
Physiological Basis of High Nighttime Temperature-Induced Chalkiness Formation during Early Grain-Filling Stage in Rice (Oryza sativa L.)

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and High-Temperature Treatment
2.2. Evaluation of Grain Chalkiness
2.3. Measurement of Photosynthetic Parameters
2.4. Ultrastructural Observation of Starch Granules
2.5. Measurement of Activities of Antioxidant Enzymes and Contents of Malondialdehyde
2.6. Measurement of the Activities of Enzymes Involved in Starch Biosynthesis
2.7. Measurement of the Contents of Endogenous Hormones
2.8. Rate and Duration of Grain Filling
2.9. Statistical Analysis
3. Results
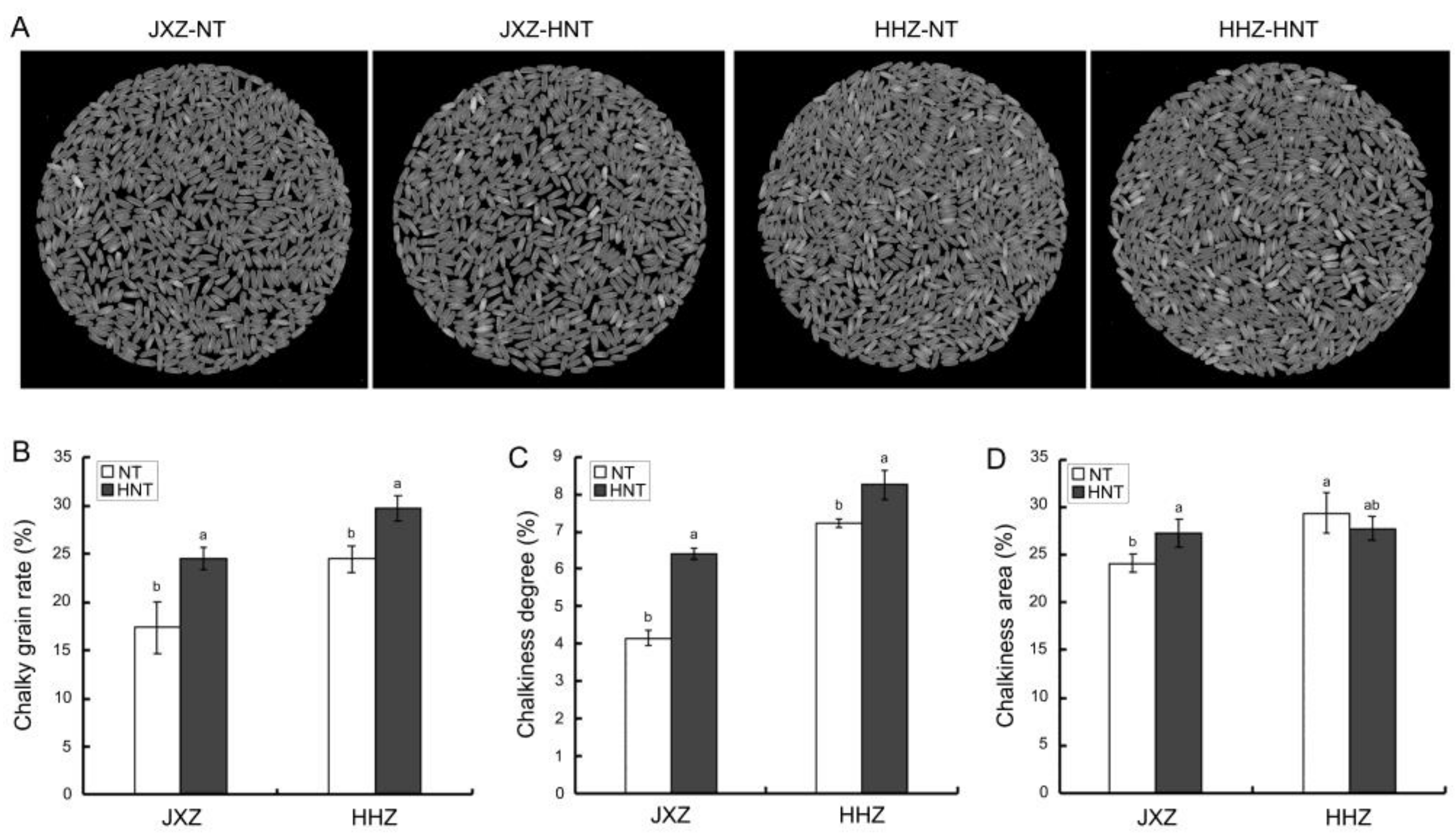
3.1. Chalky Grain Rate, Chalkiness Area, and Chalkiness Degree
3.2. Amyloplast Development in Endosperm
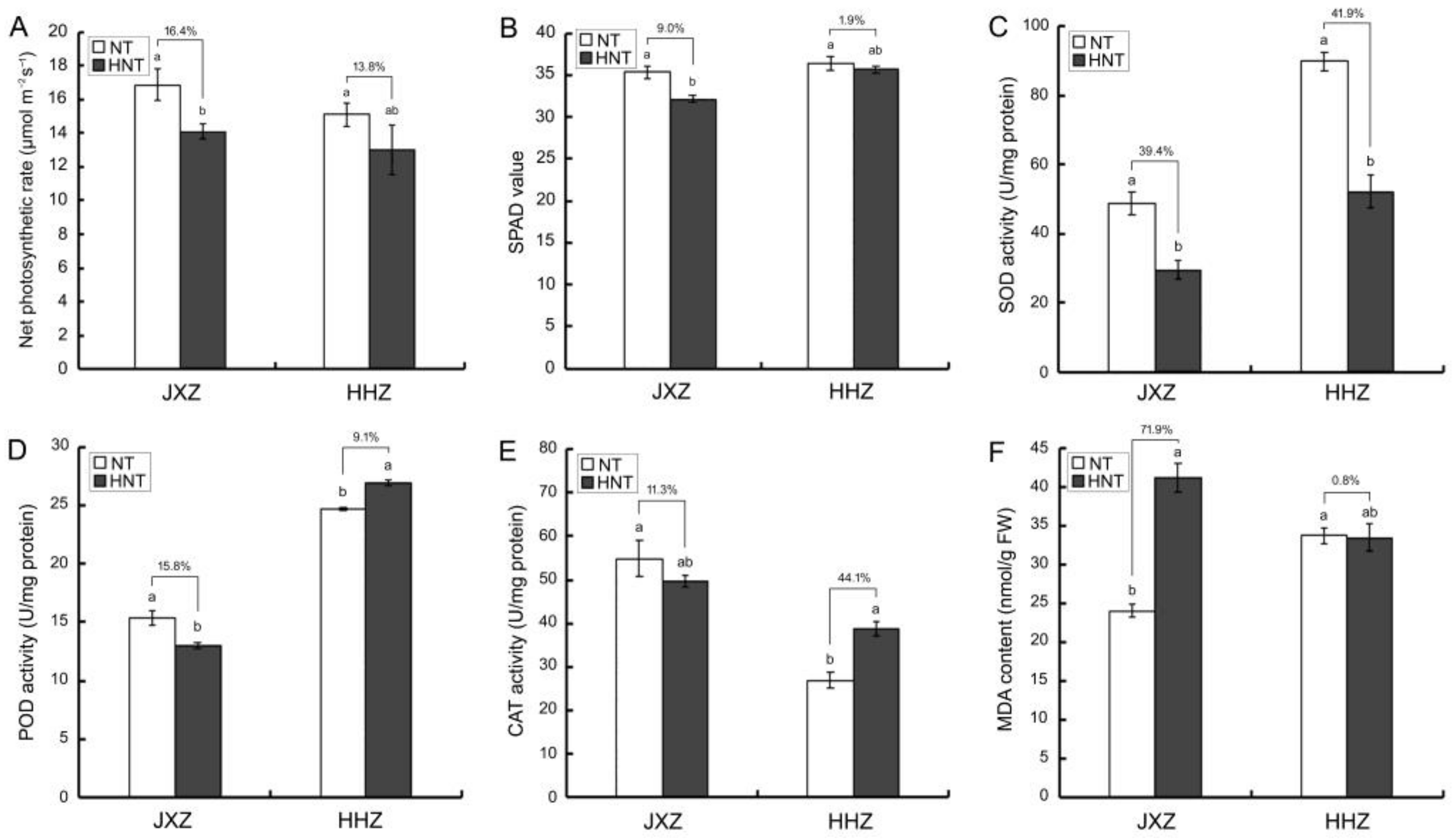
3.3. Photosynthetic Capacity and Activities of Antioxidant Enzymes
3.4. Rate and Duration of Grain Filling
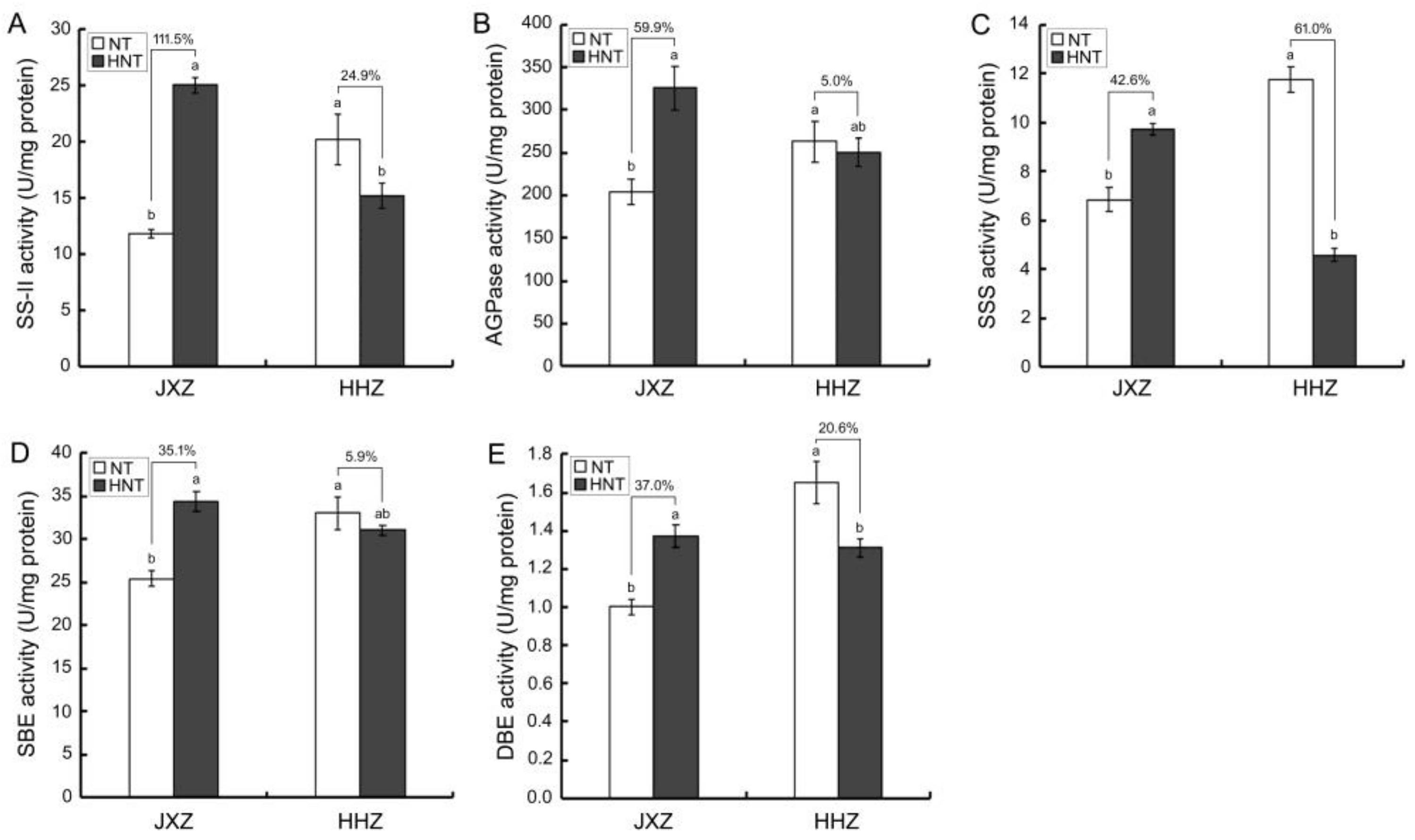
3.5. Enzymatic Activities Related to Starch Biosynthesis
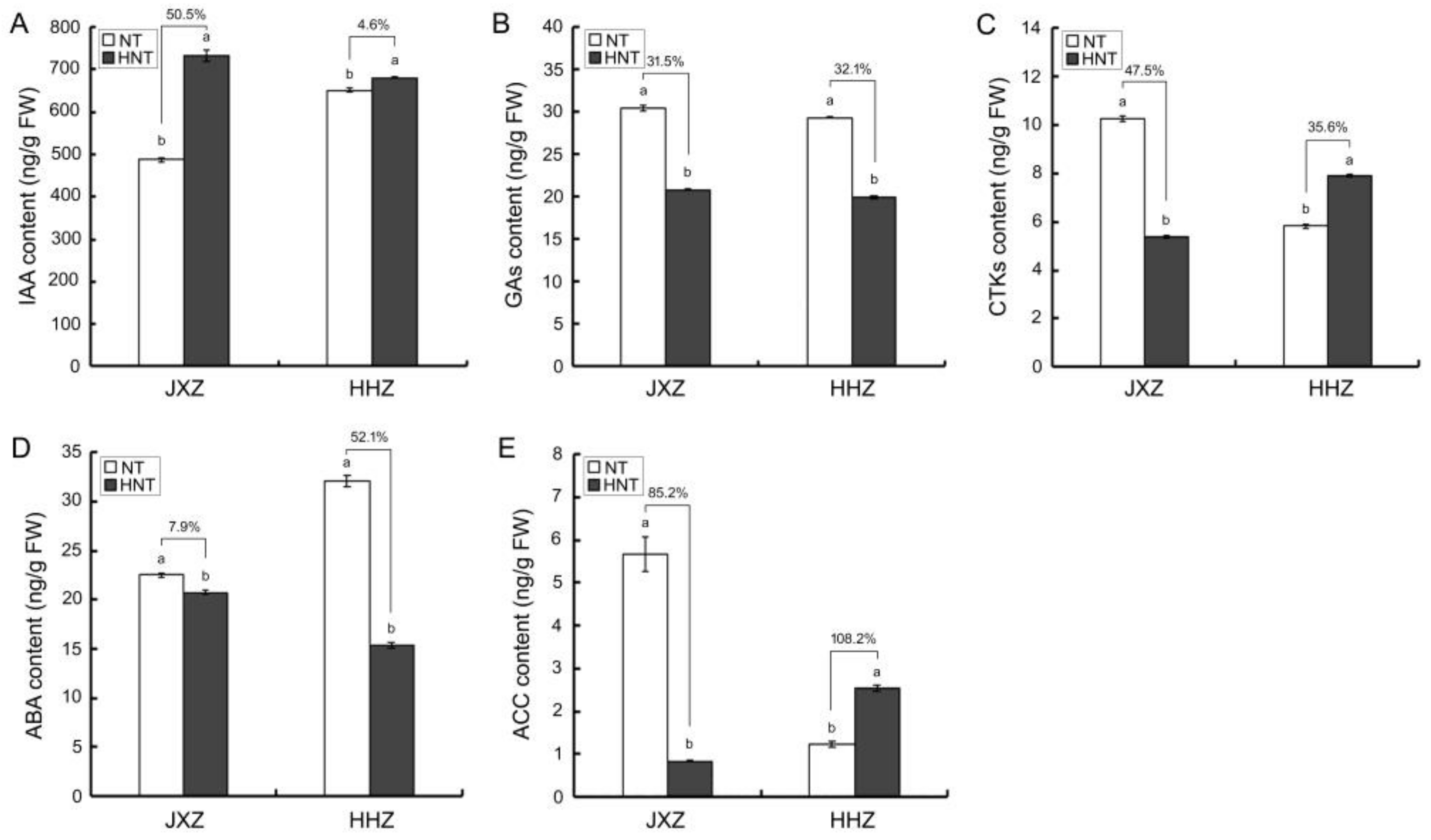
3.6. Levels of Endogenous Hormones
3.7. Relationship between Chalkiness and Physiological Indexes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Prasad, P.V.V.; Boote, K.J.; Allen, L.H.; Sheehy, J.E.; Thomas, J.M.G. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crop. Res. 2006, 95, 398–411. [Google Scholar] [CrossRef]
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V.R. High temperature effects on rice growth, yield, and grain quality. Adv. Agron. 2011, 111, 87–206. [Google Scholar]
- Xu, Y.F.; Chu, C.C.; Yao, S.G. The impact of high-temperature stress on rice: Challenges and solutions. Crop J. 2021, 9, 963–976. [Google Scholar] [CrossRef]
- Mohammed, R.; Cothren, J.T.; Tarpley, L. High night temperature and abscisic acid affect rice productivity through altered photosynthesis, respiration and spikelet fertility. Crop Sci. 2013, 53, 2603–2612. [Google Scholar] [CrossRef]
- Shi, W.; Muthurajan, R.; Rahman, H.; Selvam, J.; Peng, S.; Zou, Y.; Jagadish, K.S.V. Source-sink dynamics and proteomic reprogramming under elevated night temperature and their impact on rice yield and grain quality. New Phytol. 2013, 197, 825–837. [Google Scholar] [CrossRef]
- Shao, C.; Shen, L.; Qiu, C.; Wang, Y.; Qian, Y.; Chen, J.; Ouyang, Z.; Zhang, P.; Guan, X.; Xie, J.; et al. Characterizing the impact of high temperature during grain filling on phytohormone levels, enzyme activity and metabolic profiles of an early indica rice variety. Plant Biol. 2021, 23, 5806–5818. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Tetlow, I.J.; Nawaz, S.; Iqbal, A.; Mubin, M.; ul Rehman, M.S.N.; Butt, A.; Lightfoot, D.A.; Maekawa, M. Effect of high temperature on grain filling period, amylose content and activity of starch biosynthesis enzymes in endosperm of basmati rice. J. Sci. Food Agric. 2015, 95, 2237–2243. [Google Scholar] [CrossRef]
- Huang, M.; Zhang, H.; Zhao, C.; Chen, G.; Zou, Y. Amino acid content in rice grains is affected by high temperature during the early grain-filling period. Sci. Rep. 2019, 9, 2700. [Google Scholar] [CrossRef]
- Zhong, L.J.; Cheng, F.M.; Wen, X.; Sun, Z.X.; Zhang, G.P. The deterioration of eating and cooking quality caused by high temperature during grain filling in early-season indica rice cultivars. J. Agron. Crop Sci. 2005, 191, 218–225. [Google Scholar] [CrossRef]
- Coast, O.; Ellis, R.H.; Murdoch, A.J.; Quinones, C.; Jagadish, K.S.V. High night temperature induces contrasting responses for spikelet fertility, spikelet tissue temperature, flowering characteristics and grain quality in rice. Funct. Plant Biol. 2015, 42, 149–161. [Google Scholar] [CrossRef]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Sakai, H.; Yagi, K.; Hasegawa, T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric. Forest Meteorol. 2009, 149, 51–58. [Google Scholar] [CrossRef]
- Mohammed, A.R.; Tarpley, L. Impact of high nighttime temperature on respiration membrane stability, antioxidant capacity and yield of rice plants. Crop Sci. 2009, 49, 313–322. [Google Scholar] [CrossRef]
- Dong, W.; Tian, Y.; Zhang, B.; Chen, J.; Zhang, W. Effects of asymmetric warming on grain quality and related key enzymes activities for japonica rice (Nanjing 44) under FATI facility. Acta Agron. Sin. 2011, 37, 832–841. [Google Scholar] [CrossRef]
- Xu, J.; Misra, G.; Sreenivasulu, N.; Henry, A. What happens at night? Physiological mechanisms related to maintaining grain yield under high night temperature in rice. Plant Cell Environ. 2021, 44, 2245–2261. [Google Scholar] [CrossRef] [PubMed]
- Counce, P.A.; Bryant, R.J.; Bergman, C.J.; Bautista, R.C.; Wang, Y.J.; Siebenmorgen, T.J.; Moldenhauer, K.A.K.; Meullenet, J.F.C. Rice milling quality, grain dimensions, and starch branching as affected by high night temperatures. Cereal Chem. 2005, 82, 645–648. [Google Scholar] [CrossRef]
- Song, X.; Du, Y.; Zhao, Q.; Cui, Y. Effects of high night temperature during grain filling on formation of physicochemical properties for japonica rice. J. Cereal Sci. 2015, 66, 74–80. [Google Scholar] [CrossRef]
- Bahuguna, R.N.; Solis, C.A.; Shi, W.J.; Jagadish, S.V.K. Post-flowering night respiration and altered sink activity account for high night temperature-induced grain yield and quality loss in rice (Oryza sativa L.). Physiol. Plant. 2017, 159, 59–73. [Google Scholar] [CrossRef]
- Gong, D.; Zhang, X.; He, F.; Chen, Y.; Li, R.; Yao, J.; Zhang, M.; Zheng, W.; Yu, G. Genetic improvements in rice grain quality: A review of elite genes and their applications in molecular breeding. Agronomy 2023, 13, 1375. [Google Scholar] [CrossRef]
- Nevame, A.Y.M.; Emon, R.M.; Malek, M.A.; Hasan, M.M.; Alam, M.A.; Muharam, F.M.; Aslani, F.; Rafii, M.Y.; Ismail, M.R. Relationship between high temperature and formation of chalkiness and their effects on quality of rice. Biomed. Res. Int. 2018, 2018, 1653721. [Google Scholar] [CrossRef]
- Deng, F.; Li, Q.P.; Chen, H.; Zeng, Y.L.; Li, B.; Zhong, X.Y.; Wang, L.; Ren, W.J. Relationship between chalkiness and the structural and thermal properties of rice starch after shading during grain-filling stage. Carbohyd. Polym. 2021, 252, 117212. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhang, C.; Li, Q.; Liu, Q. Genetic control of grain appearance quality in rice. Biotechnol. Adv. 2022, 60, 108014. [Google Scholar] [CrossRef] [PubMed]
- Siddik, M.A.; Zhang, J.; Chen, J.; Qian, H.; Jiang, Y.; Raheem, A.K.; Deng, A.; Song, Z.; Zheng, C.; Zhang, W. Responses of indica rice yield and quality to extreme high and low temperatures during the reproductive period. Eur. J. Agron. 2019, 106, 30–38. [Google Scholar] [CrossRef]
- Sabaruddin, Z.; Toshiaki, M.; Shingo, T.; Youji, N. Effect of high temperature at ripening stage on the reserve accumulation in seed in some rice cultivars. Plant Prod. Sci. 2002, 5, 160–168. [Google Scholar]
- Zhang, G.; Liao, B.; Li, B.; Cai, Z. The effect of high temperature after anthesis on rice quality and starch granule structure of endosperm. Meteorol. Environ. Res. 2016, 7, 72–75. [Google Scholar]
- Tateyama, M.; Sakai, M.; Suto, M. Varietal differences in the response of the amylose content of the endosperm of low-amylose rice (Oryza sativa L.) lines to temperature during the ripening period. Breed. Res. 2005, 7, 1–7. [Google Scholar] [CrossRef]
- Umemoto, T.; Terashima, K. Activity of granule-bound starch synthase is an important determinant of amylose content in rice endosperm. Funct. Plant Biol. 2002, 29, 1121–1124. [Google Scholar] [CrossRef]
- Zhu, L.; Shah, F.; Nie, L.; Cui, K.; Shah, T.; Wu, W.; Chen, Y.; Chen, C.; Wang, K.; Wang, Q.; et al. Efficacy of sowing date adjustment as a management strategy to cope with rice (Oryza sativa L.) seed quality deterioration due to elevated temperature. Aust. J. Agric. Res. 2013, 7, 543–549. [Google Scholar]
- Umemoto, T.; Nakamura, Y.; Satoh, H.; Terashima, K. Differences in amylopectin structure between two rice varieties in relation to the effects of temperature during grain-filling. Starch-Starke 1999, 51, 58–62. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Sasaki, O.; Uezono, I.; Tanaka, A. Effects of high air temperature during the ripening period on the grain quality of rice in warm regions of Japan. Jpn. J. Crop Sci. 2007, 76, 71–78. [Google Scholar] [CrossRef]
- Vose, R.S.; Easterling, D.R.; Gleason, B. Maximum and minimum temperature trends for the globe: An update through 2004. Geophys. Res. Lett. 2005, 32, L23822. [Google Scholar] [CrossRef]
- Shi, W.; Yin, X.; Struik, P.C.; Solis, C.; Xie, F.; Schmidt, R.C.; Huang, M.; Zou, Y.; Ye, C.; Jagadish, S.V.K. High day- and night-time temperatures affect grain growth dynamics in contrasting rice genotypes. J. Exp. Bot. 2017, 68, 5233–5245. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Ali, A.; Shi, Q.; Zeng, Y.; Tan, X.; Shang, Q.; Huang, S.; Xie, X.; Zeng, Y. Response of chalkiness in high-quality rice (Oryza sativa L.) to temperature across different ecological regions. J. Cereal Sci. 2019, 87, 39–45. [Google Scholar] [CrossRef]
- Liu, D.; Zeng, M.; Wu, Y.; Du, Y.; Liu, J.; Luo, S.; Zeng, Y. Comparative transcriptomic analysis provides insights into the molecular basis underlying pre-harvest sprouting in rice. BMC Genom. 2022, 23, 771. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Guo, Z.F.; Yi, Y.; Li, H.N.; Li, H.G.; Chen, L.J.; Ma, H.; Zhang, L.; Lin, J.W.; Zhong, M. Effects of cold acclimation and exogenous pytohormone abscisic acid treatment on physiological indicators of winterness wheat. J. Plant Sci. 2010, 5, 125–136. [Google Scholar] [CrossRef]
- Zhou, L.; Jiang, L.; Zhai, H.; Wan, J. Current status and strategies for improvement of rice grain chalkiness. Hereditas 2009, 31, 563–572. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef]
- Yang, W.; Wang, F.; Liu, L.N.; Sui, N. Responses of membranes and the photosynthetic apparatus to salt stress in Cyanobacteria. Front. Plant Sci. 2020, 11, 713. [Google Scholar] [CrossRef]
- Shi, W.; Yin, X.; Struik, P.C.; Xie, F.; Schmidt, R.C.; Jagadish, K.S.V. Grain yield and quality responses of tropical hybrid rice to high night-time temperature. Field Crop. Res. 2016, 190, 18–25. [Google Scholar] [CrossRef]
- Fan, X.; Li, Y.; Zhu, Y.; Wang, J.; Zhao, J.; Sun, X.; Pan, Y.; Bian, X.; Zhang, C.; Zhao, D.; et al. Characterization of physicochemical qualities and starch structures of two indica rice varieties tolerant to high temperature during grain filling. J. Cereal Sci. 2020, 93, 102966. [Google Scholar] [CrossRef]
- Viswanathan, C.; Khanna-Chopra, R. Effect of heat stress on grain growth, starch synthesis and protein synthesis in grains of wheat (Triticum aestivum L.) varieties differing in grain weight stability. J. Agron. Crop Sci. 2001, 186, 1–7. [Google Scholar] [CrossRef]
- Song, X.; Du, Y.; Song, X.; Zhao, Q. Effect of high night temperature during grain filling on amyloplast development and grain quality in Japonica rice. Cereal Chem. 2013, 90, 114–119. [Google Scholar] [CrossRef]
- Impa, S.M.; Raju, B.; Hein, N.T.; Sandhu, J.; Prasad, P.V.V.; Walia, H.; Jagadish, S.V.K. High night temperature effects on wheat and rice: Current status and way forward. Plant Cell Environ. 2021, 44, 2049–2065. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J. Grain-filling problem in ‘super’ rice. J. Exp. Bot. 2010, 61, 1–5. [Google Scholar] [CrossRef]
- Teng, Z.; Yu, H.; Wang, G.; Meng, S.; Liu, B.; Yi, Y.; Chen, Y.; Zheng, Q.; Liu, L.; Yang, J.; et al. Synergistic interaction between ABA and IAA due to moderate soil drying promotes grain filling of inferior spikelets in rice. Plant J. 2022, 109, 1457–1472. [Google Scholar] [CrossRef]
- Zhang, H.; Tan, G.; Yang, L.; Yang, J.; Zhang, J.; Zhao, B. Hormones in the grains and roots in relation to post-anthesis development of inferior and superior spikelets in Japonica/Indica hybrid rice. Plant Physiol. Biochem. 2009, 47, 195–204. [Google Scholar] [CrossRef]
- Wu, C.; Tang, S.; Li, G.; Wang, S.; Fahad, S.; Ding, Y. Roles of phytohormone changes in the grain yield of rice plants exposed to heat: A review. PeerJ 2019, 7, e7792. [Google Scholar] [CrossRef]
- Sekhar, S.; Panda, B.B.; Mohapatra, T.; Das, K.; Shaw, B.P.; Kariali, E.; Mohapatra, P.K. Spikelet-specific variation in ethylene production and constitutive expression of ethylene receptors and signal transducers during grain filling of compact- and lax-panicle rice (Oryza sativa) cultivars. J. Plant Physiol. 2015, 179, 21–34. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, Y.; Wang, J.; Yang, J.; Zhang, J. Polyamine and ethylene interactions in grain filling of superior and inferior spikelets of rice. Plant Growth Regul. 2012, 66, 215–228. [Google Scholar] [CrossRef]
- Yang, J.C.; Zhang, J.H.; Ye, Y.X.; Wang, Z.Q.; Zhu, Q.S.; Liu, L.J. Involvement of abscisic acid and ethylene in the responses of rice grains to water stress during filling. Plant Cell Environ. 2004, 27, 1055–1064. [Google Scholar] [CrossRef]
- Morita, X.; Nakano. Prospect for developing measures to prevent high-temperature damage to rice grain ripening. Jpn. J. Crop Sci. 2008, 7, 12. [Google Scholar] [CrossRef]
- Liu, X.; Guo, T.; Wan, X.; Wang, H.; Zhu, M.; Li, A.; Su, N.; Shen, Y.; Mao, B.; Zhai, H.; et al. Transcriptome analysis of grain-filling caryopses reveals involvement of multiple regulatory pathways in chalky grain formation in rice. BMC Genom. 2010, 11, 730. [Google Scholar] [CrossRef] [PubMed]
- Ashida, K.; Iida, S.; Yasui, T. Morphological, physical, and chemical properties of grain and flour from chalky rice mutants. Cereal Chem. 2009, 86, 225–231. [Google Scholar] [CrossRef]
- Xie, Q.; Xu, J.; Huang, K.; Su, Y.; Tong, J.; Huang, Z.; Huang, C.; Wei, M.; Lin, W.; Xiao, L. Dynamic formation and transcriptional regulation mediated by phytohormones during chalkiness formation in rice. BMC Plant Biol. 2021, 21, 308. [Google Scholar] [CrossRef]
- Zhu, D.; Qian, Z.; Wei, H.; Guo, B.; Xu, K.; Dai, Q.; Zhang, H.; Huo, Z. The effects of field pre-harvest sprouting on the morphological structure and physicochemical properties of rice (Oryza sativa L.) starch. Food Chem. 2019, 278, 10–16. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, L.; Lu, Y.; Yang, Y.; Feng, L.; Hao, W.; Li, Q.; Fan, X.; Zhao, D.; Liu, Q. Changes in the physicochemical properties and starch structures of rice grains upon pre-harvest sprouting. Carbohyd. Polym. 2020, 234, 115893. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef]
- Nakata, M.; Fukamatsu, Y.; Miyashita, T.; Hakata, M.; Kimura, R.; Nakata, Y.; Kuroda, M.; Yamaguchi, T.; Yamakawa, H. High temperature-induced expression of rice α-amylases in developing endosperm produces chalky grains. Front. Plant Sci. 2017, 8, 2089. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Treatment | W | GR0 | GRmax | GRmean | Tmax | D | R2 |
|---|---|---|---|---|---|---|---|---|
| JXZ | NT | 12.39a | 0.28a | 1.21b | 0.64b | 10.52a | 17.31a | 0.99 |
| HNT | 13.89a | 0.29a | 1.99a | 0.97a | 7.81b | 12.71b | 0.97 | |
| HHZ | NT | 17.39a | 0.23b | 2.78a | 1.23a | 10.62a | 14.36a | 0.99 |
| HNT | 15.49b | 0.25a | 2.32a | 1.08a | 8.66b | 11.63b | 0.99 |
| Index | Chalky Grain Rate | Chalkiness Degree |
|---|---|---|
| Net photosynthetic rate | −0.82 ** | −0.81 ** |
| SPAD | 0.08 | 0.17 |
| Superoxide dismutase (SOD) | 0.07 | 0.28 |
| Peroxidase (POD) | −0.64 * | −0.75 ** |
| Catalase (CAT) | −0.53 | −0.73 ** |
| Malondialdehyde (MDA) | 0.59 * | 0.58 * |
| Sucrose synthase (SS-II) | 0.31 | 0.33 |
| Adenosine diphosphoglucose pyrophosphorylase (AGP) | 0.41 | 0.39 |
| Soluble starch synthase (SSS) | −0.17 | −0.04 |
| Starch branching enzyme (SBE) | 0.64 * | 0.66 * |
| Debranching enzyme (DBE) | 0.54 | 0.67 * |
| Indole-3-acetic acid (IAA) | 0.77 ** | 0.77 ** |
| Gibberellins (GAs) | −0.74 ** | −0.62 * |
| Cytokinins (CTKs) | −0.50 | −0.58 * |
| Abscisic acid (ABA) | −0.34 | −0.15 |
| 1-aminocylopropane-1- carboxylicacid (ACC) | −0.61 * | −0.69 * |
| Initial grain-filling rate (GR0) | −0.08 | −0.19 |
| Maximum grain-filling rate (GRmax) | 0.61 * | 0.75 ** |
| Mean grain-filling rate (GRmean) | 0.62 * | 0.75 ** |
| Time taken to reach maximum grain-filling rate (Tmax) | −0.47 | −0.36 |
| Active grain-filling duration (D) | −0.76 ** | −0.79 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Long, C.; Deng, X.; Zhang, Z.; Liu, J.; Xu, Y.; Liu, D.; Zeng, Y. Physiological Basis of High Nighttime Temperature-Induced Chalkiness Formation during Early Grain-Filling Stage in Rice (Oryza sativa L.). Agronomy 2023, 13, 1475. https://doi.org/10.3390/agronomy13061475
Du Y, Long C, Deng X, Zhang Z, Liu J, Xu Y, Liu D, Zeng Y. Physiological Basis of High Nighttime Temperature-Induced Chalkiness Formation during Early Grain-Filling Stage in Rice (Oryza sativa L.). Agronomy. 2023; 13(6):1475. https://doi.org/10.3390/agronomy13061475
Chicago/Turabian StyleDu, Yanli, Changzhi Long, Xueyun Deng, Zhengwei Zhang, Jie Liu, Yanghaojun Xu, Dong Liu, and Yongjun Zeng. 2023. "Physiological Basis of High Nighttime Temperature-Induced Chalkiness Formation during Early Grain-Filling Stage in Rice (Oryza sativa L.)" Agronomy 13, no. 6: 1475. https://doi.org/10.3390/agronomy13061475
APA StyleDu, Y., Long, C., Deng, X., Zhang, Z., Liu, J., Xu, Y., Liu, D., & Zeng, Y. (2023). Physiological Basis of High Nighttime Temperature-Induced Chalkiness Formation during Early Grain-Filling Stage in Rice (Oryza sativa L.). Agronomy, 13(6), 1475. https://doi.org/10.3390/agronomy13061475