


Study on Screening and Degradation Effect of Autotoxin-Degrading Bacteria in Muskmelon


Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Isolation, Screening and Identification of Strains
2.2.1. Isolation and Preliminary Screening of Strains
2.2.2. Rescreening of Strains
2.2.3. Identification of Strains
2.3. Determination of the Degradation Effect of Autotoxin-Degrading Bacteria on Phenolic Acids
2.3.1. Preparation of Phenolic Acid Solution
2.3.2. HPLC Conditions
2.3.3. Standard Curve Drawing of Phenolic Acids
2.3.4. Determination of the Degradation Effect of Phenolic Acids by Autotoxin-Degrading Bacteria
2.4. Pot Experiments
2.4.1. Autotoxin-Degrading Bacteria Alleviate the Toxic Effects of Phenolic Acids on Muskmelon
2.4.2. Determination of Degradation Effect of Phenolic Acids by Autotoxin-Degrading Bacteria in the Soil
2.5. Stability Detection of Degradation Ability of Autotoxin-Degrading Bacteria
3. Results
3.1. Isolation and Screening of Autotoxin-Degrading Bacteria
3.2. Identification of Autotoxin-Degrading Bacteria
3.3. Standard Curve of Phenolic Acid Solution
3.4. Degradation of Phenolic Acids by Autotoxin-Degrading Bacteria
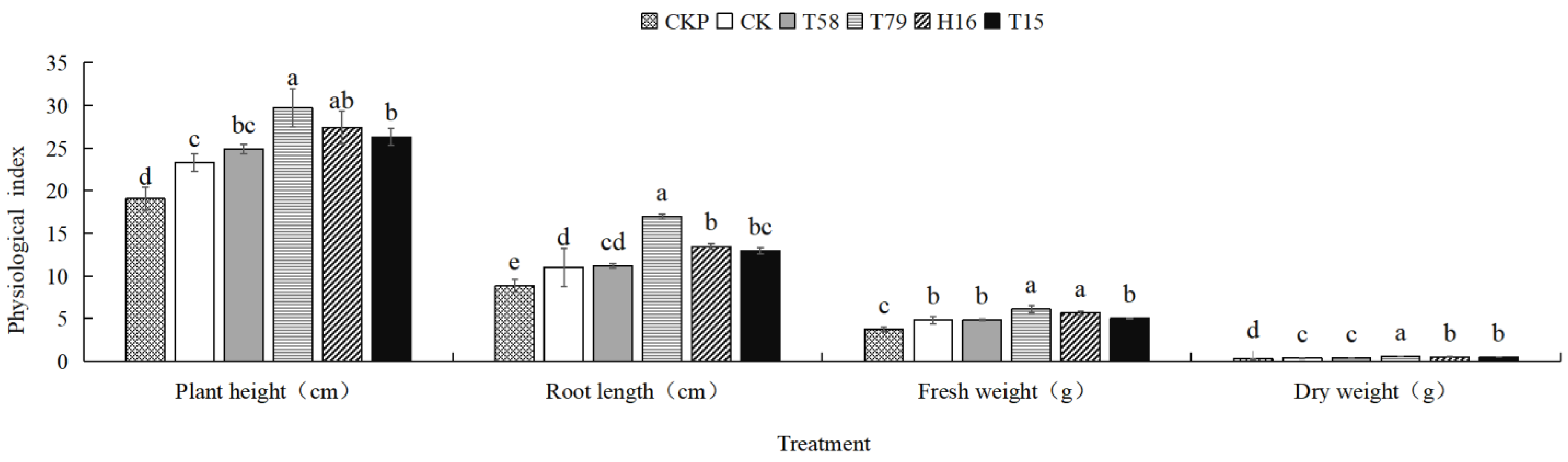
3.5. Autotoxin-Degrading Bacteria Alleviate the Toxic Effects of Phenolic Acids on Muskmelon
3.6. Degradation of Phenolic Acids by Autotoxin-Degrading Bacteria in the Soil
3.7. Stability Detection of Degradation Ability of Autotoxin-Degrading Bacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Tian, H.; Wang, P.; Xiao, Q.; Zhu, S.; Jiang, H. Variations in pH significantly affect cadmium uptake in grafted muskmelon (Cucumis melo L.) plants and drive the diversity of bacterial communities in a seedling substrate. Plant Physiol. Biochem. 2019, 139, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.N.; Gao, Y.; Yang, R.X.; Zhang, Z.R.; Cai, X.T.; Liu, B.; Gao, Z.G.; Liu, X.; Yao, Y. Screening and using allelopathic crops to increase the yield of muskmelon (Cucumis melon L.). Allelopath. J. 2017, 40, 117–131. [Google Scholar] [CrossRef]
- Li, M.; Zhang, L.Y.; Zhang, Y.J.; Zhang, Y.J.; Zhu, J.J.; Ma, H.J. Review on the microbial biodegradation and metabolism of autotoxic phenolic acids. Asian J. Ecotoxicol. 2019, 14, 72–78. [Google Scholar]
- Ding, S.; Zhou, D.P.; Wei, H.W.; Wu, S.H.; Xie, B. Alleviating soil degradation caused by watermelon continuous cropping obstacle: Application of urban waste compost. Chemosphere 2021, 262, 128387. [Google Scholar] [CrossRef]
- Tan, G.; Liu, Y.J.; Peng, S.G.; Yin, H.Q.; Meng, D.L.; Tao, J.M.; Zhou, Z.C. Soil potentials to resist continuous cropping obstacle: Three field cases. Environ. Res. 2021, 200, 111319. [Google Scholar] [CrossRef]
- Chen, P.; Wang, Y.Z.; Liu, Q.Z.; Zhang, Y.T.; Li, W.Y.; Li, H.Q.; Li, W.H. Phase changes of continuous cropping obstacles in strawberry (Fragaria × ananassa duch.) production. Appl. Soil Ecol. 2020, 155, 103626. [Google Scholar] [CrossRef]
- Wu, H.S.; Liu, D.Y.; Ling, N.; Wei, B.; Ying, R.; Shen, Q. Influence of Root Exudates of Watermelon on Fusarium oxysporum f. sp. niveum. Soil Sci. Soc. Am. J. 2009, 73, 1150–1156. [Google Scholar] [CrossRef]
- Dzafic, E.; Pongrac, P.; Likar, M.; Regvar, M.; Vogel-Miku, K. The arbuscular mycorrhizal fungus glomus mosseae alleviates autotoxic effects in maize (Zea mays L.). Eur. J. Soil Biol. 2013, 58, 59–65. [Google Scholar] [CrossRef]
- Zou, L.Y.; Ogweno, J.O.; Yao, S. Autotoxic potential of root exudates and associated phenolics in watermelon. Allelopath. J. 2006, 18, 103–109. [Google Scholar]
- Yang, R.X.; Gao, Z.G.; Liu, X.; Yao, Y.; Cheng, Y.; Huang, J.; Mcdermott, M.I. Effects of phenolic compounds of muskmelon root exudates on growth and pathogenic gene expression of Fusarium oxysporum f. sp. Melonis. Allelopath. J. 2015, 35, 175–186. [Google Scholar]
- Hao, W.Y.; Ren, L.X.; Ran, W.; Shen, Q.R. Allelopathic effects of root exudates from watermelon and rice plants on Fusarium oxysporum f. sp. niveum. Plant Soil 2010, 336, 485–497. [Google Scholar] [CrossRef]
- Jose, C.M.; Torres, L.M.B.; Torres, M.A.M.G.; Shirasuna, R.T. Phytotoxic effects of phenolic acids from Merostachys riedeliana, a native and overabundant Brazilian bamboo. Chemoecology 2016, 26, 235–246. [Google Scholar] [CrossRef]
- Sun, X.; Wang, X.F.; Wei, M.; Wang, F.; Shi, Q.H.; Zhou, B. Screening and identification of cinnamic acid-degrading fungis and the effect of degradation liquid on the cucumber germination. Acta Hortic. Sin. 2014, 41, 765–772. [Google Scholar]
- Zwetsloot, M.J.; Kessler, A.; Bauerle, T.L. Phenolic root exudate and tissue compounds vary widely among temperate forest tree species and have contrasting effects on soil microbial respiration. New Phytol. 2018, 218, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.Q.; Shou, S.Y.; Qian, Y.R.; Zhu, Z.J.; Hu, W.H. Autotoxic potential of cucurbit crops. Plant Soil 2000, 223, 147–151. [Google Scholar] [CrossRef]
- Jin, X.; Wu, F.; Zhou, X. Different toxic effects of ferulic and p-hydroxybenzoic acids on cucumber seedling growth were related to their different influences on rhizosphere microbial composition. Biol. Fertil. Soils 2020, 56, 125–136. [Google Scholar] [CrossRef]
- Asao, T.; Kitazawa, H.; Tomita, K.; Suyama, K.; Yamamoto, H.; Hosoki, T.; Pramanik, M.H.R. Mitigation of cucumber autotoxicity in hydroponic culture using microbial strain. Sci. Hortic. 2004, 9, 207–214. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Qiao, H.Y.; Gao, H.C.; Li, S. Effect of melon continuous cropping on Rhizosphere soil microorganisms and enzyme activities. North. Hortic. 2015, 19, 158–161. [Google Scholar]
- Zhou, X.; Yu, G.B.; Wu, F.Z. Responses of soil microbial communities in the rhizosphere of cucumber (Cucumis sativus L.) to exogenously applied p-hydroxybenzoic acid. J. Chem. Ecol. 2012, 38, 975–983. [Google Scholar] [CrossRef]
- Liu, Y.T.; Yang, I.C.; Lin, N.C. Evaluation of biocontrol potential for Fusarium yellows of celery by antagonistic and gallic acid-degrading bacteria. Biol. Control. 2020, 146, 104268. [Google Scholar] [CrossRef]
- Hou, H.; Dong, K.; Yang, Z.X.; Dong, Y.; Tang, L.; Zheng, Y. Advance in Mechanism of Continuous Cropping Obstacle. Soils 2016, 48, 1068–1076. [Google Scholar]
- Chen, L.; Dong, K.; Yang, Z.X.; Dong, Y.; Tang, L.; Zheng, Y. Allelopathy Autotoxcity Effect of Successive Cropping Obstacle and Its Alleviate Mechanism by Intercropping. Chin. Agric. Sci. Bull. 2017, 33, 91–98. [Google Scholar]
- Piattoni, F.; Roberti, R.; Servidio, G. Studies on the potential role of root exudates in the interaction between muskmelon roots and Fusarium oxysporum f. sp. melonis. J. Plant Dis. Prot. 2014, 121, 64–70. [Google Scholar] [CrossRef]
- Ye, S.F.; Zhou, Y.H.; Sun, Y.; Zou, L.Y. Cinnamic acid causes oxidative stress in cucumber roots, and promotes incidence of Fusarium wilt. Environ. Exp. Bot. 2006, 56, 255–262. [Google Scholar] [CrossRef]
- Ma, H.; Yang, R.X.; Gai, X.T.; Liu, C.; Zhang, Z.R.; Tang, S.S.; Gao, Z.G. Alleviating of Muskmelon Soil of Continuous Cropping Obstacles by Different Soil Treatment Agent. Agrochemicals 2018, 57, 764–767. [Google Scholar]
- Yang, R.X.; Gao, Z.G.; Yao, Y.; Liu, X.; Sun, S.Q.; Wang, Y. Allelopathic effects of phenolic compounds of melon root exudates on Fusarium oxysporum f. sp. Melonis. Chin. J. Appl. Ecol. 2014, 25, 2355–2360. [Google Scholar]
- Yang, R.X.; Gao, Z.G.; Liu, X.; Yao, Y.; Cheng, Y. Root exudates from muskmelon (Cucumis melon. L)induce autotoxicity and promote growth of Fusarium oxysporum f. sp. melonis. Allelopath. J. 2014, 33, 175–188. [Google Scholar]
- Gao, Q. Physiological and Biochemical Mechanisms of Muskmelon Allelopathy and Identification of Main Allelochemicals. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2014. [Google Scholar]
- Zhang, L.; Yang, R.X.; Wang, Y.; Yao, Y.; Yan, H.; Gao, Z.G. Identification and degradation of phenolic acids in rhizosphere soil of replanted muskmelon. North. Hortic. 2017, 41, 18–23. [Google Scholar]
- Wang, S.N.; Yang, R.X.; Liu, X. Overcoming the autotoxicity of muskmelon by Trichoderma spp. Allelopath. J. 2016, 39, 29–42. [Google Scholar]
- Liu, J.G.; Wang, X.X.; Zhang, T.L.; Li, X. Assessment of active bacteria metabolizing phenolic acids in the peanut (Arachis hypogaea L.) rhizosphere. Microbiol. Res. 2017, 205, 118–124. [Google Scholar] [CrossRef]
- Hao, H.; Zhu, W.; Noor, I.; Liu, J.W.; Li, G.H. Pseudomonas putida WH-B3 degrades benzoic acid and alleviates its autotoxicity to peach (Prunus persica L. batsch) seedlings grown in replanted soil. Sci. Hortic. 2019, 255, 183–192. [Google Scholar]
- Gauri, S.S.; Mandal, S.M.; Dey, S.; Pati, B.R. Biotransformation of p-coumaric acid and 2,4-dichlorophenoxy acetic acid by Azotobacter sp. strain SSB81. Bioresour. Technol. 2012, 126, 350–353. [Google Scholar] [CrossRef]
- Chen, S.L.; Zhou, B.L.; Lin, S.S.; Li, X.; Ye, X.L. Effects of cinnamic acid and vanillin on grafted eggplant root growth and physiological characteristics. Chin. J. Appl. Ecol. 2010, 21, 1446–1452. [Google Scholar]
- Kataoka, M.; Ueda, K.; Kudo, T. Application of the variable region in 16S rDNA to create an index for rapid species identification in the genus Streptomyces. FEMS Microbiol. Lett. 1997, 151, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Ausubel, F.M. Short Protocols in Molecular Biology; Yan, Z.Y.; Wang, H.L., Translators; Science Press: Beijing, China, 1998; pp. 39–40. [Google Scholar]
- Dalton, B.R.; Weed, S.B.; Blun, U. Plant Phenolic Acids in Soils: A Comparison of Extraction Procedures. Soil Sci. Soc. Am. J. 1987, 51, 1515–1521. [Google Scholar] [CrossRef]
- He, Z.G.; Lou, C.R.; Wang, X.J.; Dong, H.; Zhao, Y. Screening and degradation effect of tomato autotoxic substances degrading bacteria. Jiangsu Agric. Sci. 2017, 45, 114–116. [Google Scholar]
- Ma, Y.Y.; Chen, X.X.; Li, M.; Wang, J.; Zhang, X.; Yang, G.P. Degradation of cinnamic acid by Exiguobacterium sp. strain. Microbiol. China 2017, 44, 2079–2088. [Google Scholar]
- Qi, G.Z.; Mao, Z.Q.; Hu, X.N.; Du, B.H.; Ge, K.; Yang, Q.Q.; Ding, Y.Q. Isolation, identification and degradation characteristics of apple rhizosphere autotoxicitic compounds-degrading bacteria. Microbiol. China 2016, 43, 330–342. [Google Scholar]
- Wang, J.; Wang, B.B.; Shang, F.J.; Su, L.Q.; Zhao, S.G.; Hong, S.; Zhao, Q.Y. Screening, identification and antimicrobial activity of microbial strains degrading autotoxic phenolic acids in the rhizosphere of vanilla. J. Trop. Biol. 2022, 13, 595–604. [Google Scholar]
- Zhao, D.Y.; Li, Y.; Ding, W.L. Isolation and characteristics of Panax ginseng autotoxin-degrading bacterial strains. China J. Mater. Med. 2013, 38, 1703–1706. [Google Scholar]
- Liu, Y.L.; Zheng, S.H.; Ye, L.X.; Liao, G.J.; Hu, C.H. Isolation and degradation characteristics of ferulic acid-degrading fungi in rhizosphere soil of Chinese goldthread. J. Southwest Univ. (Nat. Sci. Ed.) 2019, 41, 46–50. [Google Scholar]
- Li, H.W.; Zhao, X.Y.; Li, P.K. Research on Biodegradation Effect to Phenolic Acid in Even Cook Obstacles. Chin. Agric. Sci. Bull. 2011, 27, 168–173. [Google Scholar]
- Xie, L.J.; Yin, B.Z.; Gao, F.; Qi, Y.Z. Screening of the degradation bacterium to strawberry root auto-toxic chemical and the effect of its degradation. J. Agric. Univ. Hebei 2009, 32, 76–78. [Google Scholar]
- Yu, G.H.; Xie, Y.H.; Chen, Y.H.; Chen, Y.F.; Cheng, P. Mitigating the repress of cinnamic acid to cucumber growth by microbial strain. Acta Microbiol. Sin. 2006, 46, 934–938. [Google Scholar]
- Wang, X.H.; Xue, Q.H. Screening of ferulic acid degrading actinomycetes and their degradation and inhibiting effect. J. Northwest AF Univ. (Nat. Sci. Ed.) 2011, 39, 153–158. [Google Scholar]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Yu, C.Y.; Gao, J.S. Isolation and Identification of a Phenolic Acids-Degrading Bacterium from Peanut Soil. J. Anhui Agric. Sci. 2020, 48, 93–95. [Google Scholar]
- Chen, S.; Guo, L.; Bai, J. Biodegradation of p-hydroxybenzoic acid in soil by Pseudomonas putida CSY-P1 isolated from cucumber rhizospheresoil. Plant Soil 2015, 389, 197–210. [Google Scholar] [CrossRef]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov, A.Y.; Tikhonovich, I.A.; Kluge, C.; Preisfeld, A.; et al. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2001, 47, 642–652. [Google Scholar] [CrossRef]
- Paungfoo, L.C.; Lonhienne, T.G.A.; Yeoh, Y.K. Crosstalk between sugarcane and a plant-growth promoting Burkholderia species. Sci. Rep. 2016, 6, 37389. [Google Scholar] [CrossRef]
- Mullins, A.J.; Murray, J.A.H.; Bull, M.J. Genome mining identifies cepacin as a plant-protective metabolite of the biopesticidal bacterium Burkholderia ambifaria. Nat. Microbiol. 2019, 4, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lowman, S.; Hou, G.C.; Nowak, J.; Flinn, B.; Mei, C.S. Growth promotion and colonization of switchgrass (Panicum virgatum) cv. Alamo by bacterial endophyte Burkholderia phytofirmans strain PsJN. Biotechnol. Biofuels 2012, 5, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Paungfoo, L.C.; Lonhienne, T.G.A.; Yeoh, Y.K.; Webb, R.I.; Lakshmanan, P.; Chan, C.X.; Lim, P.E.; Ragan, M.A.; Schmidt, S.; Hugenholtz, P. A new species of Burkholderia isolated from sugarcane roots promotes plant growth. Microb. Biotechnol. 2014, 7, 142–154. [Google Scholar] [CrossRef]
- Sun, Z.X.; Meng, X.J.; Long, X.Y.; Qiu, M.S.; Mao, G.Q.; Zhou, Y. Effects of Burkholderia sp. YZU-S230 on the control and growth promotion of watermelon Fusarium wilt. J. Yangtze Univ. (Nat. Sci. Ed.) 2021, 18, 82–88. [Google Scholar]
- Bernabeu, P.R.; Pistorio, M.; Torres-Tejerizo, G.; Santos, P.E.F.L.; Galar, M.L.; Boiardi, J.L.; Luna, M.F. Colonization and plant growth-promotion of tomato by Burkholderia tropica. Sci. Hortic. 2015, 191, 113–120. [Google Scholar] [CrossRef]
- de Oliveira-Longatti, S.M.; de Sousa, P.M.; Marra, L.M.; Ferreira, P.A.A.; de Souza, M.F.M. Burkholderia fungorum promotes common bean growth in a dystrophic oxisol. Ann. Microbiol. 2015, 65, 1825–1832. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Phenolic Acid | Regression Equation | Correlation | Detection Wavelength | Retention Time (min) |
|---|---|---|---|---|---|
| 1 | Ferulic acid | y = 121.34 − 85.90 | 0.99990 | 310 | 12.007 |
| 2 | Caffeic acid | y = 104.42x − 105.53 | 0.99919 | 323 | 7.773 |
| 3 | Cinnamic acid | y = 173.84x + 73.20 | 0.99975 | 270 | 19.772 |
| 4 | Salicylic acid | y = 36.60x − 2.39 | 0.99990 | 300 | 16.267 |
| 5 | Vanillin | y = 89.79x − 0.75 | 0.99999 | 275 | 9.633 |
| 6 | Gallic acid | y = 67.86x + 2.14 | 1.00000 | 273 | 3.139 |
| 7 | Protocatechuic acid | y = 74.96x + 11.96 | 0.99999 | 260 | 4.658 |
| 8 | Vanillic acid | y = 76.64x + 3.53 | 0.99997 | 260 | 7.759 |
| 9 | p-Coumaric acid | y = 247.96x + 37.29 | 0.99999 | 310 | 11.432 |
| 10 | Sinapic acid | y = 79.41x − 5.13 | 0.99998 | 320 | 12.863 |
| Phenolic Acid | Concentration of Phenolic Acid (mg/L) | Degradation Rate (%) | |||||
|---|---|---|---|---|---|---|---|
| 0 day | 7 days | 14 days | 21 days | 7 days | 14 days | 21 days | |
| Ferulic acid | 25.06 ± 0.18 d | 21.61 ± 0.33 b | 16.57 ± 0.45 d | 14.59 ± 0.30 d | 13.75 ± 0.70 cd | 33.88 ± 1.31 cd | 41.77 ± 0.77 e |
| Caffeic acid | 25.16 ± 0.33 d | 23.75 ± 0.17 a | 18.16 ± 0.82 c | 15.62 ± 0.11 c | 5.59 ± 1.88 f | 27.80 ± 4.15 e | 37.92 ± 0.48 f |
| Cinnamic acid | 25.99 ± 0.54 bc | 20.49 ± 1.14 c | 19.58 ± 0.28 b | 2.85 ± 0.11 f | 21.09 ± 5.75 ab | 24.67 ± 1.63 e | 89.04 ± 0.39 c |
| Salicylic acid | 26.09 ± 0.07 bc | 20.33 ± 0.84 c | 15.14 ± 0.13 f | 0.50 ± 0.06 g | 22.09 ± 3.03 ab | 41.98 ± 0.34 b | 98.09 ± 0.23 b |
| Vanillin | 26.88 ± 0.04 a | 21.63 ± 0.37 b | 18.52 ± 0.17 c | 16.91 ± 0.10 b | 19.54 ± 1.44 ab | 31.09 ± 0.70 d | 37.08 ± 0.44 g |
| Gallic acid | 25.80 ± 0.69 c | 20.39 ± 0.57 c | 16.59 ± 0.29 d | 0.00 ± 0.00 h | 20.87 ± 4.37 ab | 35.62 ± 2.85 c | 100 ± 0.00 a |
| Protocatechuic acid | 26.48 ± 0.29 ab | 23.45 ± 0.12 a | 19.73 ± 0.11 ab | 17.35 ± 0.05 a | 11.40 ± 1.40 de | 25.46 ± 0.78 e | 34.45 ± 0.88 h |
| Vanillic acid | 25.13 ± 0.13 d | 23.48 ± 0.54 a | 20.25 ± 0.05 a | 17.48 ± 0.04 a | 6.58 ± 2.62 ef | 19.44 ± 0.60 f | 30.44 ± 0.46 i |
| p-Coumaric acid | 26.76 ± 0.08 a | 21.97 ± 0.70 b | 15.83 ± 0.20 e | 6.60 ± 0.12 e | 17.88 ± 2.71 bc | 40.86 ± 0.84 b | 75.35 ± 0.47 d |
| Sinapic acid | 25.87 ± 0.13 c | 19.46 ± 0.21 c | 9.37 ± 0.07 g | 0.00 ± 0.00 h | 24.77 ± 0.46 a | 63.77 ± 0.28 a | 100 ± 0.00 a |
| Phenolic Acid | Concentration of Phenolic Acid (mg/L) | Degradation Rate (%) | |||||
|---|---|---|---|---|---|---|---|
| 0 day | 7 days | 14 days | 21 days | 7 days | 14 days | 21 days | |
| Ferulic acid | 25.10 ± 0.11 d | 1.09 ± 0.09 h | 0.64 ± 0.04 i | 0.00 ± 0.00 g | 95.64 ± 0.32 a | 97.44 ± 0.19 a | 100.00 ± 0.00 a |
| Caffeic acid | 25.12 ± 0.07 d | 23.18 ± 0.10 b | 18.37 ± 0.06 b | 15.87 ± 0.06 b | 7.75 ± 0.53 gh | 26.90 ± 0.36 i | 36.84 ± 0.21 f |
| Cinnamic acid | 26.59 ± 0.18 b | 19.55 ± 0.13 d | 6.88 ± 0.15 f | 2.03 ± 0.11 e | 26.47 ± 0.03 e | 74.13 ± 0.72 d | 92.38 ± 0.37 c |
| Salicylic acid | 26.10 ± 0.09 c | 17.69 ± 0.28 e | 8.92 ± 0.09 d | 0.45 ± 0.04 f | 32.24 ± 0.84 d | 65.81 ± 0.46 f | 98.27 ± 0.13 b |
| Vanillin | 26.85 ± 0.11 a | 22.7 ± 0.21 c | 17.28 ± 0.27 c | 14.44 ± 0.31 d | 15.46 ± 0.43 f | 35.66 ± 0.74 g | 46.23 ± 0.92 d |
| Gallic acid | 25.98 ± 0.12 cd | 19.42 ± 0.10 d | 7.39 ± 0.09 e | 0.40 ± 0.03 f | 25.23 ± 0.70 e | 71.55 ± 0.46 e | 98.47 ± 0.09 b |
| Protocatechuic acid | 26.43 ± 0.11 b | 24.62 ± 0.51 a | 18.38 ± 0.15 b | 15.04 ± 0.07 c | 6.85 ± 2.28 h | 30.47 ± 0.27 h | 43.08 ± 0.03 e |
| Vanillic acid | 25.04 ± 0.14 d | 22.80 ± 0.05 c | 20.38 ± 0.07 a | 16.46 ± 0.01 a | 8.95 ± 0.47 g | 18.62 ± 0.60 j | 34.29 ± 0.35 g |
| p-Coumaric acid | 26.84 ± 0.03 a | 13.29 ± 0.02 g | 6.28 ± 0.03 g | 0.00 ± 0.00 g | 50.50 ± 0.05 b | 76.62 ± 0.10 c | 100.00 ± 0.00 a |
| Sinapic acid | 25.87 ± 0.08 d | 14.06 ± 0.06 f | 3.50 ± 0.02 h | 0.00 ± 0.00 g | 45.63 ± 0.25 c | 86.46 ± 0.05 b | 100.00 ± 0.00 a |
| Phenolic Acid | Concentration of Phenolic Acid (mg/L) | Degradation Rate (%) | |||||
|---|---|---|---|---|---|---|---|
| 0 day | 7 days | 14 days | 21 days | 7 days | 14 days | 21 days | |
| Ferulic acid | 25.08 ± 0.04 f | 1.04 ± 0.04 h | 0.63 ± 0.01 j | 0.00 ± 0.00 i | 95.84 ± 0.13 a | 97.47 ± 0.05 a | 100.00 ± 0.00 a |
| Caffeic acid | 25.05 ± 0.06 f | 23.75 ± 0.09 a | 17.73 ± 0.09 b | 14.08 ± 0.07 b | 5.19 ± 0.17 j | 29.22 ± 0.20 i | 43.80 ± 0.13 h |
| Cinnamic acid | 26.56 ± 0.07 b | 19.62 ± 0.03 d | 10.98 ± 0.05 e | 8.06 ± 0.06 e | 26.13 ± 0.28 f | 58.65 ± 0.28 f | 69.63 ± 0.31 e |
| Salicylic acid | 26.08 ± 0.03 d | 18.37 ± 0.02 e | 8.23 ± 0.02 h | 1.06 ± 0.03 g | 29.57 ± 0.10 d | 68.43 ± 0.09 c | 95.94 ± 0.12 c |
| Vanillin | 26.84 ± 0.05 a | 22.8 ± 0.01 c | 15.16 ± 0.03 d | 12.65 ± 0.04 d | 15.05 ± 0.17 g | 43.53 ± 0.11 g | 52.85 ± 0.05 f |
| Gallic acid | 25.94 ± 0.07 e | 18.35 ± 0.06 e | 8.79 ± 0.03 g | 0.21 ± 0.01 h | 29.23 ± 0.38 e | 66.11 ± 0.19 d | 99.20 ± 0.02 b |
| Protocatechuic acid | 26.47 ± 0.03 c | 22.84 ± 0.02 c | 15.62 ± 0.05 c | 12.81 ± 0.03 c | 13.69 ± 0.01 h | 40.98 ± 0.15 h | 51.61 ± 0.09 g |
| Vanillic acid | 25.04 ± 0.04 f | 23.64 ± 0.03 b | 19.52 ± 0.05 a | 14.46 ± 0.05 a | 5.61 ± 0.22 i | 22.08 ± 0.06 j | 42.26 ± 0.29 i |
| p-Coumaric acid | 26.83 ± 0.08 a | 4.11 ± 0.01 g | 1.68 ± 0.03 i | 0.00 ± 0.00 i | 84.69 ± 0.05 b | 93.72 ± 0.11 b | 100.00 ± 0.00 a |
| Sinapic acid | 25.87 ± 0.03 e | 16.23 ± 0.02 f | 9.57 ± 0.04 f | 7.77 ± 0.02 f | 37.23 ± 0.13 c | 63.01 ± 0.19 e | 69.97 ± 0.02 d |
| Phenolic Acid | Concentration of Phenolic Acid (mg/L) | Degradation Rate (%) | |||||
|---|---|---|---|---|---|---|---|
| 0 day | 7 days | 14 days | 21 days | 7 days | 14 days | 21 days | |
| Ferulic acid | 25.08 ± 0.04 g | 20.64 ± 0.09 c | 15.85 ± 0.10 d | 8.41 ± 0.02 d | 17.74 ± 0.23 f | 36.79 ± 0.43 h | 66.48 ± 0.03 g |
| Caffeic acid | 25.07 ± 0.02 g | 22.23 ± 0.02 b | 18.75 ± 0.04 a | 15.32 ± 0.01 a | 11.30 ± 0.02 g | 25.19 ± 0.08 j | 38.90 ± 0.05 j |
| Cinnamic acid | 26.54 ± 0.04 b | 18.46 ± 0.05 e | 11.12 ± 0.05 e | 2.24 ± 0.03 g | 30.45 ± 0.10 e | 58.11 ± 0.25 f | 91.55 ± 0.11 d |
| Salicylic acid | 26.12 ± 0.03 d | 16.38 ± 0.05 g | 9.08 ± 0.06 g | 0.95 ± 0.02 h | 37.30 ± 0.17 c | 65.26 ± 0.19 c | 96.38 ± 0.06 c |
| Vanillin | 26.85 ± 0.07 a | 13.53 ± 0.09 i | 7.54 ± 0.08 i | 0.00 ± 0.00 j | 49.62 ± 0.32 a | 71.93 ± 0.26 a | 100.00 ± 0.00 a |
| Gallic acid | 25.92 ± 0.06 e | 14.11 ± 0.11 h | 7.70 ± 0.03 h | 0.26 ± 0.01 i | 45.58 ± 0.40 b | 70.29 ± 0.16 b | 99.00 ± 0.05 b |
| Protocatechuic acid | 26.44 ± 0.06 c | 23.68 ± 0.04 a | 16.09 ± 0.08 c | 11.97 ± 0.06 c | 10.41 ± 0.22 h | 39.12 ± 0.41 g | 54.70 ± 0.14 h |
| Vanillic acid | 25.06 ± 0.04 g | 22.26 ± 0.04 b | 18.24 ± 0.04 b | 12.94 ± 0.03 b | 11.19 ± 0.11 g | 27.21 ± 0.16 i | 48.37 ± 0.11 i |
| p-Coumaric acid | 26.81 ± 0.03 a | 18.58 ± 0.03 d | 10.70 ± 0.04 f | 5.19 ± 0.03 f | 30.69 ± 0.13 e | 60.09 ± 0.16 e | 80.65 ± 0.10 e |
| Sinapic acid | 25.82 ± 0.07 f | 17.38 ± 0.03 f | 9.08 ± 0.03 g | 7.75 ± 0.03 e | 32.67 ± 0.27 d | 64.81 ± 0.20 d | 69.99 ± 0.06 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, S.; Zhang, Z.; Liu, X.; Gao, Z. Study on Screening and Degradation Effect of Autotoxin-Degrading Bacteria in Muskmelon. Agronomy 2023, 13, 1334. https://doi.org/10.3390/agronomy13051334
Tang S, Zhang Z, Liu X, Gao Z. Study on Screening and Degradation Effect of Autotoxin-Degrading Bacteria in Muskmelon. Agronomy. 2023; 13(5):1334. https://doi.org/10.3390/agronomy13051334
Chicago/Turabian StyleTang, Shuangshuang, Zhaoran Zhang, Xiaodi Liu, and Zenggui Gao. 2023. "Study on Screening and Degradation Effect of Autotoxin-Degrading Bacteria in Muskmelon" Agronomy 13, no. 5: 1334. https://doi.org/10.3390/agronomy13051334
APA StyleTang, S., Zhang, Z., Liu, X., & Gao, Z. (2023). Study on Screening and Degradation Effect of Autotoxin-Degrading Bacteria in Muskmelon. Agronomy, 13(5), 1334. https://doi.org/10.3390/agronomy13051334