
Non-Thermal Plasma-Activated Water: A Cytogenotoxic Potential on Triticum aestivum

 ,
,  , , and
, , and
Abstract
:1. Introduction
- (a)
- the cytotoxic and genotoxic potential of PAW treatments on wheat sprouts in correlation with the number of reactive species (H2O2, NO3−) and the amount of pH in the plasma;
- (b)
- the germination rate and the length of the embryonic roots and the shoot of wheat sprouts after treatment with PAW;
- (c)
- the correlation between the genotoxic and cytotoxic potential of PAW and the germination rate, the length of the embryonic root and the length of the shoot of wheat sprouts treated with PAW.
2. Materials and Methods
2.1. Plasma Device and PAW Generation
2.2. Biological Material
2.3. Experiment Design
2.4. Cytogenetic Parameters
2.5. Measuring the Germination Rate
2.6. Measurement of the Length of theEmbryonic Root and the Shoot
2.7. Correlations between Cytogenetic Parameters and Biometric Parameters
2.8. Statistical Analysis
3. Results
3.1. Cytogenetic Parameters
3.1.1. Mitotic Index and Distribution of Mitotic Phases
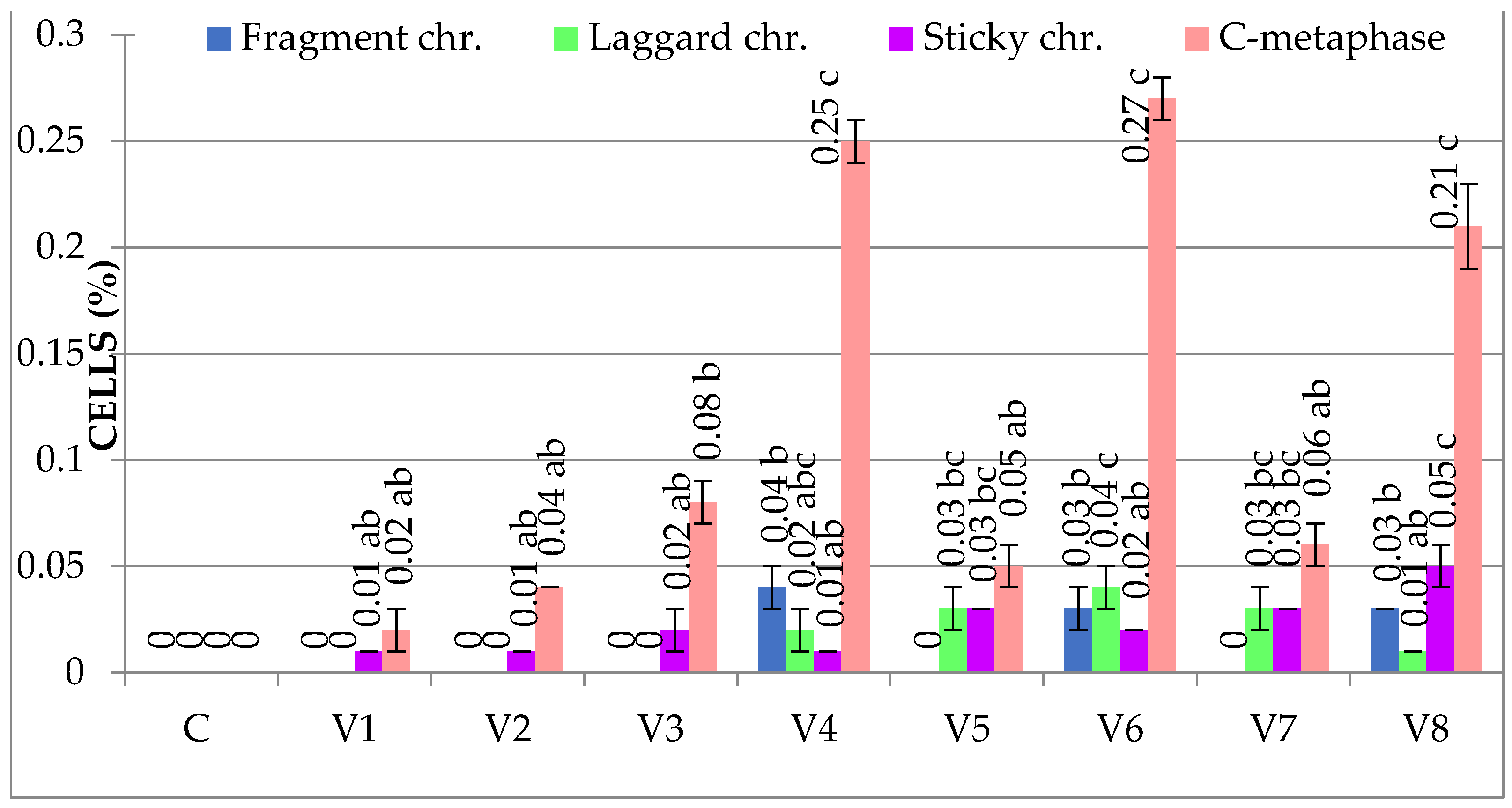
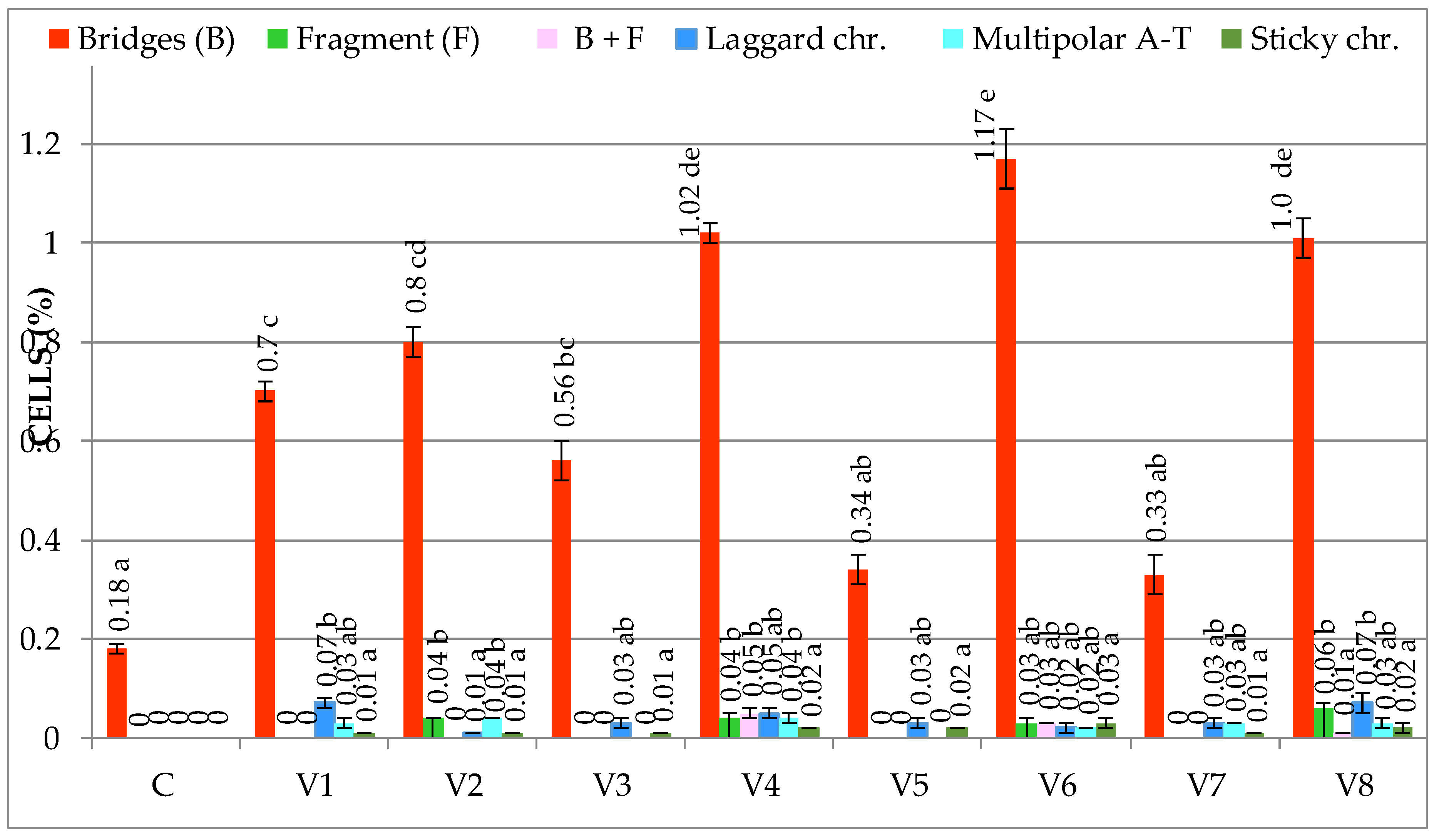
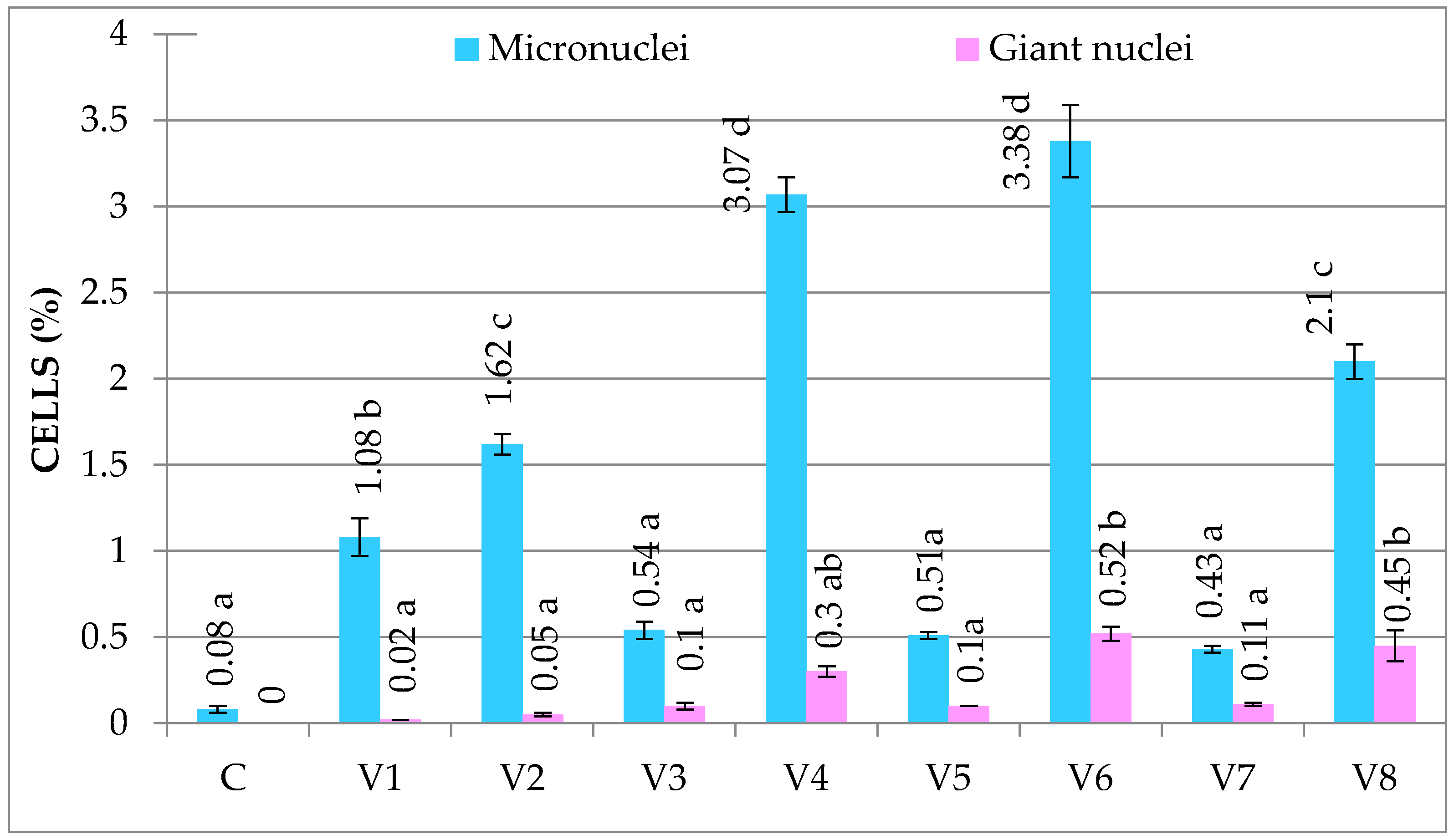
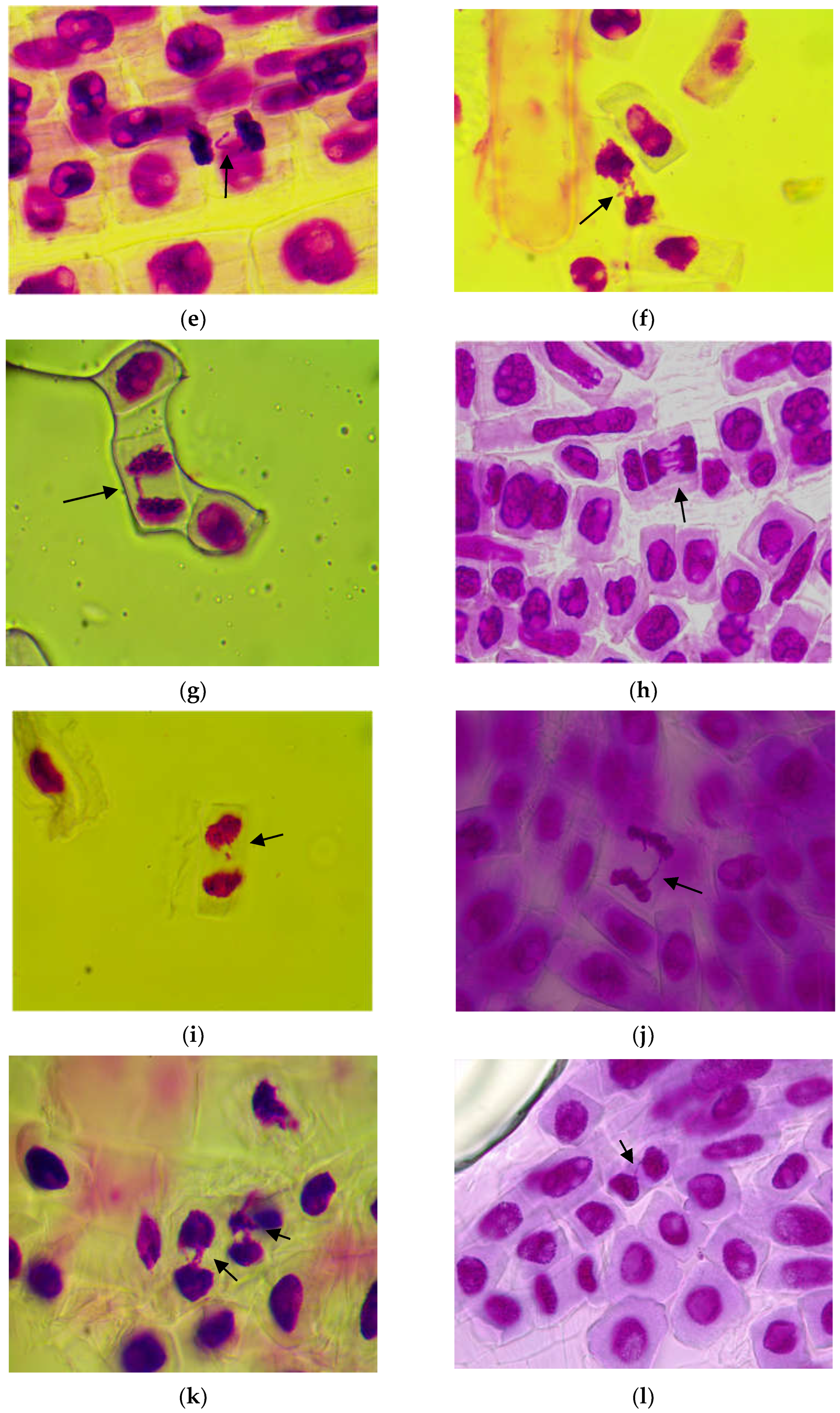
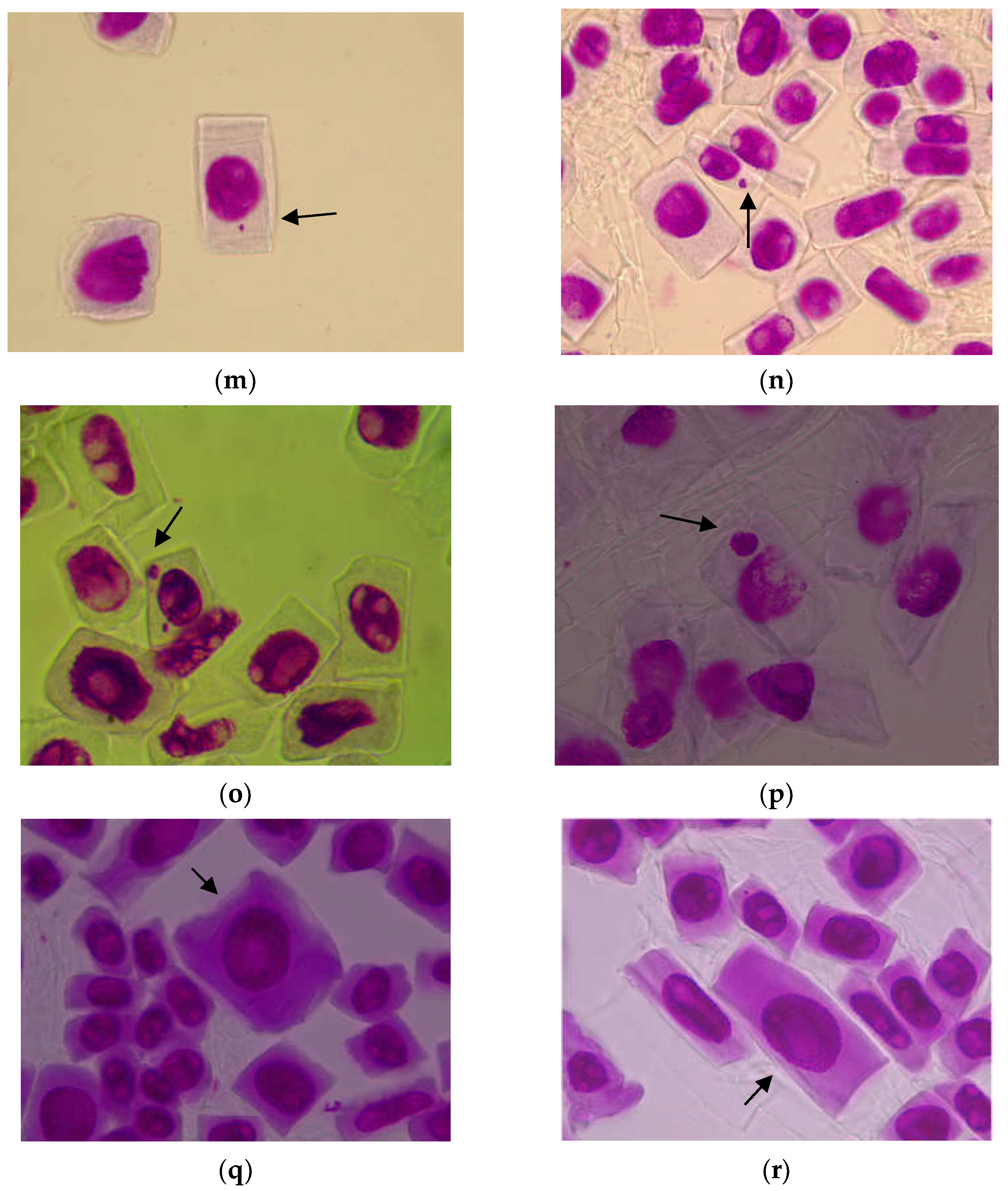
3.1.2. Genetic Abnormalities
3.2. Evaluation of the Germination Rate
3.3. Evaluation of the Length of the Embryonic Root and Shoot
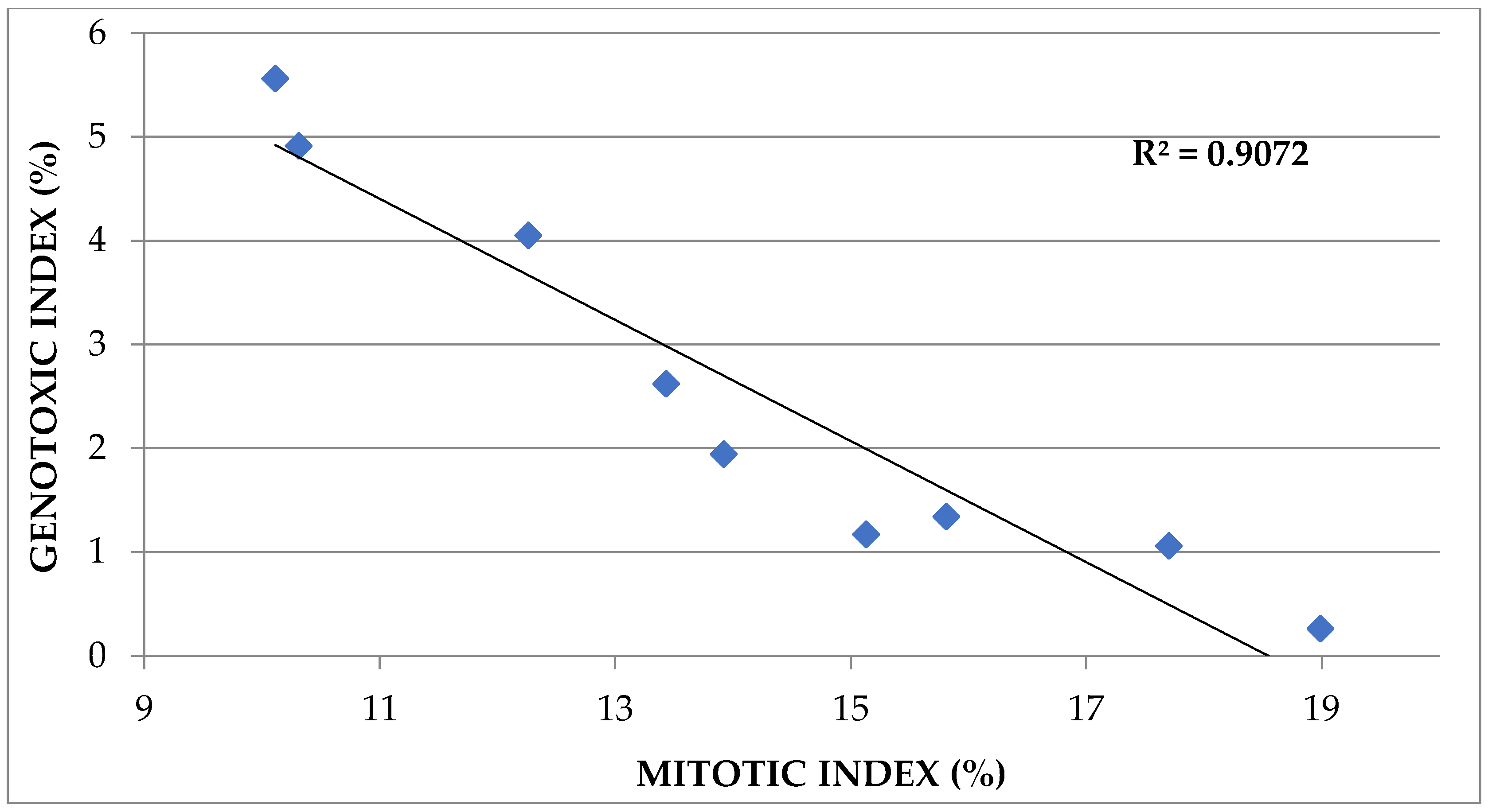
3.4. Correlations between Cytogenetic Parameters and Biometric Parameters
4. Discussion
4.1. Cytogenetic Investigations
4.2. Germination Rate and Length of the Embryonic Root and Shoot of Triticum aestivum
4.3. Correlations between Cytogenetic Parameters and Biometric Parameters
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mandici, A.; Cretu, D.E.; Burlica, R.; Astanei, D.; Beniuga, O.; Rosu, C.; Topa, D.C.; Aostacioaei, T.G.; Aprostosoaie, A.C.; Miron, A. Preliminary study on the impact of non-thermal plasma activated water on the quality in Triticum aestivum L. cv. Glosa sprouts. Horticulturae 2022, 8, 1158. [Google Scholar] [CrossRef]
- Herianto, S.; Shih, M.K.; Lin, C.M.; Hung, Y.C.; Hsieh, C.W.; Wu, J.S.; Chen, M.H.; Chen, H.L.; Hou, C.Y. The effects of glazing with plasma-activated water generated by a piezoelectric direct discharge plasma system on whiteleg shrimp (Litopenaeus vannamei). LWT 2022, 154, 112547. [Google Scholar]
- Naumova, I.K.; Maksimov, A.I.; Khlyustova, A.V. Stimulation of the germinability of seeds and germ growth under treatment with plasma-activated water. Surf. Eng. Appl. Electrochem. 2011, 47, 263–265. [Google Scholar]
- Khadre, A.; Yousef, A.E. Sporicidal action of ozone and hydrogen peroxide: A comparative study. Int. J. Food Microbiol. 2001, 71, 131–138. [Google Scholar]
- Fridman, G.; Brooks, A.D.; Balasubramanian, M.; Fridman, A.; Gutsol, A.; Vasilets, V.N.; Ayan, H.; Friedman, G. Comparison of direct and indirect effects of non-thermal atmospheric pressure plasma on bacteria. Plasma Process. Polym. 2007, 4, 370–375. [Google Scholar]
- Dobrynin, D.; Fridman, G.; Friedman, G.; Fridman, A. Physical and biological mechanisms of direct plasma interaction with living tissue. New J. Phys. 2009, 11, 115020. [Google Scholar]
- Ma, R.; Wang, G.; Tian, Y.; Wang, K.; Zhang, J.; Fang, J. Non-thermal plasma-activated water inactivation of food-borne pathogen on fresh produce. J. Hazard. Mater. 2015, 300, 643–651. [Google Scholar]
- Shen, J.; Tian, Y.; Li, Y.; Ma, R.; Zhang, Q.; Zhang, J.; Fang, J. Bactericidal effects against S. aureus and physicochemical properties of plasma activated water stored at different temperatures. Sci. Rep. 2016, 6, 28505. [Google Scholar]
- Traylor, M.J.; Pavlovich, M.J.; Karim, S.; Hait, P.; Sakiyama, Z.; Clark, D.S.; Graves, D.B. Long-term antibacterial efficacy of air plasma-activated water. J. Phys. D Appl. Phys. 2011, 44, 472001. [Google Scholar]
- Park, D.P.; Davis, K.; Gilani, S.; Alonzo, C.A.; Dobrynin, D.; Friedman, G.; Fridman, A.; Rabinovich, A.; Fridman, G. Reactive nitrogen species produced in water by non-equilibrium plasma increase plant growth rate and nutritional yield. Curr. Appl. Phys. 2013, 13, S19–S29. [Google Scholar]
- Leksakul, K.; Vichiansan, N.; Kaewkham, P.; Hattaphasu, B.; Boonyawan, D. Generating nitrate and nitrite on green oak lettuce in hydroponic farming by plasma system. Appl. Eng. Agric. 2021, 37, 105–112. [Google Scholar]
- Thirumdas, R.; Kothakota, A.; Annapure, U.S.; Siliveru, K.; Blundell, R.; Gatt, R.; Valdramidis, V.P. Plasma activated water (PAW): Chemistry, physico-chemical properties, applications in food and agriculture. Trends Food Sci. Technol. 2018, 77, 21–31. [Google Scholar]
- Oehmigen, K.; Hähnel, M.; Brandenburg, R.; Wilke, G.; Weltmann, K.-D.; Von Woedtke, T. The role of acidification for antimicrobial activity of atmospheric pressure plasma in liquids. Plasma Med. 2010, 7, 250–257. [Google Scholar]
- Leti, L.-I.; Gerber, I.C.; Mihaila, I.; Galan, P.-M.; Strajeru, S.; Petrescu, D.-E.; Cimpeanu, M.-M.; Topala, I.; Gorgan, D.-L. The modulatory effects of non-thermal plasma on seed’s morphology, germination and genetics–A Review. Plants 2022, 11, 2181. [Google Scholar] [PubMed]
- Zhang, K.; Zhang, G.; Liu, X.; Phan, A.N.; Luo, K. A study on CO2 decomposition to CO and O2 by the combination of catalysis and dielectric-barrier discharges at low temperatures and ambient pressure. Ind. Eng. Chem. Res. 2017, 56, 3204–3216. [Google Scholar]
- Liu, C.J.; Ye, J.; Jiang, J.; Pan, Y. Progresses in the preparation of coke resistant Ni-based catalyst for steam and CO2 reforming of methane. Chemcatchem 2015, 3, 529–541. [Google Scholar]
- Bolouki, N.; Kuan, W.H.; Huang, Y.Y.; Hsieh, J.H. Characterizations of a plasma-water system generated by repetitive microsecond pulsed discharge with air, nitrogen, oxygen, and argon gases species. Appl. Sci. 2021, 11, 6158. [Google Scholar]
- Graves, D.B. The emerging role of reactive oxygen and nitrogen species in redox biology and some implications for plasma applications to medicine and biology. J. Phys. D Appl. Phys. 2012, 45, 263001. [Google Scholar]
- Wang, H.; Guo, H.; Wu, Q.; Zhou, G.; Yi, C. Effect of activated carbon addition on H2O2 formation and dye decoloration in a pulsed discharge plasma system. Vacuum 2016, 128, 99–105. [Google Scholar]
- Guo, L.; Xu, R.; Gou, L.; Liu, Z.; Zhao, Y.; Liu, D.; Zhang, L.; Chen, H.; Kong, M.G. Mechanism of virus inactivation by cold atmospheric-pressure plasma and plasma-activated water. Appl. Environ. Microbiol. 2018, 84, e00726-18. [Google Scholar]
- Herianto, S.; Hou, C.Y.; Lin, C.M.; Chen, H.L. Nonthermal plasma-activated water: A comprehensive review of this new tool for enhanced food safety and quality. CRFSFS 2021, 20, 583–626. [Google Scholar]
- Puač, N.; Gherardi, M.; Shiratani, M. Plasma agriculture: A rapidly emerging field. Plasma Proces. Polym. 2018, 15, 1700174. [Google Scholar]
- Ranieri, P.; Sponsel, N.; Kizer, J.; Rojas-Pierce, M.; Hernández, R.; Gatiboni, L.; Grunden, A.; Stapelmann, K. Plasma agriculture: Review from the perspective of the plant and its ecosystem. Plasma Process. Polym. 2021, 18, 2000162. [Google Scholar]
- Waskow, A.; Howling, A.; Furno, I. Mechanisms of plasma-seed treatments as a potential seed processing technology. Front. Phys. 2021, 9, 617345. [Google Scholar]
- Imlay, J.; Chin, S.; Linn, S. Toxic DNA damage by hydrogen peroxide through the Fenton reaction in vivo and in vitro. Science 1988, 240, 640–642. [Google Scholar]
- Nakano, R.; Tashiro, K.; Aijima, R.; Hayashi, N. Effect of oxygen plasma irradiation on gene expression in plant seeds induced by active oxygen species. Plasma Med. 2016, 6, 303–313. [Google Scholar]
- Peťková, M.; Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, L.; Sevcovicova, A.; Gálová, E. The effects of cold atmospheric pressure plasma on germination parameters, enzyme activities and induction of DNA damage in barley. Int. J. Mol. Sci. 2021, 22, 2833. [Google Scholar] [PubMed]
- Burlicǎ, R.; Astanei, D.; Creţu, D.E.; Dîrlǎu, I.D.; Beniugǎ, O.; Padureanu, S.; Stoleru, V.; Patras, A. Non-thermal plasma t-shaped reactor for activated water production. EEMJ 2021, 20, 397–404. [Google Scholar]
- Sarinont, T.; Katayama, R.; Wada, Y.; Koga, K.; Shiratani, M. Plant growth enhancement of seeds immersed in plasma activated water. MRS Adv. 2017, 2, 995–1000. [Google Scholar]
- Shelar, A.; Singh, A.V.; Dietrich, P.; Maharjan, R.S.; Thissen, A.; Didwal, P.N.; Shinde, M.; Laux, P.; Luch, A.; Mathe, V.; et al. Emerging cold plasma treatment and machine learning prospects for seed priming: A step towards sustainable food production. RCS Adv. 2022, 12, 10467–10488. [Google Scholar]
- Constantin, M.J.; Owens, E.T. Introduction and perspectives plant genetic and cytogenetic assay. Mutat. Res. 1982, 99, 1–12. [Google Scholar]
- Fiskesjö, G. The Allium test as a standard in environmental monitoring. Hereditas 1985, 102, 99–112. [Google Scholar]
- Khanna, N.; Sharma, S. Allium cepa root chromosomal aberration assay. A review. Indian J. Pharm. Biol. Res. 2013, 1, 105–119. [Google Scholar]
- Sehgal, R.; Roy, S.; Kumar, V.L. Evaluation of cytotoxic potential of latex of Calotropis procera and podophyllotoxin in Allium cepa. Biocell 2006, 30, 9–13. [Google Scholar] [PubMed]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar]
- Marcano, L.; Carruyo, I.; Del Campo, A.; Montiel, X. Cytotoxicity and mode of action of maleic hydrazide in root tips of Allium cepa L. Environ. Res. 2004, 94, 221–226. [Google Scholar]
- Leme, D.M.; Marin-Morales, M.A. Allium cepa test in environmental monitoring: A review on its application. Mutat. Res. 2009, 682, 71–81. [Google Scholar]
- Mattar, M.Z.; Sammour, R.H.; Haroun, S.A.; Seada, M.L.L. Assessment of the genotoxicity of wastewater samples on Vicia faba L. Life Sci. 2014, 11, 991–998. [Google Scholar]
- Smaka-Kincl, V.; Stegnar, P.; Lovka, M.; Toman, M.J. The evaluation of waste, surface and ground water quality using the Allium test procedure. Mutat. Res. 1996, 368, 171–179. [Google Scholar]
- Da Silva Souza, T.; da Silva Figueira Barone, L.; Lacerda, D.; dos Santos Vergilio, C.; de Oliveira, B.C.V.; de Almeida, M.G.; Thompson, F.; de Rezende, C.E. Cytogenotoxicity of the water and sediment of the Paraopeba River immediately after the iron ore mining dam disaster (Brumadinho, Minas Gerais, Brazil). Sci. Total Environ. 2021, 775, 145193. [Google Scholar]
- Arjunan, K.P.; Sharma, V.K.; Ptasinska, S. Effects of atmospheric pressure plasmas on isolated and cellular DNA—A review. Int. J. Mol. Sci. 2015, 16, 2971–3016. [Google Scholar] [PubMed]
- Graves, D.B. Oxy-nitroso shielding burst model of cold atmospheric plasma therapeutics. Clin. Plasma Med. 2014, 2, 38–49. [Google Scholar]
- Naviaux, R.K. Oxidative shielding or oxidative stress? J. Pharmacol. Exp. Ther. 2012, 342, 608–618. [Google Scholar]
- Lu, X.; Naidis, G.V.; Laroussi, M.; Reuter, S.; Graves, D.B.; Ostrikov, K. Reactive species in non-equilibriu atmospheric-pressure plasmas: Generation, transport, and biological effects. Phys. Rep. 2016, 630, 1–84. [Google Scholar]
- Tero, R.; Yamashita, R.; Hashizume, H.; Suda, Y.; Takikawa, H.; Hori, M.; Ito, M. Nanopore formation process in artificial cell membrane induced by plasma-generated reactive oxygen species. Arch. Biochem. Biophys. 2016, 605, 26–33. [Google Scholar] [PubMed]
- Holubová, L.; Kyzek, S.; Ďurovcová, I.; Fabová, J.; Horváthová, E.; Ševčovičová, A.; Gálová, E. Non-Thermal Plasma—A New Green Priming Agent for Plants? Int. J. Mol. Sci. 2020, 21, 9466. [Google Scholar]
- Wende, K.; Landsberg, K.; Lindequist, U.; Weltmann, K.-D.; von Woedtke, T. Distinctive activity of nonthermal atmospheric-pressure plasma jet on eukaryotic and prokaryotic cells in a cocultivation approach of keratinocytes and microorganisms. IEEE Trans. Plasma Sci. 2010, 38, 2479–2485. [Google Scholar]
- Lunov, O.; Zablotskii, V.; Churpita, O.; Jäger, A.; Polívka, L.; Syková, E.; Dejneka, A.; Kubinová, Š. The interplay between biological and physical scenarios of bacterial death induced by non-thermal plasma. Biomaterials 2016, 82, 71–83. [Google Scholar] [PubMed]
- Morrison, K.A.; Akintayo, R.; Jin, J.; Kaymakcalan, O.; Weinreb, R.; Dong, X.; Westblade, L.F.; Golkowski, C.; Spector, J.A. Abstract: Non-thermal plasma treatment safely and rapidly eradicates MRSA from infected wounds. Plast. Reconstr. Surg.-Glob. Open 2016, 4, 52–53. [Google Scholar]
- Waskow, A.; Betschart, J.; Butscher, D.; Oberbossel, G.; Klöti, D.; Büttner-Mainik, A.; Adamcik, J.; Von Rohr, P.R.; Schuppler, M. Characterization of efficiency and mechanisms of cold atmospheric pressure plasma decontamination of seeds for sprout production. Front. Microbiol. 2018, 9, 3164. [Google Scholar]
- Yasuda, H.; Hashimoto, M.; Rhaman, M.M.; Takashima, K.; Mizuno, A. States of biological components in bacteria and bacteriophages during inactivation by atmospheric dielectric barrier discharges. Plasma Process. Polym. 2008, 5, 615–621. [Google Scholar]
- Mai-Prochnow, A.; Clauson, M.; Hong, J.; Murphy, A.B. Gram positive and gram negative bacteria differ in their sensitivity to cold plasma. Sci. Rep. 2016, 6, 38610. [Google Scholar] [PubMed]
- Ptasinska, S.; Bahnev, B.; Stypczynska, A.; Bowden, M.; Mason, N.J.; Braithwaite, N.S.J. DNA strand scission induced by a non-thermal atmospheric pressure plasma jet. Phys. Chem. Chem. Phys. 2010, 12, 7779–7781. [Google Scholar]
- Stypczyńska, A.; Ptasińska, S.; Bahnev, B.; Bowden, M.; Braithwaite, N.S.J.; Mason, N.J. The influence of amino acids on DNA damage induced by cold plasma radiation. Chem. Phys. Lett. 2010, 500, 313–317. [Google Scholar]
- Kyzek, S.; Holubová, L.; Medvecká, V.; Zahoranová, A.; Sevcovicová, A.; Gálová, E. Genotoxic effect of low temperature plasma treatment on plant seeds. Toxicol. Lett. 2017, 280, S119. [Google Scholar]
- Kyzek, S.; Holubová, Ľ.; Medvecká, V.; Tomeková, J.; Gálová, E.; Zahoranová, A. Cold atmospheric pressure plasma can induce adaptive response in pea seeds. Plasma Chem. Plasma Process. 2019, 39, 475–486. [Google Scholar]
- Crawford, D.R.; Davies, K.J. Adaptive response and oxidative stress. EHP 1994, 102, 25–28. [Google Scholar]
- Tomeková, J.; Kyzek, S.; Medvecká, V.; Gálová, E.; Zahoranová, A. Influence of cold atmospheric pressure plasma on pea seeds: DNA damage of seedlings and optical diagnostics of plasma. Plasma Chem. Plasma Process. 2020, 40, 1571–1584. [Google Scholar]
- Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, Ľ.; Gálová, E.; Zahoranová, A. Novel insight at the effect of cold atmospheric pressure plasma on the activity of enzymes essential for the germination of pea (Pisum sativum L. cv. Prophet) seeds. Plasma Chem. Plasma Process. 2020, 40, 1221–1240. [Google Scholar]
- Švubová, R.; Slováková, Ľ.; Holubová, Ľ.; Rovňanová, D.; Gálová, E.; Tomeková, J. Evaluation of the impact of cold atmospheric pressure plasma on soybean seed germination. Plants 2021, 10, 177. [Google Scholar]
- Yemeli, G.B.N.; Švubová, R.; Kostolani, D.; Kyzek, S.; Machala, Z. The effect of water activated by nonthermal air plasma on the growth of farm plants: Case of maize and barley. Plasma Process. Polym. 2021, 18, 2000205. [Google Scholar]
- Yi, H.; Meng, Z. Genotoxicity of hydrated sulfur dioxide on root tips of Allium sativum and Vicia faba. Mutat. Res. 2003, 537, 109–114. [Google Scholar] [PubMed]
- Luo, L.Z.; Werner, K.M.; Gollin, S.M.; Saunders, W.S. Cigarette smoke induces anaphase bridges and genomic imbalances normal cells. Mutat. Res. 2004, 554, 375–385. [Google Scholar]
- Türkoğlu, S. Genotoxicity of five food preservatives tested on root tips of Allium cepa L. Mutat. Res. 2007, 626, 4–14. [Google Scholar] [PubMed]
- Adams, T.; Kerisew, B. Assessment of cytotoxicity and genotoxicity potential of effluents from Bahir Dar tannery using Allium cepa. Adv. Public Health 2022, 2022, 1–10. [Google Scholar]
- Anirban, P.; Nag, S.; Sinha, K. Cytological effects of blitox on root mitosis of Allium cepa L. IJSRP 2013, 3, 1–7. [Google Scholar]
- Daphedar, A.; Taranath, T.C. Cytotoxic and genotoxic effect of biofabricated zinc nanoparticles on mitotic chromosomes of Drimia polyantha (Roxb.) Jessop. IJPCBS 2018, 8, 53–63. [Google Scholar]
- Gomes, J.V.; dos Santos Teixeira, J.T.; Moreira de Lima, V.; Borba, R.H. Induction of cytotoxic and genotoxic effects of Guandu river waters in the Allium cepa system. Ambient. Água 2015, 10, 48–58. [Google Scholar]
- Badar, A. Mitodepressive and chromotoxic activities of two herbicides in Allium cepa. Cytologia 1983, 48, 451–457. [Google Scholar]
- Türkoğlu, Ş. Genotoxic effects of mono-, di-, and trisodiumphosphate on mitotic activity, DNA content, and nuclear volume in Allium cepa L. Caryologia 2009, 62, 171–179. [Google Scholar]
- Fernandes, T.C.C.; Mazzeo, D.E.C.; Marin-Morales, M.A. Mechanism of micronuclei formation in polyploidizated cells of Allium cepa exposed to trifluralin herbicide. Pest. Biochem. Physiol. 2007, 88, 252–259. [Google Scholar]
- Kaushik, N.K.; Uhm, H.; Ha Choi, E.H. Micronucleus formation induced by dielectric barrier discharge plasma exposure in brain cancer cells. Appl. Phys. Lett. 2012, 100, 084102. [Google Scholar]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar]
- Shrivastav, M.; de Haro, L.P.; Nickolof, J.A. Regulation of DNA double-strand break repair pathway choice. Cell Res. 2008, 18, 134–147. [Google Scholar] [PubMed]
- Sivachandiran, L.; Khacef, A. Enhanced seed germination and plant growth by atmospheric pressure cold air plasma: Combined effect of seed and water treatment. RSC Adv. 2017, 7, 1822–1832. [Google Scholar]
- Arc, E.; Galland, M.; Godin, B.; Cueff, G.; Rajjou, L. Nitric oxide implication in the control of seed dormancy and germination. Front. Plant Sci. 2013, 4, 1–13. [Google Scholar]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar]
- Mildažiene, V.; Pauzaite, G.; Nauciené, Z.; Malakauskiene, A.; Zukiene, R.; Januskaitiene, I.; Jakstas, V.; Ivanauskas, L.; Filatova, I.; Lyushkevich, V. Pre-sowing seed treatment with cold plasma and electromagnetic field increases secondary metabolite content in purple coneflower (Echinacea purpurea) leaves. Plasma Process. Polym. 2018, 15, 1700059. [Google Scholar]
- Šera, B.; Šery, M. Non-thermal plasma treatment as a new biotechnology in relation to seeds, dry fruits, and grains. Plasma Sci. Technol. 2018, 20, 044012. [Google Scholar]
- Hayashi, N.; Ono, R.; Nakano, R.; Shiratani, M.; Tashiro, K.; Kuhara, S.; Yasuda, K.; Hagiwara, H. DNA microarray analysis of plant seeds irradiated by active oxygen species in oxygen plasma. Plasma Med. 2016, 6, 459–471. [Google Scholar]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar]
- Ghasempour, M.; Iranbakhsh, A.; Ebadi, M.; Oraghi Ardebili, Z. Seed priming with cold plasma improved seedling performance, secondary metabolism, and expression of deacetylvindoline O-acetyltransferase gene in Catharanthus roseus. Contrib. Plasma Phys. 2020, 60, e201900159. [Google Scholar]
- Noormohammadi, Z.; Mohammadzadeh-Shahir, M.; Fahmi, D.; Atyabi, S.M.; Farahani, F. Induced genetic and morphological changes in Catharanthus roseus L. by cold atmospheric plasma. Nova Biol. Reper. 2019, 6, 302–310. [Google Scholar]
- Stolárik, T.; Henselová, M.; Martinka, M.; Novák, O.; Zahoranová, A.; Cernák, M. Effect of low-temperature plasma on the structure of seeds, growth and metabolism of endogenous phytohormones in pea (Pisum sativum L.). Plasma Chem. Plasma Proc. 2015, 35, 659–676. [Google Scholar]
- Zhang, J.J.; Jo, J.O.; Huynh, D.L.; Mongre, R.K.; Ghosh, M.; Singh, A.K.; Lee, S.B.; Mok, I.S.; Hyuk, P.; Jeong, D.K. Growth-inducing effects of argon plasma on soybean sprouts via the regulation of demethylation levels of energy metabolism-related genes. Sci. Rep. 2017, 7, 41917. [Google Scholar] [PubMed] [Green Version]
- Jiang, J.; He, X.; Li, L.; Li, J.; Shao, H.; Xu, Q.; Ye, R.; Dong, Y. Effect of cold plasma treatment on seed germination and growth of wheat. Plasma Sci. Technol. 2014, 16, 54. [Google Scholar]
- Mildažienė, V.; Aleknavičiūtė, V.; Žūkienė, R.; Paužaitė, G.; Naučienė, Z.; Filatova, I.; Lyushkevich, V.; Haimi, P.; Tamošiūnė, I.; Baniulis, D. Treatment of common sunflower (Helianthus annus L.) seeds with radio-frequency electromagnetic field and cold plasma induces changes in seed phytohormone balance, seedling development and leaf protein expression. Sci. Rep. 2019, 9, 6437. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant of PAW | H2O2 (mg/L) ±5% | NO3− (mg/L) ±10% | Frequency (Hz) | Pulse (ms) | Q Air (L/min) | Q Water (mL/min) | pH |
|---|---|---|---|---|---|---|---|
| C | 0 | 0 | 0 | 0 | 0 | 0 | 7 |
| V1 | 7 | 30 | 150 | 2 | 1 | 10 | 4.6 |
| V2 | 8 | 40 | 250 | 2 | 1 | 10 | 4.4 |
| V3 | 3 | 14 | 60 | 2 | 1 | 20 | 5.1 |
| V4 | 13 | 49 | 250 | 2 | 1 | 3.33 | 4.1 |
| V5 | 5 | 15 | 60 | 1.5 | 1 | 25 | 5.1 |
| V6 | 14 | 55 | 250 | 1.5 | 1 | 3.33 | 3.9 |
| V7 | 1 | 8 | 250 | 2 | 2.5 | 85 | 5.5 |
| V8 | 22 | 68 | 250 | 2 | 2.5 | 4 | 3.8 |
| PAW Treatment Variant | Mean Number of Cells/Variant | Mitotic Index (%) | Cells in Prophase (%) | Cells in Metaphase (%) | Cells in Anaphase (%) | Cells in Telophase (%) |
|---|---|---|---|---|---|---|
| C | 7603.33 | 18.99 ± 3.00 e | 7.95 ± 1.00 e | 4.16 ± 0.78 c | 2.54 ± 1.00 bc | 4.34 ± 0.34 b |
| V1 | 7296.00 | 13.92 ± 1.00 bc | 6.21 ± 0.87 cd | 3.53 ± 1.01 c | 1.95 ± 0.69 ab | 2.23 ± 1.15 a |
| V2 | 7706.33 | 13.43 ± 1.94 bc | 7.34 ± 0.54 de | 2.22 ± 0.95 a | 1.16 ± 0.93 a | 2.71 ± 0.44 a |
| V3 | 7652.33 | 15.81 ± 0.61 cd | 6.83 ± 0.50 cde | 4.12 ± 0.13 c | 2.15 ± 0.02 ab | 2.71 ± 0.02 a |
| V4 | 7666.33 | 10.31 ± 0.89 a | 4.60 ± 0.30 b | 1.91 ± 0.35 a | 1.51 ± 0.44 ab | 2.29 ± 0.60 a |
| V5 | 7653.67 | 15.13 ± 1.17 c | 6.56 ± 0.40 cd | 3.40 ± 0.40 bc | 2.52 ± 0.52 bc | 2.65 ± 0.65 a |
| V6 | 7762.66 | 10.11 ± 0.59 a | 3.50 ± 0.50 a | 2.44 ± 0.04 ab | 1.82 ± 0.40 ab | 2.35 ± 0.35 a |
| V7 | 7747.66 | 17.70 ± 0.91 de | 6.12 ± 0.12 c | 4.28 ± 0.28 c | 3.37 ± 0.37 c | 3.93 ± 0.27 b |
| V8 | 7734.00 | 12.26 ± 0.06 ab | 5.84 ± 0.84 c | 2.33 ± 0.33 a | 1.70 ± 0.70 ab | 2.39 ± 0.25 a |
| PAW Treatment Variant | Cells in Metaphase (%) | Cells in A-T (%) | Cells in Interphase | GI (%) | |||
|---|---|---|---|---|---|---|---|
| Normal Metaphase (%) | Aberrant Metaphase (%) | Normal A-T (%) | Aberrant A-T (%) | Normal Interphase (%) | Aberrant Interphase (%) | ||
| C | 4.16 ± 0.78 b | 0.00 ± 0.00 a | 6.70 ± 1.33 c | 0.18 ± 0.03 a | 80.93 ± 3.01 a | 0.08 ± 0.02 a | 0.26 ± 0.03 a |
| V1 | 3.50 ± 1.00 b | 0.03 ± 0.00 ab | 3.37 ± 0.52 ab | 0.81 ± 0.09 c | 84.98 ± 1.06 c | 1.10 ± 0.11 bc | 1.94 ± 0.12 c |
| V2 | 2.17 ± 0.94 a | 0.05 ± 0.01 abc | 2.97 ± 0.22 ab | 0.90 ± 0.10 c | 84.90 ± 2.02 c | 1.67 ± 0.10 c | 2.62 ± 0.14 d |
| V3 | 4.02 ± 0.11 b | 0.1 ± 0.03 bc | 4.26 ± 0.22 ab | 0.60 ± 0.10 b | 83.55 ± 0.65 bc | 0.64 ± 0.09 ab | 1.34 ± 0.26 bc |
| V4 | 1.59 ± 0.33 a | 0.32 ± 0.05 d | 2.58 ± 0.54 a | 1.22 ± 0.02 d | 86.32 ± 1.09 c | 3.37 ± 0.86 e | 4.91 ± 0.87 f |
| V5 | 3.29 ± 0.42 b | 0.11 ± 0.04 c | 4.72 ± 1.22 b | 0.45 ± 0.05 b | 84.26 ± 1.23 bc | 0.61 ± 0.09 ab | 1.17 ± 0.05 b |
| V6 | 2.08 ± 0.10 a | 0.36 ± 0.06 d | 2.87 ± 0.09 a | 1.30 ± 0.05 d | 85.99 ± 0.38 c | 3.90 ± 0.36 e | 5.56 ± 0.41 g |
| V7 | 4.16 ± 0.25 b | 0.12 ± 0.07 c | 6.90 ±0.73 c | 0.40 ± 0.02 b | 81.76 ± 0.99 ab | 0.54 ± 0.08 ab | 1.06 ± 0.14 b |
| V8 | 2.03 ± 0.34 a | 0.30 ± 0.04 d | 2.89 ± 0.41 a | 1.20 ± 0.10 d | 85.19 ± 0.34 c | 2.55 ± 0.39 d | 4.05 ± 0.45 e |
| PAW Treatment Variant | Germination Rate of Grains (%) | |||
|---|---|---|---|---|
| After 48 h | After 72 h | After 96 h | After 120 h | |
| C | 78.00 ± 1.73 bc | 89.33 ± 2.08 c | 91.33 ± 5.03 b | 92.33 ± 2.08 b |
| V1 | 72.00 ± 7.00 ab | 82.00 ± 4.58 b | 90.00 ± 4.36 b | 92.66 ± 4.93 b |
| V2 | 75.66 ± 4.93 abc | 87.33 ± 1.53 bc | 91.33 ± 2.52 b | 91.66 ± 2.31 b |
| V3 | 79.00 ± 1.73 bc | 87.66 ± 4.04 bc | 89.00 ± 3.46 b | 92.00 ± 1.73 b |
| V4 | 68.00 ± 2.00 a | 70.00 ± 6.00 a | 71.00 ± 3.00 a | 79.00 ± 2.00 a |
| V5 | 71.66 ± 4.04 ab | 89.00 ± 3.46 bc | 89.00 ± 8.54 b | 93.33 ± 6.51b |
| V6 | 68.33 ± 5.03 a | 71.00 ± 2.00 a | 73.00 ± 3.00 a | 73.00 ± 3.00 a |
| V7 | 81.33 ± 5.13 c | 90.33 ± 5.77 c | 92.33 ± 6.81b | 92.33 ± 5.03 b |
| V8 | 69.66 ± 0.58 a | 71.33 ± 1.15 a | 72.33 ± 1.15 a | 74.00 ± 1.00 a |
| PAW Treatment Variant | Root Length (mm) | |||
|---|---|---|---|---|
| After 48 h | After 72 h | After 96 h | After 120 h | |
| C | 17.57 ± 2.94 bcd | 34.39 ± 2.11 b | 43.14 ± 3.19 bcd | 59.92 ± 5.65 bc |
| V1 | 15.08 ± 2.52 bc | 40.71 ± 2.82 b | 41.60 ± 1.91bc | 55.40 ± 1.17 bc |
| V2 | 19.10 ± 0.46 cd | 40.02 ± 2.93 b | 47.25 ± 5.49 cd | 49.62 ± 3.30 ab |
| V3 | 21.61 ± 1.09 d | 40.64 ± 2.13 b | 55.56 ± 4.29 d | 71.10 ± 5.79 d |
| V4 | 14.24 ± 2.54 b | 17.38 ± 2.26 a | 31.88 ± 4.24 ab | 49.29 ± 3.30 ab |
| V5 | 15.47 ± 0.21 bc | 37.80 ± 0.40 b | 51.33 ± 2.15 cd | 65.38 ± 3.83 cd |
| V6 | 1.58 ± 0.22 a | 15.52 ± 1.20 a | 25.21 ± 1.35 a | 39.15 ± 4.31a |
| V7 | 17.97 ± 1.91 bcd | 42.72 ± 1.83 b | 54.94 ± 7.17 d | 74.44 ± 4.26 d |
| V8 | 17.54 ± 2.29 bcd | 37.55 ± 1.85 b | 40.79 ± 2.23 bc | 52.56 ± 2.09 b |
| PAW Treatment Variant | Shoot Length (mm) | |||
|---|---|---|---|---|
| After 48 h | After 72 h | After 96 h | After 120 h | |
| C | 7.01 ± 1.03 bc | 18.76 ± 3.18 a | 29.09 ± 1.13 b | 43.5 ± 5.73 ab |
| V1 | 7.91 ± 0.05 c | 14.97 ± 1.75 a | 28.37 ± 2.12 b | 41.10 ± 4.33 ab |
| V2 | 7.45 ± 1.23 bc | 15.60 ± 0.73 a | 16.33 ± 0.99 a | 34.51 ± 0.31 ab |
| V3 | 7.25 ± 0.92 bc | 19.77 ± 1.76 a | 34.24 ± 1.59 b | 46.27 ± 1.37 ab |
| V4 | 6.91 ± 0.27 bc | 17.19 ± 1.32 a | 29.00 ± 2.88 b | 29.77 ± 3.91 a |
| V5 | 7.39 ± 0.45 bc | 15.7 ± 2.17 a | 29.04 ± 1.89 b | 44.92 ± 2.27 ab |
| V6 | 0.50 ± 0.02 a | 17.19 ± 1.32 a | 17.91 ± 0.21 a | 34.90 ± 2.26 ab |
| V7 | 7.42 ± 0.30 bc | 17.69 ± 1.60 a | 31.83 ± 3.87 b | 50.89 ± 3.63 b |
| V8 | 6.13 ± 0.98 b | 16.33 ± 0.99 a | 28.31 ± 1.52 b | 33.26 ± 1.35 ab |
| Correlated Parameters | Correlation Coefficient (R2) | |||
|---|---|---|---|---|
| After 48 h | After 72 h | After 96 h | After 120 h | |
| MI (%) correlated to Germination Rate (%) | 0.759 | 0.772 | 0.684 | 0.612 |
| GI (%) correlated to Germination Rate (%) | −0.647 | −0.877 | −0.829 | −0.829 |
| MI (%) correlated to Root Length (%) | 0.358 | 0.474 | 0.566 | 0.643 |
| GI (%) correlated to Root Length (%) | −0.445 | −0.609 | −0.687 | −0.668 |
| MI (%) correlated to Shoot Length (%) | 0.292 | 0.164 | 0.261 | 0.719 |
| GI (%) correlated to Shoot Length (%) | −0.457 | −0.054 | −0.259 | −0.706 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padureanu, S.; Burlica, R.; Stoleru, V.; Beniuga, O.; Dirlau, D.; Cretu, D.E.; Astanei, D.; Patras, A. Non-Thermal Plasma-Activated Water: A Cytogenotoxic Potential on Triticum aestivum. Agronomy 2023, 13, 459. https://doi.org/10.3390/agronomy13020459
Padureanu S, Burlica R, Stoleru V, Beniuga O, Dirlau D, Cretu DE, Astanei D, Patras A. Non-Thermal Plasma-Activated Water: A Cytogenotoxic Potential on Triticum aestivum. Agronomy. 2023; 13(2):459. https://doi.org/10.3390/agronomy13020459
Chicago/Turabian StylePadureanu, Silvica, Radu Burlica, Vasile Stoleru, Oana Beniuga, Delicia Dirlau, Daniel Eusebiu Cretu, Dragos Astanei, and Antoanela Patras. 2023. "Non-Thermal Plasma-Activated Water: A Cytogenotoxic Potential on Triticum aestivum" Agronomy 13, no. 2: 459. https://doi.org/10.3390/agronomy13020459
APA StylePadureanu, S., Burlica, R., Stoleru, V., Beniuga, O., Dirlau, D., Cretu, D. E., Astanei, D., & Patras, A. (2023). Non-Thermal Plasma-Activated Water: A Cytogenotoxic Potential on Triticum aestivum. Agronomy, 13(2), 459. https://doi.org/10.3390/agronomy13020459