1. Introduction
Modern agriculture faces the enormous challenge of feeding a rapidly growing population without further damaging the environment [
1,
2]. The necessary increase in agricultural production can be effectively supported by conventional agrochemicals (although not exclusively), mainly synthetic fertilizers and pesticides; however, at the same time, these products are accused of having particularly harmful environmental and health effects [
3]. In this context, all non-chemical strategies that support an increase in food production are gaining or re-gaining attention.
Cereals are the most important source of food for humans and animals all over the world. They provide dietary energy, essential proteins and micronutrients, and a diverse array of non-nutrient bioactive components [
4]. Despite the raised debates over the role of cereals in the human diet, they will continue to be crucial in ensuring the food security in the world, especially in the Global South [
5]. The world’s top three cereals include maize, wheat, and rice, which comprise 38.8%, 25.4%, and 25.3% of global cereal production, respectively [
6]. These are followed by barley (5.24%), sorghum (1.96%), millet (1.02%), oats (0.84%), and triticale (0.51%), which recently overtook rye (0.50%).
Triticale (
x Triticosecale Wittm. ex A. Camus) is a human-made species developed via crossing wheat (
Triticum spp.) and rye (
Secale cereale L.). It combines favorable alleles from both progenitor species (the hardiness and nutrient use efficiency of rye and the high grain yield and nutritional qualities of wheat), which not only adapts it to environments that are less favorable for wheat but also provides its higher biomass yield and forage quality [
7]. It is claimed that triticale can be used as an alternative to other cereals, mostly wheat (
Triticum aestivum L.), in livestock feed production and has the potential to become the preferred industrial energy crop [
8]. Although triticale is a relatively new crop species, there has been growing interest in it since 1975 (when it was reported for the first time by the FAO), especially in Europe, where 93.6% of the global production of this cereal currently occurs [
6]. According to data from 2020, Poland is the leading producer of triticale (6.08 million tons per year), followed by Germany (2.04), Belarus (1.54), and France (1.20) [
6]. In Poland, triticale is currently the third most important cereal, following wheat and corn, and its share in the national cereal balance is 17.4% [
6]. Since the beginning of triticale breeding, a large number of cultivars have been developed with differences in agronomic traits including grain and straw yield, harvest index, early vigor, tillers per plant, plant height, earliness of maturity, number of grains per spike, grain weight, grain quality, nutrient and water use efficiency, and tolerance to various stresses [
9]. Breeding efforts to obtain genotypes with highly beneficial trait sets continue unabated [
10]. “The Common catalogue of varieties of agricultural plant species” (with the first supplement) currently contains 418 triticale cultivars [
11]. In the Polish National Register of Cultivars, there are 67 cultivars of triticale, of which 51 are cultivars of winter triticale and 16 are cultivars of spring triticale [
12]. In Poland, winter triticale is more popular than spring triticale, mainly because of its higher yield potential [
13]. A wide array of cultivars allows farmers to choose the right cultivar for the environmental conditions and management practices in place, increasing the chance of realizing the full yield potential of triticale.
Since the potential to increase crop acreage is limited, cereal production based on increasing yields and reducing losses due to abiotic and biotic factors will be necessary to address global food security issues [
14].
Yield losses due to weeds are of critical concern in cereal-producing areas [
15]. They were previously estimated at 32% (with a range of 26–40%), exceeding those due to pests (18%) and pathogens (15%) [
16]. However, left uncontrolled, weeds can result in yield losses of even 100% [
17]. They compete with crops for sunlight, water, nutrients, and space. In addition, they harbor insects and pathogens, which attack crop plants. Weed harmfulness depends on several factors, including weed emergence time, weed density, the species composition of the weed community, and weed and crop species competitiveness [
17,
18]. The demonstration of competitive abilities against weeds is a species and cultivar attribute of cereals [
18,
19]. Winter cereals are generally more effective at inhibiting weed growth than spring cereals [
20,
21]. Among winter cereals, rye is the most competitive against weeds [
20], while the competitive ability of triticale falls between its progenitor species [
21,
22]. The literature has reported a large number of relationships between competitive ability and plant traits, including plant height, development rate, canopy architecture, and resource partitioning [
23]. In general, the vigorous growth habit of triticale and its leafiness and height allow triticale to compete well with weeds [
10]; however, competitiveness varies widely between cultivars [
23]. It was proved that the competitive ability of tall triticale cultivars can parallel that of rye [
21,
22]. Moreover, the importance of traits related to cultivar competitiveness may vary between years [
23]. Competitive cultivars can be used in weed management strategies as an inexpensive non-chemical option [
23].
Crop rotation has been used as an effective tool for weed management since the distant past [
24]. The diversity of the crop species in the rotation and their proper sequencing are critical [
25]. A rotation system involving three or more crops is often referred to as a diversified crop rotation (DCR) [
26]. In a well-designed DCR, an appropriate sequence of crops with varying patterns of resource competition, allelopathic interference, and soil disturbance and mechanical damage caused by crop-associated operations provides weeds with an unstable and often inhospitable environment that effectively prevents the proliferation of weed species [
27]. These factors support the regulation of weed populations. However, the role of crop rotation in weed control has depreciated significantly in the 20th century due to the easy access to synthetic inputs [
24]. With the introduction of selective herbicides in the late 1940s and a constant influx of new active ingredients in the succeeding decades, farmers were provided with a new tool for weed control independent of the crop production system [
28]. So, over the past half-century, crop rotations have become increasingly simplified, frequently producing only one or two crops in succession [
29]. The simplification of crop rotations results in the repeated exposure of weeds to the same set of ecological and agronomic conditions. This can result in increased abundances of several competitive, highly adapted weed species in crops and promote the development of herbicide resistance [
28,
29,
30].
At present, agricultural scientists are advocating for a return to crop rotation, stressing its productive and environmental functions [
24]. The implementation of diversified crop rotation (DCR) is recommended on many pathways toward sustainable agriculture [
2]. DCR is indicated as a non-chemical agricultural practice [
31] that enables farmers to reduce their reliance on external inputs [
32], including herbicides [
29].
In addition to a new perspective on the old practice, it is worth emphasizing the change in the approach to weeds that has taken place recently. Weed species have begun to be seen as an element of biodiversity, and the important ecosystem services provided by this type of vegetation have been appreciated. Primarily, the key trophic and paratrophic functions of weeds in the agroecosystem have been noticed [
30,
33]. Furthermore, the importance of weed diversity in mitigating yield losses has been acknowledged. Adeux et al. [
34] demonstrated that diversified weed communities can limit the negative impact of competitive and dominant species on the productivity of the crop. Opinions have emerged that weeds have an underappreciated value to biodiversity [
35]. Thus, the progressive loss of weed diversity due to agricultural intensification has begun to be viewed with concern [
36]. Not only has a significant decrease in the species diversity of weeds been recorded [
36] but reductions in their taxonomic and functional diversities have also been recorded [
37].
The species diversity of weeds is most often expressed as the number of species (species richness) or via selected diversity indexes (mostly Shannon–Wiener or Simpson) that additionally include weed species abundance (density and biomass) [
38,
39,
40,
41]. These measures of species diversity have their limitations. Species richness treats all species as if they are equally abundant, and other single indicators may be more sensitive to abundant or rare species [
42]. A more comprehensive view and comparison of the species diversity of weed communities can be obtained using index families, such as Renyi diversity profiles [
39,
42]. According to this concept, a community can be regarded as more diverse if its diversity at all values of the scaling parameter (α) is higher than those of other communities. An important limitation of the species-based approach to weed diversity is that it treats all species found in the community equivalently, no matter their taxonomic affinity (taxonomic relationships among species) or role in the ecosystem [
43]. The latter two aspects fall under the taxonomic and functional dimensions of biodiversity, respectively. Taxonomic diversity refers to the representation of lower-rank taxa within higher-rank taxa, and it can be a measure of the stress exerted on weed communities by various environmental factors since the taxonomic spectrum of organism communities is often reduced when disturbances occur [
44]. At the extreme, weed communities may contain only closely related species, even species of the same genus [
44]. Functional diversity is understood through the concept of ecological niche and is based on the organisms’ traits (morphological, anatomical, physiological, reproductive, or behavioral) that directly or indirectly affect the functioning of the ecosystem [
45]. Species with a similar impact on a particular process (or more than one process) in an ecosystem (effect traits) or those that present similar responses to environmental factors (response traits) are grouped into functional groups [
46]. Making an informed choice of traits based on ecological functions is crucial. The functional structure of the weed community governs the processes of competition for resources or complementarity in their use in space and time [
34]. Competitive weeds have attributes related to rapid resource acquisition (i.e., high seed mass, high height, high specific leaf area, high leaf nitrogen content, and the same phenology as the crop). When the weed community occupies the same niche as the crop, weed–crop competition is the most intense. A higher level of functional diversity within the weed community is expected to promote complementarity in resource utilization, thereby reducing the probability of intensive niche overlap with the crop and yield losses due to dominant and competitive weeds. The shift in focus from weed taxonomy to function has yielded successful predictions of competitive outcomes; hence, a functional group approach rather than a single-species approach has been recommended in designing weed regulation [
47,
48].
Modern, sustainable weed management strategies are expected to minimize weed competitiveness while promoting weed diversity, i.e., keeping beneficial species at acceptable levels [
48,
49]. The role of DCR in weed control is evident [
24,
50]. However, DCR alone may not be sufficient to achieve optimal weed control, and other practices may need to be used in conjunction, such as applying herbicides or introducing more competitive crop genotypes [
29].
Previous long-term studies by Zawiślak [
20] proved the redundancy of herbicide use in the cultivation of winter rye cultivation under DCR. A strong root system of rye, capable of intensive water and nutrient uptake followed by a fast-growing and rapidly closing canopy, made this agrophytocenosis unfavorable for weeds. Thus, there was no yield threat from weed competition. In addition, the application of a herbicide under these conditions resulted in a reduction in rye yield, demonstrating the phytotoxic effect of the herbicide. In turn, according to a previous study by Jastrzębska et al. [
51], DCR was sufficient to provide a satisfactory winter rye yield and reduce the abundance of weeds without decreases in weed species diversity or functional diversity. Moreover, when rye was grown under DCR conditions, the application of a herbicide did not increase rye yield, but it was harmful to weed biodiversity. Since the growth behavior and competitive ability of some triticale cultivars are similar to those of rye [
19,
21,
22], it can be assumed that there is no need for herbicides when growing triticale in a DCR system, especially if the appropriate cultivar is selected for cultivation. This work focused on determining (1) whether herbicide application in triticale grown in a DCR system will result in an increase in yield due to weed biomass reduction or whether its application will no longer be necessary, (2) how the herbicide will affect the species, taxonomic, and functional diversities of weeds, and (3) how the choice of cultivar is important in the above issues. It was expected that growing a high-yield and competitive cultivar of triticale in a DCR system without herbicide support would maintain the weed biomass at a low level that would not compromise the triticale yield and weed diversity. In addition, the relationships between weed diversity, weed biomass, and triticale yield were investigated.
4. Discussion
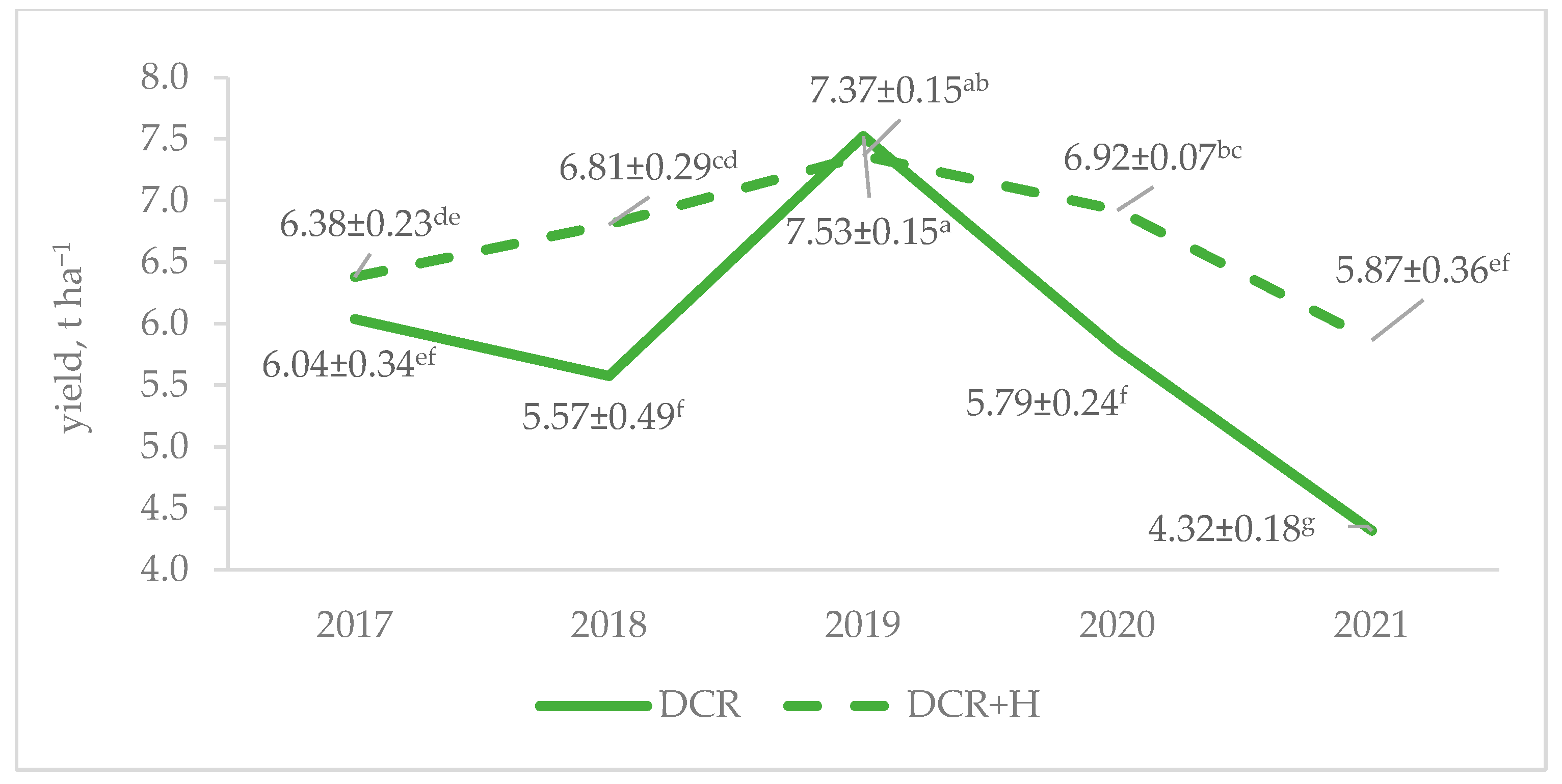
Regardless of the cultivar and year, the DCR + H strategy resulted in a higher triticale yield compared to the DCR strategy, a difference that was attributed to its effective weed biomass reduction. At the same time, however, the DCR + H strategy reduced weed diversity.
The superior efficacy of the DCR + H strategy over DCR in reducing weed biomass was expected, given the results of previous studies [
51,
66]. Although it was assumed that there would be no increase in triticale yield due to herbicide application, the observed increase is also not surprising considering the studies by other authors [
67,
68,
69]. The higher yield achieved under the DCR + H strategy was associated with increases in the spike density and grain number per spike, indicating an increase in resource availability to the triticale crop once weed competition was reduced by the herbicide [
48]. Other authors have also found an increase in 1000-grain weight under such conditions [
67]. However, in the present study, the opposite effect was observed, which may be explained by compensation mechanisms between the different yield components. Often, yield components are negatively correlated, with a higher spike density resulting in fewer and/or smaller grains per individual spike [
70]. This is the response of plants to light conditions modified by the spike density.
Over the years, the Borowik cultivar consistently showed a higher yield than the Trapero cultivar, regardless of the weed management strategy used. The generally lower number of productive tillers of the former was effectively compensated for by the higher grain number per spike and the greater 1000-grain weight. Genetic differences among triticale cultivars with respect to yield components are well documented in the literature [
18,
69,
70,
71,
72,
73,
74], and the Borowik cultivar has been noted for its particularly high 1000-grain weight [
18].
The yield response of triticale to the weed management strategy was cultivar-dependent. This was related to the different competitiveness of the cultivars grown against weeds observed under the conditions of the DCR strategy. The competitive ability of winter triticale against weeds is a cultivar attribute. Tall cultivars with better winter hardiness and higher tillering coefficients are considered to have competitive abilities comparable to those of rye and compete well with both monocotyledonous and dicotyledonous weeds [
21,
75]. In the present study, the greater competitiveness of the Borowik cultivar against weeds in a DCR system strategy was explained by its faster growth rate after the spring vegetation restart than that of the Trapero cultivar, as well as by its larger leaves and longer tillers, although they are slightly less numerous. Early season crop ground cover confers later competitiveness against weeds [
76]. The high competitive ability of the Borowik cultivar under organic cropping conditions was confirmed by the study of Feledyn-Szewczyk et al. [
18]. When weeds are controlled with herbicides, differences in weed biomass between cultivars or years tend to blur [
77]. Such an effect was noted in the present study. The less-competitive and lower-yielding cultivar, Trapero, responded to the reduction in weed biomass caused by the herbicide (DCR + H strategy) with a greater increase in its yield. Most of all, the reduction in weed competition allowed this cultivar to produce more spikes, which was not observed in the case of the Borowik cultivar. However, the yield of Trapero in a DCR + H system barely matched that of Borowik in a DCR system.
The inter-annual variability peculiarly determined the triticale yield and weed biomass and interacted with weed management strategy in shaping these variables.
The reduction in weed biomass under the conditions of the DCR + H strategy was consistent across years with the exception of 2017, when there were no differences between the DCR and DCR + H strategies. It was the first year of a new crop rotation cycle and the growth of the Trapero and Borowik cultivars. Cultivar rotation has been previously suggested as a tool for weed control strategies in crop fields [
78]. Moreover, in 2017, the herbicide proved to be less effective in controlling
Agropyron repens, and by limiting other weed species, it provided more space for the latter’s biomass to develop. The lack of a difference in triticale yield between the DCR and DCR + H strategies this year seems to be a natural consequence of the statistically equal weed biomass.
Regardless of the weed management strategy, 2019 proved to be the most favorable year for productivity for both cultivars over the five-year study period. This year, after a February and March that were much warmer than usual (
Table S1), the vegetation started unusually early, and triticale took advantage of these conditions for intensive tillering [
79]. A high spike density was the most important yield-determining component in 2019. The grain number per spike and 1000-grain weight, although they demonstrated the lowest values of the five years, were high enough to not contribute to a reduction in the yield of triticale. An early spring restart and increased tillering allowed the triticale to escape weed competition under the DCR strategy. Although the higher spike density in 2019 was followed by a significant reduction in grain numbers per spike and the 1000-grain weight, the triticale yield under the DCR strategy was not lower than the triticale yield under the DCR + H strategy that year. On the other hand, Brzozowska and Brzozowski [
67] claim that in years characterized by an even distribution of precipitation, the competitiveness of triticale against weeds is enhanced, and the effect of herbicide application becomes insignificant. It is noteworthy that in 2020, after a warm winter and an early spring, the yield of triticale under the DCR strategy decreased significantly, although the weed biomass was not higher than in 2019. This can be explained by a more complementary resource use by crops and weeds in 2019 but a more intense niche overlap and competition in 2020 [
34]. The Trapero cultivar made better use of the early spring in 2019 and 2020 for tillering than the Borowik cultivar, but due to its generally lower grain number per spike and 1000-grain weight, it did not match the productivity of the Borowik cultivar, regardless of the weed management strategy. A late spring in 2021 worsened triticale tillering, especially in the Trapero cultivar, which weeds took advantage of by building up a large biomass under the conditions of the DCR strategy. A rather wet July was not favorable for grain filling by triticale. A low spike density and relatively low 1000-grain weight in 2021 contributed to the low yield of the triticale (the lowest of the study period), while the yield increase due to the use of a herbicide (DCR + H) in that year was the highest of the study period.
The contribution of the individual yield components to the yield volume varied across years; however, for the entire study period (2017–2021), only a positive correlation between yield and spike density was confirmed. Other authors have also pointed to spike density as the most important component of triticale yield [
67]. However, the present study and other studies show that it is difficult to establish a strict ranking of the individual components of triticale yield. This is because they are determined by both genetic factors (cultivar) and environmental conditions [
67].
The reduction in weed species diversity under the DCR + H strategy is an obvious result of the long-term use of herbicides not only in triticale but also in any crop previously grown on a given plot according to the rotation cycle pattern. Each year, species sensitive to a particular herbicide have been eliminated or prevented from producing seeds. At the same time, the soil seed bank has been depleted [
80]. This has promoted a reduction in species composition and changes in weed community structures in herbicide-treated fields. The decline in weed species diversity as a result of herbicide use in conventional crop rotations has been confirmed by many other studies, which measured diversity using species richness or the Shannon–Wiener index [
41,
51,
81,
82]. Renyi profiles are considered to provide a more comprehensive picture of species diversity than individual measures (indexes) [
42]; however, they are not yet used very often in comparing weed communities [
38,
83,
84]. In the present study, Renyi profiles show that abandoning chemical weed control in triticale (DCR strategy) results in a consistently higher weed species diversity (at all values of the scaling parameter) or at least a higher number of species among which functionally valuable species can be found. In an earlier study involving this family of indexes, Jastrzębska et al. [
38] also proved a consistently higher weed diversity was found in triticale grown in a Norfolk-type rotation without herbicides (organic farming) than in the same rotation with herbicide application (integrated farming).
The declines observed in the taxonomic and functional diversities of weed communities were the results of either the complete eradication of weeds in individual plots by the herbicide or a reduction in the number of species to one or a small group of taxonomically or functionally close species. Such a direction of weed structure simplification under chemical control has been noted in the literature [
50,
85]. The simplification of weed community structure has often been linked to emerging herbicide resistance in weeds, and a decline in weed diversity has been reported from many cropping systems, while weed biomass has not decreased [
30]. Such a phenomenon was not observed in the present study as the herbicides were exchanged quite frequently during the experiment.
The number of reports on the impacts of agricultural practices on the taxonomic and functional diversities of weeds has been increasing in recent years [
37,
48,
86,
87,
88,
89]. However, there are still few studies comparable to the present study in terms of the research factors and methods used (e.g., formulas of indexes, weed abundance measures, and selection of functional traits) as various methodological approaches were used by other authors [
37,
88,
89]. A general opinion has been established that agricultural intensification reduces the taxonomic and functional diversities of weed communities [
37,
90], and herbicide use is among the indicators of this intensification at the individual field scale [
37]. The findings from the present study are broadly in line with this opinion, although in some years, the negative effects of herbicides (DCR + H strategy) on taxonomic distinctness were not proven. In a methodologically similar study with winter rye, Jastrzębska et al. [
51] found similar differential effects.
In the present study, there was no effect of cultivar on weed species richness (
S) and Shannon–Wiener species diversity (
H′), and Renyi profiles also show an inconclusive and inconsistent effect of this factor on weed species diversity across the study years. Such findings are not very surprising given the reports of other authors. Wesołowski et al. [
91] and Urban et al. [
92] showed that the
H′ values of weed communities varied depending on the cereal species and cultivar, but the differences were small. In other studies, small differences between cereal cultivars were recorded in weed species richness [
91,
93]. In the available literature, we failed to find articles on the taxonomic and functional diversities of weeds depending on crop cultivar, leaving room for further study. Currently, it is recommended that the assessment of plant community diversity should be based on a set of indicators that simultaneously capture different components of the multidimensional concept of biodiversity [
42]. The present study, which used Renyi profiles and taxonomic and functional diversity indexes to assess weed diversity, addresses this postulate.
Inter-annual variations in triticale yield and weed biomass and diversity and the interaction of these variations with the chosen weed management strategy and/or cultivar may be explained by the fact that many factors play a role in the competitive interactions between crops and weeds, which may vary from year to year [
75]. This is also why the observations carried out in long-term field experiments are of great importance. Weather conditions appear to be of primary importance. The amount and pattern of rainfall and temperatures over time not only determine crop development and yield [
67] but also strongly influence seed germination and weed plant development [
94]. The latter is due to the specific requirements of different weed species and can explain differences in both weed biomass and weed diversity [
41,
65,
94,
95].
Weed biomass is the qualitative measure of weed competitiveness, and most of the previous studies relating crop yield and weed biomass showed a negative linear function [
96]. In the present study, the negative correlation between triticale yield and weed biomass was also statistically confirmed except in 2017 and 2019. In these two years, competition from weeds proved to be a weak yield-limiting factor. It was particularly surprising in the case of 2019, in which the weed biomass in the Trapero field under the DCR strategy was even higher than in 2020. In addition,
Apera spica-venti, a species that is strongly competitive with winter cereals, accounted for 69.4% of the total weed community (
Table S5). This demonstrates that understanding the ecological processes that drive weed abundance (density and biomass) and how to influence these factors is still an open challenge in sustainable weed management [
48].
In addition to the negative correlation with weed biomass, triticale yield showed a negative correlation with at least one of the weed diversity dimensions. This relationship was particularly evident under the DCR strategy conditions, in which weather conditions played a greater role in determining weed biomass and diversity. The strong reduction in weed biomass by the DCR + H strategy to almost the same level in all years made the relationship between weed biomass and yield insignificant.
The relationship between weed community diversity and crop performance has yet to be fully elucidated [
97]. More diverse weed communities are thought to buffer agroecosystems against dominance by one or a few aggressive, resistance-prone species [
98]. On the other hand, in diverse communities, there is a greater chance that one or more species would be able to survive a given control strategy and gain a competitive advantage over the crop, lowering its productivity [
99]. The present study supports the opinion that high weed abundance provides great opportunities for weed species diversity [
100]. Moreover, it seems that it was the link between weed diversity and weed biomass that directed the relationship between weed diversity and triticale yield. Evidence of both a positive [
34,
101] and negative [
102] relationship between crop productivity and weed diversity, as well as a lack of a relationship between these variables [
51,
103], can be found in the literature. Some authors [
97,
103] claim that the relationship between weed diversity and crop productivity is a complex phenomenon and cannot be reflected adequately by the common weed crop interference models.
Finally, it should be considered optimistic that under the DCR strategy (without herbicide), the Borowik cultivar limited the weed biomass without reducing weed diversity. Although the yield of the Borowik cultivar under the DCR strategy was lower than under the DCR + H strategy on average for the 5 years of the study, the difference was only 0.5 t ha
−1, which represents a loss of 7.2% compared to the yield under the DCR + H strategy. The analogous difference for the Trapero cultivar was 1.13 t/ha, i.e., 17.7%, and the yield of Trapero under the DCR + H strategy did not exceed the yield of Borowik under the DCR strategy. Weed competition yield losses of 2.5–10% are accepted in integrated weed management programs [
104]; thus, the loss of 7.2% for Borowik can be considered acceptable. On the positive side, there is less pollution from herbicides and increased biodiversity. These are still not economically quantifiable for the farmer [
105]. However, governmental subsidies and other support programs can compensate for the decreases in yields and provide additional incentives to abandon pesticide use [
106]. Previous studies have also shown that choosing the right cultivar is one of the key factors in determining triticale productivity [
18,
68,
71,
72,
107]. The findings from the present and other studies [
18,
108] prove that the Borowik cultivar demonstrates high yield potential. This potential, in addition to its high level of competitiveness against weeds, predestines this cultivar for organic production in which DCR plays a central role and synthetic herbicides are prohibited [
18], or for other crop production systems that rely on a non-chemical approach [
109].












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}