
Potassium Application Alleviated Negative Effects of Soil Waterlogging Stress on Photosynthesis and Dry Biomass in Cotton


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Leaf Relative Water Content
2.3. Leaf Gas Exchange Parameters
2.4. Agronomic Traits
2.5. Biomass
2.6. Statistical Analysis
3. Results
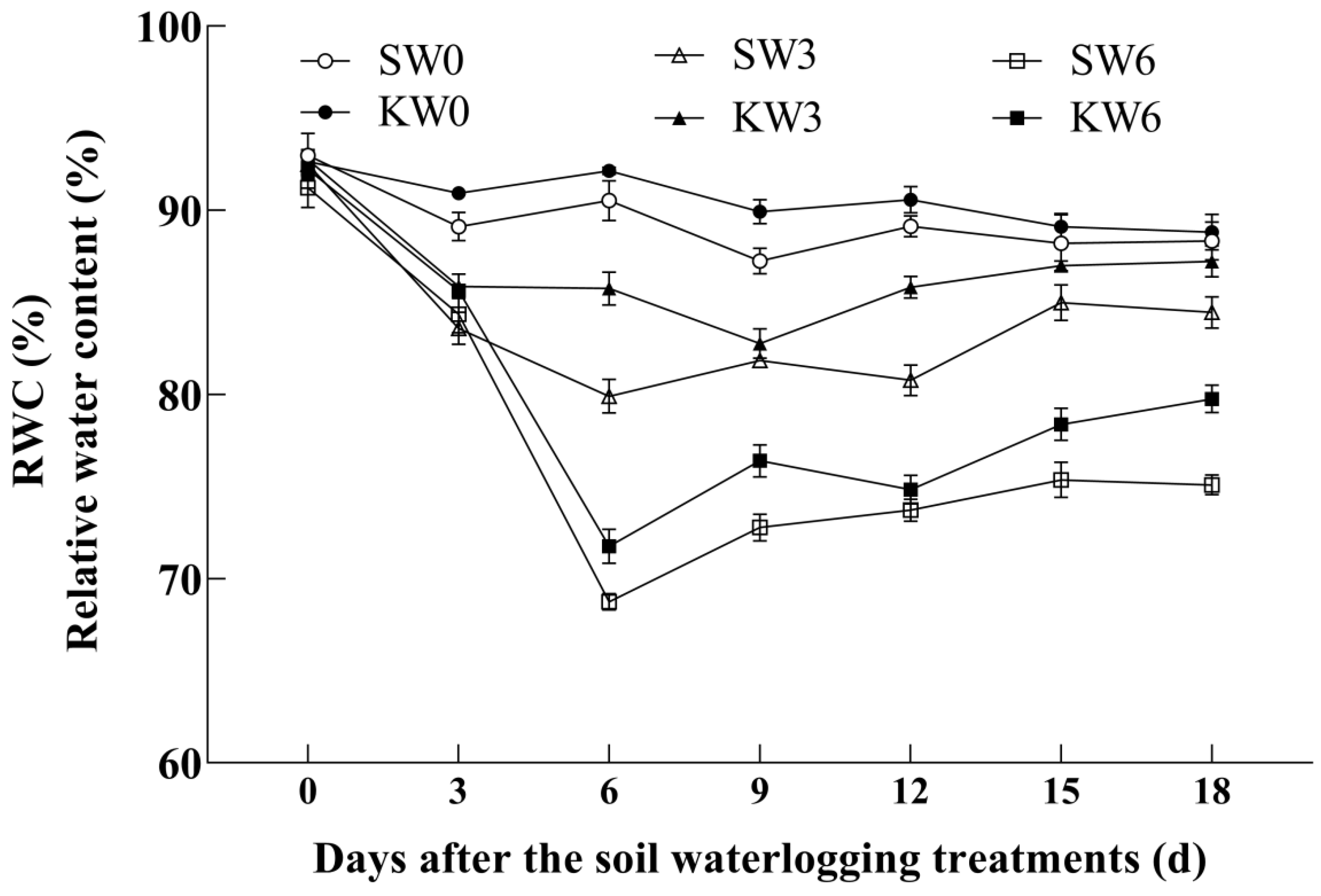
3.1. Leaf Water Status Traits
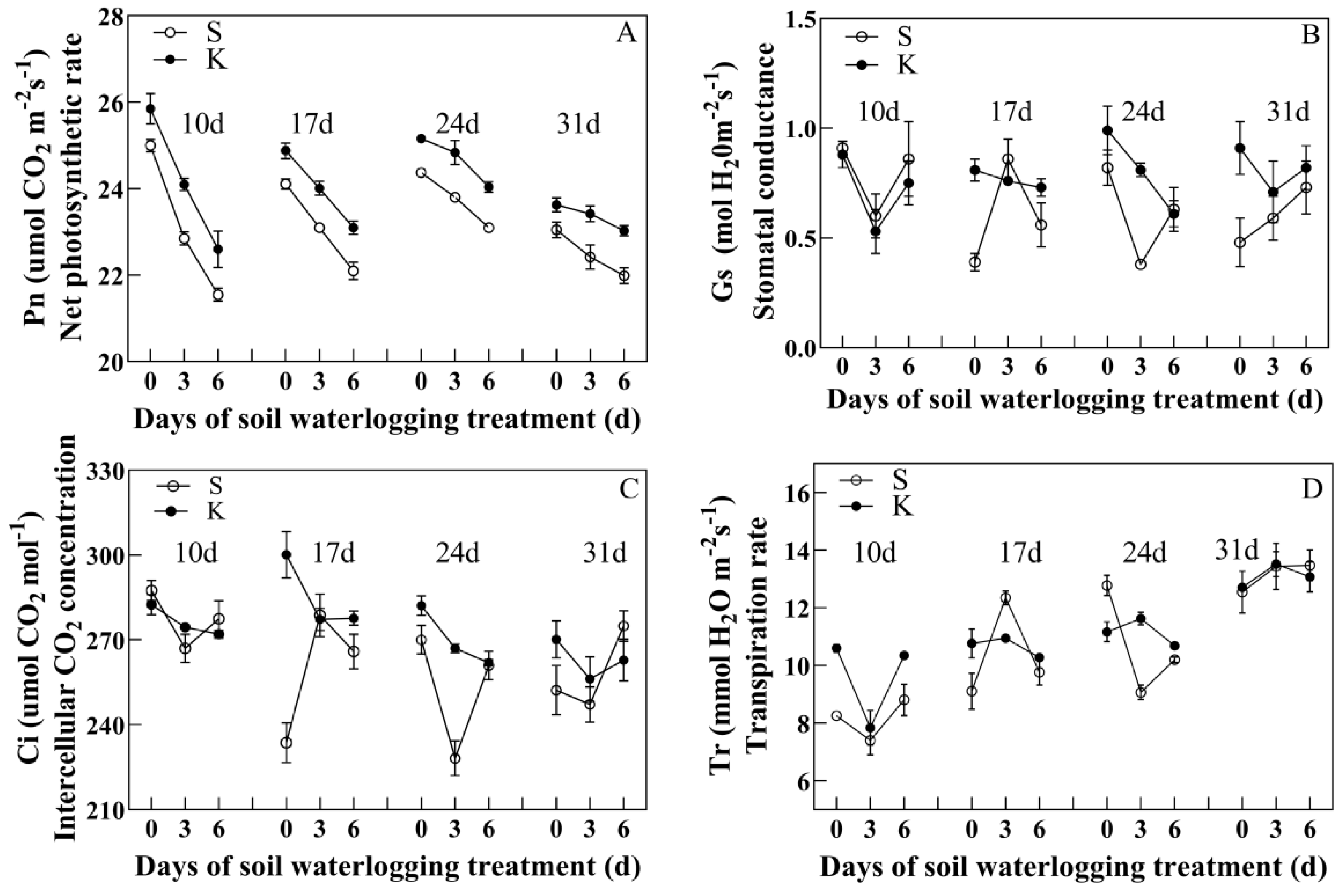
3.2. Changes in Leaf Gas Exchange Parameters
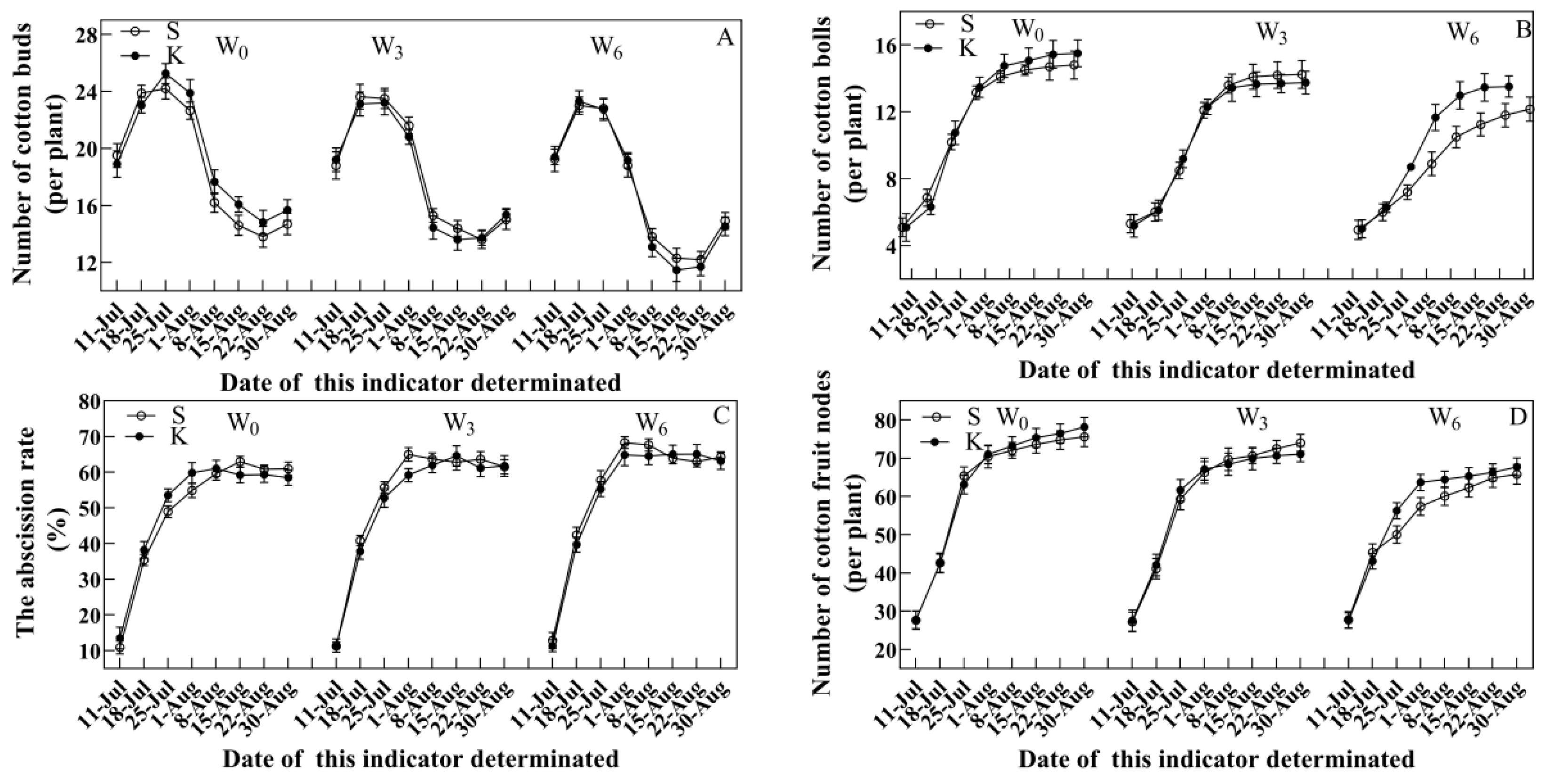
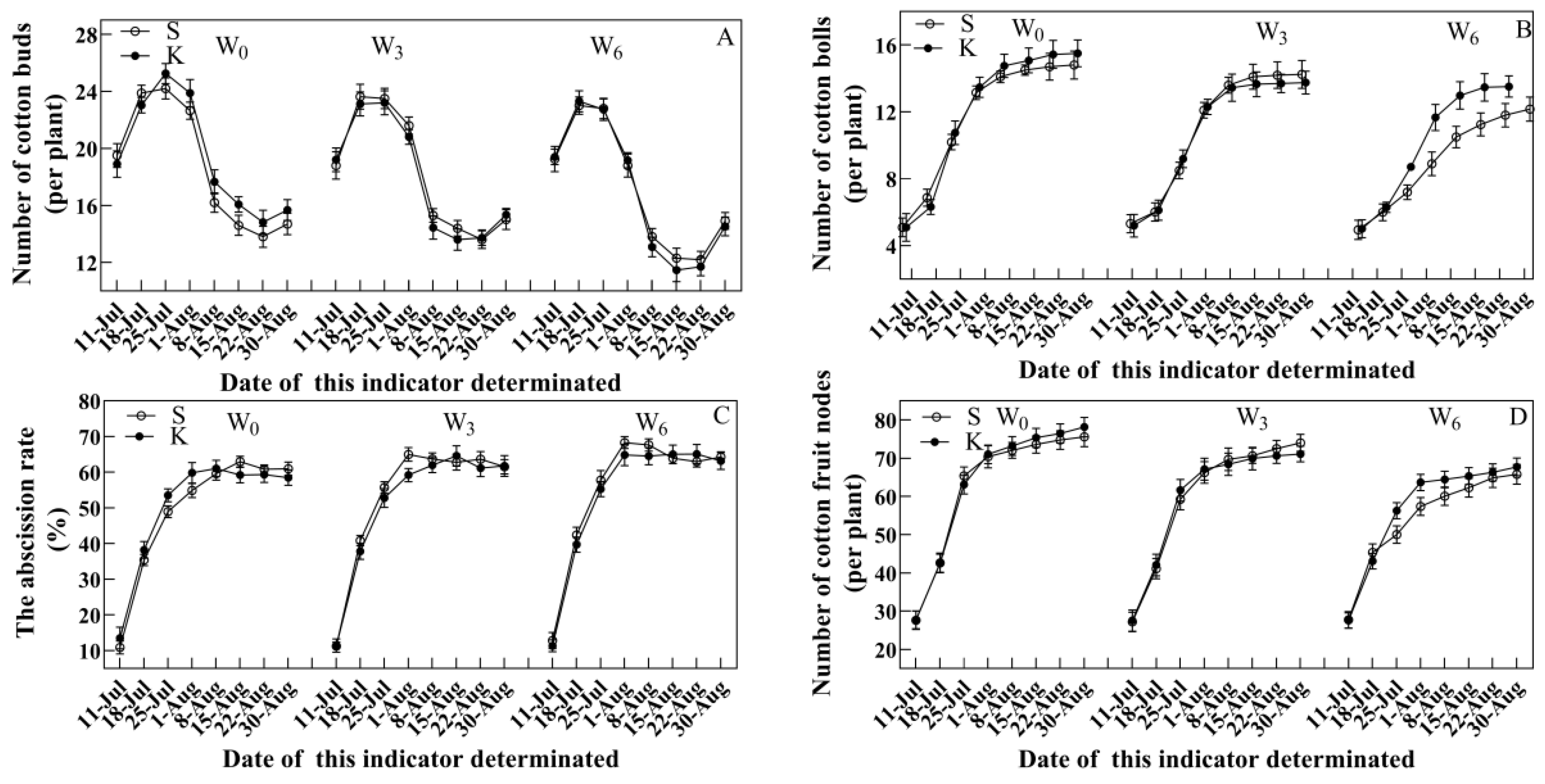
3.3. Agronomic Traits
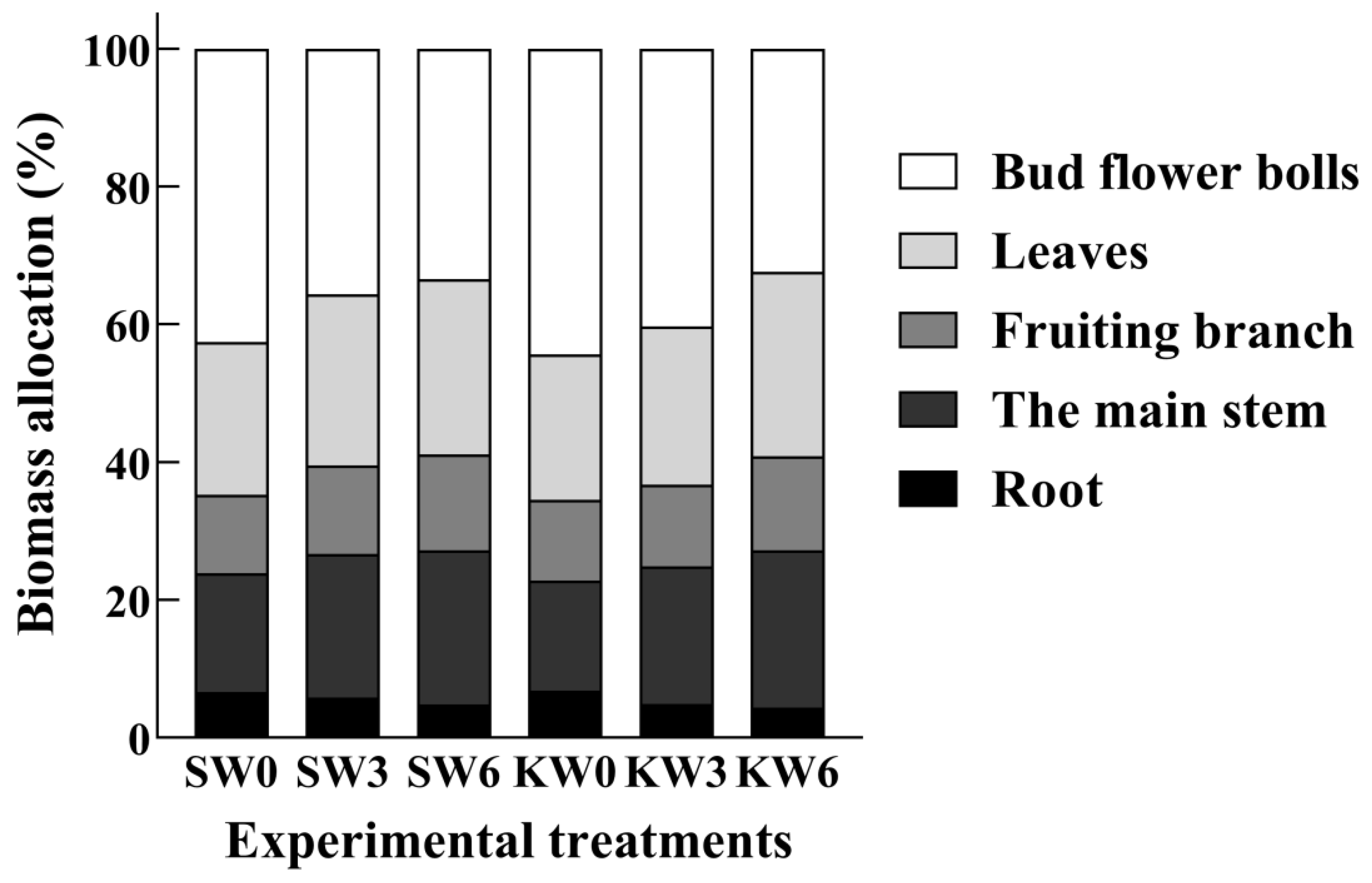
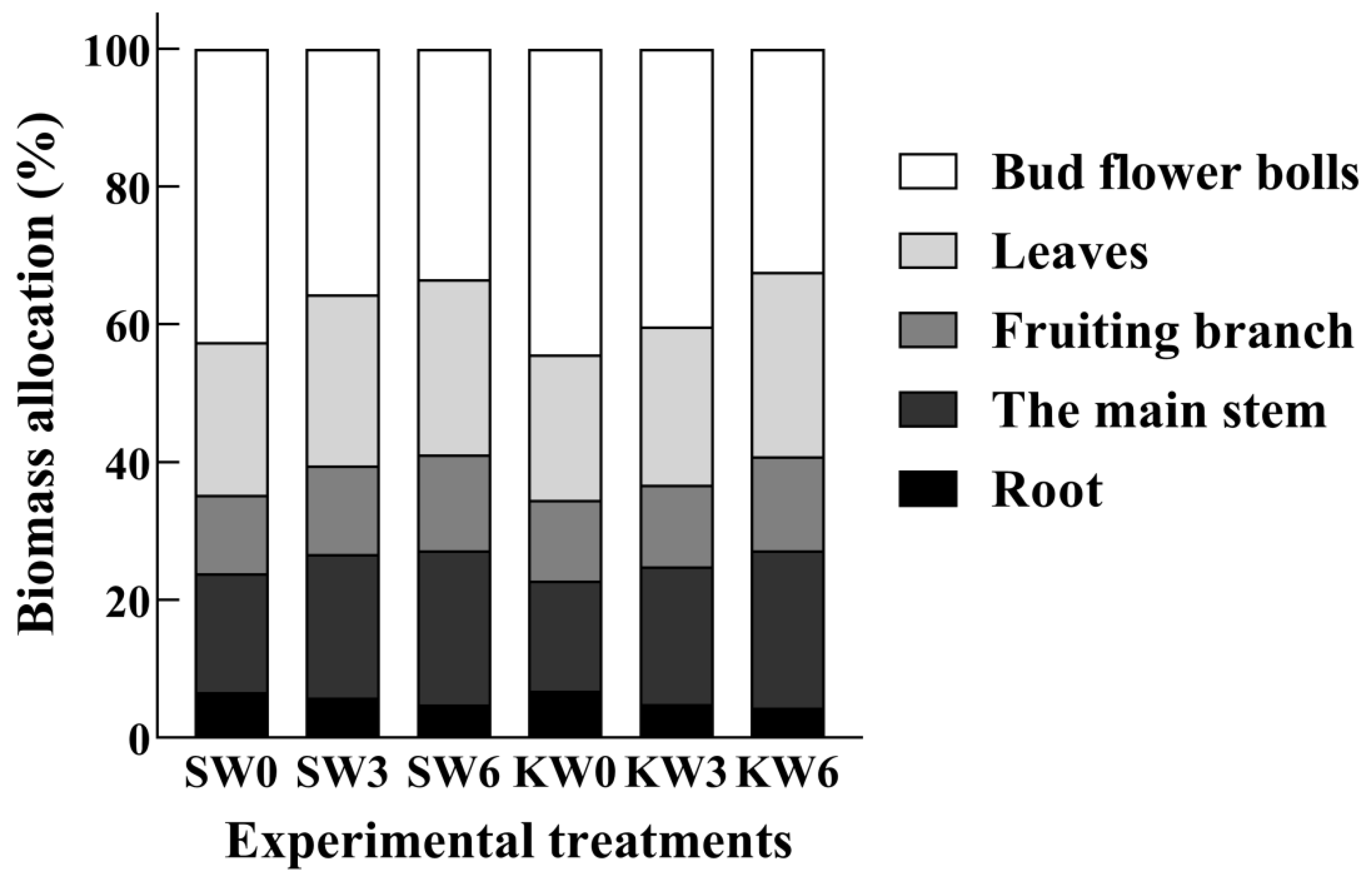
3.4. Dry Matter Accumulation and Allocation
4. Discussion
4.1. Potassium Application Improves Leaf Relative Water Content and Photosynthetic Capacity in Cotton Subjected to Soil Waterlogging Stress
4.2. Potassium Application Developed Better Agronomic Traits in Cotton Suffering from Soil Waterlogging Stress
4.3. Effect of Potassium Application on Dry Matter Accumulation and Distribution in Cotton Undergoing Soil Waterlogging Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, K.; Harrison, M.T.; Yan, H.; Liu, D.; Meinke, H.; Hoogenboom, G.; Wang, B.; Peng, B.; Guan, K.; Jaegermeyr, J.; et al. Silver lining to a climate crisis in multiple prospects for alleviating crop waterlogging under future climates. Nat. Commun. 2023, 14, 765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fueglistaler, S. Mechanism for Increasing Tropical Rainfall Unevenness With Global Warming. Geophys. Res. Lett. 2019, 46, 14836–14843. [Google Scholar] [CrossRef]
- Kuai, J.; Zhou, Z.G.; Wang, Y.H.; Meng, Y.L.; Chen, B.L.; Zhao, W.Q. The effects of short-term waterlogging on the lint yield and yield components of cotton with respect to boll position. Eur. J. Agron. 2015, 67, 61–74. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Liu, G.Y.; Dong, H.Z.; Li, C.D. Waterlogging stress in cotton: Damage, adaptability, alleviation strategies, and mechanisms. Crop J. 2021, 9, 257–270. [Google Scholar] [CrossRef]
- Hossain, M.A.; Uddin, S.N. Mechanisms of waterlogging tolerance in wheat: Morphological and metabolic adaptations under hypoxia or anoxia. Aust. J. Crop Sci. 2011, 5, 1094–1101. [Google Scholar]
- Drew, M.C. Plant injury and adaptation to oxygen deficiency in the root environment: A review. Plant Soil 1983, 75, 179–199. [Google Scholar] [CrossRef]
- Khan, A.; Pan, X.D.; Najeeb, U.; Tan, D.K.Y.; Fahad, S.; Zahoor, R.; Luo, H.H. Coping with drought: Stress and adaptive mechanisms, and management through cultural and molecular alternatives in cotton as vital constituents for plant stress resilience and fitness. Biol. Res. 2018, 51, 2–17. [Google Scholar] [CrossRef]
- Wang, H.; Liu, X.; Yang, P.; Wu, R.; Wang, S.; He, S.; Zhou, Q. Potassium application promote cotton acclimation to soil waterlogging stress by regulating endogenous protective enzymes activities and hormones contents. Plant Physiol. Biochem. 2022, 185, 336–343. [Google Scholar] [CrossRef]
- Zhang, R.; Yue, Z.; Chen, X.; Wang, Y.; Zhou, Y.; Xu, W.; Huang, R. Foliar applications of urea and melatonin to alleviate waterlogging stress on photosynthesis and antioxidant metabolism in sorghum seedlings. Plant Growth Regul. 2022, 97, 429–438. [Google Scholar] [CrossRef]
- Zahoor, R.; Dong, H.; Abid, M.; Zhao, W.; Wang, Y.; Zhou, Z. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Munsif, F.; Shah, T.; Arif, M.; Jehangir, M.; Afridi, M.Z.; Ahmad, I.; Jan, B.L.; Alansi, S. Combined effect of salicylic acid and potassium mitigates drought stress through the modulation of physio-biochemical attributes and key antioxidants in wheat. Saudi J. Biol. Sci. 2022, 29, 103294. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, G.; Motavalli, P.P.; Nelson, K.A.; Orlowski, J.M.; Golden, B.R. Impacts and management strategies for crop production in waterlogged or flooded soils: A review. Agron. J. 2019, 112, 1475–1501. [Google Scholar] [CrossRef]
- Ul-Allah, S.; Ijaz, M.; Nawaz, A.; Sattar, A.; Sher, A.; Naeem, M.; Shahzad, U.; Farooq, U.; Nawaz, F.; Mahmood, K. Potassium application improves grain yield and alleviates drought susceptibility in diverse maize hybrids. Plants 2020, 9, 75. [Google Scholar] [CrossRef]
- Zorb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture-status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef]
- Bamrungrai, J.; Polthanee, A.; Tubana, B.; Tre-loges, V.; Promkhambut, A. Effects of soil and foliar applied fertilizers on growth, yield and sugar quality of two sugarcane cultivars under rainfed conditions. Indian J. Agric. Res. 2022, 56, 545–550. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Raper, T.B. Potassium and stress alleviation: Physiological functions and management of cotton. Plant Nutr. Soil Sci. 2013, 176, 331–343. [Google Scholar] [CrossRef]
- Hu, W.; Lv, X.B.; Yang, J.S.; Chen, B.L.; Zhao, W.Q.; Meng, Y.; Wang, Y.H.; Zhou, Z.G.; Oosterhuis, D.M. Effects of potassium deficiency on antioxidant metabolism related to leaf senescence in cotton (Gossypium hirsutum L.). Field Crop Res. 2016, 191, 139–149. [Google Scholar] [CrossRef]
- Hu, W.; Coomer, T.D.; Loka, D.A.; Oosterhuis, D.M.; Zhou, Z. Potassium deficiency affects the carbon-nitrogen balance in cotton leaves. Plant Physiol. Biochem. 2017, 115, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Oosterhuis, D.; Bednarz, C. Influence of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultra structure of cotton plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Reddy, K.R.; Zhao, D. Interactive effects of elevated CO2 and potassium deficiency on photosynthesis, growth, and biomass partitioning of cotton. Field Crop Res. 2005, 94, 201–213. [Google Scholar] [CrossRef]
- Makhdum, M.L.; Pervez, H.; Ashraf, M. Dry matter accumulation and partitioning in cotton (Gossypium hirsutum L.) as influenced by potassium fertilization. Biol. Fert. Soils 2007, 43, 295–301. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Saur, E.; Constantin, J.; Porte, A.; Jordan-Meille, L. Effect of carbon assimilation on dry weight production and partitioning during vegetative growth of K-deficient cotton (Gossypium hirsutum L.) plants. Plant Soil 2009, 324, 329–343. [Google Scholar] [CrossRef]
- Hu, W.; Yang, J.; Meng, Y.; Wang, Y.; Chen, B.; Zhao, W.; Oosterhuis, D.M.; Zhou, Z. Potassium application affects carbohydrate metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll and its relationship with boll biomass. Field Crops Res. 2015, 179, 120–131. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Ahmad, M.S.A.; Ashraf, M.; Al-Qurainy, F.; Ashraf, M.Y. Alleviation of waterlogging stress in upland cotton (Gossypium hirsutum L.) by exogenous application of potassium in soil and as a foliar spray. Crop Pasture Sci. 2011, 62, 25–38. [Google Scholar] [CrossRef]
- Zahoor, R.; Zhao, W.; Abid, M.; Dong, H.; Zhou, Z. Potassium application regulates nitrogen metabolism and osmotic adjustment in cotton (Gossypium hirsutum L.) functional leaf under drought stress. J. Plant Physiol. 2017, 215, 30–38. [Google Scholar] [CrossRef]
- Zou, J.; Hu, W.; Li, Y.-x.; He, J.-q.; Zhu, H.-h.; Zhou, Z.-g. Screening of drought resistance indices and evaluation of drought resistance in cotton (Gossypium hirsutum L.). J. Integr. Agric. 2020, 19, 495–508. [Google Scholar] [CrossRef]
- Snider, J.L.; Pilon, C.; Hu, W.; Wang, H.M.; Tishchenko, V.; Slaton, W.; Chastain, D.; Parkash, V. Net photosynthesis acclimates to low growth temperature in cotton seedlings by shifting temperature thresholds for photosynthetic component processes and respiration. Environ. Exp. Bot. 2022, 196, 104816. [Google Scholar] [CrossRef]
- Kuai, J.; Liu, Z.W.; Wang, Y.H.; Meng, Y.L.; Chen, B.L.; Zhao, W.Q.; Zhou, Z.G.; Oosterhuis, D.M. Waterlogging during flowering and boll forming stages affects sucrose metabolism in the leaves subtending the cotton boll and its relationship with boll weight. Plant Sci. 2014, 223, 79–98. [Google Scholar] [CrossRef]
- Luo, Z.; Dong, H.; Li, W.; Tang, W.; Zhang, D. Combined effects of waterlogging and salinity on plant growth and some physiological parameters in cotton seedling leaves. Cotton Sci. 2008, 20, 203–206. [Google Scholar]
- Pettigrew, W.; Gerik, T.J. Cotton leaf photosynthesis and carbon metabolism. Adv. Agron. 2007, 94, 209–236. [Google Scholar]
- Li, C.Y.; Jiang, D.; Wollenweber, B.; Li, Y.; Dai, T.B.; Cao, W.X. Waterlogging pretreatment during vegetative growth improves tolerance to waterlogging after anthesis in wheat. Plant Sci. 2011, 180, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Ou, Z.; Shen, W.; Peng, Y.; Zhu, J.; Zheng, W. Photosynthetic and growth characteristics of different Excentrodendron hsienmu provenances from Karst district of southwestern Guangxi, China. J. Trop. Subtro. Bot. 2018, 22, 164–170. [Google Scholar]
- Li, X.; Li, N.; Yang, J.; Ye, F.; Chen, F.; Chen, F. Morphological and photosynthetic responses of riparian plant Distylium chinense seedlings to simulated autumn and winter flooding in three gorges reservoir region of the Yangtze River, China. Acta Ecol. Sin. 2011, 31, 31–39. [Google Scholar]
- Aasamaa, K.; Sober, A. Responses of stomatal conductance to simultaneous changes in two environmental factors. Tree Physiol. 2011, 31, 855–864. [Google Scholar] [CrossRef]
- Wang, H.; Yan, S.; Xin, H.; Huang, W.; Zhang, H.; Teng, S.; Yu, Y.C.; Fernie, A.R.; Lu, X.; Li, P.; et al. A subsidiary cell-localized glucose transporter promotes stomatal conductance and photosynthesis. Plant Cell 2019, 31, 1328–1343. [Google Scholar] [CrossRef] [PubMed]
- Jan, M.; ul Haq, M.A.; ul Haq, T.; Ali, A.; Hussain, S.; Ibrahim, M. Protective effect of potassium application on NaCl induced stress in tomato (Lycopersicon esculentum L.) genotypes. J. Plant Nutr. 2020, 43, 1988–1998. [Google Scholar] [CrossRef]
- Benlloch-González, M.; Arquero, O.; Fournier, J.M.; Barranco, D.; Benlloch, M. K+starvation inhibits water-stress-induced stomatal closure. J. Plant Physiol. 2008, 165, 623–630. [Google Scholar] [CrossRef]
- Sakarvadia, H.L.; Polara, K.B.; Davaria, R.L.; Parmar, K.B.; Babariya, N.B. Effect of potassium and zinc on growth, yield, quality parameters and nutrient uptake by Bt cotton. Soil Sci. 2012, 7, 319–323. [Google Scholar]
- Bednarz, C.W.; Bridges, D.C.; Brown, S.M. Analysis of cotton yield stability across population densities. Agron. J. 2000, 92, 128–135. [Google Scholar] [CrossRef]
- Bange, M.P.; Milroy, S.P.; Thongbai, P. Growth and yield of cotton in response to waterlogging. Field Crops Res. 2004, 88, 129–142. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, Y.; Lu, H.; Kong, X.; Dai, J.; Li, Z.; Dong, H. Growth, lint yield and changes in physiological attributes of cotton under temporal waterlogging. Field Crops Res. 2016, 194, 83–93. [Google Scholar] [CrossRef]
- Liu, K.; Harrison, M.T.; Wang, B.; Yang, R.; Yan, H.; Zou, J.; Liu, D.; Meinke, H.; Tian, X.; Ma, S.; et al. Designing high-yielding wheat crops under late sowing: A case study in southern China. Agron. Sustain. Dev. 2022, 42, 29. [Google Scholar] [CrossRef]
- Yan, H.; Harrison, M.T.; Liu, K.; Wang, B.; Feng, P.; Fahad, S.; Meinke, H.; Yang, R.; Liu, D.; Archontoulis, S.; et al. Crop traits enabling yield gains under more frequent extreme climatic events. Sci. Total Environ. 2022, 808, 152170. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, A.S. The effects of duration: Timing and chemical amelioration of short-term waterlogging during furrow irrigation of cotton in a cracking grey clay. Aust. J. Agric. Res. 1982, 33, 1019–1028. [Google Scholar] [CrossRef]
- Colmer, T.D.; Voesenek, L.A.C.J. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665. [Google Scholar] [CrossRef]
- Trought, M.C.T.; Drew, M.C. The development of waterlogging damage in wheat seedlings (Triticum aestivum L.). Plant Soil 1980, 54, 77–94. [Google Scholar] [CrossRef]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S.T.; Liu, Y.; Cui, Y.; Zahoor, R.; Jiang, D.; Dai, T. Improved tolerance to post-anthesis drought stress by pre-drought prim priming at vegetative stages in drought-tolerant and -sensitive wheat cultivars. Plant Physiol. Biochem. 2016, 106, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Mengel, K.; Haeder, H.E. Effect of potassium supply on the rates of phloem sap exudation and the composition of phloem sap of Ricinus communis. Plant Physiol. 1977, 59, 282–284. [Google Scholar] [CrossRef]
- Patrick, J.W.; Zhang, W.; Tyerman, S.D.; Offler, C.E.; Walker, N.A. Role of membrane transport in phloem translocation of assimilates and water. Funct. Plant Biol. 2001, 28, 697–709. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Treatments | Root | Main Stem | Fruiting Branch | Leaf | Bud Flower Bolls | Total Biomass |
|---|---|---|---|---|---|---|
| SW0 | 23.72 ± 0.64 b | 61.88 ± 3.74 a | 79.38 ± 1.51 a | 40.93 ± 2.41 b | 152.69 ± 3.68 b | 358.58 ± 4.14 b |
| SW3 | 15.23 ± 1.32 d | 62.26 ± 4.90 a | 71.57± 6.00 c | 37.10 ± 1.05 c | 125.75 ± 5.69 c | 311.90 ± 4.86 cd |
| SW6 | 11.31 ± 0.35 e | 58.95 ±0.48 b | 69.04 ± 0.86 d | 35.18 ± 6.52 e | 83.54 ± 1.73 d | 258.01 ± 7.52 e |
| KW0 | 25.33 ± 1.35 a | 59.60 ± 5.55 a | 78.30 ± 5.02 a | 43.59 ± 1.55 a | 165.14 ± 10.77 a | 371.94 ± 0.00 a |
| KW3 | 17.21 ± 0.88 c | 61.17 ± 3.83 a | 73.12± 5.59 b | 37.75 ± 1.32 c | 104.84 ± 2.67 c | 294.09 ± 6.31 d |
| KW6 | 12.58 ± 0.66 e | 58.43 ± 0.77 b | 66.43 ± 8.82 d | 36.31 ± 5.89 d | 87.31 ± 5.37 d | 261.05 ± 3.87 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Li, J.; Yang, P.; Zeng, X.; Chen, Y.; Wang, H. Potassium Application Alleviated Negative Effects of Soil Waterlogging Stress on Photosynthesis and Dry Biomass in Cotton. Agronomy 2023, 13, 1157. https://doi.org/10.3390/agronomy13041157
Huang L, Li J, Yang P, Zeng X, Chen Y, Wang H. Potassium Application Alleviated Negative Effects of Soil Waterlogging Stress on Photosynthesis and Dry Biomass in Cotton. Agronomy. 2023; 13(4):1157. https://doi.org/10.3390/agronomy13041157
Chicago/Turabian StyleHuang, Li, Jinxiang Li, Pan Yang, Xianghua Zeng, Yinyi Chen, and Haimiao Wang. 2023. "Potassium Application Alleviated Negative Effects of Soil Waterlogging Stress on Photosynthesis and Dry Biomass in Cotton" Agronomy 13, no. 4: 1157. https://doi.org/10.3390/agronomy13041157
APA StyleHuang, L., Li, J., Yang, P., Zeng, X., Chen, Y., & Wang, H. (2023). Potassium Application Alleviated Negative Effects of Soil Waterlogging Stress on Photosynthesis and Dry Biomass in Cotton. Agronomy, 13(4), 1157. https://doi.org/10.3390/agronomy13041157