



Endophytic Biostimulants for Smart Agriculture: Burkholderia seminalis 869T2 Benefits Heading Leafy Vegetables In-Field Management in Taiwan

 , , , and
, , , and 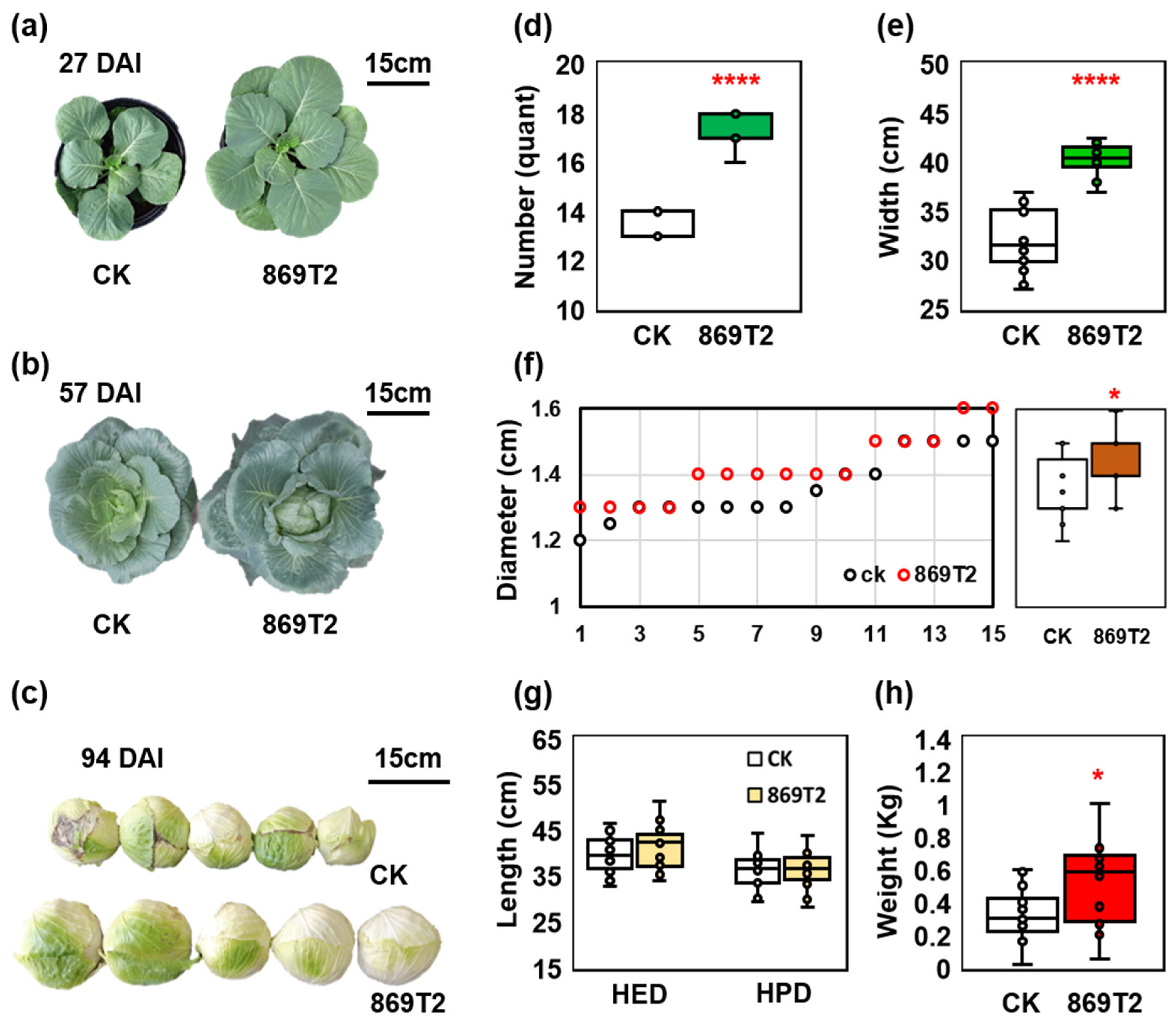{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Endophyte and Plant Materials
2.2. Plant–Endophyte Symbiosis Assays
2.3. Endophyte Inoculation Timing and Concentration Evaluation
2.4. Fermentation of Endophytic Strain for Lettuce In-Field Experiments
2.5. Smart Agriculture Management
2.6. Rheological Property Measurement
2.7. Chlorophyll Content Measurements
2.8. Measurement of Total Soluble Sugars and the Starch Content
2.9. Nutrient Element Measurement
2.10. Statistics
3. Results
3.1. B. seminalis 869T2 Improves the Growth of Cabbage in a Greenhouse
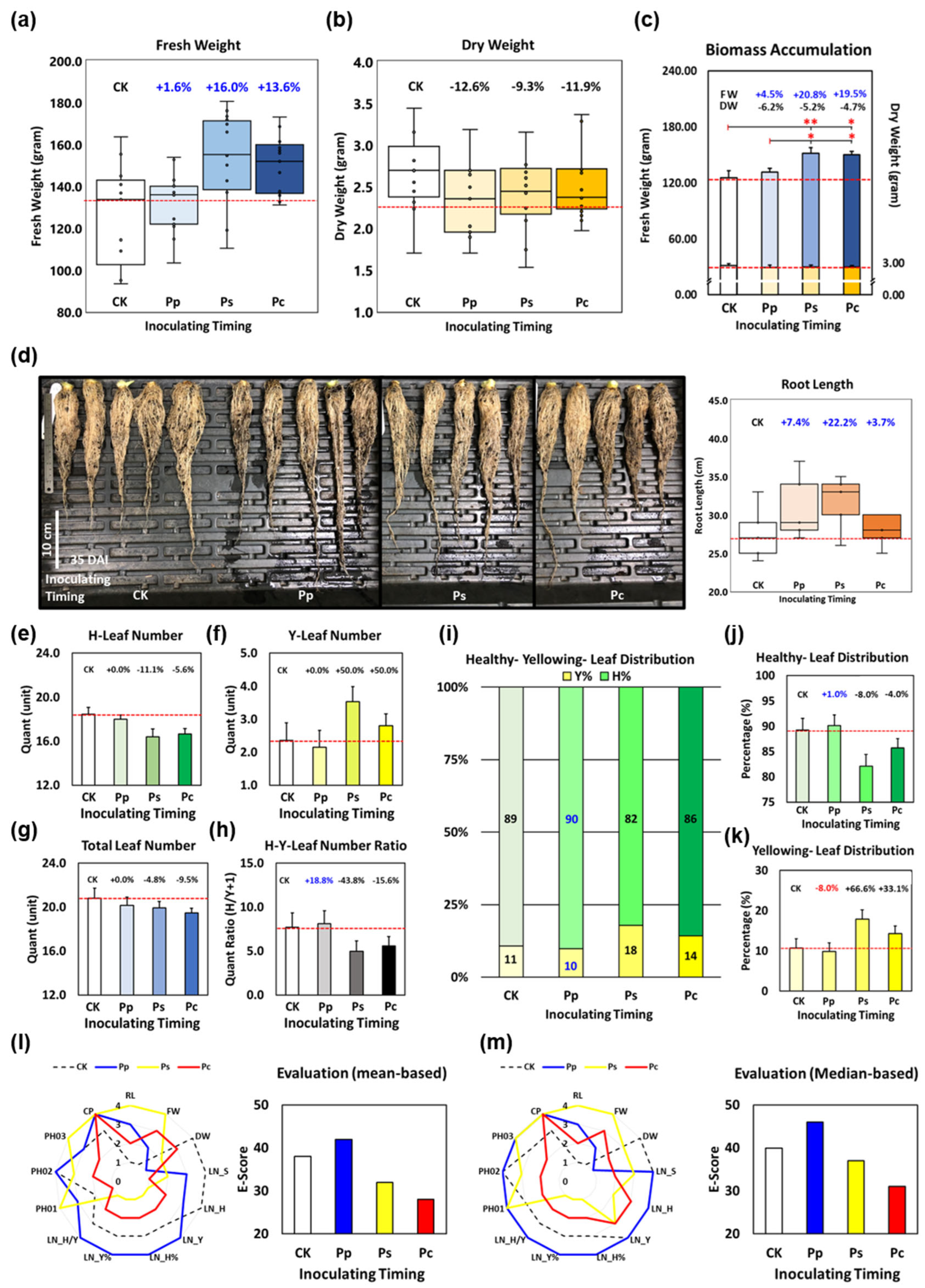
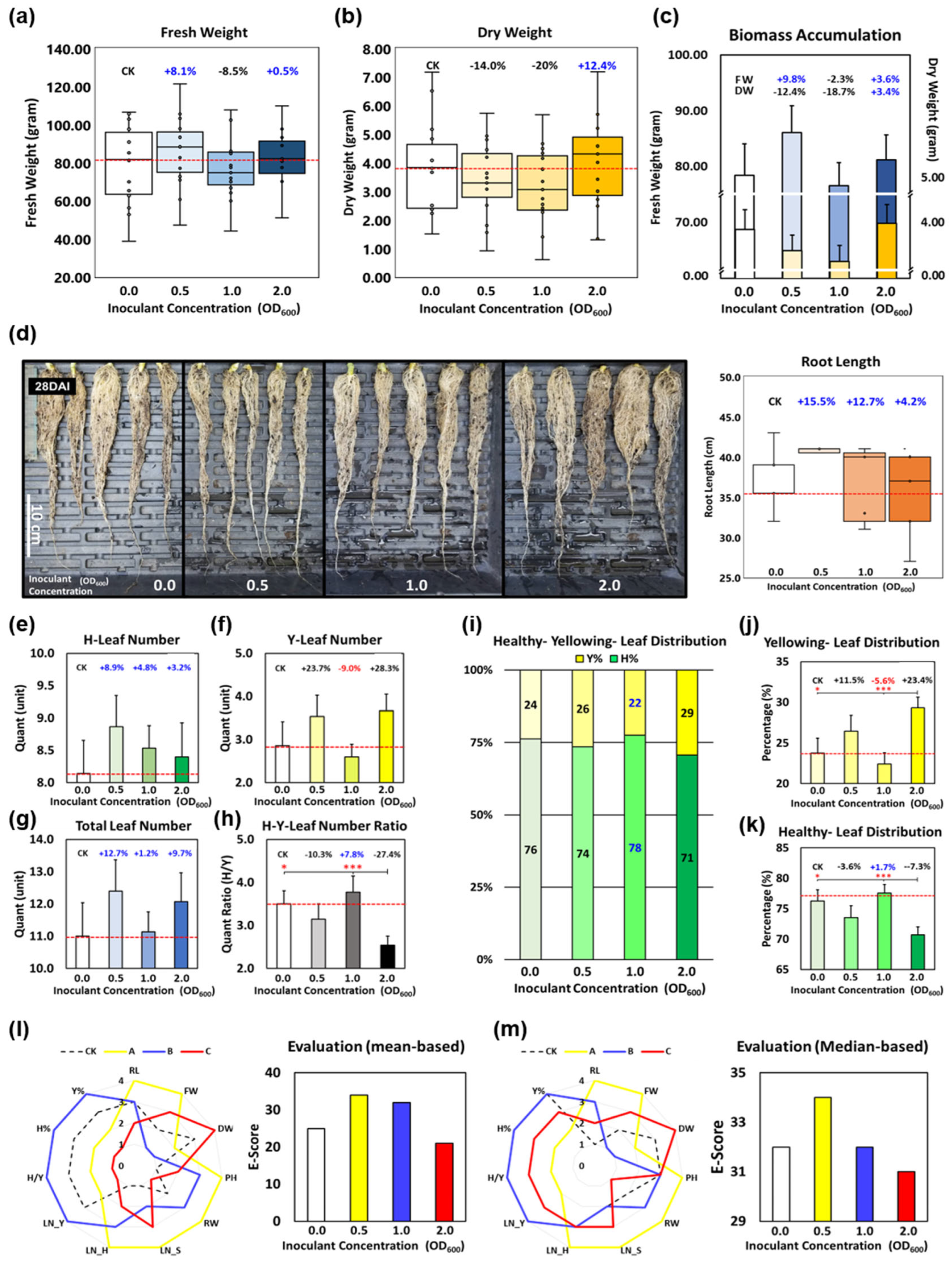
3.2. The Inoculation Timing and Dosage Matter in Endophytic Biostimulants Application
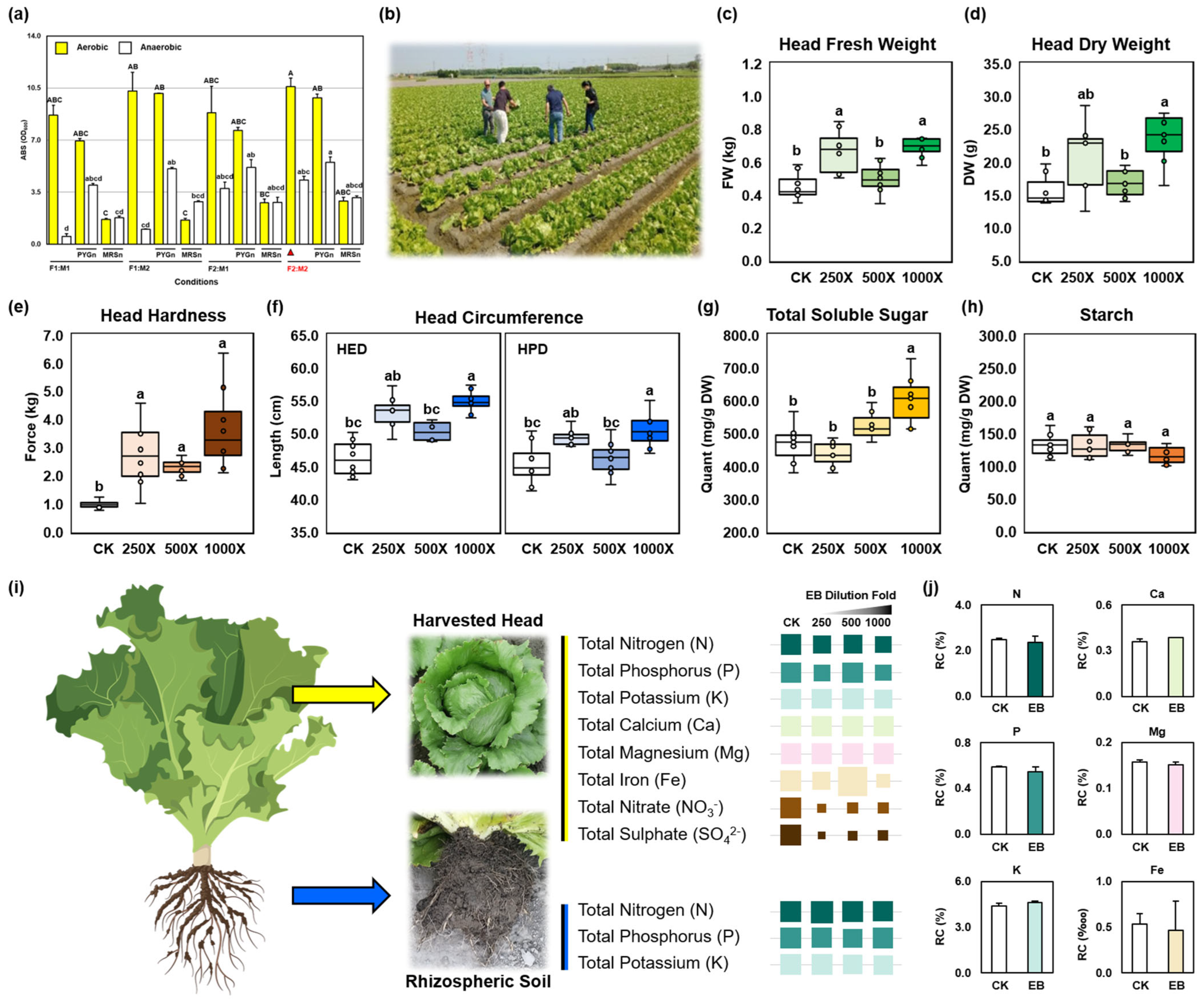
3.3. In-Field Lettuce Cultivation Improvement with Endophytic Biostimulants
3.4. Endophytic Biostimulants Save Costs in Agro-Industry
4. Discussion
4.1. B. seminalis Improves the Growth of Heading Vegetables and Protects against Stresses
4.2. Endophytic Biostimulant-Assisted Management for Sustainable Agriculture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Food Policy Research Institute. 2022 Global Food Policy Report: Climate Change and Food Systems; International Food Policy Research Institute: Washington, DC, USA, 2022. [Google Scholar]
- Swinnen, J.; Arndt, C.; Vos, R. Climate Change and Food Systems: Transforming Food Systems for Adaptation, Mitigation, and Resilience; International Food Policy Research Institute: Washington, DC, USA, 2022. [Google Scholar]
- Stads, G.-J.; Wiebe, K.D.; Nin-Pratt, A.; Sulser, T.B.; Benfica, R.; Reda, F.; Khetarpal, R. Research for the Future: Investments for Efficiency, Sustainability, and Equity; International Food Policy Research Institute: Washington, DC, USA, 2022. [Google Scholar]
- Falck-Zepeda, J.B.; Biermayr-Jenzano, P.; Roca, M.M.; Fuentes-Campos, E.; Kikulwe, E.M. Bio-Innovations: Genome-Edited Crops for Climate-Smart Food Systems; International Food Policy Research Institute: Washington, DC, USA, 2022. [Google Scholar]
- White, J.F.; Kingsley, K.L.; Zhang, Q.; Verma, R.; Obi, N.; Dvinskikh, S.; Elmore, M.T.; Verma, S.K.; Gond, S.K.; Kowalski, K.P. Review: Endophytic Microbes and Their Potential Applications in Crop Management. Pest. Manag. Sci. 2019, 75, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef]
- Elsheikh, E.A.E.; El-Keblawy, A.; Mosa, K.A.; Okoh, A.I.; Saadoun, I. Role of Endophytes and Rhizosphere Microbes in Promoting the Invasion of Exotic Plants in Arid and Semi-Arid Areas: A Review. Sustainability 2021, 13, 13081. [Google Scholar] [CrossRef]
- Ha-Tran, D.M.; Nguyen, T.T.M.; Hung, S.-H.; Huang, E.; Huang, C.-C. Roles of Plant Growth-Promoting Rhizobacteria (PGPR) in Stimulating Salinity Stress Defense in Plants: A Review. Int. J. Mol. Sci. 2021, 22, 3154. [Google Scholar] [CrossRef] [PubMed]
- Verma, H.; Kumar, D.; Kumar, V.; Kumari, M.; Singh, S.K.; Sharma, V.K.; Droby, S.; Santoyo, G.; White, J.F.; Kumar, A. The Potential Application of Endophytes in Management of Stress from Drought and Salinity in Crop Plants. Microorganisms 2021, 9, 1729. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted Interactions Between Endophytes and Plant: Developments and Prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Baldani, J.; Caruso, L.; Baldani, V.L.D.; Goi, S.R.; Döbereiner, J. Recent Advances in BNF with Non-Legume Plants. Soil Biol. Biochem. 1997, 29, 911–922. [Google Scholar] [CrossRef]
- James, E.K.; Olivares, F.L.; Baldani, J.I.; Döbereiner, J. Herbaspirillum, an Endophytic Diazotroph Colonizing Vascular Tissue 3 Sorghum bicolor L. Moench. J. Exp. Bot. 1997, 48, 785–798. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Hwang, H.-H.; Chien, P.-R.; Huang, F.-C.; Hung, S.-H.; Kuo, C.-H.; Deng, W.-L.; Chiang, E.-P.I.; Huang, C.-C. A Plant Endophytic Bacterium, Burkholderia Seminalis Strain 869T2, Promotes Plant Growth in Arabidopsis, Pak Choi, Chinese Amaranth, Lettuces, and Other Vegetables. Microorganisms 2021, 9, 1703. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Seo, J.; Peng, Y.; Jeon, D.; Park, S.J.; Kim, C.Y.; Kim, P.I.; Kim, C.H.; Lee, J.H.; Lee, J. Genome Insights into the Plant Growth-Promoting Bacterium Saccharibacillus brassicae ATSA2T. AMB Express 2023, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Szymańska, S.; Lis, M.I.; Piernik, A.; Hrynkiewicz, K. Pseudomonas stutzeri and Kushneria marisflavi Alleviate Salinity Stress-Associated Damages in Barley, Lettuce, and Sunflower. Front. Microbiol. 2022, 13, 788893. [Google Scholar] [CrossRef] [PubMed]
- Frommel, M.I.; Nowak, J.; Lazarovits, G. Growth Enhancement and Developmental Modifications of In Vitro Grown Potato (Solanum tuberosum spp. tuberosum) as Affected by a Nonfluorescent Pseudomonas Sp. 1. Plant Physiol. 1991, 96, 928–936. [Google Scholar] [CrossRef]
- Sessitsch, A.; Coenye, T.; Sturz, A.V.; Vandamme, P.; Barka, E.A.; Salles, J.F.; Van Elsas, J.D.; Faure, D.; Reiter, B.; Glick, B.R.; et al. Burkholderia phytofirmans Sp. Nov., a Novel Plant-Associated Bacterium with Plant-Beneficial Properties. Int. J. Syst. Evol. Microbiol. 2005, 55, 1187–1192. [Google Scholar] [CrossRef]
- Weilharter, A.; Mitter, B.; Shin, M.V.; Chain, P.S.G.; Nowak, J.; Sessitsch, A. Complete Genome Sequence of the Plant Growth-Promoting Endophyte Burkholderia phytofirmans Strain PsJN. J. Bacteriol. 2011, 193, 3383–3384. [Google Scholar] [CrossRef]
- Mitter, B.; Petric, A.; Shin, M.; Chain, P.; Hauberg-Lotte, L.; Reinhold-Hurek, B.; Nowak, J.; Sessitsch, A. Comparative Genome Analysis of Burkholderia phytofirmans PsJN Reveals a Wide Spectrum of Endophytic Lifestyles Based on Interaction Strategies with Host Plants. Front. Plant Sci. 2013, 4, 120. [Google Scholar] [CrossRef]
- Ali, S.; Duan, J.; Charles, T.C.; Glick, B.R. A Bioinformatics Approach to the Determination of Genes Involved in Endophytic Behavior in Burkholderia spp. J. Theor. Biol. 2014, 343, 193–198. [Google Scholar] [CrossRef]
- Pinski, A.; Betekhtin, A.; Hupert-Kocurek, K.; Mur, L.A.J.; Hasterok, R. Defining the Genetic Basis of Plant–Endophytic Bacteria Interactions. Int. J. Mol. Sci. 2019, 20, 1947. [Google Scholar] [CrossRef]
- Mesny, F.; Miyauchi, S.; Thiergart, T.; Pickel, B.; Atanasova, L.; Karlsson, M.; Hüttel, B.; Barry, K.W.; Haridas, S.; Chen, C.; et al. Genetic Determinants of Endophytism in the Arabidopsis Root Mycobiome. Nat. Commun. 2021, 12, 7227. [Google Scholar] [CrossRef]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- EU EUR-Lex-L:2019:170:TOC-EN-EUR-Lex. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=OJ%3AL%3A2019%3A170%3ATOC (accessed on 19 October 2022).
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and Fulvic Acids as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein Hydrolysates as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed Extracts as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Savvas, D.; Ntatsi, G. Biostimulant Activity of Silicon in Horticulture. Sci. Hortic. 2015, 196, 66–81. [Google Scholar] [CrossRef]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; Pascale, S.D.; Bonini, P.; Colla, G. Arbuscular Mycorrhizal Fungi Act as Biostimulants in Horticultural Crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant Growth-Promoting Rhizobacteria Act as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Mishra, A.K.; Choi, J.; Choi, S.-J.; Baek, K.-H. Cyclodipeptides: An Overview of Their Biosynthesis and Biological Activity. Molecules 2017, 22, 1796. [Google Scholar] [CrossRef]
- Carreño-López, R.; Alatorre-Cruz, J.M.; Marín-Cevada, V. Pyrroloquinoline Quinone (PQQ): Role in Plant-Microbe Interactions. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms: Discovery and Applications; Singh, H.B., Keswani, C., Reddy, M.S., Sansinenea, E., García-Estrada, C., Eds.; Springer: Singapore, 2019; pp. 169–184. ISBN 9789811358623. [Google Scholar]
- Ho, Y.-N.; Chiang, H.-M.; Chao, C.-P.; Su, C.-C.; Hsu, H.-F.; Guo, C.; Hsieh, J.-L.; Huang, C.-C. In Planta Biocontrol of Soilborne Fusarium Wilt of Banana through a Plant Endophytic Bacterium, Burkholderia cenocepacia 869T2. Plant Soil 2015, 387, 295–306. [Google Scholar] [CrossRef]
- Hung, S.-H.W.; Chiu, M.-C.; Huang, C.-C.; Kuo, C.-H. Complete Genome Sequence of Curtobacterium Sp. C1, a Beneficial Endophyte with the Potential for In-Plant Salinity Stress Alleviation. MPMI 2022, 35, 731–735. [Google Scholar] [CrossRef]
- Hwang, H.-H.; Chien, P.-R.; Huang, F.-C.; Yeh, P.-H.; Hung, S.-H.W.; Deng, W.-L.; Huang, C.-C. A Plant Endophytic Bacterium Priestia Megaterium StrainBP-R2 Isolated from the Halophyte Bolboschoenus Planiculmis Enhances Plant Growth under Salt and Drought Stresses. Microorganisms 2022, 10, 2047. [Google Scholar] [CrossRef]
- Ho, Y.-N.; Mathew, D.C.; Hsiao, S.-C.; Shih, C.-H.; Chien, M.-F.; Chiang, H.-M.; Huang, C.-C. Selection and Application of Endophytic Bacterium Achromobacter xylosoxidans Strain F3B for Improving Phytoremediation of Phenolic Pollutants. J. Hazard. Mater. 2012, 219–220, 43–49. [Google Scholar] [CrossRef]
- Ho, Y.-N.; Huang, C.-C. Draft Genome Sequence of Burkholderia cenocepacia Strain 869T2, a Plant-Beneficial Endophytic Bacterium. Genome Announc. 2015, 3, e01327-15. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.-A.T.; Hsieh, J.-L.; Lo, S.-C.; Wang, S.-Y.; Hung, C.-H.; Huang, E.; Hung, S.-H.; Chin, W.-C.; Huang, C.-C. Biodegradation of Dioxins by Burkholderia cenocepacia Strain 869T2: Role of 2-Haloacid Dehalogenase. J. Hazard. Mater. 2021, 401, 123347. [Google Scholar] [CrossRef] [PubMed]
- Araújo, F.D.d.S.; Araújo, W.L.; Eberlin, M.N. Potential of Burkholderia seminalis TC3.4.2R3 as Biocontrol Agent Against Fusarium oxysporum Evaluated by Mass Spectrometry Imaging. J. Am. Soc. Mass Spectrom. 2017, 28, 901–907. [Google Scholar] [CrossRef]
- BACI-CEPII CEPII—BACI: International Trade Database at the Product-Level. The 1994–2007 Version. Available online: http://www.cepii.fr/CEPII/en/publications/wp/abstract.asp?NoDoc=2726 (accessed on 19 October 2022).
- Hsieh, M.H.; Jiang, W.J. Application of Controlled Released Fertilizers for Iceberg Lettuce Cultivation. Res. Bull. Tainan Dist. Agric. Improv. Stn. 2014, 64, 20–35. [Google Scholar]
- Hsieh, M.H.; Lin, J.W.; Jiang, W.J.; Lee, C. Efficiency evaluation on single application of fertilizer in ridge to substitute multiple topdressings for iceberg lettuce production. Res. Bull. Tainan Dist. Agric. Improv. Stn. 2018, 71, 34–45. [Google Scholar]
- Hsieh, M.H.; Jiang, W.J.; Lin, C.W. Application of Controlled Release Fertilizers in Iceburg Lettuce to Reduce Fertilizer Consumption. Res. Bull. Tainan Dist. Agric. Improv. Stn. 2015, 66, 39–51. [Google Scholar]
- Hsieh, M.H.; Lin, C.W. Application of Fertilizer in Lettuce to Reduce Tipburn Rate. Res. Bull. Tainan Dist. Agric. Improv. Stn. 2016, 67, 24–37. [Google Scholar]
- Chen, Y.-J.; Lin, H.-L. Effects of Blanching on Piquancy of Garlic (Allium sativum L. Cv. ‘Ta-Pen Hey’) Clove. Hortic. NCHU 2016, 41, 83–89. [Google Scholar]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Doran, L.; Souza, A.P.D. Total Soluble Sugar Quantification from Ethanolic Plant Extracts. Available online: https://www.protocols.io/view/total-soluble-sugar-quantification-from-ethanolic-b2nsqdee (accessed on 20 October 2022).
- Doran, L.; Souza, A.P.D. Extraction of Non-Structural Carbohydrates (Total Soluble Sugars + Starch) in Plant Tissues. Available online: https://www.protocols.io/view/extraction-of-non-structural-carbohydrates-total-s-b2jmqck6 (accessed on 20 October 2022).
- Agriculture Fertilizer Standards of Taiwan. Available online: https://www.afa.gov.tw/cht/index.php?act=download&ids=4829 (accessed on 25 April 2021).
- Theunissen, J.; Sins, A. Growth Stages of Brassica Crops for Crop Protection Purposes. Sci. Hortic. 1984, 24, 1–11. [Google Scholar] [CrossRef]
- Shen, H.; He, X.; Liu, Y.; Chen, Y.; Tang, J.; Guo, T. A Complex Inoculant of N2-Fixing, P- and K-Solubilizing Bacteria from a Purple Soil Improves the Growth of Kiwifruit (Actinidia chinensis) Plantlets. Front. Microbiol. 2016, 7, 841. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Sahu, P.K.; Kumar, K.; Pal, G.; Gond, S.K.; Kharwar, R.N.; White, J.F. Endophyte Roles in Nutrient Acquisition, Root System Architecture Development and Oxidative Stress Tolerance. J. Appl. Microbiol. 2021, 131, 2161–2177. [Google Scholar] [CrossRef]
- Grabka, R.; d’Entremont, T.W.; Adams, S.J.; Walker, A.K.; Tanney, J.B.; Abbasi, P.A.; Ali, S. Fungal Endophytes and Their Role in Agricultural Plant Protection against Pests and Pathogens. Plants 2022, 11, 384. [Google Scholar] [CrossRef]
- NCDR. Climate Change in Taiwan 2017: Scientific Report—The Physical Science Basis; National Science and Technology Center for Disaster Reduction: Taipei, Taiwan, 2017. [Google Scholar]
- Hersbach, H.; Bell, B.; Berrisford, P.; Hirahara, S.; Horányi, A.; Muñoz-Sabater, J.; Nicolas, J.; Peubey, C.; Radu, R.; Schepers, D.; et al. The ERA5 Global Reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- Park, H.J.; Jung, W.Y.; Lee, S.S.; Song, J.H.; Kwon, S.-Y.; Kim, H.; Kim, C.; Ahn, J.C.; Cho, H.S. Use of Heat Stress Responsive Gene Expression Levels for Early Selection of Heat Tolerant Cabbage (Brassica oleracea L.). Int. J. Mol. Sci. 2013, 14, 11871–11894. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant Growth-Promoting Bacterial Endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- CEPAL. The 2030 Agenda and the Sustainable Development Goals: An Opportunity for Latin America and the Caribbean; CEPAL: Santiago, Chile, 2018. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, S.-H.W.; Huang, T.-C.; Lai, Y.-C.; Wu, I.-C.; Liu, C.-H.; Huarng, Y.-F.; Hwang, H.-H.; Chiang, E.-P.I.; Kuo, C.-H.; Huang, C.-C. Endophytic Biostimulants for Smart Agriculture: Burkholderia seminalis 869T2 Benefits Heading Leafy Vegetables In-Field Management in Taiwan. Agronomy 2023, 13, 967. https://doi.org/10.3390/agronomy13040967
Hung S-HW, Huang T-C, Lai Y-C, Wu I-C, Liu C-H, Huarng Y-F, Hwang H-H, Chiang E-PI, Kuo C-H, Huang C-C. Endophytic Biostimulants for Smart Agriculture: Burkholderia seminalis 869T2 Benefits Heading Leafy Vegetables In-Field Management in Taiwan. Agronomy. 2023; 13(4):967. https://doi.org/10.3390/agronomy13040967
Chicago/Turabian StyleHung, Shih-Hsun Walter, Tsai-Ching Huang, Yu-Chih Lai, I-Chen Wu, Chia-Ho Liu, Yuh-Feng Huarng, Hau-Hsuan Hwang, En-Pei Isabel Chiang, Chih-Horng Kuo, and Chieh-Chen Huang. 2023. "Endophytic Biostimulants for Smart Agriculture: Burkholderia seminalis 869T2 Benefits Heading Leafy Vegetables In-Field Management in Taiwan" Agronomy 13, no. 4: 967. https://doi.org/10.3390/agronomy13040967
APA StyleHung, S.-H. W., Huang, T.-C., Lai, Y.-C., Wu, I.-C., Liu, C.-H., Huarng, Y.-F., Hwang, H.-H., Chiang, E.-P. I., Kuo, C.-H., & Huang, C.-C. (2023). Endophytic Biostimulants for Smart Agriculture: Burkholderia seminalis 869T2 Benefits Heading Leafy Vegetables In-Field Management in Taiwan. Agronomy, 13(4), 967. https://doi.org/10.3390/agronomy13040967