



Development of Lycium barbarum–Forage Intercropping Patterns

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Experimental Site Description
2.2. Experimental Design: Wolfberry–Forage Plant Interactions
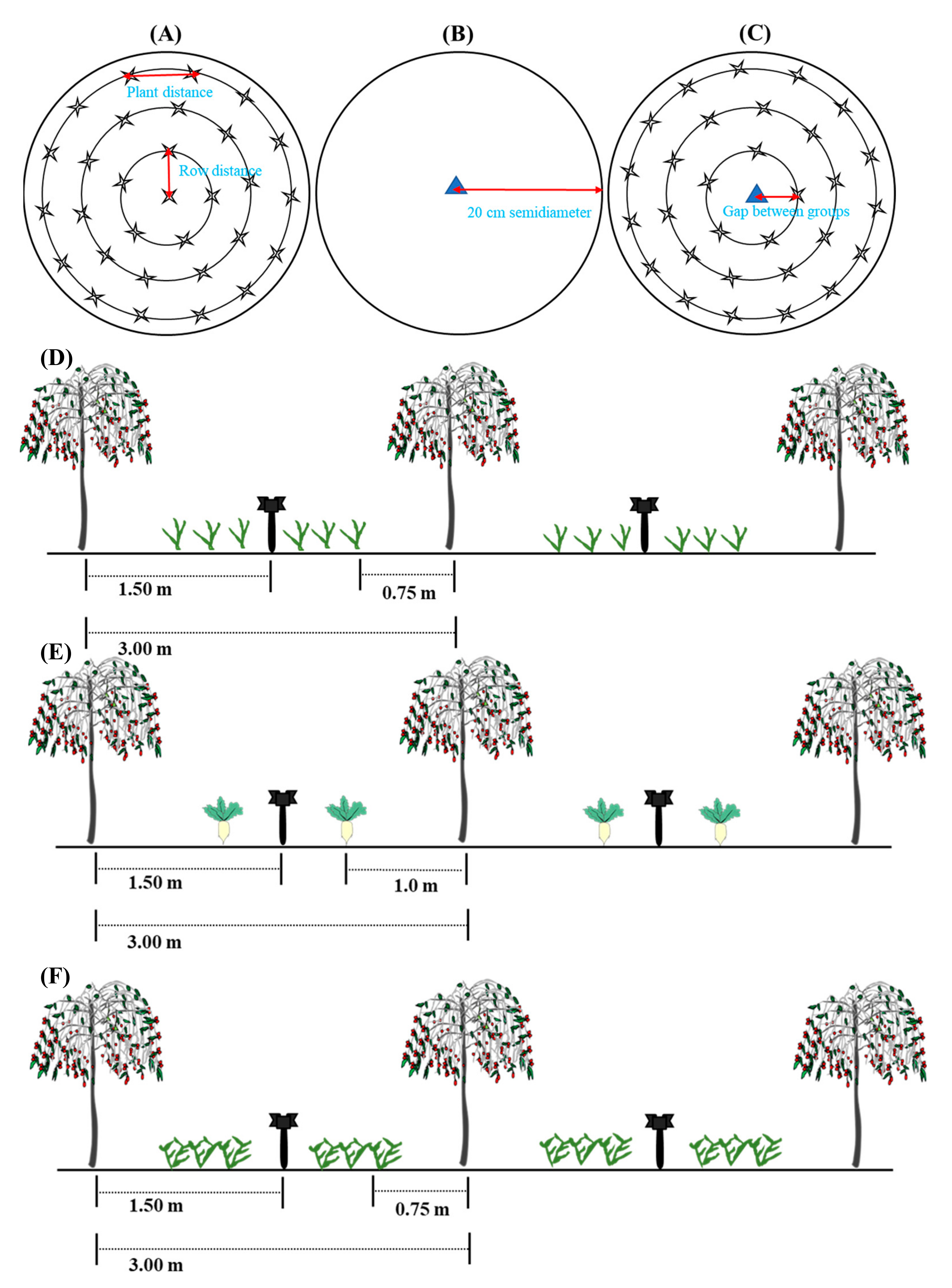
2.2.1. Experimental Design and Management
2.2.2. Experimental Sample Collection
2.3. Relative Index Measurements of Wolfberry and Forage
2.3.1. Determination of Above-Ground Biomass and Yield of Forage
2.3.2. Determination of Added Biomass and Yield of Wolfberry
2.4. Evaluation of Wolfberry–Forage Intercropping Patterns
2.4.1. Contribution of Wolfberry and Forage to Productivity Advantage
2.4.2. Interspecific Competitiveness Evaluation of Wolfberry–Forage Intercropping
2.5. Statistical Analyses
3. Results and Analyses
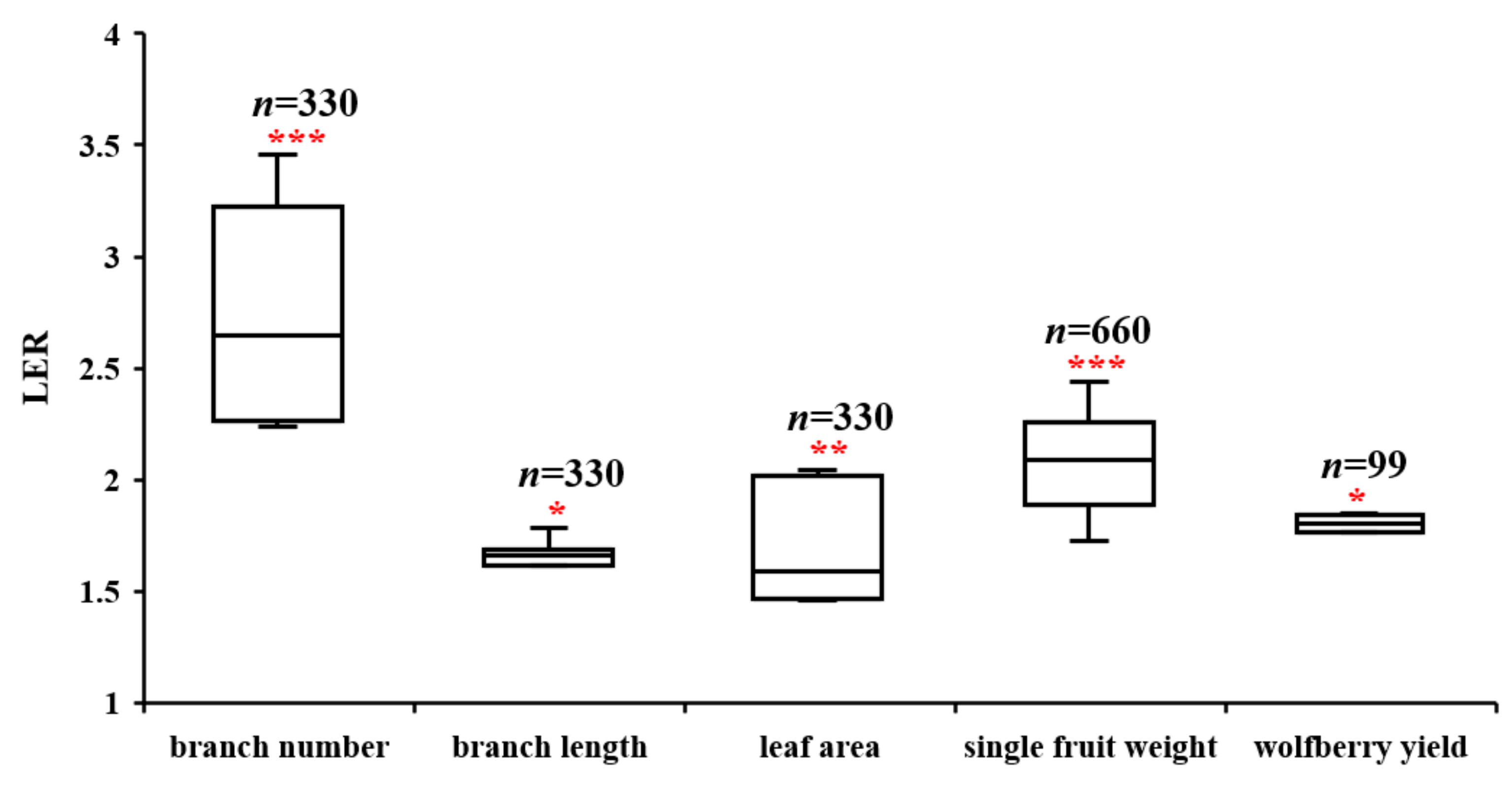
3.1. Growth and Biological Yield Advantage of Intercropping Patterns
3.2. Yield and Intercropping Advantage of Different Intercropping Patterns
3.2.1. Yield and Intercropping Advantage of Different Intercropping Patterns in Greenhouse
3.2.2. Yield and Intercropping Advantage of Different Intercropping Patterns in the Field
3.2.3. Evaluation of Interspecific Competitiveness
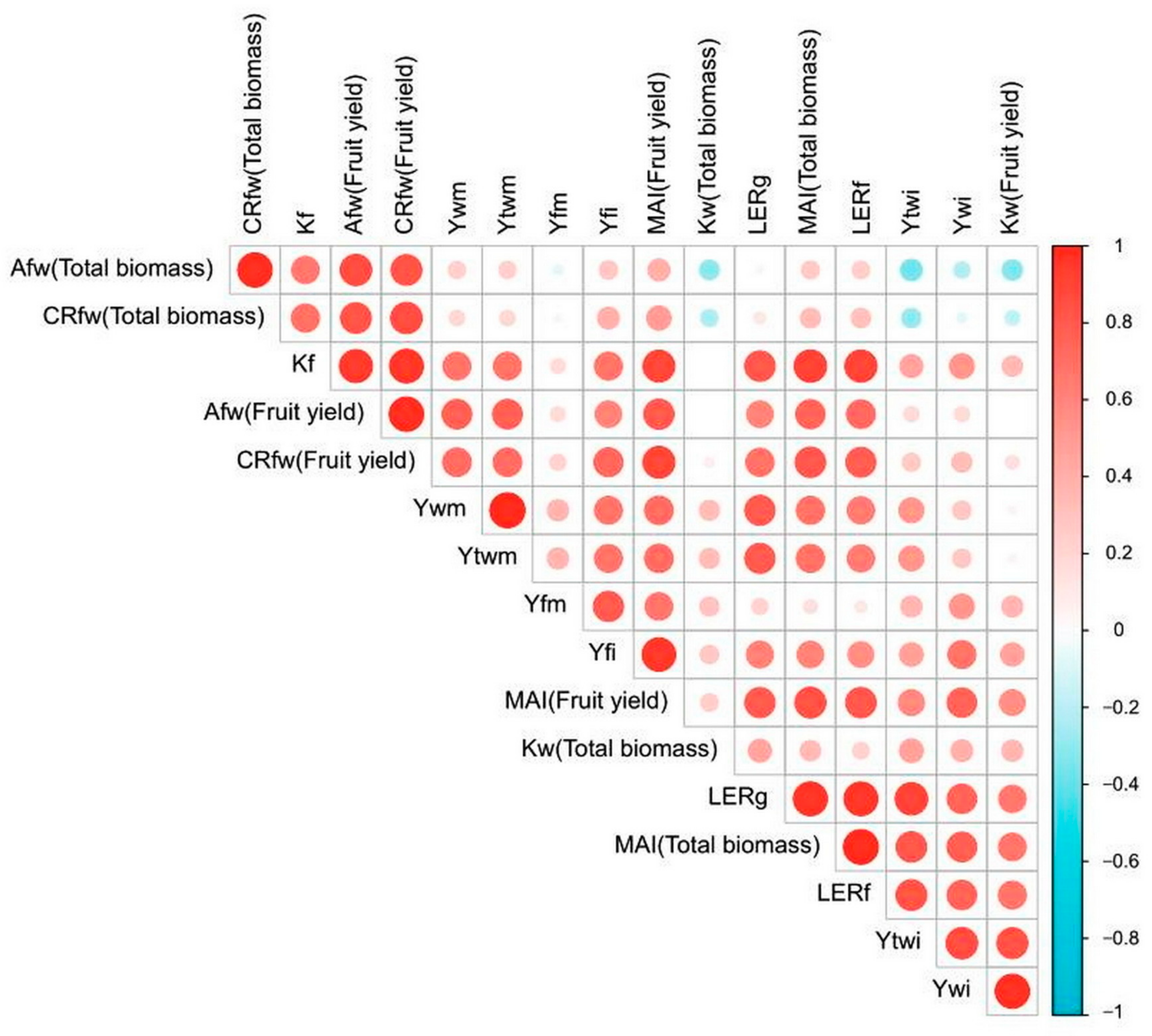
3.2.4. Descriptive and Correlational Analyses of Growth and Yield Factors
4. Discussion
4.1. Yield Differences under Different Wolfberry–Forage Intercropping Patterns
4.2. Competitiveness Differences under Different Wolfberry–Forage Intercropping Patterns
4.3. Production Efficiency Differences under Different Wolfberry–Forage Intercropping Patterns
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Amagase, H.; Farnsworth, N.R. A review of botanical characteristics, phytochemistry, clinical relevance in efficasscy and safety of Lycium barbarum fruit (Goji). Food Res. Int. 2011, 44, 1702–1717. [Google Scholar] [CrossRef]
- Wen, S.P.; Wang, C.; Zhang, Z.H. Lycium Barbarum Industry Dvevlopment and Technological Innovation Trends in the Whole Industry Chain in Ningxia. Agric. Outlook 2018, 14, 53–58+64. [Google Scholar]
- Zhu, L.; He, J.; Tian, Y.; Li, X.; Li, Y.; Wang, F.; Qin, K.; Wang, J. Intercropping Wolfberry with Gramineae plants improves productivity and soil quality. Sci. Hortic. 2022, 292, 110632. [Google Scholar] [CrossRef]
- Rajib, M.A.; Ahiablame, L.; Paul, M. Modeling the effects of future land use change on water quality under multiple scenarios: A case study of low-input agriculture with hay/pasture production. Sustain. Water Qual. Ecol. 2016, 8, 50–66. [Google Scholar] [CrossRef]
- Carberry, P.S.; Liang, W.L.; Twomlow, S.; Holzworth, D.P.; Dimes, J.P.; Mcclelland, T.; Huth, N.I.; Chen, F.; Hochman, Z.; Keating, B.A. Scope for improved eco-efficiency varies among diverse cropping systems. Proc. Natl. Acad. Sci. USA 2013, 110, 8381–8386. [Google Scholar] [CrossRef]
- Padgham, J. Agricultural Development under a Changing Climate: Opportunities and Challenges for Adaptation; World Bank: Washington, DC, USA, 2012. [Google Scholar]
- Martin-Guay, M.O.; Paquette, A.; Dupras, J.; Rivest, D. The new Green Revolution: Sustainable intensification of agriculture by intercropping. Sci. Total Environ. 2017, 615, 767–772. [Google Scholar] [CrossRef]
- Link, C.M.; Thevathasan, N.V.; Gordon, A.M.; Isaac, M.E. Determining tree water acquisition zones with stable isotopes in a temperate tree-based intercropping system. Agrofor. Syst. 2015, 89, 611–620. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, P.A.; Verma, D.K.; Mir, N.H.; Sharma, A.; Wani, S.A. Forage Production and Orchard Floor Management through Grass/Legume Intercropping in Apple Based Agroforestry Systems; AkiNik Publications: New Delhi, India, 2018. [Google Scholar]
- Köhl, J.; Wenneker, M.; Haas, B.H.D.; Anbergen, R.H.N.; Plas, C.H.; Kastelein, P. Inoculum Sources of the Post-Harvest Pathogens Neofabraea spp. and Cadophora spp. in Dutch Apple and Pear Orchards; Wageningen University: Wageningen, The Netherlands, 2015. [Google Scholar]
- Forey, O.; Temani, F.; Wery, J.; Jourdan, C.; Metay, A. Effect of combined deficit irrigation and grass competition at plantation on peach tree root distribution. Eur. J. Agron. 2017, 91, 16–24. [Google Scholar] [CrossRef]
- Liu, X.; Rahman, T.; Song, C.; Yang, F.; Yang, W. Relationships among light distribution, radiation use efficiency and land equivalent ratio in maize-soybean strip intercropping. Field Crops Res. 2018, 224, 91–101. [Google Scholar] [CrossRef]
- Hu, F.; Tan, Y.; Yu, A.; Zhao, C.; Fan, Z.; Yin, W.; Chai, Q.; Coulter, J.A.; Cao, W. Optimizing the split of N fertilizer application over time increases grain yield of maize-pea intercropping in arid areas. Eur. J. Agron. 2020, 119, 126117. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, P.; Zhao, X.; Wang, Z. Growth, yield, and nitrogen use in the wheat/maize intercropping system in an arid region of northwestern China. Field Crops Res. 2014, 167, 19–30. [Google Scholar] [CrossRef]
- Zhang, R.; Huang, Q.; Yan, T.; Yang, J.; Li, M. Effects of intercropping mulch on the content and composition of soil dissolved organic matter in apple orchard on the loess plateau. J. Environ. Manag. 2019, 250, 109531. [Google Scholar] [CrossRef]
- Borden, K.A.; Isaac, M.E.; Thevathasan, N.V.; Gordon, A.M.; Thomas, S.C. Estimating coarse root biomass with ground penetrating radar in a tree-based intercropping system. Agrofor. Syst. 2014, 88, 657–669. [Google Scholar] [CrossRef]
- Bouttier, L.; Paquette, A.; Messier, C.; Rivest, D.; Olivier, A.; Cogliastro, A. Vertical root separation and light interception in a temperate tree-based intercropping system of Eastern Canada. Agrofor. Syst. 2014, 88, 693–706. [Google Scholar] [CrossRef]
- Evers, A.K.; Bambrick, A.; Lacombe, S.; Dougherty, M.C.; Bradley, R.L. Potential Greenhouse Gas Mitigation through Temperate Tree-Based Intercropping Systems. Open Agric. J. 2010, 4, 49–57. [Google Scholar] [CrossRef]
- Bakshi, M.; Singh, M.P.; Wadhwa, M.; Singh, B. Evaluation of Forest Grasses as Livestock Feed. Livest. Res. Rural. Dev. 2005, 17, 1–15. [Google Scholar]
- Knörzer, H.; Graeff-Hönninger, S.; Guo, B.; Pu, W.; Claupein, W. The Rediscovery of Intercropping in China: A Traditional Cropping System for Future Chinese Agriculture—A Review; Springer: Amsterdam, The Netherlands, 2009. [Google Scholar] [CrossRef]
- Chen, Q.H.; Wang, C.L. Advantage of Ningxia Wolfberry Industry and Strategy of Improving Its Export Competitive Power. Res. Agric. Mod. 2008, 29, 151–154. [Google Scholar]
- Zhang, Z.; Wen, S.; Wang, J. Current situation, existing problems countermeasures in development of top quality chinese wolfberry products in Ningxia. J. Agric. Sci. 2014, 35, 46–50. [Google Scholar]
- Li, J.Y.; Xu, B.; Yang, X.C.; Jin, Y.X.; Gao, T.; Yu, H.D.; Ma, H.L.; Qin, Z.H.; Zhao, L.N. Temporal and spatial variations of grassland desertification in Lingwu and Yanchi of Ningxia, China. In Proceedings of the Second International Conference on Agro-geoinformatics IEEE, Fairfax, VA, USA, 12–16 August 2013. [Google Scholar]
- Wit, C. On Competition; Pudoc: Wageningen, The Netherlands, 1960. [Google Scholar]
- Willey, R.W.; Rao, M.R. A competitive ratio for quantifying competition between intercrops. Exp. Agric. 1980, 16, 117–125. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Wrigh, J.P.; Cadotte, M.W.; Carroll, I.T.; Hector, A.; Srivastava, D.S.; Loreau, M.; Weis, J.J. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc. Nat. Acad. Sci. USA 2007, 104, 18123–18128. [Google Scholar] [CrossRef]
- Karkanis, A.; Ntatsi, G.; Lepse, L.; Fernandez, J.A.; Savvas, D. Faba Bean Cultivation—Revealing Novel Managing Practices for More Sustainable and Competitive European Cropping Systems. Front. Plant Sci. 2018, 9, 1115. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Stomph, T.J.; Makowski, D.; van Der Werf, W. Temporal niche differentiation increases the land equivalent ratio of annual intercrops: A meta-analysis. Field Crops Res. 2015, 184, 133–144. [Google Scholar] [CrossRef]
- Hamzei, J.; Seyyedi, M. Energy use and input–output costs for sunflower production in sole and intercropping with soybean under different tillage systems. Soil Tillage Res. 2016, 157, 73–82. [Google Scholar] [CrossRef]
- Yufeng, L.; Hongliang, L.; Sicun, Y.; Jianguo, S.; Long, L.I. Yield and inter-specific interactions in maize/chickpea intercrop under different application rates of P in irrigated sierozem along the Yellow River in Northwest China. Chin. J. Eco-Agric. 2010, 18, 6. [Google Scholar]
- Xia, W.; Wu, F.; Zhou, X.; Fu, X.; Yue, T.; Xu, W.; Kai, P.; Liu, S. Effects of Intercropping with Potato Onion on the Growth of Tomato and Rhizosphere Alkaline Phosphatase Genes Diversity. Front. Plant Sci. 2016, 7, 846. [Google Scholar]
- Ringeval, B.; Nowak, B.; Nesme, T.; Delmas, M.; Pellerin, S. Contribution of anthropogenic phosphorus to agricultural soil fertility and food production. Glob. Biogeochem. Cycles 2015, 28, 743–756. [Google Scholar] [CrossRef]
- Akhalkatsi, M. Climate Global Change on Reproduction and Diversity of Agricultural Plants in Semi-Arid Regions of Georgia (Caucasus Ecoregion). Agric. Res. Technol. Open Access J. 2017, 3, 555619. [Google Scholar] [CrossRef]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in European grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Reich, P.B.; Scheu, S. Increasing plant diversity effects on productivity with time due to delayed soil biota effects on plants. Basic Appl. Ecol. 2012, 13, 571–578. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, C.; Tang, X.; Li, H.; Zhang, F.; Rengel, Z.; Whalley, W.R.; Davies, W.J.; Shen, J. Increased soil phosphorus availability induced by faba bean root exudation stimulates root growth and phosphorus uptake in neighbouring maize. New Phytol. 2016, 209, 823–831. [Google Scholar] [CrossRef]
- Shi, X.W.; Guang-Ping, Q.I.; Wang, J.H.; Kang, Y.X.; Yan-Lin, M.A.; Wang, J.H.; Zhang, Z.P. Effects of wolfberry-legume intercropping on soil temperature and wolfberry fruit yield in Yellow River Pumping Irrigation Region of Gansu. J. Gansu Agric. Univ. 2018, 6, 111–119. [Google Scholar]
- Sadeghpour, A.; Jahanzad, E.; Esmaeili, A.; Hosseini, M.B.; Hashemi, M. Forage yield, quality and economic benefit of intercropped barley and annual medic in semi-arid conditions: Additive series. Field Crops Res. 2013, 148, 43–48. [Google Scholar] [CrossRef]
- Abtahi, M.; Majidi, M.M.; Mirlohi, A.; Saeidnia, F. Association analysis for seed yield, forage yield and traits related to drought tolerance in orchardgrass (Dactylis glomerata). Crop Pasture Sci. 2018, 69, 1150. [Google Scholar] [CrossRef]
- Majidi, M.M.; Araghi, B.; Barati, M.; Mirlohi, A. Polycross Genetic Analysis of Forage Yield and Related Traits in Dactylis glomerata. Crop Sci. 2015, 55, 203–210. [Google Scholar] [CrossRef]
- Fatemeh, S.; Mahdi, M.M.; Aghafakhr, M.; Benyamin, A. Physiological responses of drought tolerance in orchardgrass (Dactylis glomerata) in association with persistence and summer dormancy. Crop Pasture Sci. 2018, 69, 515–526. [Google Scholar]
- Lin, F.; Liu, X.J.; Tong, C.C.; Yong, W.U.; University, G.A. A study of root system characteristics and carbon and nitrogen metabolism of alfalfa and four grass forages in monoculture or intercropped. Acta Prataculturae Sin. 2019, 28, 45–54. [Google Scholar]
- Lin, F.; Liu, X.J.; Tong, C.C.; Wu, Y. Characteristics of light energy utilization of intercropping alfalfa/gramineae forage based on yield effect. Chin. J. Appl. Ecol. 2020, 31, 2963–2976. [Google Scholar]
- Ren, Y.Y.; Wang, Z.L.; Wang, X.L.; Zhang, S.Q. The effect and mechanism of intercropping pattern on yield and economic benefit on the Loess Plateau. Acta Ecol. Sin. 2015, 35, 4168–4177. [Google Scholar]
- Nedunchezhiyan, M. Evaluation of sweet potato (Ipomoea batatas) based strip intercropping systems for yield, competition indices and nutrient uptake. Indian J. Agron. 2011, 56, 98–103. [Google Scholar]
- Mousavi, S.R.; Eskandari, H. A General Overview on Intercropping and Its Advantages in Sustainable Agriculture. J. Appl. Environ. Biol. 2011, 1, 482–486. [Google Scholar]
- Zhu, L.; Tian, Y.; Wang, J.; Qin, K.; He, J. Plant allelopathy and its application in grassland agroecosystem. Soils Crops 2021, 9, 1–14. [Google Scholar]
- Li, L. Intercropping enhances agroecosystem services and functioning: Current knowledge and perspectives. Chin. J. Eco-Agric. 2016, 24, 403–415. [Google Scholar]
- Zheng, W.; Li, Y.; Gong, Q.; Zhang, H.; Wang, Z. Improving yield and water use efficiency of apple trees through intercrop-mulch of crown vetch (Coronilla varia L.) combined with different fertilizer treatments in the Loess Plateau. Span. J. Agric. Res. 2016, 14, e1207. [Google Scholar] [CrossRef]
- Gautam, P.; Lal, D.B.; Rana, R. Intercropping: An Alternative Pathway for Sustainable Agriculture. Pop. Kheti 2014, 1, 216–220. [Google Scholar]
- Alemayehu, D.; Shumi, D.; Afeta, T. Effect of Variety and Time of Intercropping of Common Bean (Phaseolus vulgaris L.) With Maize (Zea mays L.) on Yield Components and Yields of Associated Crops and Productivity of the System at Mid-Land of Guji, Southern Ethiopia. Adv. Crop Sci. Technol. 2018, 6, 1–9. [Google Scholar] [CrossRef]
- Jahanzad, E.; Sadeghpour, A.; Hoseini, M.B.; Barker, A.V.; Hashemi, M.; Afshar, R.K. Competition, Nitrogen Use Efficiency, and Productivity of Millet–Soybean Intercropping in Semiarid Conditions. Crop Sci. 2015, 55, 2842–2851. [Google Scholar] [CrossRef]
- Amanullah; Khalid, S.; Khalil, F. Imranuddin. Influence of irrigation regimes on competition indexes of winter and summer intercropping system under semi-arid regions of Pakistan. Sci. Rep. 2020, 10, 8129. [Google Scholar] [CrossRef]
- Lin, F. Study on the Mechanism of Improvement of Production Potential and Nutrient Quality of Alfalfa/Gramineae Grasses Intercropping and the Utilization Effect of Livestock; Gansu Agricultural University: Lanzhou, China, 2019. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cropping System | Treatment | Sampling Sites | |
|---|---|---|---|
| Greenhouse (Yinchuan) | Field (Zhongning) | ||
| intercropping | Wolfberry–ryegrass | О ∆ ✯ | О ∆ ✯ |
| Wolfberry–oats | О ∆ | О ∆ ✯ | |
| Wolfberry–sweet sorghum | О ∆ | О ∆ ✯ | |
| Wolfberry–Kudouzi | О ∆ | О ∆ ✯ | |
| Wolfberry–Lvyuan 5 | О ∆ ✯ | О ∆ ✯ | |
| Wolfberry–Stipas | О ∆ | О ∆ ✯ | |
| Wolfberry–wheatgrass | О ∆ | О ∆ ✯ | |
| Wolfberry–white clover | О ∆ ✯ | О ∆ ✯ | |
| Wolfberry–mangold | О ∆ ✯ | О ∆ ✯ | |
| Wolfberry–alfalfa | О ∆ ✯ | О ∆ ✯ | |
| monocropping | Ryegrass | О ∆ ✯ | О ∆ ✯ |
| Oats | О ∆ | О ∆ ✯ | |
| Sweet sorghum | О ∆ | О ∆ ✯ | |
| Kudouzi | О ∆ | О ∆ ✯ | |
| Lvyuan 5 | О ∆ ✯ | О ∆ ✯ | |
| Stipas | О ∆ | О ∆ ✯ | |
| Wheatgrass | О ∆ | О ∆ ✯ | |
| White clover | О ∆ ✯ | О ∆ ✯ | |
| Mangold | О ∆ ✯ | О ∆ ✯ | |
| Alfalfa | О ∆ ✯ | О ∆ ✯ | |
| Wolfberry | О ∆ ✯ | О ∆ ✯ | |
| Cropping System | Greenhouse Test | Field Test | ||||||
|---|---|---|---|---|---|---|---|---|
| Row Distance (cm) | Plant Distance (cm) | Seeding Rate (g/m2) | Depth (cm) | Row Distance (cm) | Plant Distance (cm) | Seeding Rate (g/m2) | Depth (cm) | |
| ryegrass | 2 | -- | 7.5 | 2 | 10 | -- | 4.5 | 3 |
| oats | 4 | -- | 45 | 3 | 15 | -- | 22.5 | 4 |
| sweet sorghum | 5 | -- | 7.5 | 3 | 30 | -- | 3.5 | 4 |
| kudouzi | 3 | -- | 7.5 | 3 | 20 | -- | 3 | 3 |
| lvyuan 5 | 2 | -- | 7.5 | 2 | 10 | -- | 4.5 | 3 |
| stipas | 1 | -- | 8 | 0.5 | 10 | -- | 3.5 | 1 |
| wheatgrass | 4 | -- | 4.5 | 1.5 | 20 | -- | 2.5 | 1.5 |
| white clover | 3 | -- | 15 | 1 | 15 | -- | 5 | 1.5 |
| mangold | 10 | 10 | 6 | 2 | 30 | 20 | 3 | 3 |
| alfalfa | 3 | -- | 6.5 | 1.5 | 15 | -- | 3.5 | 2 |
| Cropping System | Field Test | |||
|---|---|---|---|---|
| Row Distance (cm) | Plant Distance (cm) | Number of Lines | Gap between Groups (cm) | |
| wolfberry–ryegrass | 10 | -- | 11 | 100 |
| wolfberry–oats | 15 | -- | 8 | 97.5 |
| wolfberry–sweet sorghum | 30 | -- | 4 | 105 |
| wolfberry–kudouzi | 20 | -- | 6 | 100 |
| wolfberry–lvyuan 5 | 10 | -- | 11 | 100 |
| wolfberry–stipas | 10 | -- | 11 | 100 |
| wolfberry–wheatgrass | 10 | -- | 11 | 100 |
| wolfberry–white clover | 15 | -- | 8 | 97.5 |
| wolfberry–mangold | 50 | 20 | 3 | 100 |
| wolfberry–alfalfa | 15 | -- | 8 | 97.5 |
| Treatment | Year | Gramineous Yield (kg/T) | Increased Biomass (kg/T) | Above-Ground Biomass (kg/T) | ||||
|---|---|---|---|---|---|---|---|---|
| Monocropping | Intercropping | Monocropping | Intercropping | Monocropping Weighted Average | Intercropping Yield | LER | ||
| 2019 | 1.12 ± 0.08 a | 0.56 ± 0.01 b | 1.66 ± 0.16 a | 0.98 ± 0.08 b | 1.31 ± 0.07 b | 1.54 ± 0.02 a | 1.09 ns | |
| wolfberry– lvyuan 5 | 2020 | 1.30 ± 0.01 a | 0.68 ± 0.04 b | 1.90 ± 0.19 a | 1.19 ± 0.06 b | 1.51 ± 0.10 b | 1.87 ± 0.06 a | 1.15 * |
| 2021 | 1.36 ± 0.06 a | 0.80 ± 0.06 b | 2.30 ± 0.19 a | 1.25 ± 0.09 b | 1.69 ± 0.07 b | 2.05 ± 0.12 a | 1.13 ns | |
| 2019 | 1.44 ± 0.16 a | 0.52 ± 0.03 b | 1.66 ± 0.16 a | 0.82 ± 0.13 b | 1.52 ± 0.11 a | 1.34 ± 0.10 a | 0.85 * | |
| wolfberry– oats | 2020 | 1.20 ± 0.08 a | 0.49 ± 0.01 b | 1.90 ± 0.19 a | 1.06 ± 0.13 b | 1.45 ± 0.05 a | 1.55 ± 0.08 a | 0.96 ns |
| 2021 | 1.23 ± 0.03 a | 0.46 ± 0.06 b | 2.30 ± 0.19 a | 1.29 ± 0.16 b | 1.61 ± 0.04 a | 1.75 ± 0.03 a | 0.94 ns | |
| 2019 | 1.72 ± 0.01 a | 0.56 ± 0.03 b | 1.66 ± 0.16 a | 0.51 ± 0.10 b | 1.70 ± 0.12 a | 1.07 ± 0.05 b | 0.63 ** | |
| wolfberry– wheatgrass | 2020 | 1.58 ± 0.14 a | 0.54 ± 0.07 b | 1.90 ± 0.19 a | 0.58 ± 0.11 b | 1.69 ± 0.09 a | 1.12 ± 0.01 b | 0.65 ** |
| 2021 | 1.86 ± 0.12 a | 0.70 ± 0.01 b | 2.30 ± 0.19 a | 0.87 ± 0.12 b | 2.02 ± 0.13 a | 1.57 ± 0.03 b | 0.75 * | |
| 2019 | 1.32 ± 0.10 a | 0.76 ± 0.09 b | 1.66 ± 0.16 a | 1.24 ± 0.08 b | 1.44 ± 0.03 b | 2.00 ± 0.12 a | 1.32 ** | |
| wolfberry–stipas | 2020 | 1.74 ± 0.12 a | 0.80 ± 0.03 b | 1.90 ± 0.19 a | 1.55 ± 0.08 b | 1.80 ± 0.07 b | 2.35 ± 0.12 a | 1.27 * |
| 2021 | 1.50 ± 0.02 a | 0.54 ± 0.05 b | 2.30 ± 0.19 a | 1.56 ± 0.10 b | 1.78 ± 0.06 b | 2.10 ± 0.07 a | 1.04 ns | |
| 2019 | 2.02 ± 0.18 a | 1.30 ± 0.07 b | 1.66 ± 0.16 a | 1.86 ± 0.08 a | 1.90 ± 0.09 b | 3.16 ± 0.23 a | 1.76 *** | |
| wolfberry– ryegrass | 2020 | 2.42 ± 0.13 a | 1.20 ± 0.02 b | 1.90 ± 0.19 b | 2.07 ± 0.10 a | 2.24 ± 0.15 b | 3.27 ± 0.26 a | 1.59 *** |
| 2021 | 2.16 ± 0.10 a | 1.40 ± 0.10 b | 2.30 ± 0.19 a | 2.18 ± 0.08 a | 2.21 ± 0.19 b | 3.58 ± 0.19 a | 1.60 *** | |
| 2019 | 21.02 ± 2.37 a | 5.82 ± 1.02 b | 1.66 ± 0.16 a | 1.23 ± 0.13 b | 14.25 ± 2.52 a | 7.05 ± 1.07 b | 1.02 ns | |
| wolfberry– sweet sorghum | 2020 | 19.87 ± 2.08 a | 6.03 ± 0.89 b | 1.90 ± 0.19 a | 1.52 ± 0.09 b | 13.58 ± 2.01 a | 7.55 ± 0.90 b | 1.10 ns |
| 2021 | 24.75 ± 3.69 a | 5.46 ± 0.77 b | 2.30 ± 0.19 a | 1.69 ± 0.12 b | 16.89 ± 1.69 a | 7.15 ± 1.62 b | 0.95 ns | |
| W-G | Mean | 4.98 ± 0.52 A | 1.59 ± 0.21 B | 1.96 ± 0.18 A | 1.30 ± 0.09 B | 3.92 ± 0.61 A | 2.89 ± 0.28 B | 1.10 ns |
| 2019 | 0.87 ± 0.09 a | 0.13 ± 0.01 b | 1.66 ± 0.16 a | 0.78 ± 0.02 b | 1.15 ± 0.06 a | 0.91 ± 0.01 a | 0.62 ** | |
| wolfberry– kudouzi | 2020 | 1.02 ± 0.03 a | 0.10 ± 0.02 b | 1.90 ± 0.19 a | 1.41 ± 0.01 b | 1.33 ± 0.08 a | 1.51 ± 0.07 a | 0.84 * |
| 2021 | 0.93 ± 0.10 a | 0.16 ± 0.01 b | 2.30 ± 0.19 a | 1.69 ± 0.09 b | 1.41 ± 0.19 b | 1.85 ± 0.12 a | 0.90 ns | |
| wolfberry– alfalfa | 2020 | 1.08 ± 0.09 a | 0.69 ± 0.03 a | 1.90 ± 0.19 a | 1.52 ± 0.10 a | 1.37 ± 0.07 b | 2.21 ± 0.14 a | 1.44 *** |
| 2021 | 1.32 ± 0.10 a | 0.73 ± 0.09 b | 2.30 ± 0.19 a | 1.58 ± 0.12 b | 1.66 ± 0.10 b | 2.31 ± 0.12 a | 1.24 * | |
| 2019 | 0.68 ± 0.01 a | 0.28 ± 0.06 b | 1.66 ± 0.16 a | 1.70 ± 0.09 a | 1.02 ± 0.02 b | 1.98 ± 0.09 a | 1.43 ** | |
| wolfberry– white clover | 2020 | 1.00 ± 0.05 a | 0.18 ± 0.00 b | 1.90 ± 0.19 a | 1.81 ± 0.05 a | 1.32 ± 0.08 b | 1.99 ± 0.10 a | 1.13 ns |
| 2021 | 1.08 ± 0.07 a | 0.20 ± 0.02 b | 2.30 ± 0.19 a | 1.90 ± 0.11 b | 1.51 ± 0.06 b | 2.10 ± 0.14 a | 1.01 ns | |
| W-L | Mean | 1.08 ± 0.07 A | 0.36 ± 0.03 B | 1.96 ± 0.18 A | 1.54 ± 0.07 B | 1.35 ± 0.07 B | 1.90 ± 0.10 A | 1.12 ns |
| 2019 | 14.22 ± 1.06 a | 10.89 ± 1.37 b | 1.66 ± 0.16 a | 1.75 ± 0.11 a | 9.83 ± 1.79 b | 12.64 ± 2.66 a | 1.82 *** | |
| wolfberry– mangold | 2020 | 17.01 ± 2.19 a | 14.68 ± 2.01 b | 1.90 ± 0.19 a | 1.77 ± 0.09 a | 11.72 ± 1.04 b | 16.45 ± 0.09 a | 1.79 *** |
| 2021 | 15.66 ± 1.77 b | 18.32 ± 2.42 a | 2.30 ± 0.19 a | 2.23 ± 0.12 a | 10.99 ± 2.01 b | 20.55 ± 2.47 a | 2.14 *** | |
| W-C | Mean | 15.63 ± 1.67 A | 14.63 ± 1.93 B | 1.96 ± 0.18 A | 1.92 ± 0.11 B | 10.84 ± 1.61 B | 16.55 ± 1.74 A | 1.92 *** |
| Treatment | Year | Gramineous Yield (kg/667 m2) | Increased Biomass (kg/667 m2) | Above-Ground Biomass (kg/667 m2) | ||||
|---|---|---|---|---|---|---|---|---|
| Monocropping | Intercropping | Monocropping | Intercropping | Monocropping Weighted Average | Intercropping Yield | LER | ||
| 2019 | 3293.38 ± 140.49 a | 2138.92 ± 110.72 b | 4412.50 ± 495.30 a | 2606.09 ± 216.53 b | 4613.30 ± 202.45 b | 4745.01 ± 326.77 a | 1.26 * | |
| wolfberry–lvyuan 5 | 2020 | 2937.95 ± 136.77 a | 1957.37 ± 63.92 b | 5051.51 ± 388.98 a | 3157.42 ± 201.33 b | 4994.24 ± 142.09 b | 5114.80 ± 121.45 a | 1.31 ** |
| 2021 | 2770.60 ± 144.24 a | 1917.65 ± 70.11 b | 6111.01 ± 526.37 a | 3322.20 ± 208.88 b | 5121.74 ± 206.87 b | 5239.854 ± 208.77 a | 1.26 * | |
| 2019 | 3610.48 ± 124.01 a | 2397.72 ± 154.08 b | 4412.50 ± 495.30 a | 2169.77 ± 177.09 b | 4011.49 ± 162.45 b | 4567.49 ± 165.96 a | 1.11 ns | |
| wolfberry–oats | 2020 | 3156.19 ± 116.33 a | 1867.04 ± 140.00 b | 5051.51 ± 388.98 a | 2802.98 ± 363.44 b | 4103.85 ± 78.89 b | 4670.00 ± 277.53 a | 1.11 ns |
| 2021 | 3320.71 ± 107.99 a | 2010.35 ± 132.66 b | 6111.01 ± 526.37 a | 3428.75 ± 350.88 b | 4715.86 ± 233.78 b | 5439.09 ± 466.89 a | 1.13 ns | |
| 2019 | 2312.78 ± 164.60 a | 917.42 ± 104.07 b | 4412.50 ± 495.30 a | 1351.69 ± 88.45 b | 3362.64 ± 102.37 a | 2269.11 ± 178.96 b | 0.69 ** | |
| wolfberry–wheatgrass | 2020 | 2106.64 ± 163.97 a | 708.02 ± 66.77 b | 5051.51 ± 388.98 a | 1545.48 ± 101.67 b | 3579.07 ± 232.74 a | 2253.50 ± 130.62 b | 0.63 ** |
| 2021 | 1926.98 ± 161.44 a | 691.72 ± 99.35 b | 6111.01 ± 526.37 a | 2303.85 ± 78.22 b | 4019.00 ± 121.96 a | 2995.57 ± 128.99 b | 0.73 * | |
| 2019 | 2091.95 ± 179.23 a | 896.74 ± 95.18 b | 4412.50 ± 495.30 a | 3290.06 ± 190.52 b | 3252.23 ± 98.89 b | 4186.81 ± 301.56 a | 1.19 * | |
| wolfberry–stipas | 2020 | 1791.96 ± 169.81 a | 892.85 ± 103.66 b | 5051.51 ± 388.98 a | 4102.50 ± 177.43 b | 3421.73 ± 77.34 b | 4995.35 ± 266.84 a | 1.33 ** |
| 2021 | 1688.15 ± 188.92 a | 710.51 ± 45.29 b | 6111.01 ± 526.37 a | 4140.89 ± 281.76 b | 3899.58 ± 135.64 b | 4851.40 ± 199.35 a | 1.12 ns | |
| 2019 | 3611.79 ± 100.01 a | 3123.62 ± 259.56 b | 4412.50 ± 495.30 a | 4945.70 ± 178.78 b | 4012.14 ± 201.87 b | 8069.326 ± 377.56 a | 1.96 *** | |
| wolfberry–ryegrass | 2020 | 3786.48 ± 166.88 a | 3038.74 ± 281.32 b | 5051.51 ± 388.98 a | 5502.54 ± 233.09 b | 4418.99 ± 199.07 b | 8541.28 ± 368.54 a | 1.87 *** |
| 2021 | 3740.65 ± 125.97 a | 3213.45 ± 89.07 b | 6111.01 ± 526.37 a | 5792.25 ± 201.22 b | 4925.83 ± 207.43 b | 9005.70 ± 299.87 a | 1.78 *** | |
| 2019 | 10,639.36 ± 800.30 a | 3158.61 ± 157.09 b | 4412.50 ± 495.30 a | 3264.41 ± 308.44 b | 7525.93 ± 566.89 a | 6423.02 ± 465.99 b | 1.04 ns | |
| wolfberry–sweet sorghum | 2020 | 10,242.39 ± 916.43 a | 3082.51 ± 140.21 b | 5051.51 ± 388.98 a | 4020.30 ± 291.88 b | 7646.95 ± 394.08 a | 7102.84 ± 407.29 b | 1.10 ns |
| 2021 | 9388.89 ± 567.00 a | 2713.80 ± 201.88 b | 6111.01 ± 526.37 a | 4487.31 ± 288.89 b | 7749.95 ± 521.46 a | 7201.11 ± 366.55 b | 1.02 ns | |
| W-G | Mean | 4186.85 ± 277.96 a | 2004.80 ± 122.22 b | 5191.68 ± 361.14 a | 3457.45 ± 137.56 b | 4689.26 ± 234.98 b | 5462.26 ± 269.88 a | 1.20 * |
| 2019 | 3828.33 ± 104.76 a | 1390.33 ± 87.56 b | 4412.50 ± 495.30 a | 2076.65 ± 89.96 b | 4120.42 ± 88.32 a | 3466.98 ± 244.31 b | 0.75 * | |
| wolfberry–kudouzi | 2020 | 3516.87 ± 121.02 a | 808.90 ± 45.99 b | 5051.51 ± 388.98 a | 3749.36 ± 101.43 b | 4284.19 ± 155.46 a | 4558.26 ± 261.80 a | 0.92 ns |
| 2021 | 3476.68 ± 89.16 a | 801.19 ± 77.54 b | 6111.01 ± 526.37 a | 4478.61 ± 252.37 b | 4793.85 ± 174.08 b | 5279.80 ± 197.56 a | 0.91 ns | |
| 2019 | 5907.25 ± 169.73 a | 5182.18 ± 150.19 b | 4412.50 ± 495.30 a | 3991.26 ± 197.88 b | 5159.88 ± 227.99 b | 9173.43 ± 343.19 a | 1.74 *** | |
| wolfberry–alfalfa | 2020 | 5279.17 ± 125.10 a | 4311.06 ± 143.87 b | 5051.51 ± 388.98 a | 4033.01 ± 188.56 b | 5165.34 ± 209.17 b | 8344.07 ± 277.43 a | 1.58 *** |
| 2021 | 5676.63 ± 69.77 a | 4625.43 ± 109.77 b | 6111.01 ± 526.37 a | 4178.85 ± 102.33 b | 5893.82 ± 155.44 b | 8804.28 ± 256.38 a | 1.46 *** | |
| 2019 | 3240.63 ± 169.16 a | 1383.73 ± 111.08 b | 4412.50 ± 495.30 a | 4509.68 ± 200.67 a | 3826.56 ± 125.60 b | 5893.42 ± 200.67 a | 1.39 ** | |
| wolfberry–white clover | 2020 | 2936.30 ± 99.16 a | 993.32 ± 67.33 b | 5051.51 ± 388.98 a | 4812.09 ± 213.43 a | 3993.90 ± 277.86 b | 5805.61 ± 188.44 a | 1.24 * |
| 2021 | 2910.56 ± 116.55 a | 945.52 ± 69.45 b | 6111.01 ± 526.37 a | 5036.56 ± 300.59 b | 4510.78 ± 263.98 b | 5982.08 ± 277.30 a | 1.10 ns | |
| W-L | Mean | 4059.03 ± 133.26 a | 2220.20 ± 97.44 b | 5191.67 ± 452.53 a | 4096.23 ± 194.71 b | 4625.35 ± 179.86 b | 6316.43 ± 245.68 a | 1.23 * |
| 2019 | 10,980.08 ± 466.70 a | 8416.92 ± 411.09 b | 4412.50 ± 495.30 a | 4645.33 ± 127.31 a | 7696.29 ± 267.99 b | 13,062.25 ± 571.32 a | 1.86 *** | |
| wolfberry–mangold | 2020 | 9739.57 ± 301.19 a | 8209.06 ± 204.00 b | 5051.51 ± 388.98 a | 4700.40 ± 266.06 b | 7395.53 ± 233.07 b | 12,909.47 ± 406.38 a | 1.82 *** |
| 2021 | 9332.63 ± 355.87 a | 7647.04 ± 368.99 b | 6111.01 ± 526.37 a | 5915.80 ± 197.90 a | 7721.82 ± 401.90 b | 13,562.85 ± 476.51 a | 1.83 *** | |
| W-C | Mean | 10,017.43 ± 367.92 a | 8091.04 ± 327.56 b | 5191.68 ± 450.66 a | 5087.18 ± 197.35 a | 7604.55 ± 307.56 b | 13,178.21 ± 480.21 a | 1.84 *** |
| Treatment | Year | Gramineous Yield (kg/667 m2) | Increased Biomass (kg/667 m2) | Fruit Yield (kg/667 m2) | ||||
|---|---|---|---|---|---|---|---|---|
| Monocropping | Intercropping | Monocropping | Intercropping | Monocropping Weighted Average | Intercropping Yield | LER | ||
| 2019 | 3293.38 ± 140.49 a | 2138.92 ± 110.72 b | 902.18 ± 103.68 a | 711.62 ± 32.32 b | 2097.788 ± 127.41 b | 2850.55 ± 209.67 a | 1.46 *** | |
| wolfberry–lvyuan 5 | 2020 | 2937.95 ± 136.77 a | 1957.37 ± 63.92 b | 856.39 ± 99.67 a | 694.64 ± 37.66 b | 1897.17 ± 133.08 b | 2652.01 ± 177.09 a | 1.50 *** |
| 2021 | 2770.60 ± 144.24 a | 1917.65 ± 70.11 b | 783.05 ± 45.32 a | 663.45 ± 55.80 b | 1776.82 ± 106.54 b | 2581.10 ± 159.03 a | 1.56 *** | |
| 2019 | 3610.48 ± 124.01 a | 2397.72 ± 154.08 b | 892.18 ± 46.17 a | 743.33 ± 38.09 b | 2251.33 ± 187.95 b | 3141.05 ± 344.01 a | 1.45 *** | |
| wolfberry–oats | 2020 | 3156.19 ± 116.33 a | 1867.04 ± 140.00 b | 856.39 ± 39.88 a | 708.62 ± 24.99 b | 2006.29 ± 111.00 b | 2575.67 ± 209.60 a | 1.38 ** |
| 2021 | 3320.71 ± 107.99 a | 2010.35 ± 132.66 b | 783.05 ± 50.09 a | 694.64 ± 47.99 a | 2051.88 ± 146.57 b | 2704.99 ± 177.35 a | 1.45 ** | |
| 2019 | 2312.78 ± 164.60 a | 917.42 ± 104.07 b | 822.18 ± 67.34 a | 638.72 ± 37.54 b | 1567.48 ± 98.32 a | 1556.14 ± 176.45 a | 1.16 * | |
| wolfberry–wheatgrass | 2020 | 2106.64 ± 163.97 a | 708.02 ± 66.77 b | 856.39 ± 86.11 a | 611.19 ± 29.87 b | 1481.52 ± 121.56 a | 1319.21 ± 89.66 b | 1.04 ns |
| 2021 | 1926.98 ± 161.44 a | 691.72 ± 99.35 b | 783.05 ± 37.62 a | 567.05 ± 34.77 b | 1355.02 ± 49.87 a | 1258.77 ± 59.33 b | 1.07 ns | |
| 2019 | 2091.95 ± 179.23 a | 896.74 ± 95.18 b | 922.18 ± 107.55 a | 708.62 ± 59.83 b | 1507.07 ± 166.90 a | 1605.37 ± 201.77 a | 1.22 * | |
| wolfberry–stipas | 2020 | 1791.96 ± 169.81 a | 892.85 ± 103.66 b | 856.39 ± 55.47 a | 694.64 ± 77.19 b | 1324.18 ± 103.65 b | 1587.49 ± 78.55 a | 1.33 ** |
| 2021 | 1688.15 ± 188.92 a | 710.51 ± 45.29 b | 783.05 ± 65.42 a | 660.45 ± 45.80 b | 1235.60 ± 100.52 a | 1370.96 ± 19.67 a | 1.28 * | |
| 2019 | 3611.79 ± 100.01 a | 3123.62 ± 259.56 b | 862.18 ± 58.54 a | 858.83 ± 123.98 a | 2236.98 ± 19.67 a | 3982.45 ± 306.00 a | 1.84 *** | |
| wolfberry–ryegrass | 2020 | 3786.48 ± 166.88 a | 3038.74 ± 281.32 b | 856.39 ± 37.99 a | 836.33 ± 98.59 a | 2321.43 ± 169.88 b | 3875.07 ± 207.42 a | 1.76 *** |
| 2021 | 3740.65 ± 125.97 a | 3213.45 ± 89.07 b | 830.00 ± 51.09 a | 790.08 ± 99.45 a | 2285.32 ± 163.24 b | 4003.54 ± 188.56 a | 1.79 *** | |
| 2019 | 10,639.36 ± 800.30 a | 3158.61 ± 157.09 b | 902.18 ± 103.68 a | 772.83 ± 103.77 b | 5770.77 ± 374.89 a | 3931.44 ± 337.69 b | 1.15 * | |
| wolfberry–sweet sorghum | 2020 | 10,242.39 ± 916.43 a | 3082.51 ± 140.21 b | 856.39 ± 99.67 a | 733.33 ± 81.43 b | 5549.39 ± 403.66 a | 3815.87 ± 288.15 b | 1.16 * |
| 2021 | 9388.89 ± 567.00 a | 2713.80 ± 201.88 b | 783.05 ± 45.32 a | 703.69 ± 76.33 a | 5085.97 ± 261.90 a | 3417.49 ± 271.46 b | 1.19 * | |
| W-G | Mean | 4186.85 ± 277.96 a | 2004.80 ± 122.22 b | 843.70 ± 48.79 a | 710.67 ± 66.37 b | 2515.28 ± 111.05 a | 2715.48 ± 201.01 a | 1.38 ** |
| 2019 | 3828.33 ± 104.76 a | 1390.33 ± 87.56 b | 902.18 ± 50.12 a | 694.64 ± 37.08 b | 2365.26 ± 127.34 a | 2084.97 ± 59.81 b | 1.05 ns | |
| wolfberry–kudouzi | 2020 | 3516.87 ± 121.02 a | 808.90 ± 45.99 b | 856.39 ± 49.08 a | 660.45 ± 58.66 b | 2186.63 ± 87.91 a | 1469.35 ± 43.22 b | 0.95 ns |
| 2021 | 3476.68 ± 89.16 a | 801.19 ± 77.54 b | 783.05 ± 33.56 a | 608.62 ± 56.22 b | 2129.87 ± 91.56 a | 1409.81 ± 36.37 b | 0.95 ns | |
| 2019 | 5907.25 ± 169.73 a | 5182.18 ± 150.19 b | 868.45 ± 67.44 a | 740.33 ± 38.96 b | 3387.85 ± 203.22 b | 5922.51 ± 269.88 a | 1.69 *** | |
| wolfberry–alfalfa | 2020 | 5279.17 ± 125.10 a | 4311.06 ± 143.87 b | 866.54 ± 104.37 a | 700.69 ± 51.34 b | 3072.85 ± 179.60 b | 5011.75 ± 209.46 a | 1.59 *** |
| 2021 | 5676.63 ± 69.77 a | 4625.43 ± 109.77 b | 834.19 ± 76.52 a | 698.62 ± 44.88 b | 3255.41 ± 153.22 b | 5324.05 ± 237.50 a | 1.62 *** | |
| 2019 | 3240.63 ± 169.16 a | 1383.73 ± 111.08 b | 808.64 ± 66.09 a | 810.13 ± 77.42 a | 2024.64 ± 80.99 a | 2193.86 ± 95.66 a | 1.37 ** | |
| wolfberry–white clover | 2020 | 2936.30 ± 99.16 a | 993.32 ± 67.33 b | 786.83 ± 57.86 a | 772.83 ± 107.09 a | 1861.56 ± 131.67 a | 1766.35 ± 101.74 a | 1.27 * |
| 2021 | 2910.56 ± 116.55 a | 945.52 ± 69.45 b | 753.69 ± 44.92 a | 753.69 ± 76.53 a | 1832.12 ± 97.33 a | 1699.21 ± 43.87 b | 1.28 * | |
| W-L | Mean | 4059.03 ± 133.26 a | 2220.20 ± 97.44 b | 828.88 ± 60.89 a | 715.56 ± 63.27 b | 2443.95 ± 127.61 b | 2935.75 ± 113.42 a | 1.31 ** |
| 2019 | 10,980.08 ± 466.70 a | 8416.92 ± 411.09 b | 923.59 ± 101.54 a | 872.83 ± 127.66 a | 5951.84 ± 307.03 b | 9289.75 ± 399.80 a | 1.75 *** | |
| wolfberry–mangold | 2020 | 9739.57 ± 301.19 a | 8209.06 ± 204.00 b | 910.13 ± 144.21 a | 853.69 ± 49.67 a | 5324.85 ± 182.96 b | 9062.75 ± 277.46 a | 1.83 *** |
| 2021 | 9332.63 ± 355.87 a | 7647.04 ± 368.99 b | 872.83 ± 89.66 a | 843.33 ± 88.58 a | 5102.72 ± 201.44 b | 8490.37 ± 301.20 b | 1.83 *** | |
| W-C | Mean | 10,017.43 ± 367.92 a | 8091.04 ± 327.56 b | 902.18 ± 110.44 a | 856.62 ± 80.66 a | 5459.80 ± 233.54 b | 8947.65 ± 192.33 a | 1.80 *** |
| Cropping Pattern | Aggressivity | Relative Crowding Coefficient of Forage | Relative Crowding Coefficient of Wolfberry | Competitive Ratio | Monetary Advantage Index | ||||
|---|---|---|---|---|---|---|---|---|---|
| Afw | Afw | Kf | Kw | Kw | CRfw | CRfw | MAI | MAI | |
| (Fruit Yield) | (Total Biomass) | (Forage Biomass) | (Fruit Yield) | (Total Biomass) | (Fruit Yield) | (Total Biomass) | (Fruit Yield) | (Total Biomass) | |
| wolfberry–lvyuan 5 | −0.25 | 0.14 | 2.20 | 4.39 | 1.60 | 0.84 | 1.12 | 429.75 | 652.05 |
| wolfberry–oats | −0.53 | 0.00 | 1.39 | 5.58 | 1.40 | 0.69 | 1.00 | 514.24 | 437.53 |
| wolfberry–wheatgrass | −0.76 | −0.07 | 0.55 | 2.82 | 0.64 | 0.48 | 0.91 | 73.42 | −590.43 |
| wolfberry–stipas | −0.67 | −0.56 | 0.89 | 4.14 | 3.00 | 0.58 | 0.63 | 213.53 | 523.63 |
| wolfberry–ryegrass | −0.31 | −0.43 | 4.56 | 39.25 | −31.1 | 0.84 | 0.79 | 827.63 | 2106.54 |
| wolfberry–sweet sorghum | −1.15 | −0.95 | 0.42 | 6.66 | 3.38 | 0.34 | 0.38 | 449.24 | 320.18 |
| wolfberry–kudouzi | −1.12 | −0.93 | 0.27 | 3.40 | 2.10 | 0.27 | 0.31 | −11.07 | −301.12 |
| wolfberry–alfalfa | −0.07 | 0.02 | 3.97 | 4.98 | 3.77 | 0.96 | 1.01 | 994.18 | 1706.27 |
| wolfberry–white clover | −1.36 | −1.24 | 0.46 | 186.72 | 13.58 | 0.31 | 0.34 | 195.86 | 633.3 |
| wolfberry–mangold | −0.20 | −0.25 | 5.69 | 18.80 | 39.61 | 0.90 | 0.87 | 1918.57 | 3103.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Li, X.; He, J.; Zhou, X.; Wang, F.; Zhao, Y.; Liang, X.; Nan, X.; Li, Y.; Qin, K.; et al. Development of Lycium barbarum–Forage Intercropping Patterns. Agronomy 2023, 13, 1365. https://doi.org/10.3390/agronomy13051365
Zhu L, Li X, He J, Zhou X, Wang F, Zhao Y, Liang X, Nan X, Li Y, Qin K, et al. Development of Lycium barbarum–Forage Intercropping Patterns. Agronomy. 2023; 13(5):1365. https://doi.org/10.3390/agronomy13051365
Chicago/Turabian StyleZhu, Lizhen, Xiaoying Li, Jun He, Xiaoping Zhou, Fang Wang, Yan Zhao, Xiaojie Liang, Xiongxiong Nan, Yonghua Li, Ken Qin, and et al. 2023. "Development of Lycium barbarum–Forage Intercropping Patterns" Agronomy 13, no. 5: 1365. https://doi.org/10.3390/agronomy13051365
APA StyleZhu, L., Li, X., He, J., Zhou, X., Wang, F., Zhao, Y., Liang, X., Nan, X., Li, Y., Qin, K., & Cao, Y. (2023). Development of Lycium barbarum–Forage Intercropping Patterns. Agronomy, 13(5), 1365. https://doi.org/10.3390/agronomy13051365