
Manipulation of CBTS1 Expression Alters Tobacco Resistance to Spodoptera frugiperda and Phytophthora nicotianae

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
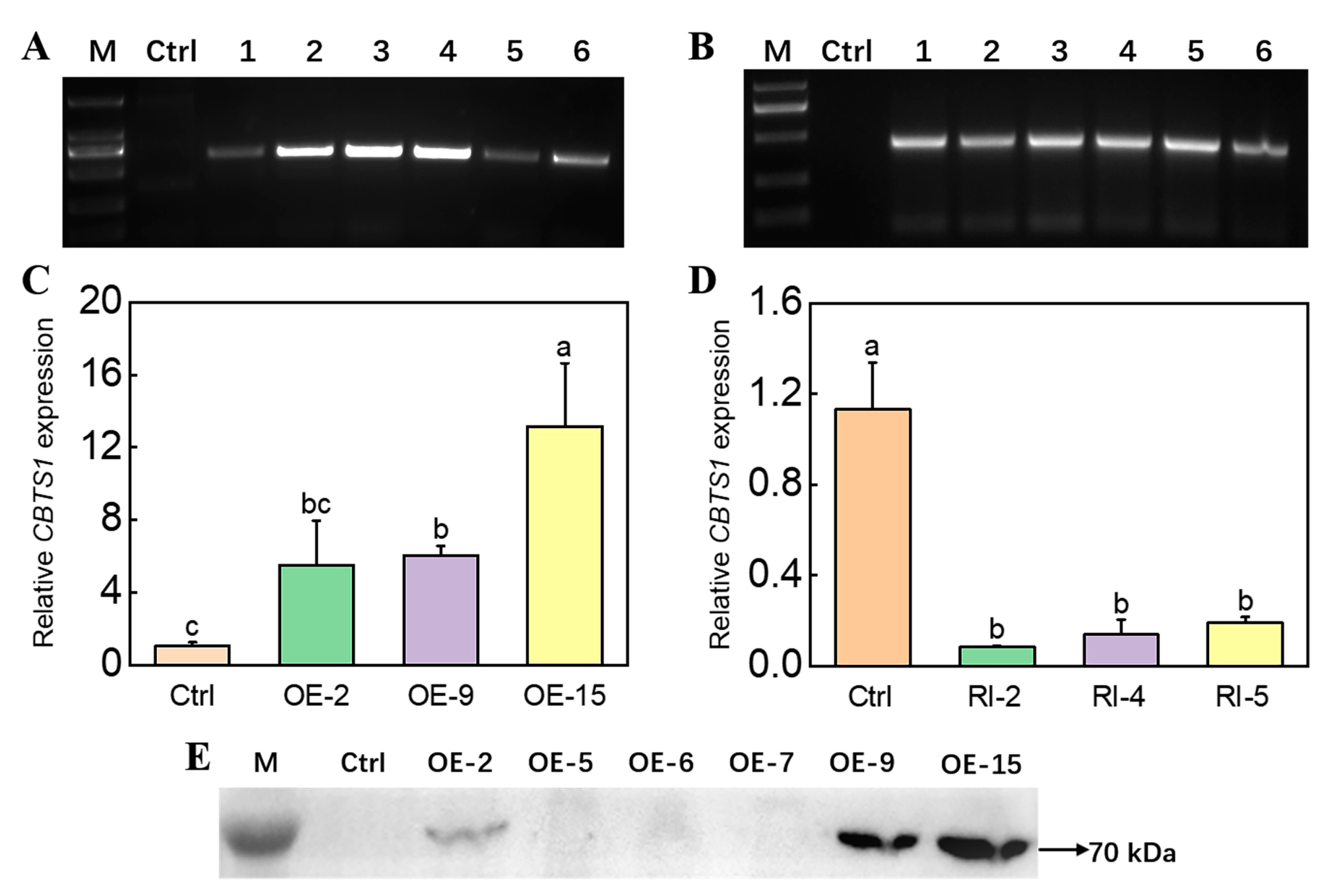
2.1. Development of CBTS1 Gene-Overexpressing and Gene-Silencing Plants
2.2. Changing the Expression of CBTS1 Altered the Cembranoid Content of Tobacco
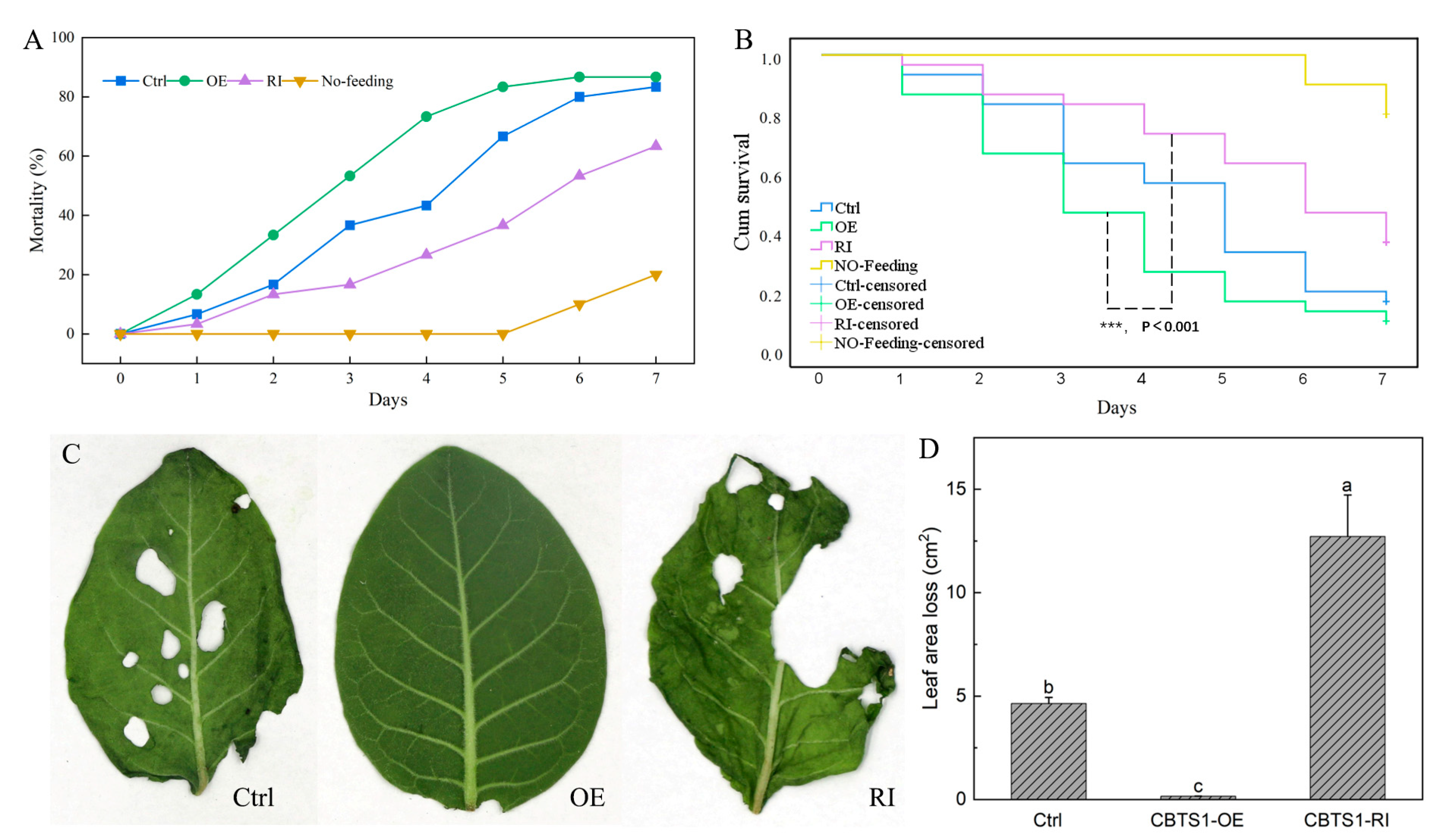
2.3. Manipulating the Expression of CBTS1 Altered Tobacco Resistance to Insect Pest S. frugiperda
2.4. Manipulating the Expression of CBTS1 Changed Tobacco Resistance to Oomycete Pathogen P. nicotianae
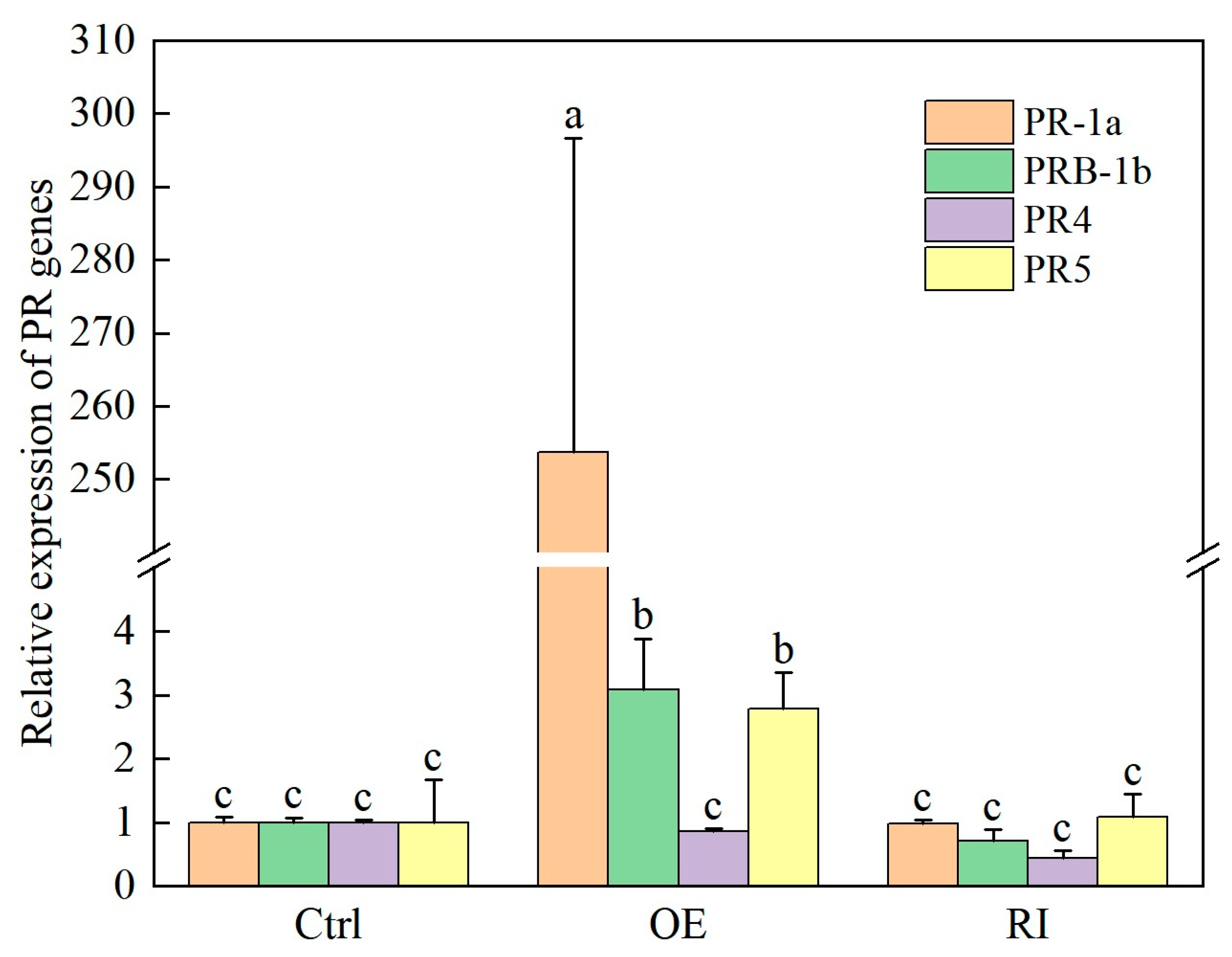
2.5. Expression of PR Genes were Changed in CBTS1-OE and CBTS1-RI Plants
3. Discussion
4. Materials and Methods
4.1. Growth Conditions for Tobacco Plants
4.2. Vectors Construction and Transgenic Plant Development
4.3. PCR and qRT-PCR
4.4. Western Blotting Analysis
4.5. Extraction and Determination of Cembranoid Diterpenes from Tobacco
4.6. Insect Toxicity and Feeding Preference Test
4.7. Oomycete Pathogen Challenge of Tobacco Leaves
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yan, N.; Liu, Y.H.; Gong, D.P.; Du, Y.M.; Zhang, H.B.; Zhang, Z.F. Solanesol: A review of its resources, derivatives, bioactivities, medicinal applications, and biosynthesis. Phytochem. Rev. 2015, 14, 403–417. [Google Scholar] [CrossRef]
- Ennajdaoui, H.; Vachon, G.; Giacalone, C.; Besse, I.; Sallaud, C.; Herzog, M.; Tissier, A. Trichome specific expression of the tobacco (Nicotiana sylvestris) cembratrien-ol synthase genes is controlled by both activating and repressing cis-regions. Plant Mol. Biol. 2010, 73, 673–685. [Google Scholar] [CrossRef]
- El Sayed, K.A.; Laphookhieo, S.; Baraka, H.N.; Yousaf, M.; Hebert, A.; Bagaley, D.; Rainey, F.A.; Muralidharan, A.; Thomas, S.; Shah, G.V. Biocatalytic and semisynthetic optimization of the anti-invasive tobacco (1S,2E,4R,6R,7E,11E)-2,7,11-cembratriene-4,6-diol. Bioorg. Med. Chem. 2008, 16, 2886–2893. [Google Scholar] [CrossRef]
- Sun, H.D.; Li, S.H. Diterpene Chemistry; Chemical Industry Press: Beijing, China, 2011; pp. 367–372. ISBN 9787122121394. [Google Scholar]
- Yan, N.; Du, Y.M.; Liu, X.M.; Zhang, H.B.; Liu, Y.H.; Zhang, P.; Gong, D.P.; Zhang, Z.F. Chemical structures, biosynthesis, bioactivities, biocatalysis and semisynthesis of tobacco cembranoids: An overview. Ind. Crop. Prod. 2016, 83, 66–80. [Google Scholar] [CrossRef]
- El Sayed, K.A.; Sylvester, P.W. Biocatalytic and semisynthetic studies of the anticancer tobacco cembranoids. Expert. Opin. Investig. Drugs 2007, 16, 877–887. [Google Scholar] [CrossRef]
- Kennedy, B.S.; Nielsen, M.T.; Severson, R.F.; Sisson, V.A.; Stephenson, M.K.; Jackson, D.M. Leaf surface chemicals from Nicotiana affecting germination of Peronospora tabacina (adam) sporangia. J. Chem. Ecol. 1992, 18, 1467–1479. [Google Scholar] [CrossRef]
- Aqil, F.; Zahin, M.; El Sayed, K.A.; Ahmad, I.; Orabi, K.Y.; Arif, J.M. Antimicrobial, antioxidant, and antimutagenic activities of selected marine natural products and tobacco cembranoids. Drug Chem. Toxicol. 2011, 34, 167–179. [Google Scholar] [CrossRef]
- Yuan, X.L.; Mao, X.X.; Du, Y.M.; Yan, P.Z.; Hou, X.D.; Zhang, Z.F. Anti-tumor activity of cembranoid-type diterpenes isolated from Nicotiana tabacum L. Biomolecules 2019, 9, 45. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Li, H.; Zhang, Q.; Yang, M.; Gu, Y.C.; Liang, L.F.; Tang, W.; Guo, Y.W. Rare Cembranoids from Chinese Soft Coral Sarcophyton ehrenbergi: Structural and Stereochemical Studies. J. Org. Chem. 2019, 84, 5091–5098. [Google Scholar] [CrossRef]
- Song, R.; Tan, Y.; Ahmed, W.; Zhou, G.; Zhao, Z. Unraveling the expression of differentially expressed proteins and enzymatic activity in response to Phytophthora nicotianae across different flue-cured tobacco cultivars. BMC Microbiol. 2022, 22, 112. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, S.T.; Yang, H.J.; Ji, H.; Wang, X.J. Gene expression profile analysis of tobacco leaf trichomes. BMC Plant Biol. 2011, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Tang, F.H.M.; Lenzen, M.; McBratney, A.; Maggi, F. Risk of pesticide pollution at the global scale. Nat. Geosci. 2021, 14, 206–210. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.; Liu, Q.; Tang, G.; Wang, H.; Fan, C.; Yin, S. Bioactive cembranoids from the South China Sea soft coral Sarcophyton elegans. Molecules 2015, 20, 13324–13335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.; Wagner, G.J. Elucidation of the functions of genes central to diterpene metabolism in tobacco trichomes using posttranscriptional gene silencing. Planta 2003, 216, 686–691. [Google Scholar] [CrossRef]
- Guo, Z.; Wagner, G.J. Biosynthesis of cembratrienols in cell-free extracts from trichomes of Nicotiana tabacum. Plant Sci. 1995, 110, 1–10. [Google Scholar] [CrossRef]
- Wang, E.; Wang, R.; De Parasis, J.; Loughrin, J.H.; Gan, S.; Wagner, G.J. Suppression of a P450 hydroxylase gene in plant trichome glands enhances natural-product-based aphid resistance. Nat. Biotechnol. 2001, 19, 371–374. [Google Scholar] [CrossRef]
- Biere, A.; Marak, H.B.; Damme, J.M. Plant chemical defense against herbivores and pathogens: Generalized defense or trade-offs. Oecologia 2004, 140, 430–441. [Google Scholar] [CrossRef]
- Makgoba, M.C.; Tshikhudo, P.P.; Nnzeru, L.R.; Makhado, R.A. Impact of fall armyworm (Spodoptera frugiperda) (J.E. Smith) on small-scale maize farmers and its control strategies in the Limpopo province, South Africa. Jamba 2021, 13, 1016. [Google Scholar] [CrossRef]
- Xu, P.J.; Zhang, D.D.; Wang, J.; Wu, K.M.; Wang, X.W.; Wang, X.F.; Ren, G.W. The host preference of Spodoptera frugiperda on maize and tobacco. Plant Prot. 2019, 45, 61–65. (In Chinese) [Google Scholar] [CrossRef]
- Paredes-Sánchez, F.A.; Rivera, G.; Bocanegra-García, V.; Martínez-Padrón, H.Y.; Berrones-Morales, M.; Niño-García, N.; Herrera-Mayorga, V. Advances in control strategies against Spodoptera frugiperda. A Review. Molecules 2021, 26, 5587. [Google Scholar] [CrossRef]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2021, 28, 602–610. [Google Scholar] [CrossRef]
- Zhang, X.G.; Sun, W.X.; Guo, L.; Yu, J.F.; Chang, C.J. Genetic and pathogenic variation among tobacco black shank strains of Phytophthora parasitica var. nicotianae from the main tobacco growing in China. J. Phytopathol. 2003, 151, 259–266. [Google Scholar] [CrossRef]
- Ji, P.; Csinos, A.S.; Hickman, L.L.; Hargett, U. Efficacy and application methods of oxathiapiprolin for management of black shank on tobacco. Plant Dis. 2014, 98, 1551–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonopoulos, D.F.; Melton, T.; Mila, A.L. Effects of chemical control, cultivar resistance, and structure of cultivar root system on black shank incidence of tobacco. Plant Dis. 2010, 94, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, J.K.; Wang, C.K.; Liu, X.F.; Fang, N.; Liu, Y.H.; Wang, W.J.; Yan, N.; Zhang, H.B.; Du, Y.M.; Liu, X.M.; et al. Formation of α- and β-Cembratriene-Diols in Tobacco (Nicotiana tabacum L.) Is Regulated by jasmonate-signaling components via manipulating multiple cembranoid synthetic genes. Molecules 2018, 23, 2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atici, O.; Nalbantoglu, B. Antifreeze proteins in higher plants. Phytochemistry 2003, 64, 1187–1196. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.M.; Liu, X.M.; Zhang, H.B.; Liu, Y.H.; Zhang, Z.F. A review on bioactivities of tobacco cembranoid diterpenes. Biomolecules 2019, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, S.; Yang, Y.; Jia, H.; Cui, H. Metabolic flux engineering of cembratrien-ol production in both the glandular trichome and leaf mesophyll in Nicotiana tabacum. Plant Cell. Physiol. 2018, 59, 566–574. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.H.; Hu, J.; Xu, Z.; Mu, C.; Alvarez, P.; Ford, B.D.; El Sayed, K.; Eterovic, V.A.; Ferchmin, P.A.; Hao, J. Neuroprotective activity of (1S,2E,4R,6R,-7E,11E)-2,7,11-cembratriene-4,6-diol (4R) in vitro and in vivo in rodent models of brain ischemia. Neuroscience 2015, 291, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Pham, N.B.; Butler, M.S.; Quinn, R.J. Naturally occurring cembranes from an Australian Sarcophyton species. J. Nat. Prod. 2002, 65, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Mischko, W.; Hirte, M.; Roehrer, S.; Engelhardt, H.; Mehlmer, N.; Minceva, M.; Brück, T. Modular biomanufacturing for a sustainable production of terpenoid-based insect deterrents. Green Chem. 2018, 20, 2637–2650. [Google Scholar] [CrossRef]
- Cui, L.; Bing, C.; Li, Y.; Wang, J.; Yang, D.; Yan, X.; Guo, Y.; Yuan, H. Research and application of chemical control technology against Spodoptera frugiperda (Lepidoptera: Noctuidae) in foreign countries. Plant Prot. 2019, 45, 7–13. (In Chinese) [Google Scholar] [CrossRef]
- Duan, S.Z.; Du, Y.M.; Hou, X.D.; Yan, N.; Dong, W.J.; Mao, X.X.; Zhang, Z.F. Chemical basis of the fungicidal fctivity of tobacco extracts against Valsa mali. Molecules 2016, 21, 1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sels, J.; Mathys, J.; De Coninck, B.M.; Cammue, B.P.; De Bolle, M.F. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar] [CrossRef]
- Zhang, H.B.; Bokowiec, M.T.; Rushton, P.J.; Han, S.C.; Timko, M.P. Tobacco transcription factors NtMYC2a and NtMYC2b form nuclear complexes with the NtJAZ1 repressor and regulate multiple jasmonate-inducible steps in nicotine biosynthesis. Mol. Plant. 2012, 5, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, M.D.; Grossniklaus, U. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef]
- Navarro-González, S.S.; Ramírez-Trujillo, J.A.; Peña-Chora, G.; Gaytán, P.; Roldán-Salgado, A.; Corzo, G.; Lina-García, L.P.; Hernández-Velázquez, V.M.; Suárez-Rodríguez, R. Enhanced tolerance against a fungal pathogen and insect resistance in transgenic tobacco plants overexpressing an endochitinase gene from Serratia marcescens. Int. J. Mol. Sci. 2019, 20, 3482. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, J.; Du, Z.; Tian, T.; Wang, W.; Ju, F.; Lin, X.; Zhang, Z.; Cao, Y.; Zhang, H. Manipulation of CBTS1 Expression Alters Tobacco Resistance to Spodoptera frugiperda and Phytophthora nicotianae. Agronomy 2023, 13, 845. https://doi.org/10.3390/agronomy13030845
Guan J, Du Z, Tian T, Wang W, Ju F, Lin X, Zhang Z, Cao Y, Zhang H. Manipulation of CBTS1 Expression Alters Tobacco Resistance to Spodoptera frugiperda and Phytophthora nicotianae. Agronomy. 2023; 13(3):845. https://doi.org/10.3390/agronomy13030845
Chicago/Turabian StyleGuan, Jian, Zaifeng Du, Tian Tian, Wenjing Wang, Fuzhu Ju, Xiaoyang Lin, Zhongfeng Zhang, Yi Cao, and Hongbo Zhang. 2023. "Manipulation of CBTS1 Expression Alters Tobacco Resistance to Spodoptera frugiperda and Phytophthora nicotianae" Agronomy 13, no. 3: 845. https://doi.org/10.3390/agronomy13030845
APA StyleGuan, J., Du, Z., Tian, T., Wang, W., Ju, F., Lin, X., Zhang, Z., Cao, Y., & Zhang, H. (2023). Manipulation of CBTS1 Expression Alters Tobacco Resistance to Spodoptera frugiperda and Phytophthora nicotianae. Agronomy, 13(3), 845. https://doi.org/10.3390/agronomy13030845