Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage

 ,
,  ,
, 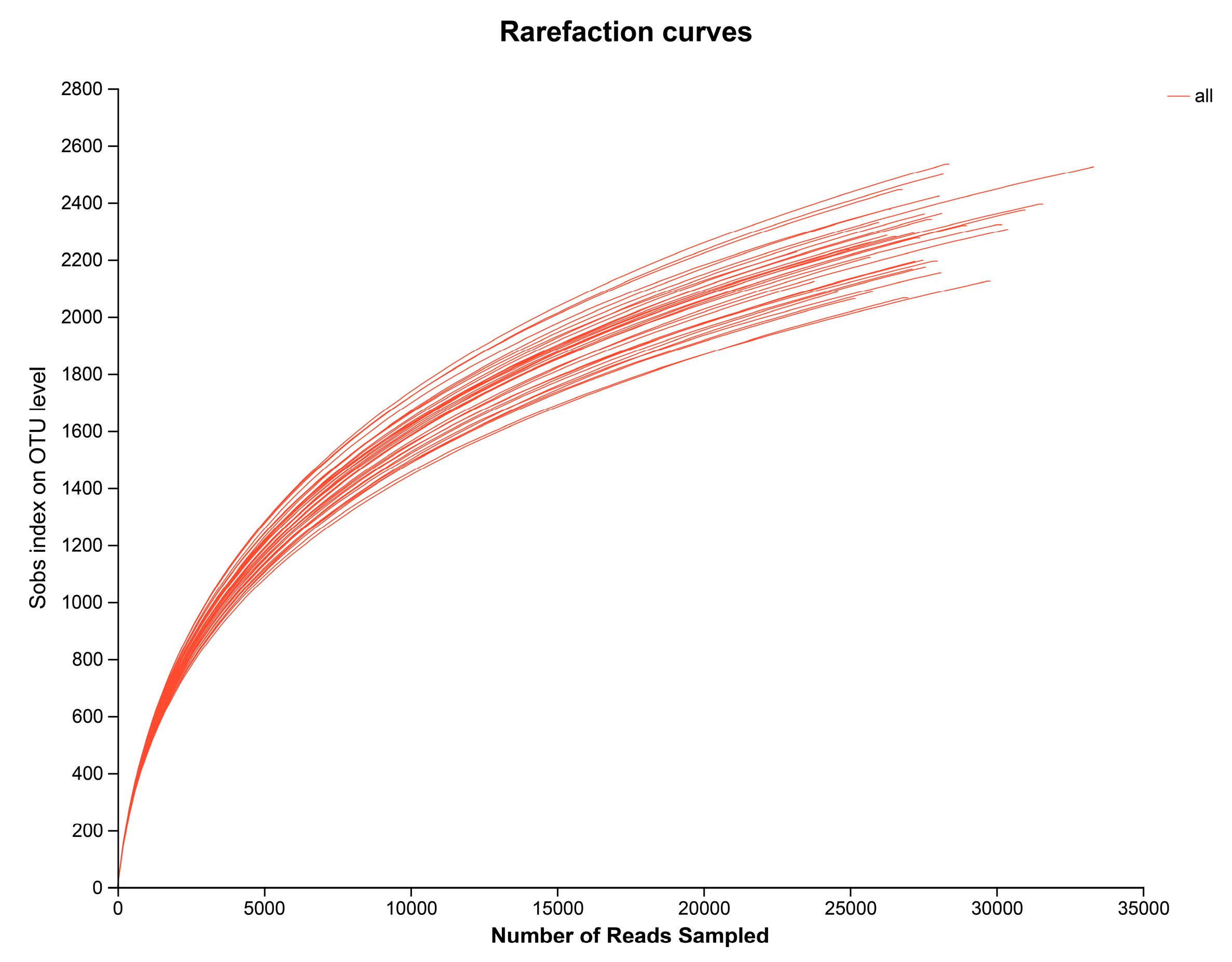{kind=link}
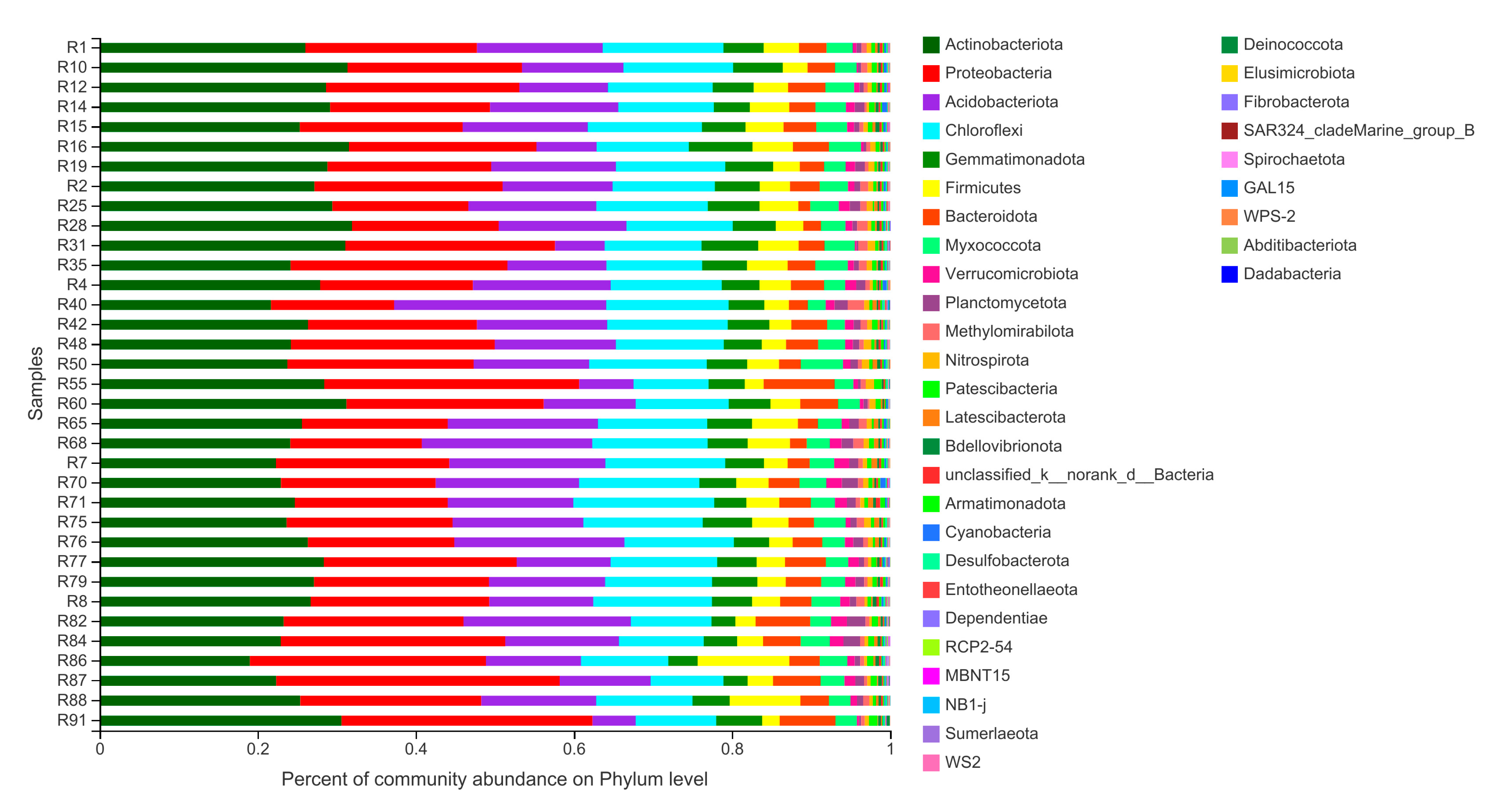{kind=link}
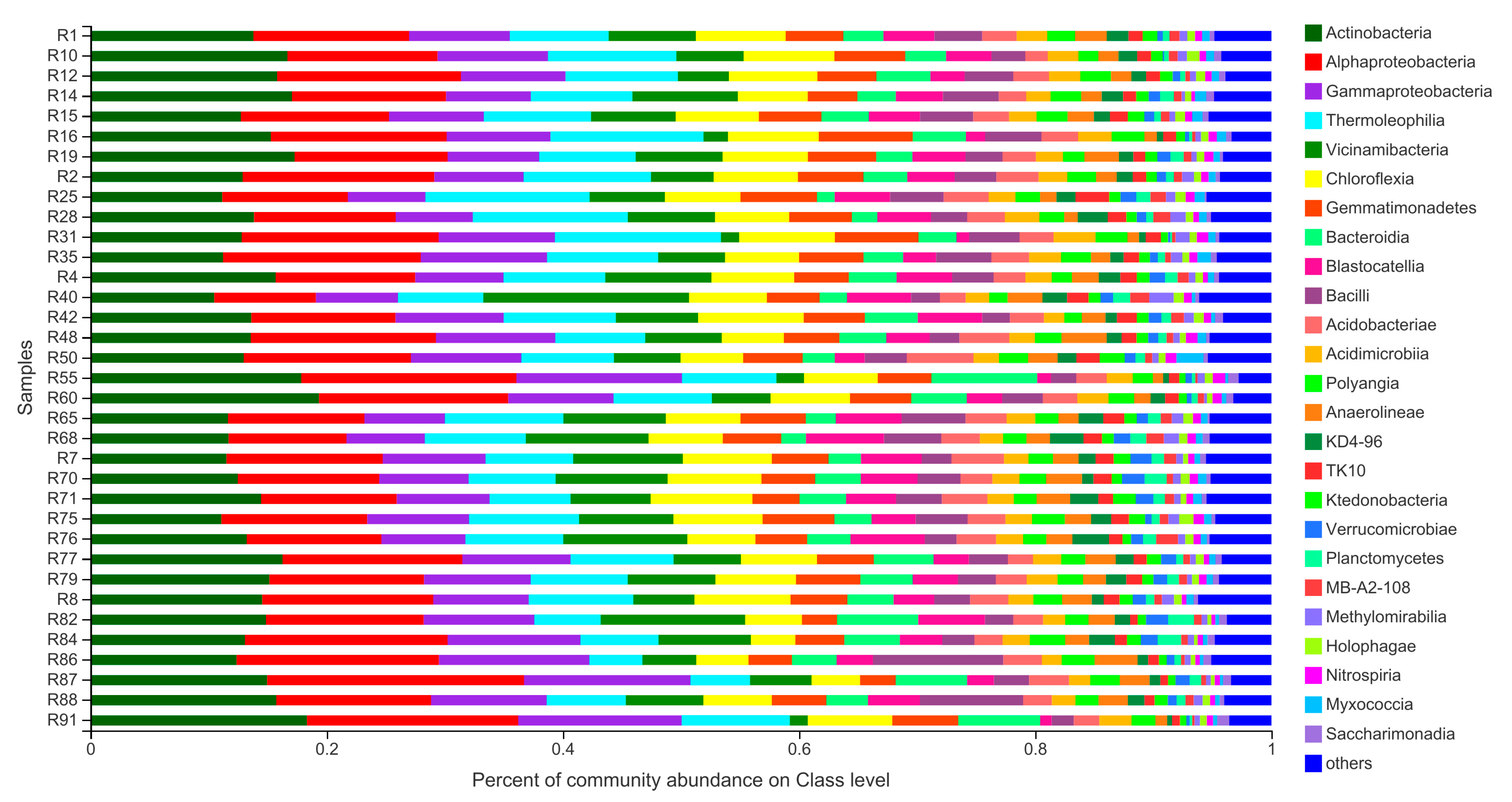{kind=link}
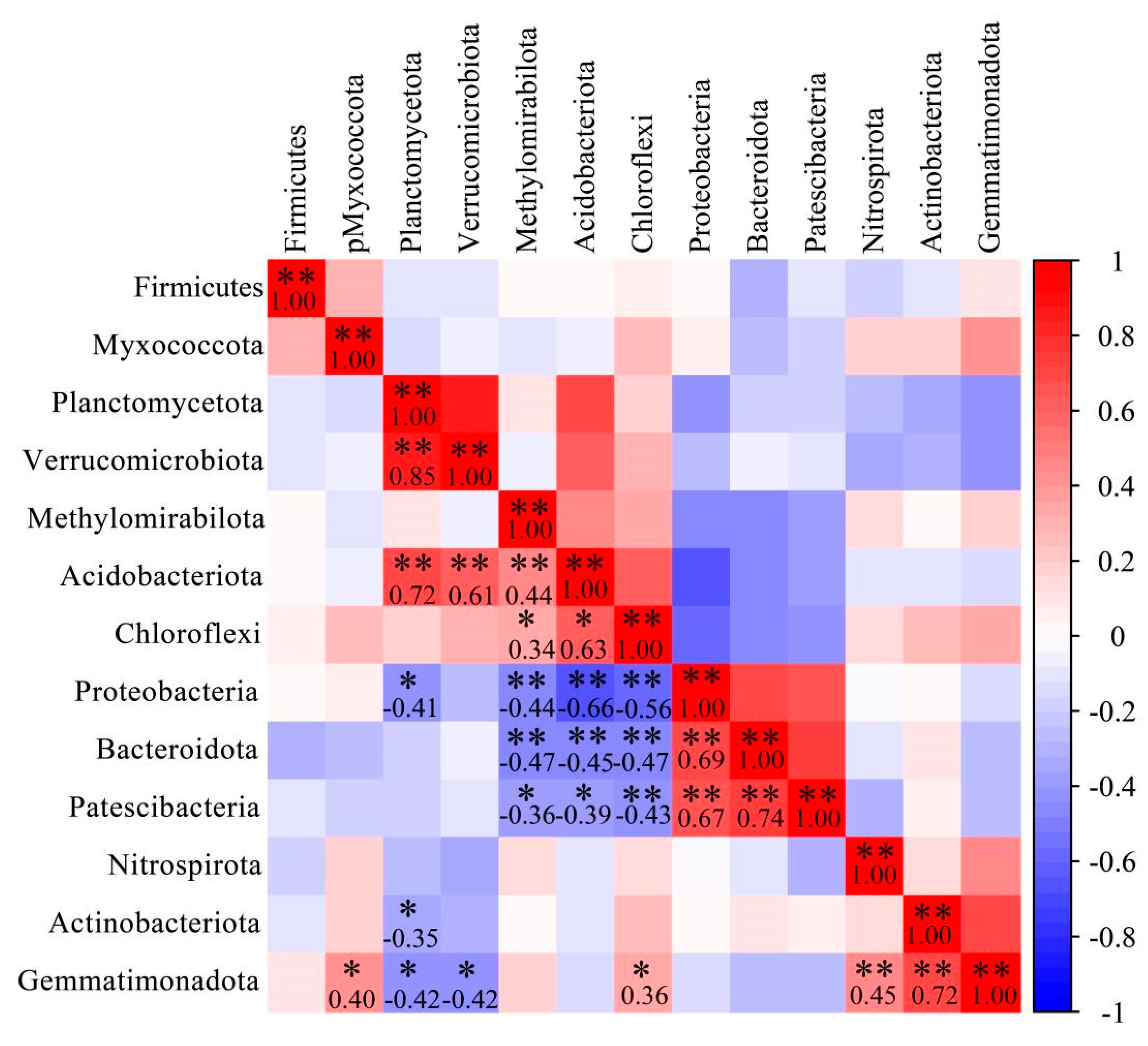{kind=link}
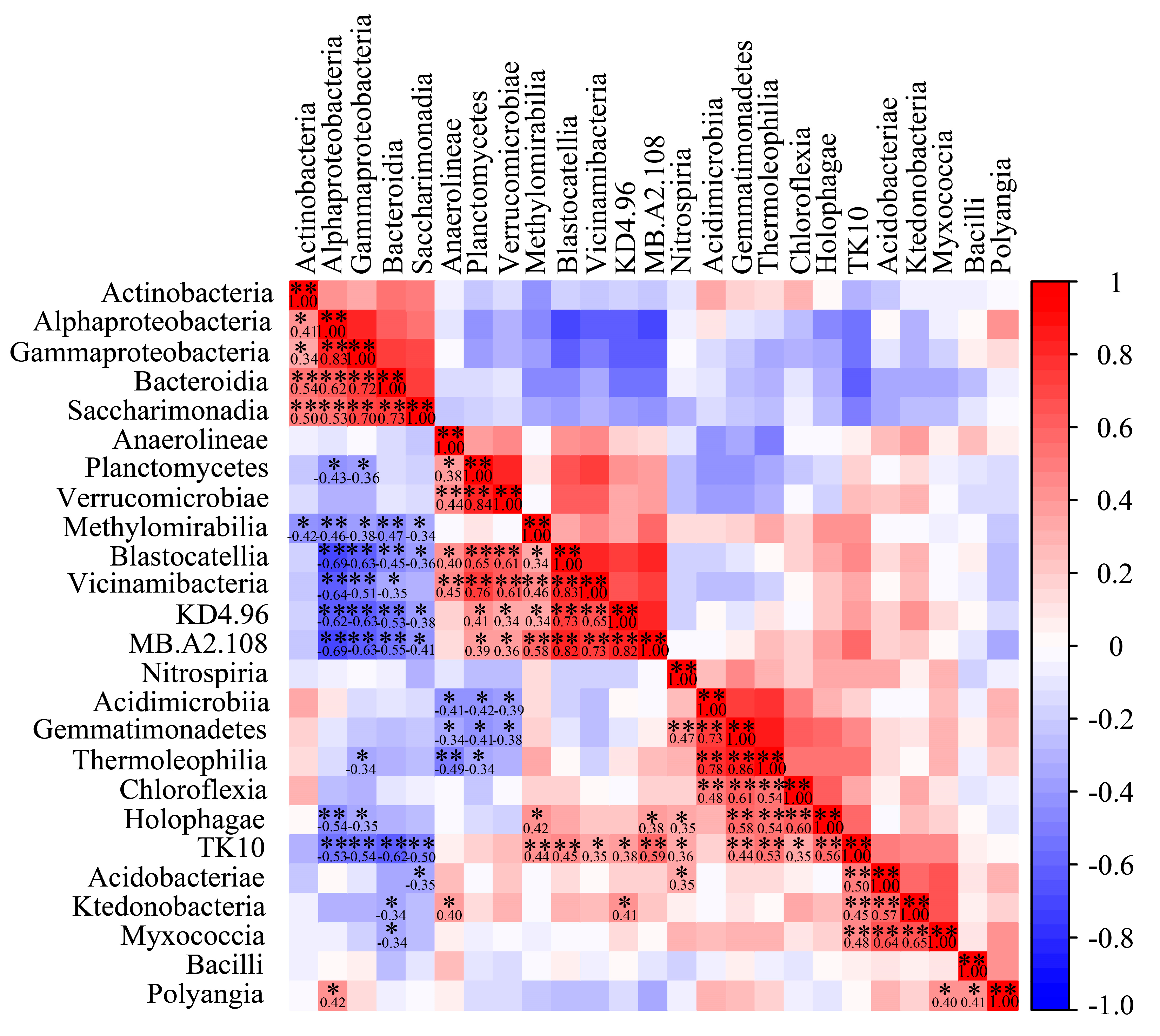{kind=link}
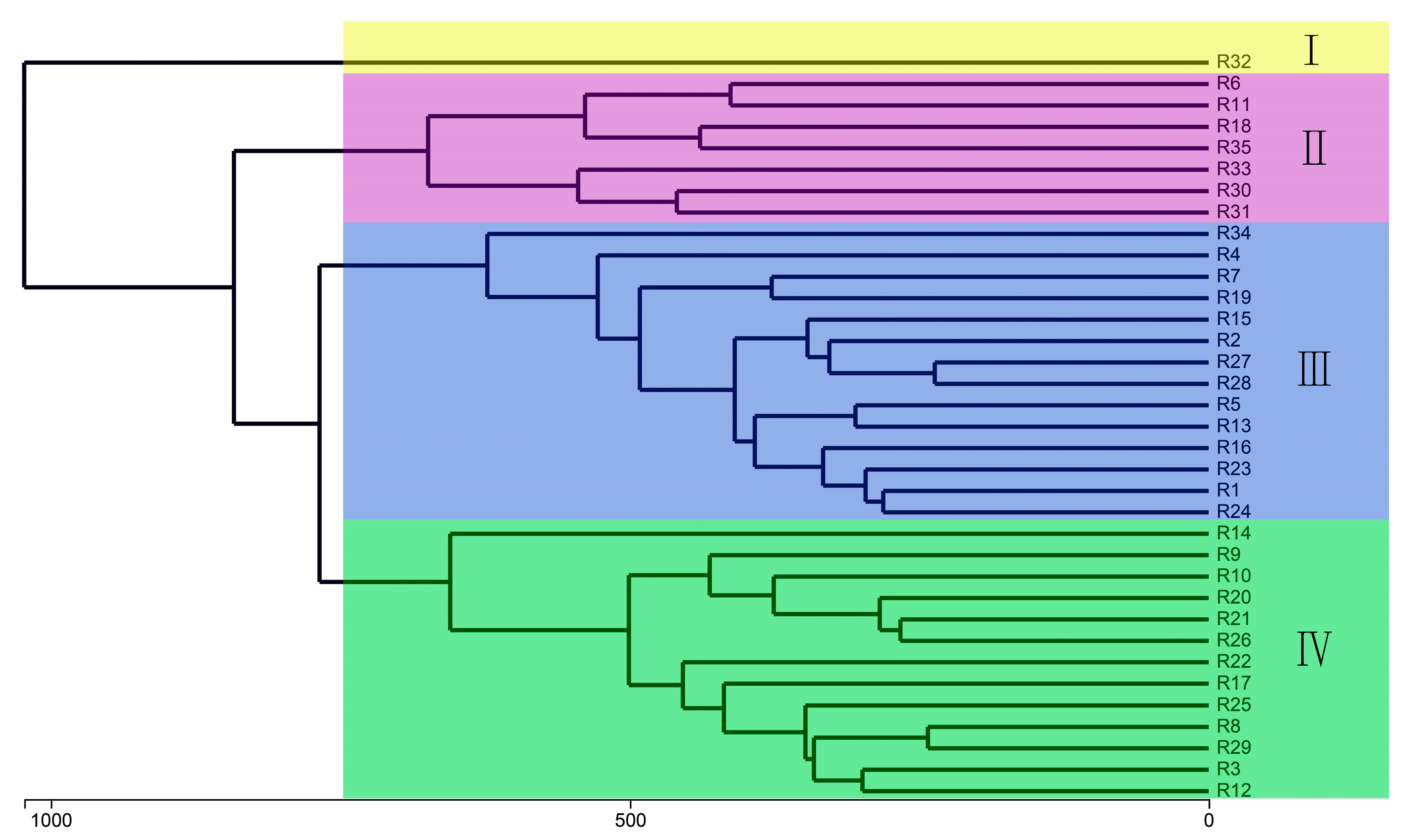{kind=link}
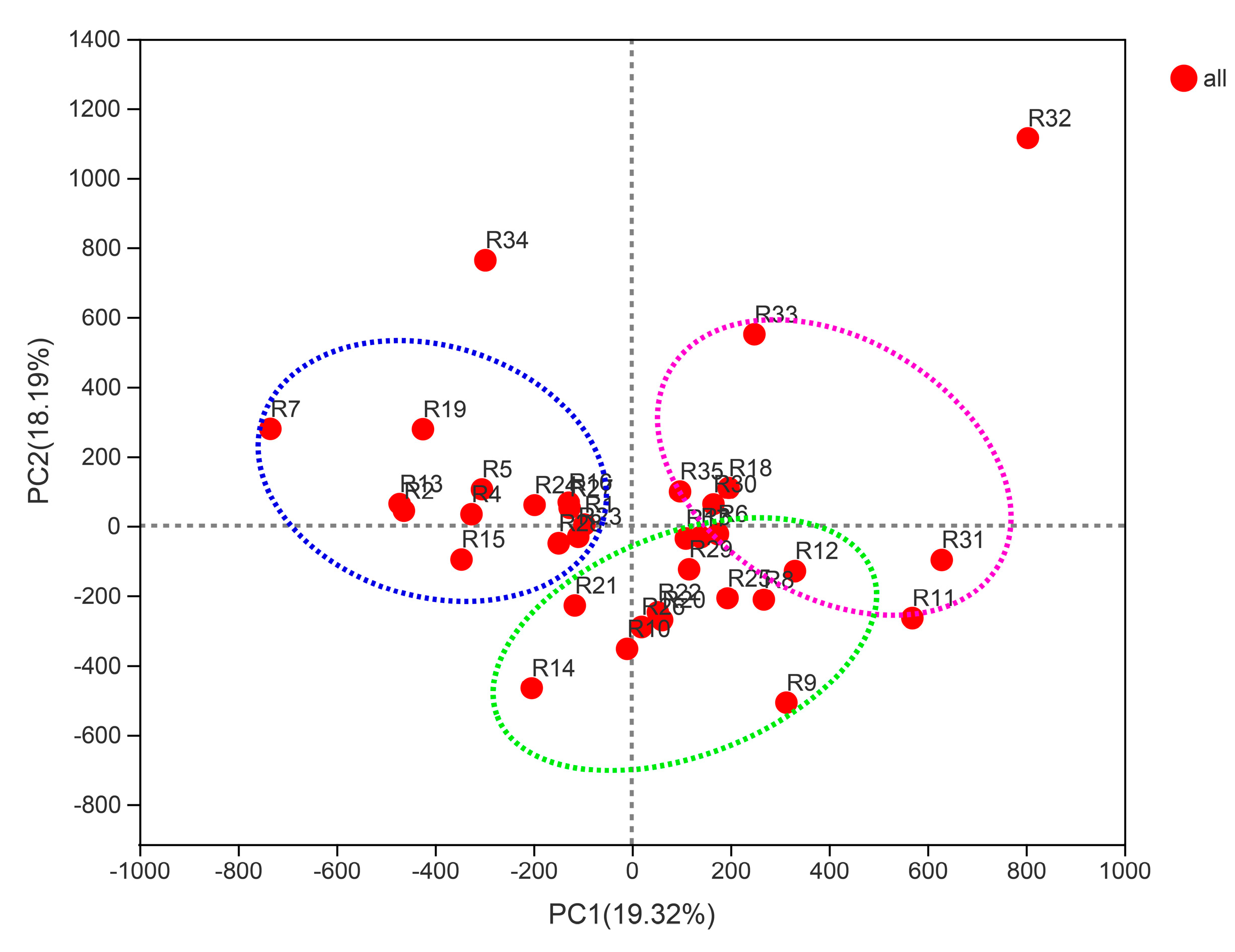{kind=link}
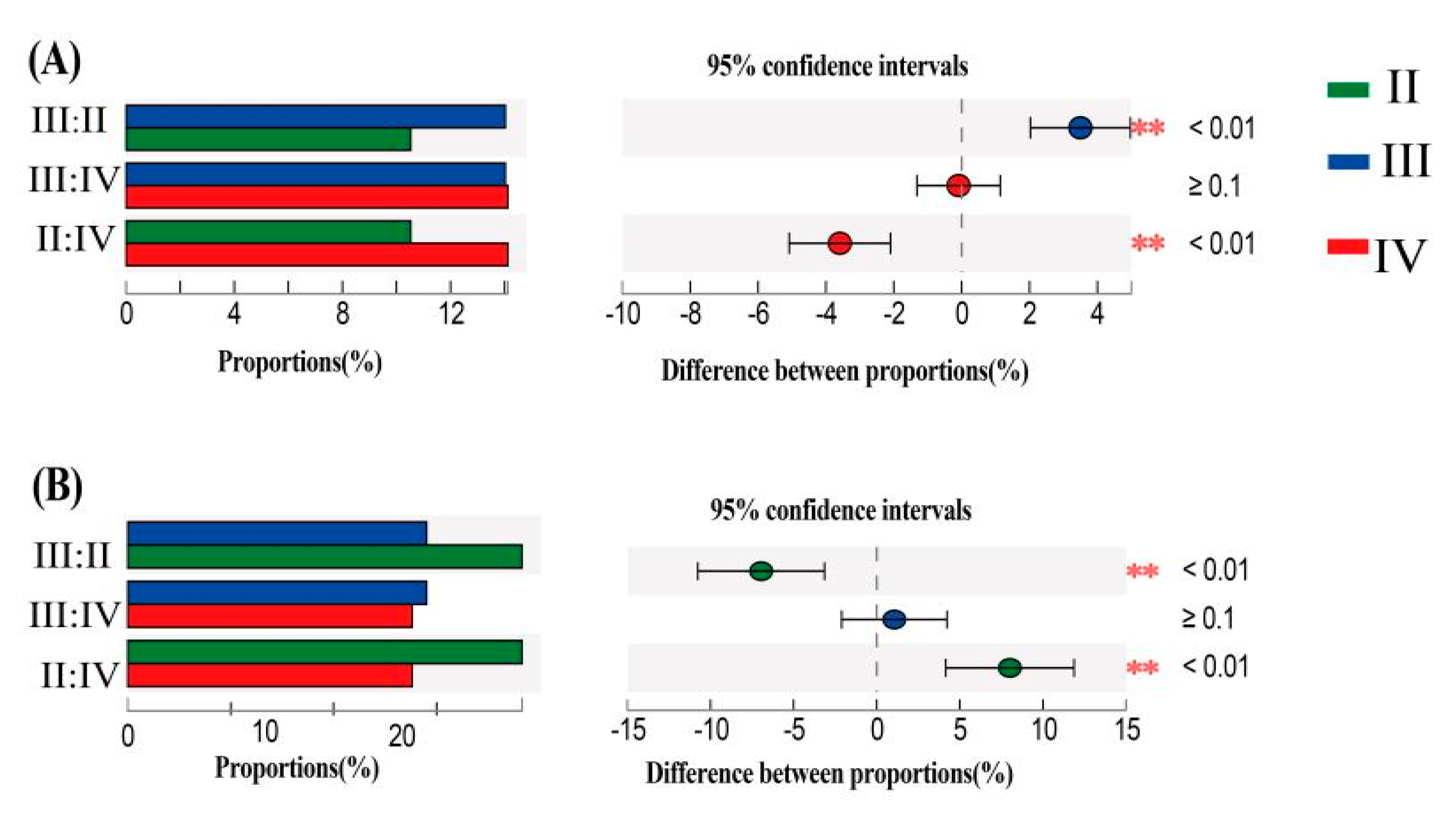{kind=link}
Abstract
Share and Cite
Zhang, S.; An, Y.; Zhou, Y.; Wang, X.; Tang, Y.; Zhang, D.; Sun, G.; Wang, Q.; Ren, X. Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage. Agronomy 2023, 13, 2825. https://doi.org/10.3390/agronomy13112825
Zhang S, An Y, Zhou Y, Wang X, Tang Y, Zhang D, Sun G, Wang Q, Ren X. Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage. Agronomy. 2023; 13(11):2825. https://doi.org/10.3390/agronomy13112825
Chicago/Turabian StyleZhang, Siyu, Yue An, Yu Zhou, Xiaofang Wang, Yiqing Tang, Daorong Zhang, Genlou Sun, Qifei Wang, and Xifeng Ren. 2023. "Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage" Agronomy 13, no. 11: 2825. https://doi.org/10.3390/agronomy13112825
APA StyleZhang, S., An, Y., Zhou, Y., Wang, X., Tang, Y., Zhang, D., Sun, G., Wang, Q., & Ren, X. (2023). Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage. Agronomy, 13(11), 2825. https://doi.org/10.3390/agronomy13112825