Abstract
Cassava breeding is a critical aspect of sustainable agriculture to address the challenges of food security and climate change. The present study aimed to assess the combining ability of six Thai cassava elite varieties for four key traits, including fresh root yield (FRY), fresh shoot yield (FSY), harvest index (HI) and starch content (SC) through an analysis of the diallel mating design over two years. Parental lines HB80 and R5 showed significant positive general combining ability (GCA) effects for high SC; R1 and HB80 for low FSY; and R1 for high FRY and HI. The genetic effects for FSY, SC and HI were primarily influenced by the GCA of the progenitors, while specific combining ability (SCA) was more important for the FRY than the other three traits. Cultivation conditions were also found to impact the evaluation of genetic effects, with a poor growing environment affecting the expression of the genotype. This study provides a useful basis for the selection of parental lines for cassava breeding programs and insights into the genetic control of yield and related traits in the Thai elite cassava germplasm.
1. Introduction
Cassava (Manihot esculenta Crantz) is a globally significant commercial food crop that is widely cultivated in tropical and subtropical regions. It serves as an essential source of food, feed, starch and starch-related products worldwide [1,2]. In Southeast Asia, particularly in Thailand, Cambodia and Vietnam, cassava is primarily grown for industrial starch production. Over the past 40 years, efforts have been made to breed high-yield varieties that are well-adapted to local soil and climate conditions [3,4]. Despite the extensive crossing and breeding efforts, newly selected varieties face an increasing challenge to outperform released cultivars such as the highly successful KU50 [3]. This situation suggests that alternative approaches may be necessary to enhance cassava breeding potential in Thailand.
Cassava breeding in Thailand has primarily involved inter-varietal crossing and selection at various trial stages [3]. Traditionally, breeding decisions have been based on the individual performance of different crossing combinations, with little or no information regarding the segregating progenies at the family level. To date, quantitative genetic analysis, including diallel analysis and combining ability analysis, has not been performed in Thai cassava breeding populations. The lack of genetic information on parents and segregating progenies may be the underlying reason for the limitations in our breeding selections for superior progenies. With the increasing availability of outstanding breeding progenitors, it is crucial to obtain genetic information about the parents and the hybrids they produce. This will enable the effective allocation of resources toward specific crosses with the highest breeding potential for recurrent phenotypic selection.
The selection of suitable progenitors for hybridization is a critical aspect of cassava breeding programs, and a combining ability analysis is a proven method for selecting potential parents [5]. Evaluating the performance of hybrid progenies commonly involves estimating the general (GCA) and specific (SCA) combining ability effects [6]. While diallel crossing and combining ability analyses have been utilized in cassava breeding programs in South America and Africa, these methods have yet to be applied in Thai cassava breeding populations. Previous studies have shown that these genetic effects can significantly influence root yield and disease resistance, making this information valuable for guiding effective breeding selection [7,8,9,10,11,12,13,14]. Therefore, it is crucial to determine these genetic effects in the Thai cassava population to improve the breeding scheme.
Cassava breeding is a demanding process that requires significant investments in terms of time, labor and resources. However, insights into the genetic basis of breeding crosses can make the breeding process more efficient and streamline expected outcomes. In particular, the use of combining ability analysis can help identify key parents and mating pairs for targeted breeding populations. In this study, six Thai elite cassava varieties were selected for diallel crossing to estimate GCA and SCA effects for four productivity traits: fresh root yield (FRY), fresh shoot yield (FSY), harvesting index (HI) and starch content (SC). The 15 F1 progenies and parents were evaluated using an augmented RCBD design in Northeastern Thailand. Productivity trials were conducted over two consecutive years, with rainfall, temperature and soil conditions monitored. This study aims to provide guidelines for cassava breeding in Thailand by inferring genetic effects on the four traits. By leveraging the Thai elite varieties, the potential for improving breeding efficiency and streamlining the breeding process can be realized.
2. Materials and Methods
2.1. Parental Lines and Hybridization
The selection of six progenitors was based on yield and the ability to make crosses such as flowering ability and flowering period. Two progenitors were landraces: Rayong 1 and Hanatee (R1 and HNT, respectively). Four were improved cultivars: Rayong 5, Rayong 90, Huay Bong 80 and Kasetsart 50 (R5, R90, HB80 and KU50, respectively). These progenitors were utilized in a 6 × 6 diallel mating design (Table 1). The direct and reciprocal crosses from each F1 family were pooled together. These varieties are the main ones frequently utilized in breeding programs in Thailand [15,16]. R1 is known for its large root crown and adaptability to various environments, although it generally produces low starch content [16]. HNT is the most widely cultivated variety for food in Thailand. R5 and R90 are high-yielding varieties developed by the Department of Agriculture, Ministry of Agriculture and Cooperatives, Thailand, known for their high starch content. HB80 was released by The Thai Tapioca Development Institute (TTDI) and Kasetsart University [17]. KU50 has been widely propagated in Thailand for 31 years since its release and is still highly popular.

Table 1.
Six parental lines used for 6 × 6 diallel crosses.
2.2. Experimental Design and Cultivation Conditions
Controlled pollinations of 6 × 6 diallel design were conducted, following the methods described in [18]. Botanical seeds were germinated in plastic bags for two months and then transplanted into the experimental field. A single row spacing of 1.2 × 1.0 m between and within rows was used. No evaluations were performed during the seedling stages. At ten months of age, the stems were harvested and cut into 20 cm cuttings before being planted in the experimental field for evaluation. Seedlings that displayed stunted growth or were small were discarded. For the evaluation, an augmented randomized complete block design (augmented in RCBD) [19] was employed. Each experimental block consisted of 30 genotypes including 24 seedling genotypes and 6 parental lines. Within each row, there were 10 cutting stems (plants). Each genotype was grown in duplicate in separate blocks. The experiment was conducted for two consecutive years at the same location.
The trials were conducted at the Thai Tapioca Development Institute (TTDI), located in Huay Bong, Dan Khun Thot District, Nakhon Ratchasima, Thailand. The geographical coordinates of the location are 15.157942 latitude and 101.505531 longitude. The site has an average elevation of 368 m above sea level. The predominant soil type is sand loam.
Cultivation was carried out using stem cuttings (20 cm) inserted into soil ridges with a spacing of 1.2 × 1.0 m. Twenty plants along the border were designated as guard rows. No irrigation was employed during the trials. The trials spanned two consecutive years: year 1 from June 2015 to May 2016 and year 2 from June 2016 to May 2017. Soil properties were monitored before planting (Table S1). The rainy season in Thailand typically occurs from mid-May to October. During the field trial period, the total rainfall recorded was 810.60 and 690.90 mm in year 1 and year 2, respectively (Figure S1). No precipitation was observed for a three-month period in 2017. The average monthly temperature was also monitored and indicated that the overall temperature was higher in the second year compared to the first (Figure S2).
2.3. Data Collection
At the 12th month after planting (MAP), eight plants from each genotype were harvested for evaluation. FSY and FRY were weighed by pooling the above-ground biomass and roots, respectively, and the results were averaged per plant. Starch content (% w/w) was directly measured on-site using a Reimann scale based on the specific gravity of 5 kg samples [20]. HI was calculated as the ratio of FRY to the total plant weight, expressed as FRY/(FRY + FSY).
2.4. Data Analysis
The data were analyzed using the R programming language [21], employing an augmented randomized complete block design. To estimate GCA and SCA variances, the analysis model proposed by Gardner and Eberhart (1966) in Analysis III [22] was utilized. The GCA and SCA values for each trait obtained from the diallel analysis were compared using the method described in [23]. The difference between GCA and SCA was calculated by considering the standard error and referring to the tabulated t-value. Additionally, the GCA:SCA ratio was determined by comparing the mean squares of GCA and SCA [24,25]. The Baker’s (BR) ratio was also calculated, with a value above 0.5 indicating that additive gene effects are more prominent than dominant effects [26]. However, it should be noted that this article will primarily focus on the genetic variation between families, while the within-family genetic variation will be the subject of a different article as conducted in similar studies [7,9,12,13,27,28].
3. Results
3.1. Distributions of Productivity Traits among 15 F1 Diallel Families
The diallel crosses were performed among the six cassava progenitors to generate 15 full sib (F1) families. Efforts were made to obtain as many botanical seeds as possible to properly represent within-family genetic variation (Table 2). Only healthy plants from the seedling nursery were selected for evaluating FSY, FRY, HI and SC. Unavoidably, there was considerable variation in the number of genotypes representing each F1 family. Figure 1 shows the distribution of the four traits for each cross compared to the respective progenitors using data collected from the two years. The segregations among the F1 for FSY appeared to be narrow and generally within the ranges of the progenitors (3–5 kg/plant). Segregations for FRY were mostly lower in F1 than in the progenitors, except in R5 × R90 and crosses involving HNT. The F1 distribution for HI was within the range of the progenitors. The SC of F1 showed an upward distribution and was even higher than the progenitors in many crosses (R1 × R5, R1 × HNT, R5 × R90, R5 × KU50, R5 × HNT, R90 × HB80, KU50 × HNT and HB80 × HNT). The outliers of the F1 progenies also demonstrated the potential for selecting among segregating genotypes for the four traits.
Table 2.
Numbers of F1 progenies from each of 15 crosses used for evaluation in two consecutive years (cross number, number of progenies in Year 1/Year 2).
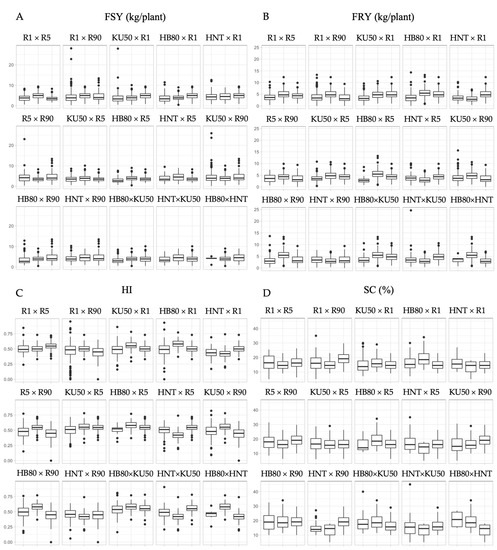
Figure 1.
Distribution fresh shoot yield (FSY) (A), fresh root yield (FRY) (B), harvest index (HI) (C) and starch content (SC) (D) for each cross compared to their respective progenitors using data collected from the two years. In each family, data are presented with F1 progenies followed by their respective parents indicated for each box.
3.2. Analysis of Variance
The genotype effect was statistically significant for the four traits. The effect of genotype × year was statistically significant for three traits, except for SC. The year effect was statistically significant only for FSY (Table 3). The weather conditions during the two years of evaluation showed major differences including a drought period and higher average temperature in the second year (Figures S1 and S2). The impact of these differences on the performance of each trait in the 15 families was analyzed (Figure 2). Despite unclear patterns in the performance of FSY and HI between the years, FRY was mostly reduced, except in crosses R1 × R5, R5 × HB80 and KU50 × HNT, which suggests the potential in these F1 families for drought tolerance. All crosses and progenitors showed an increase in SC in the second year, which could be a result of having abundant rains during the growing period (Jun–Oct) and severe drought before harvesting (Feb–Apr).
Table 3.
Analysis of variance with Gardner and Eberhart analysis III. *, **, *** indicate significant at the 0.05, 0.01 and 0.001 probability levels, respectively.
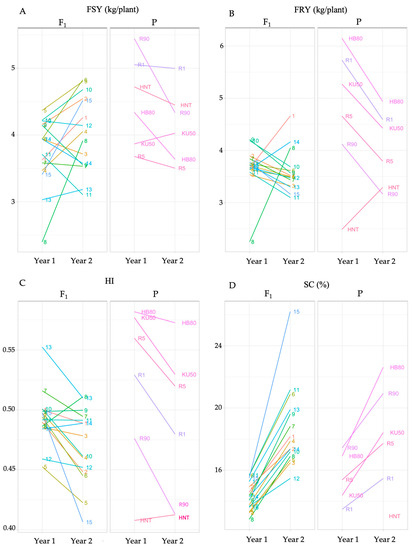
Figure 2.
Genotype–environment interaction of crosses and progenitors for FSY (A), FRY (B), HI (C) and SC (D). Mean values of the four traits from each cross and progenitor were plotted between the two years of the trial. The cross numbers are based on Table 2.
The analysis of variance conducted on FSY, FRY, HI and SC for each of the two years indicated significant genotype effects for all four traits, except for HI in the second year (Table 4). There were significant differences among progenitors for SC in both years and for HI in the first year. Interestingly, the contrasts between progenitors and F1 crosses were significant, except for FRY in the first year. There were significant differences in SC among F1 crosses (both years) and for FRY in the first year (Table 4).
Table 4.
Combining ability of the 6 × 6 diallel for fresh shoot yield (FSY), fresh root yield (FRY), harvest index (HI) and starch content (SC) in Year 1 (June 2015–May 2016) and Year 2 (June 2016–May 2017). *, **, *** indicate significant at the 0.05, 0.01 and 0.001 probability levels, respectively.
The GCA effects were significant for SC in both years (p < 0.01 and 0.001), as well as for FSY, FRY and HI in the first year (p < 0.05). The SCA effects were significant only for FRY (p < 0.01) and SC (p < 0.05) in the first year. The GCA:SCA ratios indicated that HI and SC were strongly influenced by the breeding value of the progenitors (e.g., “additive effects”), as indicated by values above one for both years, while the ratio for FSY and FRY showed inconsistency between the two years.
Based on the significant GCA effects for FSY and FRY in the first year, it can be inferred that FSY was influenced by the breeding value of the progenitors, while FRY was clearly dependent on non-additive genetic effects. GCA effects for SC, accounted for 73.1% and 67.9% of the total variation in the first and second year, respectively. Results for FSY, FRY and HI in the first year indicated that the variations in FSY and HI were mostly governed by GCA effects, while FRY was more influenced by SCA effects. Furthermore, the BR ratios of all evaluated traits were above 0.5, suggesting that these traits were primarily under the influence of additive effects rather than non-additive effects.
3.3. GCA Effects
An analysis of the GCA effects among the six parents revealed that R90 and R1 had high effects for FRY, with R1 having higher GCA effects compared to KU50, HB80 and HNT for HI. Additionally, HB80 and R5 showed similar effects for SC, but they had the higher GCA effects than the other parents (Table 5).
Table 5.
GCA effects of the 6 × 6 diallel for fresh shoot yield (FSY), fresh root yield (FRY), harvest index (HI) and starch content (SC) in two consecutive years. Columns with no significant difference among the data are presented without superscript. Among those significant, values without a common letter differ (p < 0.05).
In this study, in which only vigorous growing genotypes were pre-selected for the clonal trials, it may be desirable to have negative or lower GCA effects for FSY in order to achieve higher HI values. R1 and HB80 are potential progenitors for low FSY. Progenitor R1 showed an adequate breeding value for FRY, FSY and HI but was weak for SC. On the other hand, HB80 showed good breeding value for FSY and a higher SC. The progenies of R5 showed an above-average breeding value for SC. The significant GCA effects for SC in the second year further support the suitability of R5 and HB80 for SC.
3.4. SCA Effects
Significant SCA effects among the 15 F1 families were only observed for FRY and SC in the first year (Table 6). FSY and HI did not show significant SCA effects in either year. Notably, R5 × HNT and KU50 × HB80 had positive and higher SCA effects for FRY, and R5 × HNT had a higher SCA effect for FRY compared to other crosses. HB80 × HNT, R1 × R90 and R5 × HB80 showed positive and high SCA effects for SC. While not statistically significant, the high SCA effects of HB80 × HNT and R1 × R90 for SC in the second year may support the result observed in the first year. However, none of the above five crosses exhibited simultaneously positive and high SCA effects for both FRY and SC. Alternatively, R5 × KU50 and R90 × KU50 could be of interest due to their well-balanced SCA effects for both FRY and SC traits.
Table 6.
SCA effects of the 6 × 6 diallel for FSY, FRY, HI and SC in two consecutive years. Columns with no significant difference are without superscript. Among those significant, values without a common letter differ (p < 0.05).
4. Discussion
In this study, we evaluated the performances and combining ability in a six-progenitor diallel analysis for four productivity traits over two consecutive years. The progenitors were elite Thai varieties. The experimental design employed an augmented randomized complete block design, and the storage roots were harvested at 12 MAP. The average values of F1 for FSY and HI were generally distributed between the respective parents, while the SC averages of F1 surpassed those of the progenitors in some cases. On the other hand, the average FRY values of F1 generally showed lower performance compared to their progenitors across all crosses. This outcome is not unexpected since the progenitors were selected for their exceptional performance over the years, while each F1 family represents a random sample of all potential genotypes that could result from a particular cross. Nevertheless, outstanding individual genotypes were observed for each of the four traits, providing opportunities to select desirable progenies.
The analysis of combining ability in our study confirmed that non-additive SCA effects were relatively more important for the FRY trait, while GCA (breeding value) effects were prevalent for FSY, HI and SC. The significance of non-additive genetic effects for FRY in cassava has been well documented in previous studies summarized by Ceballos et al. [5]. Numerous diallel analysis studies [7,9,12,13,27,28] and combining ability analyses have consistently demonstrated higher ratios of SCA to GCA for FRY [8,10,11,14,29,30,31,32,33,34,35,36]. Epistasis effects, which involve the interaction of multiple genes, have also been shown to have a profound impact on FRY [5]. Our study aligns with this consensus, as we observed a GCA:SCA ratio below 1.0 and a higher sum of squares of SCA compared to GCA. Our study, using locally developed germplasm grown under subtropical conditions in Southeast Asia, further supports this notion from studies conducted in South America or Africa. This highlights the importance of considering specific families or approved cross pairs for successful selection of high FRY. Furthermore, the relevance of non-additive genetic effects for FRY also emerged from ongoing efforts to implement genomic selection in cassava breeding [37].
Previous studies have consistently indicated that GCA effects play a more significant role than SCA effects for HI [7,10,14,28,36] and FSY [10]. Although our results align with this conclusion, the mode of inheritance for HI and FSY is rather inconsistent, as SCA effects were reported to be more important for these traits in some trial locations [11,33,36]. In contrast, our two-year combining ability analysis strongly indicates that SC is influenced by additive effects. To select progenies with desirable FSY, HI and SC, one can test parents with desirable GCA effects for each trait. For example, HB80 and R5 can be utilized for high SC; R1 and HB80 for low FSY; and R1 for high HI. The substantial genetic gains achieved for SC over the years in Southeast Asia provide strong evidence for the influence of additive effects for this trait [38,39].
Cultivation conditions may affect the evaluation of genetic effects. Poor growing conditions or unsuitable environments may result in lower SCA effects and a closer resemblance to GCA effects [10]. The study by Calle et al. [28] also demonstrated a strong genotype–environment interaction for FRY in cassava. In our study, we observed notable differences between the two years of evaluation. Specifically, in the second year, there was a prolonged dry period and a decrease in FRY. These conditions resulted in no significance in SCA and an almost equal value of GCA. This disparity between the two years could be primarily attributed to differences in rainfall patterns during the critical period for starch deposition in the storage root. The poor performance in root yield under such conditions may impact the expression of the genotype.
Cassava breeding involves making direct crosses between desirable varieties, followed by clonal propagation of selected genotypes through stem cuttings. As a result, inter-varietal crossing results in a huge genetic segregation that requires large numbers of individuals to isolate exceptional genotypes. The presence of extensive within-family genetic variation further weakens the predictive value of GCA [3]. In addition, the evaluation of traits such as FRY, SC, HI, dry matter content and those related to diseases and environmental changes requires long-term clonal evaluation, further prolonging the breeding and selection schemes. Some of the parental lines used in this study have been partly evaluated by [40], which reported that more than 40% of selected hybrids are coming from crosses between R90, KU50, R60 and R5, demonstrating their potential as good breeding parents.
Determining the combining ability of parents through mating design analysis is a valuable approach that enhances the likelihood of systematically generating high-yielding breeding lines while minimizing the number of unsuccessful crosses. This study provides insights into the combining ability of six Thai cassava elite varieties for breeding purposes. The findings of this study emphasize the potential for improving cassava productivity through strategic parent selection based on desirable traits, focusing on traits such as FRY as well as FSY and HI for optimal root production and SC for the highest value of root products. However, obtaining an individual with multiple desired traits such as disease resistance and environmental adaptability remains challenging.
The difference in rainfall during the two-year trial allowed analyzing the impact of drought during the 9-11 MAP on the four traits. Crosses such as R1 × R5, R5 × HB80 and KU50 × HNT were identified as potential sources of drought tolerance based on the FRY result in the second year under drought conditions. R1 and HB80 have been identified as general combiners for FSY; R1 as a general combiner for HI and HB80; and R5 as a general combiner for SC.
The findings of this study have important implications for cassava breeding programs, providing valuable insights into the selection of parental lines. By understanding the genetic control of yield and related traits in Thai cassava germplasm, breeders can make informed decisions when designing breeding strategies and selecting suitable parents to improve cassava varieties. These insights contribute to the advancement of cassava breeding efforts, ultimately leading to the development of high-yielding and superior cassava cultivars.
In summary, this study assessed the combining ability of six Thai cassava varieties for FRY, FSY, HI and SC in a diallel-mating design over two years. The results demonstrate that non-additive SCA effects play a crucial role in determining FRY, while GCA effects are predominant for FSY, HI and SC. The study also highlights the influence of growing conditions on genetic effects and the need to consider specific crossing pairs for a successful selection of high FRY. The findings provide insights for selecting parental lines in cassava breeding programs and understanding the genetic control of yield and related traits in Thai cassava germplasm.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy13061546/s1: Figure S1: Average monthly rainfall at The Thai Tapioca Development Institute from June 2015 to May 2017; Figure S2: Average monthly temperature of the northeast Thailand data collected from June 2015 to May 2017; Table S1: Soil analysis of the plantation at The Thai Tapioca Development Institute.
Author Contributions
Conceptualization, P.Y. and P.K.; methodology, C.J., E.K. and P.K.; formal analysis, P.Y., S.V., P.W., H.C. and P.K.; resources, C.J.; figure preparation, S.V. and P.W.; writing, P.Y., S.V., H.C. and P.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Kasetsart University Research and Development Institute (KURDI), Office of Higher Education Commission, Ministry of Education (OHEC) and Center for Advanced Studies of Agriculture and Food (CASAF). PY was supported by The Graduate School, Kasetsart University. PK was supported by National Science and Technology Development Agency (NSTDA) Thailand: P1852170. SV was supported by the National Research Council of Thailand (NRCT): NRCT5-RSA63002-02. HC and SV were supported by the Office of the Ministry of Higher Education, Science, Research and Innovation and the Thailand Science Research and Innovation through the Kasetsart University Reinventing University Program 2021.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Thai Tapioca Development Institute (TTDI) for locations and help with the yield trial study. We are thankful for the support from the Kasetsart University Research and Development Institute (KURDI) and Center for Advanced Studies of Agriculture and Food (CASAF).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, S.; Cui, Y.; Zhou, Y.; Luo, Z.; Liu, J.; Zhao, M. The industrial applications of cassava: Current status, opportunities and prospects. J. Sci. Food Agric. 2017, 97, 2282–2290. [Google Scholar] [CrossRef] [PubMed]
- Parmar, A.; Sturm, B.; Hensel, O. Crops that feed the world: Production and improvement of cassava for food, feed, and industrial uses. Food Secur. 2017, 9, 907–927. [Google Scholar] [CrossRef]
- Ceballos, H.; Rojanaridpiched, C.; Phumichai, C.; Kittipadakul, P.; Iglesias, C.; Gracen, V.E. Excellence in cassava breeding: Perspectives for the future. Crop Breed Genet. Genom. 2020, 2, e200008. [Google Scholar] [CrossRef]
- Malik, A.I.; Kongsil, P.; Nguyễn, V.A.; Ou, W.; Srean, P.; López-Lavalle, L.A.B.; Utsumi, Y.; Lu, C.; Kittipadakul, P.; Nguyễn, H.H.; et al. Cassava breeding and agronomy in Asia: 50 years of history and future directions. Breed. Sci. 2020, 70, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, H.; Kawuki, R.S.; Gracen, V.E.; Yencho, G.C.; Hershey, C.H. Conventional breeding, marker assisted selection, genomic selection and inbreeding in clonally propagated crops: A case study for cassava. Theor. Appl. Genet. 2015, 128, 1647–1667. [Google Scholar] [CrossRef] [PubMed]
- Griffing, B. Concept of general and specific combining ability in relation to diallel crossing system. Aust. J. Exp. Biol. Med. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Cach, N.T.; Lenis, J.I.; Perez, J.C.; Morante, N.; Calle, F.; Ceballos, H. Inheritance of useful traits in cassava grown in subhumid conditions. Plant Breed. 2006, 125, 177–182. [Google Scholar] [CrossRef]
- Chalwe, A.; Melis, R.; Shanahan, P.; Chiona, M. Inheritance of resistance to cassava green mite and other useful agronomic traits in cassava grown in Zambia. Euphytica 2015, 205, 103–119. [Google Scholar] [CrossRef]
- Jaramillo, G.; Morante, N.; Perez, J.C.; Calle, F.; Ceballos, H.; Arias, B.; Bellotti, A.C. Diallel analysis in cassava adapted to the midaltitude valleys environment. Crop Sci. 2005, 45, 1325–1335. [Google Scholar] [CrossRef]
- Kulembeka, H.P.; Ferguson, M.; Herselman, L.; Kanju, E.; Mkamilo, G.; Masumba, E.; Fregene, M.; Labuschagne, M.T. Diallel analysis of field resistance to brown streak disease in cassava (Manihot esculenta Crantz) landraces from Tanzania. Euphytica 2012, 187, 277–288. [Google Scholar] [CrossRef]
- Parkes, E.Y.; Fregene, M.; Dixon, A.; Peprah, B.B.; Labuschagne, M.T. Combining ability of cassava genotypes for cassava mosaic disease and cassava bacterial blight, yield and its related components in two ecological zones in Ghana. Euphytica 2013, 194, 13–24. [Google Scholar] [CrossRef]
- Pérez, J.C.; Ceballos, H.; Calle, F.; Morante, N.; Gaitan, W.; Liano, G.; Alvarez, E. Within-family genetic variation and epistasis in cassava (Manihot esculenta Crantz) adapted to the acid-soils environment. Euphytica 2005, 145, 77–85. [Google Scholar] [CrossRef]
- Pérez, J.C.; Ceballos, H.; Jaramillo, G.; Morante, N.; Calle, F.; Arias, B.; Bellotti, A.C. Epistasis in cassava adapted to mid-altitude valley environments. Crop Sci. 2005, 45, 1491–1496. [Google Scholar] [CrossRef]
- Zacarias, A.M.; Labuschagne, M.T. Diallel analysis of cassava brown streak disease, yield and yield-related characteristics in Mozambique. Euphytica 2010, 176, 309–320. [Google Scholar] [CrossRef]
- Kittipadakul, P.; Kongsil, P.; Phumichai, C.; Jansky, S.H. Chapter 7: Breeding cassava for higher yield. In Achieving Sustainable Cultivation of Cassava Volume 2: Genetics, Breeding, Pests, and Diseases; Hershey, C., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2017; pp. 139–170. [Google Scholar]
- Kawano, K.; Sarakarn, S.; Limsila, A.; Tongglum, A.; Suparhan, D. Cassava cultivar evolution viewed through harvest index and biomass production. In Proceedings of the 8th Symposium of the International Society for Tropical Root Crops, Bangkok, Thailand, 30 October–5 November 1988; Howeler, R.H., Ed.; International Society for Tropical Root Crops: Nairobi, Kenya, 1990; pp. 202–221. [Google Scholar]
- Kittipadakul, P.; Vichukit, V.; Rodjanaridpiched, C.; Phumichai, C.; Kongsila, P.; Chanthaworn, J.; Boonma, S.; Jansky, S.H. “Huay Bong 80” a New Variety with High Yield and High Stability for Starch Content. In Proceedings of the Ninth Regional Workshop, Nanning, China, 27 November–3 December 2009; pp. 89–95. [Google Scholar]
- Kawano, K. Cassava. In Hybridization of Crop Plants; Walter, F., Henry, H., Eds.; American Society of Agronomy and Crop Science Society of America: Madison, WI, USA, 1980; pp. 225–233. [Google Scholar]
- Federer, W.T. Augmented (or hoonuiaku) design. Hawaii. Plant. Rec. 1956, 55, 191–208. [Google Scholar]
- Wholey, D.W.; Booth, R.H. A comparison of simple methods for estimating starch content of cassava roots. J. Sci. Food Agric. 1979, 30, 158–164. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org/ (accessed on 5 April 2023).
- Gardner, C.O.; Eberhart, S.A. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 1966, 22, 439–452. [Google Scholar] [CrossRef]
- Singh, R.K.; Chaudhary, B.D. Biometrical Methods in Quantitative Genetic Analysis; Kalyani: Ludhiana, India, 1977. [Google Scholar]
- Beil, G.; Atkins, R.E. Estimates of general and specific combining ability in F1 hybrids for grain yield and its components in grain sorghum, Sorghum vulgare pers. 1. Crop Sci. 1967, 7, 225–228. [Google Scholar] [CrossRef]
- Haussmann, B.I.G.; Obilana, A.B.; Ayiecho, P.O.; Blum, A.; Schipprack, W.; Geiger, H.H. Quantitative-genetic parameters of sorghum [Sorghum bicolor (L.) Moench] grown in semi-arid areas of Kenya. Euphytica 1999, 105, 109–118. [Google Scholar] [CrossRef]
- Baker, R.J. Issues in diallel analysis. Crop Sci. 1978, 18, 533–536. [Google Scholar] [CrossRef]
- Cach, N.T.; Pérez, J.C.; Lenis, J.I.; Calle, F.; Morante, N.; Ceballos, H. Epistasis in the expression of relevant traits in cassava (Manihot esculenta Crantz) for subhumid conditions. J. Hered. 2005, 96, 586–592. [Google Scholar] [CrossRef]
- Calle, F.; Perez, J.C.; Gaitan, W.; Morante, N.; Ceballos, H.; Lhano, G.; Alvarez, E. Diallel inheritance of relevant traits in cassava (Manihot esculenta Crantz) adapted to acid-soil savannas. Euphytica 2005, 144, 177–186. [Google Scholar] [CrossRef]
- Bueno, A. Estimation of genetic parameters in cassava. In Mejoramiento Genético de la Yuca en América Latina; Centro Internacional de Agricultura Tropical (CIAT): Cali, CO, USA, 1991; pp. 197–220. [Google Scholar]
- Chipeta, M.M.; Bokosi, J.M.; Saka, V.W.; Benesi, I.R. Combining ability and mode of gene action in cassava for resistance to cassava green mite and cassava mealy bug in Malawi. J. Plant Breed. Crop Sci. 2013, 5, 195–202. [Google Scholar]
- Lokko, Y.; Dixon, A.G.; Offei, S.K.; Danquah, B.Y. Combining ability analysis of resistance to mosaic virus disease in cassava. Afr. Crop Sci. J. 2006, 14, 221–230. [Google Scholar]
- Owolade, O.F.; Dixon, A.G.O.; Adeoti, A.Y.A. Diallel Analysis of Cassava Genotypes to Anthracnose Disease. World J. Agric. Sci. 2006, 2, 98–104. [Google Scholar]
- Kamau, J.; Melis, R.; Laing, M.; Derera, J.; Shanahan, P.; Ngugi, E. Combining the yield ability and secondary traits of selected cassava genotypes in the semi-arid areas of Eastern Kenya. J. Plant Breed. Crop Sci. 2010, 2, 181–191. [Google Scholar]
- Were, W.V.; Shanahan, P.; Melis, R.; Omari, O.O. Gene action controlling farmer preferred traits in cassava varieties adapted to mid-altitude tropical climatic conditions of western Kenya. Field Crops Res. 2012, 133, 113–118. [Google Scholar] [CrossRef]
- Peninah, N.; Richard, E.; Joseph, K. Combining ability for beta-carotene and important quantitative traits in a cassava F1 population. J. Plant Breed. Crop Sci. 2014, 6, 24–30. [Google Scholar]
- Esuma, W.; Kawuki, R.; Herselman, L.; Labuschagne, M.; Ferguson, M.E. Identification of genomic regions associated with cassava resistance to green mite infestation and whitefly in Uganda using genotyping-by-sequencing. Front. Plant Sci. 2016, 7, 1786. [Google Scholar]
- Wolfe, M.D.; Kulakow, P.; Rabbi, I.Y.; Jannink, J.L. Marker-based estimates reveal significant nonadditive effects in clonally propagated cassava (Manihot esculenta): Implications for the prediction of total genetic value and the selection of varieties. G3 2016, 6, 3497. [Google Scholar] [CrossRef]
- Kawano, K.; Narintaraporn, P.; Narintaraporn, S.; Sarakarn, S.; Limsila, A.; Limsila, J.; Suphaharn, D.; Sarawat, V.; Wattananonta, W. Yield improvement in a multistage breeding program for cassava. Crop Sci. 1998, 38, 325–332. [Google Scholar] [CrossRef]
- Kawano, K.; Cock, J.H. Breeding cassava for underprivileged: Institutional, socio-economic and biological factors for success. J. Crop Improv. 2005, 14, 197–219. [Google Scholar] [CrossRef]
- Ceballos, H.; Fregene, M.; Perez, J.C.; Morante, N.; Calle, F. Chapter 12 Cassava genetic improvement. In Breeding Major Food Staples; Kang, M.S., Priyadarshan, P.M., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 365–391. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).