m2 for tomato. In addition, the plants were separated by placing plastic film in the middle of the DI and MSDI treatments.
The drip irrigation system was used to supply water to tomato and cucumber plants throughout the growing seasons during four stages: seedling, flowering, fruiting, and fruit picking. The amount of applied water was 40 mm and was supplied in 2–5 days intervals after transplanting to ensure deeper roots and vigorous plant growth. The daily irrigation volume of the drip irrigation system was referred to as the cumulative pan evaporation (
Epan), measured using two evaporation pans (20 cm in diameter). The pans were placed inside the greenhouse and 5 cm above the plant canopy at two locations [
31]. The evaporated volume from each pan was measured at 6:00 pm daily. Before the next reading, the pans were cleaned and refilled with fresh water up to 20 mm depth.
In addition, for MSDI treatment, atomization nozzles were used to micro-spray the water. The nozzles were placed 2.5 m above the soil surface. The droplet sizes of the selected nozzles were 65 µm, and the flow rate of the nozzles was 7.5 L/h. For MSDI, amount of drip irrigation was same with DI treatment. The nozzle spraying time and spraying interval time were divided into different levels: for cucumbers (2017 and 2018), the spraying time was 2 min, four times a day (after each hour spraying interval at 9:00, 10:00, 11:00, and 12:00). For tomatoes (2021 and 2022), the spraying time was 1 min, seven times a day (after each hour spraying interval at 9:00, 10:00, 11:00, 12:00, 13:00, 14:00, and 15:00. The nozzles were only opened when the air temperature was above 30 °C (named “high temperature”) [
6].
On the other hand, no additional cooling methods were used during the experiment apart from natural ventilation, which ensured that the control variables were observed. The experimental setup is shown in
Figure 1.
2.3. Measurements
2.3.1. Meteorological Data
The average daily air temperature (
Ta) and relative humidity (
RH) inside the greenhouse were collected using a data logger (CR1000, Campbell, Logan, UT, USA). The photosynthetically active radiation (PAR) was measured using a PQS 1 (Kipp & Zonen, Delft, The Netherlands), mounted 2 m above the soil surface. The air temperature and relative humidity probes (HMP155A, Campbell, Logan, UT, USA) were set at the same height as the plant canopy and adjusted over plant growth to measure the air temperature and relative humidity near the plants from both treatments. The accuracy of the data recording sensors was 0.1 °C for air temperature and ±1.7% for relative humidity. All data were collected and averaged for 10 min. The environmental factors recorded during the experimental period are shown in
Figure 2.
2.3.2. Plant Growth, Yield, Total Soluble Solids, Irrigation Water Use Efficiency, and Fruit Shape Index
The plant height and stem diameter of plants grown under DI and MSDI were measured at the end of the experiments. Plant height (cm) was measured with a measuring tape. Stem diameter (mm) was measured at a marked place (5 cm above the soil surface) with a vernier caliper. The yield of plants was calculated using a weight balance (accuracy 0.1 g). For total soluble solids (TSS, degrees Brix), three fruits per plant from the same ripening batch with no external defects were selected and tested using the abbe refractometer (WYA-3S, Yidian, Shanghai, China) method. Before the test, the fruits were peeled and pressed, and the paste was filtered through four layers of fine cotton gauze. Each sample was measured three times to minimize observation errors. In addition, the irrigation water use efficiency was calculated using the following equation:
where
WUE is irrigation water use efficiency (kg/m
3),
is tomato yield (t/ha), and
is irrigation water amount (mm). The weight per fruit is the average value of each cucumber and tomato plant.
The fruit shape index is the ratio of the longitudinal to the transverse diameter of the fruit and was measured with an electronic vernier caliper (accuracy of 0.01 mm) and a meter ruler (accuracy of 1 mm).
2.3.3. Leaf Gas Exchange
During the micro-spray irrigation treatment, a photosynthesis system (GFS-3000, WALZ, Effeltrich, Germany) was used to measure the rate of photosynthesis (Pn), the rate of water loss (Tr), and the conductance of the stomata (Gs). The Pn, Tr, and Gs were measured on a clear, cloudless, sunny day during both the cucumber and tomato ripening and harvesting periods. For measuring leaf gas exchange parameters, six plants with uniform size and growth were randomly selected; further, three plant leaves were randomly chosen from the fifth functional leaf below the top leaf of each plant. Each leaf was measured three times, and the average values were taken every 2 h between 08:00 and 18:00. The RH and leaf Tl in the Walz leaf cuvette are the same as the environment in which the crop is located. It ensures that the photosynthetic rate is measured under actual conditions because the CO2 variable in the greenhouse does not vary much during the day (around 466 ppm). To avoid interference by human movement, we used a CO2 buffer tank to ensure that the ambient CO2 variable is the same when measuring the gas exchange under MSDI and DI.
2.3.4. Chlorophyll Fluorescence, Photosynthetically Active Radiation, and Leaf Temperature
Chlorophyll fluorescence parameters, including the effective quantum yield of PSII (
ΦPSII and Equation (2)), were monitored using a PAM WinControl-3 controlled MONI-PAM multi-channel chlorophyll fluorometer (Monitoring-PAM, WALZ, Effeltrich, Germany). The chlorophyll fluorometer consists of 4 measuring heads, a data collection system, and a personal computer interface box. Two plants under DI and MSDI were selected to monitor the chlorophyll fluorescence parameters. The fifth functional leaf below the top leaf of two randomly chosen plants was used for the measurements. The selected plants were changed every morning before 9:00 am to eliminate individual differences. The photosynthetically active radiation (PAR) and leaf temperature (
Tl) were also recorded with
ΦPSII.
ΦPSII can reflect the actual photosynthetic efficiency measured under natural light conditions. The MONI-PAM data collector was set up to collect data automatically every 10 min.
where
ΦPSII is the effective quantum yield of photosystem II,
F’m is the maximal fluorescence yield, and
F’s is the steady-state fluorescence yield.
2.3.5. K-Means Clustering Algorithm
The K-means clustering algorithm is an unsupervised algorithm usually used to classify an area. This study used the K-means clustering algorithm to classify changes in the
ΦPSII in different PARs. Clustering takes
k points in the space as the centroid and minimizes the sum of squares of the distance from each data point to its assigned cluster center [
32]. The centroid value of each cluster is updated successively using an iterative optimization procedure until the best clustering result is obtained.
The calculation process of the algorithm is as follows:
We randomly selected
k points as the initial center
Ci (
i = 1, 2, 3, …,
k). In this experiment, the
k values were 2 and 3. Then, we calculated the distance from any sample point
Pt (
t = 1, 2, 3, …,
n) to
Ci, where
n is the number of sample points.
The sample point was classified into the cluster with the centroid and the smallest distance. The mean was used to update the centroid value of each group.
Equations (2) to (4) were repeated until the centroid value of each cluster stopped changing.
The value of the contour coefficient is in the range of (−1, 1). The closer the samples of the same category are to each other, and the further the samples of different types are from each other, the higher the score and the better the clustering effect.
2.4. Data Analysis
Statistical analyses were performed using SPSS 22 (SPSS Inc., Chicago, IL, USA) to detect outliers and to perform the least significant difference (LSD) test to determine the statistically significant difference between two irrigation treatments at a significance level of p < 0.05. Figures were produced using Origin 2022 (OriginLab Inc., Hampden, MA, USA).
4. Discussion
In the context of climate change, extreme events are listed among the primary abiotic stresses that negatively affect crop growth and yield, and the plant response to this stress varies according to the length and size of the temperature rise and the crop stage at which the stress occurs [
34]. The heat stress problem has always been a complicated problem that needs to be solved in daylight conditions in greenhouses. Therefore, an appropriate cooling method is necessary for successful crop production in greenhouses. Evaporative cooling is the most effective cooling method among various cooling technologies [
10] that is applied with roof evaporative cooling, condensing fan cooling systems, micro-spray cooling systems, etc. It has been pointed out that the effects of different micro-spray water volumes and frequencies on crops under high-temperature stress are not the same [
14] and also depend on the effectiveness of different crops in responding to the micro-spray [
16]. This indicates that the appropriate water volume and frequency of micro-spray are also what we need to study at present. For this reason, the discussion is structured into three sub-sections. The first sub-section discusses the effects of MSDI on crop growth as well as fruit quality. The second sub-section discusses how MSDI on a daily scale affects the crop micro-climate as well as leaf temperature (
Tl) during multiple irrigations. The third sub-section discusses the effects of MSDI on short-term leaf gas exchange and chlorophyll fluorescence on a daily scale and investigates the effects of MSDI on cucumbers and tomatoes during the growth stage based on physiological conditions.
4.1. Effects of Micro-Spray on Crop Growth and Fruit Quality
Plant height is a function of the number of nodes and the length of each internode, both of which are strongly influenced by greenhouse temperatures [
35], and the overgrowth of plants at high temperatures occurs [
36]. Micro-spray changes the air temperature (
Ta) during the day, thus affecting internode spacing. Stem diameter has been considered the best predictor of plant growth and is usually discussed with water status or drought stress [
37,
38]. In this study, it was found that the plant height and stem thickness of cucumbers and tomatoes increased under MSDI in four planting seasons. Comparing plant height and stem thickness under the two irrigation treatments, there was a significant difference (
p < 0.05) in the plant height of cucumbers in 2018 and tomatoes in 2022, and in the stem thickness of cucumbers in 2017 and tomatoes in 2022. The stem diameter of cucumbers and tomatoes under MSDI was greater than it was under DI, probably due to the improved environment caused by the micro-spray, which reduced limitation due to heat stress.
Crop yield is most affected by fertilizer and irrigation. In this study, the same fertilizer was applied to cucumbers and tomatoes under both treatments and other field management was carried out in the same way. The effects of micro-spray on plant yield and quality have been extensively studied, and it was found that micro-spray increased grape and globe artichoke yields by 27.3% and 51.9%, respectively [
10,
15]. MSDI increased the yield of cucumbers and tomatoes by 18.9% and 40.4%, respectively, compared with DI in this study, which showed that MSDI could improve the growing environment of cucumbers and tomatoes. Although micro-spray increased water consumption, it improved crop yield under MSDI, which improved the economic efficiency of the crops. In general, crop yield can be increased by promoting leaf photosynthesis. In this study, weight per fruit (W
f) did not increase, but total yield increased in cucumbers (2017) and tomatoes (2021) under MSDI compared to DI, suggesting that MSDI mitigated heat stress, avoided flower scorching, and increased fruit number.
Total soluble solids (TSS) have been used by many researchers to estimate the quality of fruits [
39]. TSS is mostly made up of sugars and acids and has been linked to water stress [
40] (Zhang et al., 2017). It was found that there was a decreasing trend in TSS with increasing water use [
41], which can be used as an index to quickly evaluate the comprehensive quality of fruits [
30]. In this study, TSS was higher under DI than it was under MSDI, but there was no significant difference (
p > 0.05) between the two treatments over the four seasons. This suggests that the tomato plants did not really absorb the water from the micro-spray because it mostly evaporated.
4.2. Effects of Micro-Spray on Crop Micro-Climate and Leaf Temperature in Daily Scaled Blades
Suitable air temperature (
Ta) and relative humidity (
RH) are important prerequisites for ensuring the photosynthetic capacity of crops, and high temperatures interfere with vegetative development, thereby adversely affecting the growth stage of crops [
42]. The high temperature received by different crops varies with phenological stage, organ, and involved tissue. The degree of change in
Ta and
RH is not the same for different sprinkler durations or sprinkler frequencies in a single pass. Different amounts of water in a single pass are set for different crops with different maximum temperatures that they can tolerate so that the
Ta at which different crops are exposed reaches their acceptance level. Prolonged evaporative cooling in greenhouses, such as micro-spray, also increases air humidity [
43]. High humidity inhibits plant transpiration, making plants more susceptible to disease. So, we need to study how to set the right duration of sprinkler irrigation to achieve the best cooling effect while saving water. Changes in environmental factors outside the crops are the indirect effects of micro-spray on the crops. A plant’s response is a direct estimate of the cooling method, and leaf temperature (
Tl) is a better way to determine the status of a crop than air temperature because it is more directly related to the plant’s physiological response. Micro-spray cooling has been shown to have a positive effect on changes in
Tl [
15].
In this study, two sets of single micro-spray irrigation water volumes were set for different crops, and the changes in Ta and Tl for the single micro-spray duration of one minute and two minutes did not differ much. A single micro-spray duration of 1 min reduced Ta and Tl by 0.76 °C and 4.6 °C, respectively. A single micro-spray duration of 2 min reduced Ta and Tl by 0.8 °C and 4.9 °C, respectively. Both spray durations of the micro-spray were off for ten minutes to maximize the cooling effect. However, the difference in RH between one minute and two minutes of spray was only 2.7% and 4.3%, respectively, and the RH in the greenhouse after micro-spray did not exceed 60%, which proved that micro-spray showed an excellent ability to improve greenhouse heat stress and avoid high humidity.
4.3. Effects of Micro-Spray on Leaf Gas Exchange and ΦPSII
Crop physiological conditions such as the transpiration rate (
Tr), photosynthetic rate (
Pn), and stomatal conductance (
Gs) can directly reflect the real condition of crops, and micro-spray can increase the
Pn and
Gs of the crop [
14,
15]. However, the response of
Tr to micro-spray is not clear, and it has been pointed out that a change in the micro-climate leads to a reduction in water transpired by the leaves.
Gs prevents an increase in
Tl and a decrease in photosynthesis [
7]. There are studies that point out that micro-spray increases the
Tr of crops. In this study, a single micro-spray with different amounts of water increased the
Tr of cucumbers and tomatoes but did not show significant differences. MSDI increased the daily mean
Pn by 84.6% and 82.7% (2017 and 2018) and by 55.3% and 29.1% (2021 and 2022) in cucumber leaves (2017, 2018) and tomato leaves (2021, 2022), and increased the daily mean
Gs by 112.3% and 148.1% (2017 and 2018) and by 45.1% and 46.6% (2021 and 2022), respectively. The changes in the
Pn and
Gs for cucumber leaves with high air temperature showed a single peak trend of increasing and then decreasing under the two irrigation treatments, but the changes in the
Pn and
Gs of tomato leaves showed a double peak, with a small increase in
Pn and
Gs at 16:00 after experiencing a photosynthesis midday break. This proved that the spray irrigation periods of 9:00–12:00 for cucumbers and 9:00–15:00 for tomatoes are scientifically effective at high temperatures.
ΦPSII is widely used as a parameter to reflect the light use efficiency of plants and can reflect the degree of stress suffered by plants [
44,
45]. Some studies have shown that PAR is the most influential parameter on
ΦPSII. In this study, the values of
ΦPSII varied with PAR, which is similar to previous studies [
46]. The daily average
ΦPSII of cucumbers and tomatoes increased under MSDI. Before the micro-spray irrigation was turned on, the
ΦPSII of cucumbers and tomatoes under two irrigation treatments were different, and the
ΦPSII of MSDI was higher than that of DI. The variability in the
ΦPSII of cucumbers and tomatoes decreased with time under the two irrigation treatments, but the difference still existed at the end of one day of micro-spray irrigation. This demonstrated that the effect of MSDI on enhancing
ΦPSII was not transient when the spray irrigation was switched on. The
ΦPSII of cucumbers and tomatoes was also affected when the spray irrigation was not switched on.
The K-means clustering algorithm is considered to be one of the most influential and popular data mining algorithms in the research community. It has the remarkable feature of being fast and easy to implement and can be widely used in the field of data mining [
28,
29]. Changes in temperature and substrate water content have been explored using the K-means clustering method [
30]. In this study, we analyzed the relationship between the PAR and the change in
ΦPSII and opted to use the K-means clustering method for analysis. When K = 2, the classification level was clear (
Table 2), and there were significant differences in the PAR variables between the categories classified with the cluster analysis, proving that it is scientifically feasible to classify the effect of PAR on
ΦPSII into two parts. The PAR and
ΦPSII corresponding to the center coordinates of the clusters of cucumbers and tomatoes under MSDI were increased. It was demonstrated that MSDI increased the
ΦPSII of cucumbers and tomatoes and alleviated light stress. The first cluster of cucumbers and tomatoes had a smaller temperature range but the largest change in
ΦPSII, indicating that the
ΦPSII of cucumbers and tomatoes responded more to PAR. The second cluster of cucumbers and tomatoes floated less with the change in PAR, and the trend line was very flat (
Figure 9). This result suggests that PAR is not the main factor affecting the
ΦPSII of cucumbers and tomatoes in the second cluster.
5. Conclusions
In this study, two irrigation treatments (DI and MSDI) were designed for cucumbers (2017 and 2018) and tomatoes (2021 and 2022) grown in a greenhouse for four seasons. The experiments demonstrated the following:
MSDI increased the plant height and stem diameter of cucumbers and tomatoes in the four seasons, indicating a positive effect of the MSDI treatment on plant growth.
A single micro-spray duration of 2 min decreased Ta and Tl by 0.8 °C and 4.9 °C, respectively, and increased RH by 4.3%. A single micro-spray duration of 1 min decreased the average daily Ta and Tl by 0.76 °C and 4.6 °C, respectively, but the increase in RH was only 2.7%, which was much less than the single micro-spray duration of 2 min.
The Pn and Gs of cucumbers and tomatoes increased under MSDI, with the greatest enhancement at 10:00. At the same time, MSDI also improved the ΦPSII of cucumbers and tomatoes. The K-means analysis showed that MSDI could eliminate the negative effects of light stress on cucumbers and tomatoes.
MSDI increased the yield of cucumbers and tomatoes by 18.9% and 40.4%, respectively. MSDI mitigated heat stress, prevented flowers from being burned, and increased the number of fruits. The result showed that the TSS of cucumbers and tomatoes showed no significant difference (p > 0.05) between the two treatments in the four seasons.
In conclusion, MSDI can be an appropriate strategy for the irrigation needs and climate control of plants grown in greenhouses during the hot season. What needs to be investigated in the future is the adjustment of the micro-spray running times and the frequency of micro-sprays to cope with more severe environmental changes under different environmental factors.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}