Abstract
Among the mycoparasites, Ampelomyces strains are studied in detail, particularly regarding their use as biocontrol agents (BCAs) of powdery mildew (PM) fungi, including their potential to replace conventional agrochemicals. Ampelomyces strains are characterized morphologically; their ribosomal DNA internal transcribed spacer (rDNA-ITS) regions and actin gene (ACT) fragments were sequenced and their mycoparasitic activity was analyzed. In the interaction between Ampelomyces strains and PM fungi, the spores of the mycoparasites germinate on plant leaves, and their hyphae then penetrate the hyphae of PM fungi. Ampelomyces hyphae continue their growth internally, initiating the atrophy of PM conidiophores and eventually their complete collapse. Following the successful destruction of PM hyphae by Ampelomyces, the mycoparasite produces new intracellular pycnidia in PM conidiophores. The progeny spores released by mature pycnidia become the sources of subsequent infections of intact PM hyphae. As a result, the number of Ampelomyces-inoculated PM colonies gradually declines, and the conidial release of PM colonies is inhibited after the first treatment. Almost all conidiophores of 5- and 10-day-old Ampelomyces-inoculated PM colonies undergo complete atrophy or collapse. Methodological advances and in-depth analyses of the Ampelomyces–PM interaction were recently published. In this review, we summarize the genetic and phylogenetic diversity, the timing of mycoparasitism and pycnidiogenesis, the results of quantitative and visual analyses using electrostatic and digital microscopy technologies, the PM biocontrol potential of Ampelomyces, and the potential commercialization of the mycoparasites. The information provided herein can support further biocontrol and ecological studies of Ampelomyces mycoparasites.
1. Introduction
Powdery mildew (PM) is a serious disease affecting many crops [1,2]. The leaf damage caused by the fungus significantly reduces crop productivity [3,4]. While fungicides can be sprayed before or after PM colonies appear on host leaves to control the disease, frequent application of commercial fungicides can lead to resistance [5,6,7]. To avoid fungicide resistance and the environmental problems caused by fungicide residues, new control strategies that are independent of chemical methods are needed to control PM. Biological control offers an alternative method to prevent or suppress PM in crops by exploiting the antagonism between micro-organisms. Mycoparasitic fungi parasitize other fungi and they include a diverse group of parasites. These fungi absorb nutrients from their mycohosts through haustoria or other special interfaces between their cell walls and membranes. Alternatively, they invade the hyphae of their mycohosts, growing from cell to cell in the latter’s hyphae, conidia, and conidiophores while absorbing nutrients from the infection structures [8,9].
Kiss [10,11] examined all known fungal antagonists of PM, whether found in the field or tested as potential biocontrol agents (BCAs) of PM infections, including species without any record of a natural antagonistic relationship. More than 40 fungal taxa were shown to suppress the growth and sporulation of PM fungi. Of these, Aphanocladium album [12], Pseudozyma flocculosa, Moesziomyces rugulosus [13,14], Gjaerumia minor [15,16], Lecanicillium lecanii [14], and Ampelomyces quisqualis [17,18] are well known as BCAs against PM.
The mycoparasitic fungus Ampelomyces quisqualis Cesati ex Schlechtend (syn. Cicinnobolus cesatii de Bary; [17,18]) is a slow-growing pycnidial fungus widely distributed in PM colonies and naturally occurring worldwide [19,20,21], where it acts as a hyperparasite of strains infecting cultivated and wild plants [10,21,22,23]. The Ampelomyces strains produce progeny spores in mature pycnidia, which develop intracellularly in the hyphae of PM fungi in nature and then suppress mycelial growth, sporulation, and conidial germination of their mycohosts [9,10,11]. Therefore, this mycoparasitic fungus gained much attention as BCAs for controlling the PMs. The life cycle, mode of action, and biocontrol potential of hyperparasitic fungi were reviewed [9,11,24] with the aim of guiding future research in fungal and plant ecology, as well as in the development of products for the control of plant diseases [25]. However, quantitative data on the impact of hyperparasitism on host fungi are lacking. Thus, in this work, we review (1) the interactions between Ampelomyces strains and PM fungi (mycohosts) with respect to the morphological and physiological characteristics and phylogenetic placement of Ampelomyces strains; (2) the visualization and impact of fungal hyperparasitism (infection process and pycnidiogenesis) on mycohost survival by using a digital microscopic technique; (3) the quantitative impact of fungal hyperparasitism on the suppression of conidial release from PM colonies infected with Ampelomyces strains by incorporating a recent methodological advance; and (4) the practical aspects of using Ampelomyces strains as BCAs. Finally, (5) summarizing experimental results, we provide an ideal spray inoculation system for the effective use of Ampelomyces as a BCA, as well as in research.
2. Powdery Mildew Fungi
PM fungi (Erysiphaceae) are obligate biotrophic pathogens of more than 10,000 host plant species, including important crops, and are responsible for serious losses in agriculture, horticulture, and forestry [1,26,27,28]. The sporulation of many PM anamorphs is intense (Figure 1A), and the produced conidia (Figure 1B) spread rapidly [29,30]. While regularly applied fungicides are used to control PM, its frequent and inadequate use can lead to the emergence of fungicide resistance [7,31,32], as demonstrated in cucurbit PM fungi [5,6,33,34,35,36]. In addition, plant leaves retain fungicides that are not completely decomposed by microorganisms, and the fungicides may also have negative side effects on plant physiology [37] as well as biodiversity [38]. Thus, to avoid drug resistance and environmental problems, new strategies for the control of PM that are independent of chemical methods are needed.

Figure 1.
Photograph of powdery mildew (PM) disease caused by Podosphaera xanthii on melon leaves, and a micrograph of conidiophores formed in the fungal colonies. (A) Melon PM disease involving the whole leaf. (B) Melon PM conidiophores observed using a digital microscope (KH-2700 DM). The conidiophores have normal catenate conidia, forming chains. Bar: 60 µm.
3. Ampelomyces, a Genus in Need of Taxonomic Revision
Ampelomyces quisqualis was first described from the PM fungus Erysiphe necator [17]. However, de Bary [39] used the name Cicinnobolus cesatii for the same fungus instead, and the latter became the most commonly used name until the 1970s. However, based on priority, the former name is taxonomically correct [40]. Based on the presumed, but experimentally unproven specialization to host fungi, or based on the host plant of PM infected with Ampelomyces, a number of different species were described in the genus. There are more than 40 formally valid descriptions of Ampelomyces species in the literature (see [41]). However, as it seems there is no narrow host specificity (see below) in the genus, in recent decades, the name A. quisqualis was used for the fungus, hinting that the Ampelomyces genus would be monotypic. However, considerable genetic variation characterizes the genus Ampelomyces. Between each genetic group, the sequence difference in the ribosomal DNA internal transcribed spacer (rDNA-ITS) region may be as high as 19% [41,42], with even greater variability in actin (ACT) sequences [43]. Although lineages that can be separated in the genus are presumably separate species [42,44,45], formally described species do not correspond to the phylogenetic groupings obtained on the basis of DNA sequences. In addition, for some proposed Ampelomyces species there are several published names, and a taxonomic revision of the genus is accordingly necessary [42,46]. However, a polyphasic, comprehensive analysis based on colony morphology, micromorphology, and phylogeny is yet to be conducted. In this work, therefore, we do not use the formally existing species names, but rather the terms Ampelomyces species or Ampelomyces strains, as recommended [47].
4. Ampelomyces as an Ecofriendly Biocontrol Agent against PM
Ampelomyces strains were reported in association with more than 65 species (eight genera) of Erysiphaceae from across the world [12,21,22,48,49,50,51,52,53,54,55,56,57,58,59,60,61]. The interactions between mycoparasitic fungi and their mycohosts take place exclusively on the surfaces of aerial plant organs [21,24,62].
In the 19th century, mycologists clearly recognized that some fungi were parasites of PM (e.g., A. quisqualis Cesati ex Schlechtend. [17]; Byssocystis textilis Riess [63]; and Cicinnobolus cesatii [39]). The first study of A. quisqualis was carried out by De Bary [39], who identified the fungus as an intracellular parasite of Erysiphaceae. De Bary [39] also showed that Ampelomyces hyphae grow within the mycelia of PM, spreading from cell to cell through septal pores, with pycnidia produced in one or two cells of the hyphae, conidiophores, and conidia of their mycohosts. Emmons [49] later conducted an extensive cytological study, describing in detail the penetration, growth, and sporulation of Ampelomyces in the ascomata of PM. Shortly thereafter, Yarwood [64] described the treatment of PM-infected plants using a conidial suspension of Ampelomyces, the first experiment demonstrating the biocontrol of a plant pathogenic fungus. Since then, hyperparasitic fungi of the genus Ampelomyces began to be applied as BCAs against PM fungi in various crops worldwide [9,10,23,44,46,65,66], thus demonstrating their utility as an ecofriendly method of PM disease management.
There is little on the physiological, biochemical, and molecular interactions between Ampelomyces strains and their host fungi; therefore, overall, little is known about the molecular mechanism of the mycoparasitism exerted by Ampelomyces [65,67]. A few studies reported that enzymatic, and also mechanical processes play a role during penetration into PM structures. Appressorium-like structures were observed at the penetration sites [68]. Five to ten days later, the mycoparasite degrades the cytoplasm of the host [49,69]. This suggests that the interaction becomes necrotrophic at a later stage. The activity of several hydrolytic enzymes (such as chitobiases, proteases [70], β-glucosidase, β-N-acetylglucosaminidase, acid phosphatase, ribonuclease, β-1,3-glucanase, and α-1,4-glucanase [71,72]) was demonstrated in Ampelomyces strains. It was suggested [72] that Ampelomyces probably interferes with the energy metabolism and protein and cell wall synthesis of the host. Based on transcription studies, several other genes are differentially expressed during mycoparasitism, including lipases, oxygenases, and peptidases [65]. In addition to enzymatic processes, another underexplored mechanism, toxin biosynthesis, may also take place during mycoparasitism [65].
5. Morphological and Molecular Analyses, and the Identification of Ampelomyces Strains
Based on morphological and molecular phylogenetic analyses, Németh et al. [61] recently identified hyperparasites isolated from different PM species in Japan as Ampelomyces Cesati ex Schlechtend. Spores of Japanese hyperparasitic strains were produced in pycnidia, which develop intracellularly in the mycelia of PM fungi. The spores are unicellular, hyaline, ellipsoid–ovoid to doliiform (size range: 5.7–9.2 × 2.6–5.0 μm), mostly guttulate, and embedded in a mucilaginous matrix within swollen ampulliform or pyriform pycnidia [61]. The spores germinate ca. 15–20 h after their inoculation, forming elongated hyphae that branch under conditions of high relative humidity (RH). Hyphae formed from the spores reach a length of 6.2–78.2 μm 48 h after inoculation. Fungal colonies slowly and concentrically spread after the inoculation of a single mature pycnidium in the centre of Czapek–Dox agar medium supplemented with 2% malt extract. The colony area reaches 148.4–391.3 mm2 at 20 days post-inoculation (dpi). Isolates significantly differ in their germination rate and hyphal length, but not in their colony area. The strains grow slowly, with an in vitro radial growth rate of 0.5–1.0 mm day−1. Thus, the morphological and physiological characteristics of the Japanese strains clearly resemble those of A. quisqualis isolates [41,59,60,69,73].
As noted above, molecular analyses based on the rDNA-ITS region and ACT fragment revealed considerable genetic diversity among Ampelomyces strains [42,43,44,45,62,74,75]. Using sequences from these two loci, Németh et al. [61] confirmed the existence of at least five different phylogenetic lineages within the genus Ampelomyces, and showed that the newly isolated Japanese strains belong to three major clades. The authors analyzed the phenotypic characteristics of Ampelomyces strains isolated from four different PM samples, and four different strains isolated from the same PM sample. There were no morphological characteristics that could clearly be associated with a given genotype or clade. The four strains isolated from the same PM sample, however, differed significantly in their measured hyphal lengths, germination rates, and the number of spores that developed in single pycnidia, as well as strong evidence of strain-level differences, as reported in other studies [46,70]. Whether the differences in the phenotypic characteristics of different strains of Ampelomyces are related to an as-yet unrevealed genetic diversity or are simply caused by phenotypic plasticity is currently unknown. The possibility of strain-level differences, however, needs to be considered in studies aimed at the development of Ampelomyces as BCAs.
6. Ampelomyces Strains May Be Associated with, but Are Not Specific to, Their Host PM Species
The specificity of Ampelomyces was investigated using two fundamentally different approaches: by isolating Ampelomyces from a diverse range of PM fungi and then investigating possible associations between the interacting partners and via cross-inoculation experiments.
“Some degree of mycohost specialization” and “evidence for narrow host specialization” were reported for Ampelomyces based on the genetic clustering of strains according to the mycohost [44,75,76]. However, other studies that employed a similar methodology obtained different results. Several Ampelomyces strains, all isolated from grapevine PM naturally infected by Ampelomyces, belong to four different genetic clades [43]. After a similar sampling, Ampelomyces strains isolated solely from Arthrocladiella mougeotii were assigned to three different clades [77]. These studies suggest that Ampelomyces strains isolated from a given PM fungal species can belong to genetically different groups, and isolates from different host fungi can belong to the same genetic group [41,42,43,44,47,77,78]. Taken together, these results support the lack of host specificity of Ampelomyces.
Host specificity was also experimentally investigated in other studies. In cross-inoculation experiments carried out by De Bary [39], Ampelomyces mycoparasites collected from a given PM species were shown to also produce intracellular pycnidia in the mycelia of other PM species. In other cross-inoculation experiments, including in vitro studies [42,70,74,79] and field experiments [46,77,80] involving different Ampelomyces strains and several PM species, these mycoparasitic strains did not show strict host specificity; instead, they were capable of infecting many host species irrespective of the original host, producing intracellular pycnidia in the mycelia of other species of Erysiphaceae [39,42,46,47,70,74,77,79,80,81]. Following inoculation tests with Japanese isolates and five PM species, Németh et al. [61] observed the degeneration and constriction of parasitized hyphae of all five PM species tested, as well as pycnidial formation in the hyphae and conidiophores of four PM fungi. These results show that Japanese Ampelomyces strains can infect PM hyphae irrespective of the original host, as they produced intracellular pycnidia in the mycelia of four out of the five tested mycohosts. Additional experiments showed that Ampelomyces strains from apple PM naturally infect Golovinomyces orontii (s. l.), the tobacco PM fungus, and P. xanthii causing cucumber PM [77]. These results and those of several other studies [39,70,74,77,82] support the lack of host specificity with the tested Ampelomyces strains.
Seemingly contradictory results were obtained with B. graminis. A previous study reported the lack of pycnidial production of a strain isolated from E. artemisiae in B. graminis on barley [60], which is similar to the findings of Németh et al. [61]. Other studies reported typical mycoparasitism, including the formation of intracellular pycnidia, in B. graminis conidiophores on cereals (wheat and barley) by Ampelomyces strains isolated from PM infecting dicots [59,62,79,83,84]. The contradictory results might be due to unfavourable experimental conditions, as described by Kiss [10], and not to the inability of Ampelomyces strains to infect B. graminis. It should be noted, however, that Ampelomyces strains seem to parasitize PM fungi less commonly, such as B. graminis infecting monocot plants, than PM species on dicotyledonous plants [59].
However, even in the absence of a strict host association between Ampelomyces and PM fungi, i.e., no species specificity, qualitative differences between Ampelomyces strains in their ability to infect different PM fungi cannot be ruled out. In a previous study, Ampelomyces mycoparasites formed more pycnidia in colonies of the original host than in those of other PM fungi [82]. In other studies, the opposite was observed, namely that Ampelomyces strains isolated from different PM species were similarly capable of parasitizing colonies of other PM species, regardless of the original host, both in vitro and in field experiments [46,74,77]. In their mycoparasitic tests with Japanese Ampelomyces strains, Németh et al. [61] used five PM species maintained in the greenhouse: B. graminis f. sp. hordei race 1 KBP-01 (on barley Hordeum vulgare L. cv. ‘Kobinkatagi’), E. neolycopersici (=Pseudoidium neolycopersici) KTP-03 (on tomato Solanum lycopersicum Mill. cv. ‘Moneymaker’), E. trifoliorum KRCP-4N (on red clover Trifolium pratense L., cv. ‘Megium’), P. aphanis KSP-7N (on strawberry Fragaria × ananassa Duchesne cv. ‘Sagahonoka’), and P. xanthii KMP-6N (on melon Cucumis melo L., cv. ‘Earl’s Favourite’). Then, PM-infected plants were spray-inoculated with spore suspensions and then the mycoparasitic activity was scored. Japanese Ampelomyces strains successfully infected all five PM isolates and formed mature pycnidia in four out of five mycohost colonies (E. trifoliorum, E. neolycopersici, P. aphanis, and P. xanthii). The tested strains infected melon PM more heavily than the other hosts, as reflected by the formation of a larger number of pycnidia at 14 dpi. However, there were no significant differences in the mycoparasitic activity of the eight Japanese Ampelomyces strains based on three-level scoring.
Understanding host specificity is complicated by the existence of strain-level differences between Ampelomyces strains, as in laboratory experiments, strong differences in the mycoparasitic ability of different Ampelomyces strains were observed [46,70], including with respect to the PM species [70]. However, in general, the most effective Ampelomyces strains are very effective not only against the original host, but also against other PM species [46,70]. Those observations imply that the degree of mycoparasitism does not depend on the original host fungus [46], nor is it a general characteristic of individual genetic clades; rather, it reflects differences at the strain level. Indeed, the contradictory results obtained in experimental work might be partially explained by differences at the strain level.
A summary of the available data leads to the conclusion that they do not contradict the possibility of a “certain degree of host specialization” [44] among these mycoparasites. However, as this conclusion conflicts with the experimental evidence, strict (exclusive) host specialization can in fact be ruled out, and it instead suggests structural specificity [85], defined as the ability of a given parasite to parasitize different host fungi but in different proportions or with different abundances depending on the host [85]. This holds true for Ampelomyces. Structural specificity can also result in an apparent association with host fungi without implying a narrow host specialization. This is well demonstrated by Ampelomyces strains associated with the causal agent of apple PM (P. leucotricha): while these strains are mostly found in P. leucotricha, they easily colonize other mycohosts as well [74,77].
From a practical point of view, the lack of strict host specificity [24] allows a single Ampelomyces strain to be applied as a BCA against a wide range of PM species. Several studies demonstrated the biocontrol potential of Ampelomyces species against PM on various crops, such as E. trifoliorum on red clover [64], P. leucotricha on apple [84,86], P. xanthii on cucumber [19,20,66,76,79,86,87,88,89,90,91,92] and melon [93,94,95], E. necator on grapevine [22,76,82,92,96,97], B. hordei on barley [59,83], and B. graminis on wheat [83], and several other species as well [64,76,79,86,89,98,99,100,101,102].
7. Latest Results on Ampelomyces—PM Interaction
7.1. Methodological Considerations of Spray Inoculation of Ampelomyces Spores onto PM Colonies
The effective control of PM using mycoparasitic strains requires a method for inoculating hyperparasite spores onto PM fungal colonies. Ampelomyces is usually spray-inoculated onto PM-infected plants as a spore suspension, with the applications repeated several times during the season to ensure a high level of control [79,103]. Gu and Ko [104] reported that the concentration of hyperparasite spores is an important factor affecting their germination and infection in pathogens, as spore germination decreases rapidly at spore concentrations >106 spores mL−1, due to the production of self-inhibitory compounds. In our spray inoculation system, spore suspensions of Ampelomyces are diluted to 5 × 105 spores mL−1, and polyoxyethylene sorbitan monolaurate (Tween 20) is added to a final concentration of 0.05%. With this method, spores of Ampelomyces germinate successfully 15–20 h after spray inoculation onto PM-inoculated plant leaves at high RH [61,95].
7.2. Infection Processes of Ampelomyces Strains in PM Fungi
In Németh et al. [62], A. quisqualis transformants expressing an integrated green fluorescent protein (GFP) gene could be visualized in PM fungi and PM-infected leaves, which allowed for the localization of mycoparasitic fungi in PM hyphae. The method described by Suzuki et al. [105] was used to visualize tri-trophic interactions among mycoparasites, mycohosts, and plant cells. Further insights into mycoparasitism, including direct observations of the infection process of Ampelomyces strains, were obtained in real-time using high-fidelity digital microscopy (KH-2700 DM; Hirox, Tokyo, Japan) to monitor mycoparasite–mycohost interactions and thus determine how and when mycoparasites invade PM structures. The infection process of Ampelomyces strains in tomato PM E. neolycopersici on leaf type I trichomes of common tomato (S. lycopersicum Mill. cv. ‘Moneymaker’) and in melon PM colonies was also observed using digital microscopy (KH-2700 DM). Németh et al. [61] visually followed the infection of tomato PM colonies and subsequent conidiogenesis of an Ampelomyces strain. Foot cells and generative cells (GCs) of PM conidiophores began to atrophy at 5–6 dpi, with the formation of intracellular pycnidia of the hyperparasite strain initiated in basal cells of the conidiophores at 6–8 dpi, followed by the complete collapse of the conidiophores at 10–14 dpi. Kimura et al. [95] observed the degeneration and constriction of hyphae in melon PM P. xanthii prior to intracellular pycnidial formation in the hyphae (ex. conidiophores). Infection and conidiogenesis by the tested hyperparasitic Ampelomyces strain were very similar in melon PM fungus and in tomato PM fungus, as reported by Németh et al. [61]. Interestingly, almost all intracellular pycnidia were produced in conidiophores of the mycohost.
Based on earlier work and our detailed microscopic analysis, the approximate time course of infection, the events that take place in the mycoparasites once they entered the mycohosts (PM fungi), and the morphological changes in a mycoparasite-infected mycohost can be summarized as follows: Ampelomyces hyphae within parasitized PM conidia are spread by wind [20,106,107] and spores are dispersed, e.g., by rain splash [68,108]. The processes that, after spore germination, allow Ampelomyces to penetrate and parasitize hyphae of PM fungi may be mechanical [68] or enzymatic [70,71]. Penetration of mycohost structures by the hyperparasite Ampelomyces can occur within 24 h [68,93]. The mycoparasite hyphae continue their growth in PM structures, extending from cell to cell through the septal pores, and further ramifying throughout the mycohost hyphae [68,69]. The mycohost invasion by Ampelomyces leads to atrophy in 5–6 dpi and then to complete disruption of the mycohost conidiophores at 7 dpi. Disruption of the cytoplasm of the fungal hosts causes the reduced growth and eventually the death of the host fungus [22,69,82]. During the course of infection, Ampelomyces produces intracellular pycnidia in the hyphae or conidiophores of their mycohosts at 5–10 dpi [19,68,81]. In contrast to other pycnidial mycoparasites, such as Coniothyrium minitans Campbell [109,110,111], toxin production by Ampelomyces was not detected [112].
7.3. Pycnidial Development of Ampelomyces Strains in PM Fungi
Spores of Ampelomyces strains are produced in pycnidia that develop intracellularly in the mycelia of PM fungi [92]. The pycnidia of Ampelomyces are formed ubiquitously in PM colonies (Figure 2A), with a change in colour from pale yellow (immature) to black (mature) over time [95]. The number of pycnidia of Ampelomyces per melon PM colony was shown to increase with the age of the PM colony [95]. Mature pycnidia have a size range of 40.2–84.2 × 22.6–48.1 μm, and a single mature pycnidium produces 199.4–1492.7 spores by 14 dpi [61]. Both the number of spores developed in a single pycnidium and the sizes of pycnidia among strains can significantly differ [61].
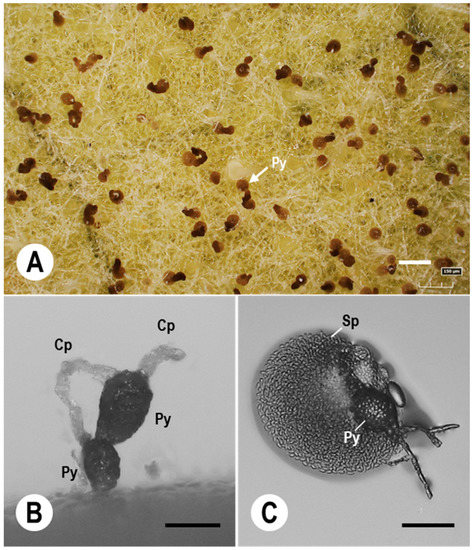
Figure 2.
Digital micrographs of P. xanthii colonies and conidiophores on melon leaves spray-inoculated with spores of a Japanese Ampelomyces strain. (A) Digital microscopy images of pycnidia (Py) of an Ampelomyces strain cultivated in plastic boxes at 70–80% relative humidity (RH) and under growth chamber conditions; the images were taken at 10 days post-inoculation (dpi) of 15-day-old melon PM colonies. (B) Pycnidia (Py) of the Ampelomyces strain that developed in melon PM conidiophores (Cp). The pycnidia were successfully produced in generative cells of the conidiophores at high RH (70–80%). (C) Ampelomyces spores released from a pycnidium after treatment with a 10 µL drop of distilled water. Mature pycnidia (Py) released abundant progeny spores (Sp). Bars: 150 µm (A,B), and 60 µm (C).
Detailed observations on pycnidial development were obtained using tomato and melon PM colonies infected with Ampelomyces strains following spray inoculation [61,95]. Almost all pycnidia were produced in conidiophores of the mycohost (Figure 2B). Following infection of tomato PM fungi, the first signs of atrophy were seen in foot cells and GCs of the conidiophores (normal noncatenate conidia) at 5–6 dpi. Intracellular pycnidia of Ampelomyces were initially produced mostly in the basal cells of the conidiophores at 6–8 dpi, during which time Ampelomyces hyphae and pycnidia continued to elongate in the host hyphae. The conidiophores completely collapsed at 10–14 dpi. In melon PM fungus, intracellular pycnidia of Ampelomyces initiated within GCs of the conidiophores (normal catenate conidia, forming chains) at 6–8 dpi. Single conidia formed at the top of the conidiophores and began to atrophy at 7–9 dpi, with complete atrophy at 10–11 dpi and complete collapse of the conidiophores at 11–12 dpi. PM hyphae containing conidiophores on melon leaves also underwent complete collapse. Melon PM colonies were therefore unable to scatter their asexual progeny conidia from the conidiophores. Pycnidia of Ampelomyces matured within 12–14 days. In the presence of water, Ampelomyces spores were released from intracellular pycnidia by the rupture of both the pycnidial and the PM cell walls (Figure 2C). The released mature spores served as sources of subsequent infections for PM hyphae.
7.4. Quantitative Analysis of PM Conidia Released from Ampelomyces-Parasitized PM Colonies under Greenhouse Conditions
In the natural environment, the asexual conidia produced by PM fungi on conidiophores (Figure 1B) are dispersed by wind over large areas and are the source of host plant infection [113,114,115,116]. Ampelomyces mycoparasites suppress both asexual and sexual sporulation of the attacked PM mycelia by colonizing and destroying conidiophores [24]. Philipp et al. [88] observed that parasitized PM colonies can continue their radial growth, but their sporulation is stopped soon after Ampelomyces penetrates their mycelia. Similarly, Shishkoff, and McGrath [91] showed that Ampelomyces could not prevent the spread of PM colonies in vitro, but the parasite caused a reduction in the inoculum produced by each colony. In addition, if Ampelomyces is to be used as a BCA, its growth and spread must outpace that of PM fungi (mycohosts). The conidiation rate of PM colonies depends on several factors, including the inoculum density, physiological patterns of the host plant, and abiotic factors [29,117,118]. Intense conidiation and spread of PM fungi will prevent their successful control by Ampelomyces mycoparasites applied as BCAs. In these cases, the effect of Ampelomyces will be limited to a reduction in disease severity and a milder impact of the PM fungus on the infected plants.
Recent methodological advances and the use of an electrostatic spore collector system facilitated the quantification of conidial release from PM colonies. In a study in which an electrostatic rotational spore collector consisting of a dielectrically polarized insulator drum was used for this purpose, a single melon and strawberry PM colony maintained under greenhouse conditions was found to release an average of 12.6 × 104 conidia and 6.7 × 104 conidia throughout its lifespan, respectively [119,120]. The collection device had no detrimental effect on the survival of the fungus; the electrostatically collected conidia produced normally elongated hyphae and formed conidiophores that produce living progeny conidia [119,120].
The same methodology was employed to study Ampelomyces–PM interactions. Using electrostatic and digital microscopy techniques, Kimura et al. [95] estimated the control effects and infection efficiency of an Ampelomyces strain against a melon PM fungus. The aim of their study was to determine whether asexual PM progeny conidia, which are a source of host plant infection, are released from Ampelomyces-inoculated melon PM colonies at different developmental stages (5-, 10-, and 15-day-old colonies). The authors found that the number of conidia released from 5- to 10-day-old melon PM colonies after spray inoculation with mycoparasite spores decreased gradually, with no release by 3–5 dpi and 4–11 dpi, respectively. Thus, conidial release from melon PM colonies was suppressed completely by the application of an Ampelomyces strain under greenhouse conditions. However, the complete prevention of progeny conidial release from 15-day-old melon PM colonies required two spray inoculations with the mycoparasite; a single treatment was insufficient. In response to a single inoculation, 5-, 10-, and 15-day-old melon PM colonies ceased their expansion at 4, 5, and 2 dpi, respectively. After electrostatic spore collection, conidiophores in uninoculated melon PM colonies had a normal morphology, forming conidial chains under greenhouse conditions, whereas conidiophores and hyphae in inoculated melon PM colonies either atrophied or collapsed, with a clear decrease in the number of normal conidiophores. There were no normal melon PM conidiophores per single 5- and 10-day-old melon PM colonies following a single spray inoculation of hyperparasite spores, unlike in 15-day-old colonies. Based on these results, for successful disease control, PM colonies should be spray-inoculated with hyperparasitic fungal spores during early developmental stages (e.g., when the colonies are 5–10 days old, or as soon as PM is detected on host leaves). If older colonies (e.g., >15 days old) are spray-inoculated with mycoparasite spores, a few normal conidiophores will persist due to suboptimal control by Ampelomyces strains, allowing PM fungi to scatter progeny conidia from the colonies. During that period, some of the conidiophores of the invaded mycelium will continue to produce fresh conidia, although they might already be infected, and will thus contain intracellular hyphae of Ampelomyces [107].
8. Practical Application of Ampelomyces Strains as Biocontrol Agents of PM
Yarwood [64] first showed the potential of Ampelomyces as a BCA by demonstrating the control of clover PM (E. polygoni) in a basic experiment that reproduced the events of a natural epidemic recorded in the previous year. On the other hand, there was also a problem with the emergence of fungicide resistance to chemical control agents. Therefore, from the 1970s, interest in the biological controls of PMs increased. Kiss [121] determined that potential BCAs need to be active in the phyllosphere because PM fungi are biotrophic pathogens infecting the aerial parts of their host plants. The first significant trial of Ampelomyces was reported by Jarvis and Slingsby [19], who used conidial suspensions of the mycoparasite to successfully control cucumber PM in greenhouse trials. Control was enhanced when application was interspersed with water sprays. The many other positive examples in which Ampelomyces was subsequently used to control PM on several crops paved the way for commercialization [24,66,96,97,122]. In addition, Sundheim and Tronsmo [123] recommended Ampelomyces-based fungal biocontrol products in plant protection practice as they can be used without any risk to human health. The absence of nontarget effects of Ampelomyces biocontrol procedures was reported as well [62,124].
Ampelomyces-based BCAs can also be applied prophylactically [125,126,127], as the mycoparasite can survive on leaves without immediate contact with the targeted PM fungus, as demonstrated experimentally [62].
The most successful biocontrol experiments using Ampelomyces were carried out in greenhouses, under high RH [19,20,103], and in the field, where free water was frequently available on the treated leaves [79,82]. However, the efficacy of Ampelomyces was shown to decrease rapidly at a RH < 85–90% [125,126] or <90–95% [88,128,129]. This high RH requirement of Ampelomyces is a major obstacle to its use as a reliable BCA. Our tested Ampelomyces strain also did not produce intracellular pycnidia in PM hyphae under greenhouse conditions that included a low RH, but it did produce them in PM hyphae in growth chambers with a high RH [95]. These results, as well as those of previous studies [19,20,22,24,61,81,82], demonstrate the importance of high-RH conditions for hyperparasitic Ampelomyces strains to produce intracellular pycnidia in mycohost hyphae. The inability to form pycnidia may explain the suboptimal biocontrol of Ampelomyces at low RH.
The high RH requirement of Ampelomyces must be addressed before the mycoparasite can be used in biocontrol [125,129]. In attempts to enhance the efficacy of Ampelomyces at lower RHs, Epton and Hamed El Nil [130] selected isolates that were able to germinate at a higher vapor deficit than wild types. In an alternative approach, a number of additives, such as an emulsion of 1% paraffin oil [128], a 0.3% mineral oil surfactant [97], 0.5% Tween 20 [46], or 0.01% Tween 80 [92], were shown to increase the biocontrol performance of Ampelomyces at lower RHs, although some of these additives can control PM directly [14,98,131,132]. To observe the effects of the BCA alone requires the use of surfactants at concentrations that do not affect the development of PM hyphae [61,95].
9. Formulation and Commercialization of Ampelomyces as BCAs
The scale-up of Ampelomyces inoculum for biocontrol purposes was one of the crucial steps towards its commercialization and practical application in plant protection. Sztejnberg et al. [133] developed and patented (European Patent Office, publ. no. 0353662/1988) a simple, inexpensive method for the production of large amounts of Ampelomyces spores in fermenters. The different formulations were tested in various crops, particularly grapevine. An improved product (AQ10TM Biofungicide) registered in 1995 for use in the control of grapevine PM was subsequently also registered for use in other fruits and vegetables in conjunction with the wetting agent (formulated as water-dispersible granules) Add-Q, a spray adjuvant recommended for use together with AQ10 biofungicides [125,126]. However, Shishkoff and McGrath [91] found that in the control of cucumber PM, Add-Q was as effective alone as when combined with AQ10. Therefore, the effect of the additives should be clearly distinguished from that of Ampelomyces when assessing the efficacy of a BCA [125].
Other studies likewise showed that the efficacy of biocontrol achieved with commercial anti-PM biofungicide products, including AQ10® (Ecogen Incorporated, Langhome, PA, USA) [97], Q-fect® (Green Biotech, Paju, South Korea), Powderycare® (AgriLife, Medak, India) [10,44,65], and Bio-Dewcon 2.00 WP (India) [66], varies significantly. Some trials reported that Ampelomyces treatment was ineffective, others suggested only very limited control of PM [90,91,129,132], others reported suboptimal control [90,134], and still others reported satisfactory results [95,102,135,136], including a level of control comparable to that using conventional fungicides [122]. These contradictions might result from experimental differences, such as humidity [10], differences in the mycoparasitic activities of individual Ampelomyces strains [10,70], and/or from physiological differences between genetically similar or uniform strains of Ampelomyces [46,70,76].
Recently, the large-scale production of a new strain, CPA-9 was reported [136]. Formulation was also developed, and the efficiency of the formulated product was demonstrated [102].
10. An Ideal Spray Inoculation System for the Effective Use of Ampelomyces as a BCA
An efficient inoculation method of Ampelomyces spores to PM colonies is needed for in vitro experimental studies of the fungus, as well as for its successful use as a BCA. Based on the experimental results and the studies conducted to date, a list of criteria for the delivery of Ampelomyces as a BCA in an optimal spray inoculation system can be compiled as follows: (1) The selected strains should have exceptional mycoparasitic abilities [46,70]. (2) Adding a surfactant as a wetting agent [46,95,97,122,128] increases efficacy. (3) Spraying should be conducted at high RH, such as in the early morning or late afternoon [122]. (4) Alternatively, although technically less feasible, PM colonies can be inoculated with mycoparasite spores at night, when conidiophores do not release progeny conidia, in contrast with during the day, when progeny conidia are actively released [95,119,120]. (5) For effective control, young colonies should be targeted, i.e., as soon as PM fungi are observable on host leaves (e.g., in 5- to 10-day-old colonies), when the leaf incidence of PM is still low (<10%) [95,122]. (6) Efficiency can be further improved by adding interspersed water sprays [19] and by repeated applications [79,95,103]. In demonstration trials, spraying with Ampelomyces was as effective as conventional fungicides [122] when conditions were optimized. In addition, because A. quisqualis tolerates a number of pesticides applied in plant protection [20,81,87,93,102], it can be included in integrated plant protection programs [102].
11. Conclusions
Ampelomyces strains were demonstrated to be able to suppress PM development. The lack of their strict host species specificity enables the use of Ampelomyces strains as a BCA against a wide range of PM fungi. There are, however, problems associated with the taxonomy of the genus, and occasional difficulties with their practical use. Considerable knowledge gaps concerning Ampelomyces include the molecular and biochemical processes during mycoparasitism, which are largely understudied. For the study these, genomic [23] and transcriptomic [65] resources are available, and an efficient transformation system [62], as well as a toolbox for gene knockout [67] were developed. These provide a basis for future studies aimed at deciphering the molecular background of mycoparasitism.
On the other hand, there were some recent advances in the study of these mycoparasites, facilitating the use of Ampelomyces strains for the effective control of PM fungi. These findings and the experimental results reported in this review lead to the development of an ideal spray inoculation system for the delivery of hyperparasitic fungi to control PM pathogens. The spray inoculation system should aid experimental research on Ampelomyces, and also its practical use as a BCA. Due to the recent methodological advancements and newest results on the biology of the fungus, Ampelomyces strains have the potential to be used as effective BCAs against PM fungi as an eco-friendly alternative to conventional fungicides.
Author Contributions
Conceptualization, T.N.; methodology, M.Z.N., D.S. and T.N.; resources, M.Z.N. and T.N.; data curation, M.Z.N., D.S. and T.N.; writing—original draft preparation, M.Z.N. and T.N.; writing—review and editing, M.Z.N. and T.N.; visualization, M.Z.N. and T.N.; supervision, M.Z.N. and T.N.; project administration, T.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences, awarded to Márk Z. Németh (BO/00221/21/4).
Data Availability Statement
Referred data sets are available in earlier publications, or from the current author on reasonable request.
Acknowledgments
The authors thank professional editors, native speakers of English, for checking the English of this document.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kiss, L.; Vaghefi, N.; Bransgrove, K.; Dearnaley, J.D.W.; Takamatsu, S.; Tan, Y.P.; Marston, C.; Liu, S.-Y.; Jin, D.-N.; Adorada, D.L.; et al. Australia: A continent without native powdery mildews? The first comprehensive catalog indicates recent introductions and multiple host range expansion events, and leads to the re-discovery of Salmonomyces as a new lineage of the Erysiphales. Front. Microbiol. 2020, 11, 1571. [Google Scholar] [CrossRef]
- Vaghefi, N.; Kusch, S.; Németh, M.Z.; Seress, D.; Braun, U.; Takamatsu, S.; Panstruga, R.; Kiss, L. Beyond nuclear ribosomal DNA sequences: Evolution, taxonomy, and closest known saprobic relatives of powdery mildew fungi (Erysiphaceae) inferred from their first comprehensive genome-scale phylogenetic analyses. Front. Microbiol. 2022, 13, 903024. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.M. Powdery mildew of strawberries. In The Powdery Mildews; Spencer, D.M., Ed.; Academic Press: New York, NY, USA, 1978; pp. 355–358. [Google Scholar]
- Keinath, T.P.; Wintermantel, W.M.; Zitter, T.A. Compendium of Cucurbit Diseases; APS Press: St. Paul, MN, USA, 2017. [Google Scholar]
- Miyamoto, T.; Hayashi, K.; Ogawara, T. First report of the occurrence of multiple resistance to Flutianil and Pyriofenone in field isolates of Podosphaera xanthii, the causal fungus of cucumber powdery mildew. Eur. J. Plant Pathol. 2020, 156, 953–963. [Google Scholar] [CrossRef]
- Miyamoto, T.; Hayashi, K.; Okada, R.; Wari, D.; Ogawara, T. Resistance to succinate dehydrogenase inhibitors in field isolates of Podosphaera xanthii on cucumber: Monitoring, cross-resistance patterns and molecular characterization. Pestic. Biochem. Physiol. 2020, 169, 104646. [Google Scholar] [CrossRef] [PubMed]
- Vielba-Fernández, A.; Polonio, A.; Ruiz-Jiménez, L.; de Vicente, A.; Pérez-García, A.; Fernández-Ortuño, D. Fungicide resistance in powdery mildew fungi. Microorganisms 2020, 8, 1431. [Google Scholar] [CrossRef]
- Jeffries, P.; Young, T.W.K. Interfungal Parasitic Relationships; CABI Publishing: Wallingford, UK, 1994. [Google Scholar]
- Prahl, R.E.; Khan, S.; Deo, R.C. Ampelomyces mycoparasites of powdery mildews–A review. Can. J. Plant Pathol. 2023. [Google Scholar] [CrossRef]
- Kiss, L. A review of fungal antagonists of powdery mildews and their potential as biocontrol agents. Pest Manag. Sci. 2003, 59, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Kiss, L. Intracellular mycoparasites in action: Interactions between powdery mildew fungi and Ampelomyces. In Stress in Yeasts and Filamentous Fungi; Avery, S.V., Stratford, M., van West, P., Eds.; Academic Press: Cambridge, MA, USA, 2008; Volume 27, pp. 37–52. [Google Scholar]
- Hijwegen, T.; Buchenauer, H. Isolation and identification of hyperparasitic fungi associated with Erysiphaceae. Neth. J. Plant Pathol. 1984, 90, 79–84. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Shaw, L.A.; Traquair, J.A. Factors affecting antagonism of cucumber powdery mildew by Stephanoascus flocculosus and S. rugulosus. Mycol. Res. 1989, 92, 162–165. [Google Scholar] [CrossRef]
- Verhaar, M.A.; Hijwegen, T.; Zadoks, J.C. Glasshouse experiments on biocontrol of cucumber powdery mildew (Sphaerotheca fuliginea) by the mycoparasites Verticillium lecanii and Sporothrix rugulosa. Biol. Cont. 1996, 6, 353–360. [Google Scholar] [CrossRef]
- Hoch, H.C.; Provvidenti, R. Mycoparasitic relationships: Cytology of the Sphaerotheca fuliginea-Tilletiopsis sp. interaction. Phytopathology 1979, 69, 359–362. [Google Scholar] [CrossRef]
- Hijwegen, T. Biological control of cucumber powdery mildew by Tilletiopsis minor. Neth. J. Plant Pathol. 1986, 92, 93–95. [Google Scholar] [CrossRef]
- Cesati, V. Ampelomyces quisqualis Ces. Bot. Ztg. 1852, 10, 301–302. [Google Scholar]
- Yarwood, C.E. An overwintering pycnidial stage of Cicinnobolus. Mycologia 1939, 31, 420–422. [Google Scholar] [CrossRef]
- Jarvis, W.R.; Slingsby, K. The control of powdery mildew of greenhouse cucumber by water sprays and Ampelomyces quisqualis. Plant Dis. Reptr. 1977, 61, 728–730. [Google Scholar]
- Sundheim, L. Control of cucumber powdery mildew by the hyperparasite Ampelomyces quisquialis and fungicides. Plant Pathol. 1982, 31, 209–214. [Google Scholar] [CrossRef]
- Kiss, L. Natural occurrence of Ampelomyces intracellular mycoparasites in mycelia of powdery mildew fungi. New Phytol. 1998, 140, 709–714. [Google Scholar] [CrossRef]
- Falk, S.P.; Gadoury, D.M.; Cortesi, P.; Pearson, R.C.; Seem, R.C. Parasitism of Uncinula necator cleistothecia by the mycoparasite Ampelomyces quisqualis. Phytopathology 1995, 85, 794–800. [Google Scholar] [CrossRef]
- Huth, L.; Ash, G.J.; Idnurm, A.; Kiss, L.; Vaghefi, N. The “bipartite” structure of the first genome of Ampelomyces quisqualis, a common hyperparasite and biocontrol agent of powdery mildews, may point to its evolutionary origin from plant pathogenic fungi. Genome Biol. Evol. 2021, 13, evab182. [Google Scholar] [CrossRef]
- Kiss, L.; Russell, J.C.; Szentiványi, O.; Xu, X.; Jeffries, P. Biology and biocontrol potential of Ampelomyces mycoparasites, natural antagonists of powdery mildew fungi. Biocont. Sci. Technol. 2004, 14, 635–651. [Google Scholar] [CrossRef]
- Manjunatha, L.; Singh, S.; Ravikumara, B.M.; Narasa Reddy, G.; Senthikumar, M. Ampelomyces. In Beneficial Microbes in Agro-ecology; Amaresan, N., Senthil Kumar, M., Annapuma, K., Kumar, K., Sankaranarayanan, A., Eds.; Academic Press: Amsterdam, The Netherlands, 2020; Chapter 44; pp. 833–860. [Google Scholar] [CrossRef]
- Matsuda, Y.; Kashimoto, K.; Takikawa, Y.; Aikami, R.; Nonomura, T.; Toyoda, H. Occurrence of new powdery mildew on greenhouse tomato cultivars. J. Gen. Plant Pathol. 2001, 67, 294–298. [Google Scholar] [CrossRef]
- White, J.F., Jr.; Johnston, S.A.; Wang, C.L.; Chin, C.K. First report of powdery mildew in greenhouse-grown tomatoes in New Jersey. Plant Dis. 1997, 81, 227. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of the Erysiphales (Powdery Mildews); CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012. [Google Scholar]
- Yarwood, C.E. Powdery mildews. Bot. Rev. 1957, 23, 235–301. [Google Scholar] [CrossRef]
- Braun, U. A monograph of the Erysiphales (powdery mildews). Beih. Nova Hedwig. 1987, 89, 1–700. [Google Scholar]
- Rallos, L.E.E.; Baudoin, A.B. Co-occurrence of two allelic variants of CYP51 in Erysiphe necator and their correlation with over-expression for DMI resistance. PLoS ONE 2016, 11, e0148025. [Google Scholar] [CrossRef]
- Pintye, A.; Németh, M.Z.; Molnár, O.; Horváth, Á.; Spitzmüller, Z.; Szalóki, N.; Pal, K.; Váczy, K.; Kovács, G. Improved DNA extraction and quantitative real-time PCR for genotyping Erysiphe necator and detecting the DMI fungicide resistance marker A495T, using single ascocarps. Phytopathol. Medit. 2020, 59, 97–106. [Google Scholar] [CrossRef]
- Huggenberger, F.; Collins, M.A.; Skylakakis, G. Decreased sensitivity of Sphaerotheca fuliginea to fenarimol and other ergosterol-biosynthesis inhibitors. Crop Prot. 1984, 3, 137–149. [Google Scholar] [CrossRef]
- McGrath, M.T.; Shishkoff, N. Resistance to triadimefon and benomyl: Dynamics and impact on managing cucurbit powdery mildew. Plant Dis. 2001, 85, 147–154. [Google Scholar] [CrossRef]
- McGrath, M.T.; Shishkoff, N. Frist report of the cucurbit powdery mildew fungus (Podosphaera xanthii) resistant to strobilurin fungicides in the United States. Plant Dis. 2003, 87, 1007. [Google Scholar] [CrossRef]
- Lebeda, A.; Sedláková, B. Fungicide resistance in population of cucurbit powdery mildew. J. Plant Pathol. 2008, 90, S2.142. [Google Scholar]
- Petit, A.N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant-Gaveau, N. Fungicide impacts on photosynthesis in crop plants. Photosyn. Res. 2012, 111, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tscharntke, T.; Winqvist, C.; et al. Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- De Bary, A. Eurotium, Erysiphe, Cicinnobolus: Nebst Bemerkungen über die Geschlechtsorgane der Ascomyceten. In Beitrage zur Morphologie und Physiologie der Pilze; De Bary, A., Woronin, M., Eds.; Verlag von C. Winter: Frankfurt, Germany, 1870; pp. 1–95. [Google Scholar]
- Rogers, D. On Cicinobolus. Mycologia 1959, 51, 96–98. [Google Scholar] [CrossRef]
- Kiss, L.; Nakasone, K.K. Ribosomal DNA internal transcribed spacer sequences do not support the species status of Ampelomyces quisqualis, a hyperparasite of powdery mildew fungi. Curr. Genet. 1998, 33, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Yang, J.; Kovács, G.M.; Szentiványi, O.; Li, B.; Xu, X.; Kiss, L. Genetic diversity of Ampelomyces mycoparasites isolated from different powdery mildew species in China inferred from analyses of rDNA ITS sequences. Fungal Div. 2007, 24, 225–240. [Google Scholar]
- Pintye, A.; Bereczky, Z.; Kovács, G.M.; Nagy, L.G.; Xu, X.; Legler, S.E.; Váczy, Z.; Váczy, K.Z.; Caffi, T.; Rossi, V.; et al. No indication of strict host associations in a widespread mycoparasite: Grapevine powdery mildew (Erysiphe necator) is attacked by phylogenetically distant Ampelomyces strains in the field. Phytopathology 2012, 102, 707–716. [Google Scholar] [CrossRef]
- Park, M.-J.; Choi, Y.-J.; Hong, S.-B.; Shin, H.-D. Genetic variability and mycohost association of Ampelomyces quisqualis isolates inferred from phylogenetic analyses of ITS rDNA and actin gene sequences. Fungal Biol. 2010, 114, 235–247. [Google Scholar] [CrossRef]
- Prahl, R.E.; Khan, S.; Deo, R.C. The role of internal transcribed spacer 2 secondary structures in classifying mycoparasitic Ampelomyces. PLoS ONE 2021, 16, e0253772. [Google Scholar] [CrossRef]
- Legler, S.E.; Pintye, A.; Caffi, T.; Gulyás, S.; Bohár, G.; Rossi, V.; Kiss, L. Sporulation rate in culture and mycoparasitic activity, but not mycohost specificity, are the key factors for selecting Ampelomyces strains for biocontrol of grapevine powdery mildew (Erysiphe necator). Eur. J. Plant Pathol. 2016, 144, 723–736. [Google Scholar] [CrossRef]
- Kiss, L. Genetic diversity in Ampelomyces isolates, hyperparasites of powdery mildew fungi, inferred from RFLP analysis of the rDNA ITS region. Mycol. Res. 1997, 101, 1073–1080. [Google Scholar] [CrossRef]
- Hino, I.; Kato, H. Cicinnoboli parasitic on mildew fungi. Bull. Miyazaki Coll. Agric. For. 1929, 1, 91–98. [Google Scholar]
- Emmons, C.W. Cicinnobolus cesatii, a study in host-parasite relationships. Bull. Torrey Bot. Club 1930, 57, 421–439. [Google Scholar] [CrossRef]
- Blumer, S. Die Erysiphaceen Mitteleuropas mit besonderer Berücksichtigung der Schweiz. Beitr. Zur Kryptogamenflora Schweiz 1933, 7, 1–483. [Google Scholar]
- Clare, B.G. Ampelomyces quisqualis (Cicinnobolus cesatii) on Queensland Erysiphaceae. Univ. Qld. Pap. 1964, 4, 147–149. [Google Scholar]
- Belsare, S.W.; Moniz, L.; Deo, V.B. The hyperparasite Ampelomyces quisqualis Ces. from Maharashtra State, India. Biovigyanam 1980, 6, 173–176. [Google Scholar]
- Hanlin, R.T.; Tortolero, O. Brasiliomyces, a new host for Ampelomyces. Mycotaxon 1984, 21, 459–462. [Google Scholar]
- Nagy, S.G.; Vajna, L. Ampelomyces fajok elöfordulása lisztharmatgombákban Magyarországon. Mikológiai Közlemények 1990, 29, 103–112. (In Hungarian) [Google Scholar]
- Puzanova, L.A. Distribution of hyperparasites from the genus Ampelomyces Ces. ex Schlecht. on the powdery mildew fungi in northern Caucasus and their importance in disease control. Mikol. I Fitopatol. 1991, 25, 438–442. (In Russian) [Google Scholar]
- Tsay, J.G.; Tung, B. Ampelomyces quisqualis Ces. ex Schlecht., a hyperparasite of the asparagus bean powdery mildew pathogen Erysiphe polygoni in Taiwan. Trans. Mycol. Soc. R.O.C. 1991, 6, 55–58. [Google Scholar]
- Paulech, C.; Herrera, S.; Fornet, E. Phytopathogenic micromycetes of the family Erysiphaceae distributed in Cuba. Ceska Mykol. 1993, 46, 303–314. (In Slovakian) [Google Scholar]
- Shin, H.-D. Isolation and identification of hyperparasites against powdery mildew fungi in Korea. Kor. J. Mycol. 1994, 22, 355–365. (In Korean) [Google Scholar]
- Kiss, L. Graminicolous powdery mildew fungi as new natural hosts of Ampelomyces mycoparasites. Can. J. Bot. 1997, 75, 680–683. [Google Scholar] [CrossRef]
- Ranković, B. Hyperparasites of the genus Ampelomyces on powdery mildew fungi in Serbia. Mycopathologia 1997, 139, 157–164. [Google Scholar] [CrossRef]
- Németh, M.Z.; Mizuno, Y.; Kobayashi, H.; Seress, D.; Shishido, N.; Kimura, Y.; Takamatsu, S.; Suzuki, T.; Takikawa, Y.; Kakutani, K.; et al. Ampelomyces strains isolated from diverse powdery mildew hosts in Japan: Their phylogeny and mycoparasitic activity, including timing and quantifying mycoparasitism of Pseudoidium neolycopersici on tomato. PLoS ONE 2021, 16, e0251444. [Google Scholar] [CrossRef] [PubMed]
- Németh, M.Z.; Pintye, A.; Horváth, Á.N.; Vági, P.; Kovács, G.M.; Gorfer, M.; Kiss, L. Green fluorescent protein transformation sheds more light on a widespread mycoparasitic interaction. Phytopathology 2019, 109, 1404–1416. [Google Scholar] [CrossRef] [PubMed]
- Riess, H. Uber Byssocystis textilis. Hedwigia 1852, 1, 23. [Google Scholar]
- Yarwood, C.E. Ampelomyces quisqualis on clover mildew. Phytopathology 1932, 22, 31. [Google Scholar]
- Siozios, S.; Tosi, L.; Ferrarini, A.; Ferrari, A.; Tononi, P.; Bellin, D.; Maurhofer, M.; Gessler, C.; Delledonne, M.; Pertot, I. Transcriptional reprogramming of the mycoparasitic fungus Ampelomyces quisqualis during the powdery mildew host-induced germination. Phytopathology 2015, 105, 199–209. [Google Scholar] [CrossRef]
- Sivakumar, T.; Balabaskar, P.; Renganathan, P.; Sanjeevkumar, K. To evaluate the bio-efficacy and phytotoxicty of powder formulation of Biodewcon (Ampelomyces quisqualis 2.00% Wp) against powdery mildew (Sphaerotheca fuliginea) in cucumber crop. Plant Arch. 2020, 20, 3811–3815. [Google Scholar]
- Németh, M.Z.; Li, G.; Seress, D.; Pintye, A.; Molnár, O.; Kovács, G.M.; Kiss, L.; Gorfer, M. What is the role of the nitrate reductase (euknr) gene in fungi that live in nitrate-free environments? A targeted gene knock-out study in Ampelomyces mycoparasites. Fungal Biol. 2021, 125, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Sundheim, L.; Krekling, T. Host-parasite relationships of the hyperparasite Ampelomyces quisqualis and its powdery mildew host Sphaerotheca fuliginea. Phytopath. Z. 1982, 104, 202–210. [Google Scholar] [CrossRef]
- Hashioka, Y.; Nakai, Y. Ultrastructure of pycnidial development and mycoparasitism of Ampelomyces quisqualis parasitic on Erysiphales. Trans. Mycol. Soc. Jpn. 1980, 21, 329–338. [Google Scholar]
- Angeli, D.; Puopolo, G.; Maurhofer, M.; Gessler, C.; Pertot, I. Is the mycoparasitic activity of Ampelomyces quisqualis biocontrol strains related to phylogeny and hydrolytic enzyme production? Biol. Cont. 2012, 63, 348–358. [Google Scholar] [CrossRef]
- Rotem, Y.; Yarden, O.; Sztejnberg, A. The mycoparasite Ampelomyces quisqualis expresses exgA encoding and exo-β-1,3-glucanase in culture and during mycoparasitism. Phytopathology 1999, 89, 631–638. [Google Scholar] [CrossRef]
- Philipp, W.D. Extracellular enzymes and nutritional physiology of Ampelomyces quisqualis Ces., hyperparasite of powdery mildew, in vitro. J. Phytopathol. 1985, 114, 274–283. [Google Scholar] [CrossRef]
- Vaidya, S.; Thakur, V.S. Ampelomyces quisqualis Ces.–A mycoparasite of apple powdery mildew in western Himalayas. Indian Phytopathol. 2005, 58, 250–251. [Google Scholar]
- Szentiványi, O.; Kiss, L.; Russell, J.C.; Kovács, G.M.; Varga, K.; Jankovics, T.; Lesemann, S.; Xu, X.-M.; Jeffries, P. Ampelomyces mycoparasites from apple powdery mildew identified as a distinct group based on single-stranded conformation polymorphism analysis of the rDNA ITS region. Mycol. Res. 2005, 109, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, K.K.; Khan, S.; Brooks, S.; Mortimer, P.E.; Karunarathna, S.C.; Xu, J.; Hyde, K.D. Morpho-molecular characterization of two Ampelomyces spp. (Pleosporales) strains mycoparasites of powdery mildew of Hevea brasiliensis. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Angeli, D.; Maurhofer, M.; Gessler, C.; Pertot, I. Existence of different physiological forms within genetically diverse strains of Ampelomyces quisqualis. Phytoparasitica 2012, 40, 37–51. [Google Scholar] [CrossRef]
- Kiss, L.; Pintye, A.; Kovács, G.M.; Jankovics, T.; Fontaine, M.C.; Harvey, N.; Xu, X.; Nicot, P.C.; Bardin, M.; Shykoff, J.A.; et al. Temporal isolation explains host-related genetic differentiation in a group of widespread mycoparasitic fungi. Mol. Ecol. 2011, 20, 1492–1507. [Google Scholar] [CrossRef]
- Pintye, A.; Ropars, J.; Harvey, N.; Shin, H.-D.; Leyronas, C.; Nicot, P.C.; Giraud, T.; Kiss, L. Host phenology and geography as drivers of differentiation in generalist fungal mycoparasites. PLoS ONE 2015, 10, e0120703. [Google Scholar] [CrossRef]
- Sztejnberg, A.; Galper, S.; Mazar, S.; Lisker, N. Ampelomyces quisqualis for biological and integrated control of powdery mildews in Israel. J. Phytopathol. 1989, 124, 285–295. [Google Scholar] [CrossRef]
- Paratt, S.R.; Laine, A.-L. Pathogen dynamics under both bottom-up host resistance and top-down hyperparasite attack. J. Appl. Ecol. 2018, 55, 2976–2985. [Google Scholar] [CrossRef] [PubMed]
- Philipp, W.-D.; Crüger, G. Parasitismus von Ampelomyces quisqualis auf echten Mehltaupilzenal an Gurken und anderen Gemüsearten. Z. Pflanzenkrankh. Pflanzenschutz 1979, 86, 129–142. [Google Scholar]
- Falk, S.P.; Gadoury, D.M.; Pearson, R.C.; Seem, R.C. Partial control of grape powdery mildew by the mycoparasite Ampelomyces quisqualis. Plant Dis. 1995, 79, 483–490. [Google Scholar] [CrossRef]
- Mairovich, G.; Marder, J.B.; Shtienberg, D.; Sztejnberg, A. Studies of biological control of powdery mildew in cereals by the hyperparasite Ampelomyces quisqualis. Phytoparasitica 1996, 24, 157–158. [Google Scholar]
- Szentiványi, O.; Kiss, L. Overwintering of Ampelomyces mycoparasites on apple trees and other plants infected with powdery mildews. Plant Pathol. 2003, 52, 737–746. [Google Scholar] [CrossRef]
- Poulin, R.; Krasnov, B.R.; Mouillot, D. Host specificity in phylogenetic and geographic space. Trends Parasitol. 2011, 27, 355–361. [Google Scholar] [CrossRef]
- Sztejnberg, A. Biological control of powdery mildews by Ampelomyces quisqualis. Phytopathology 1979, 69, 1047. [Google Scholar]
- Sundheim, L. Ampelomyces quisqualis, a hyperparasitic fungus in biological control of powdery mildews on greenhouse cucumber. Acta Hortic. 1984, 156, 229–236. [Google Scholar] [CrossRef]
- Philipp, W.-D.; Grauer, U.; Grossmann, F. Ergänzende Untersuchungen zur biologischen und integrierten Bekämpfung von Gurkenmehltau unter Glass durch Ampelomyces quisqualis. Z. Pflanzenkrankh. Pflanzenschutz 1984, 91, 438–443. [Google Scholar]
- Sztejnberg, A.; Mazar, S. Biocontrol of cucumber and carrot powdery mildew by Ampelomyces quisqualis. Phytopathology 1985, 75, 1301–1302. [Google Scholar]
- Dik, A.J.; Verhaar, M.A.; Bélanger, R.R. Comparison of three biological control agents against cucumber powdery mildew (Sphaerotheca fuliginea) in semi-commercial-scale glasshouse trials. Eur. J. Plant Pathol. 1998, 104, 413–423. [Google Scholar] [CrossRef]
- Shishkoff, N.; McGrath, M.T. AQ10 biofungicide combined with chemical fungicides or AddQ spray adjuvant for control of cucurbit powdery mildew in detached leaf culture. Plant Dis. 2002, 86, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Angeli, D.; Saharan, K.; Segarra, G.; Sicher, C.; Pertot, I. Production of Ampelomyces quisqualis conidia in submerged fermentation and improvements in the formulation for increased shelf-life. Crop Prot. 2017, 97, 135–144. [Google Scholar] [CrossRef]
- Sztejnberg, A.; Mazar, S. Studies on the hyperparasite Ampelomyces quisqualis and preliminary trials on biological control of powdery mildew. Phytoparasitica 1983, 11, 219–220. [Google Scholar]
- Romero, D.; Rivera, M.E.; Cazorla, F.M.; De Vicente, A.; Pérez-García, A. Effect of mycoparasitic fungi on the development of Sphaerotheca fusca in melon leaves. Mycol. Res. 2003, 107, 64–71. [Google Scholar] [CrossRef]
- Kimura, Y.; Németh, M.Z.; Numano, K.; Mitao, A.; Shirakawa, T.; Seress, D.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; Kiss, L.; et al. Hyperparasitic fungi against melon powdery mildew pathogens: Quantitative analysis of conidia released from single colonies of Podosphaera xanthii parasitized by Ampelomyces. Agronomy 2023, 13, 1204. [Google Scholar] [CrossRef]
- Hofstein, R.; Fridlender, B. Development of production, formulation and delivery systems. In Brighton Crop Protection Conference, Pest and Diseases; British Crop Protection Council: Farnham, UK, 1994; Volume 3, pp. 1273–1280. [Google Scholar]
- Hofstein, R.; Daoust, R.A.; Aeschlimann, J.P. Constrains to the development of biofungicides: The example of “AQ10”, a new product for controlling powdery mildews. Entomophaga 1996, 41, 455–460. [Google Scholar] [CrossRef]
- Pasini, C.; D’Aquila, F.; Curir, P.; Gullino, M.L. Effectiveness of antifungal compounds against rose powdery mildew (Sphaerotheca pannosa var. rosae) in glasshouses. Crop Prot. 1997, 16, 251–256. [Google Scholar] [CrossRef]
- Pertot, I.; Zasso, R.; Amsalem, L.; Baldessari, M.; Angeli, G.; Elad, Y. Integrating biocontrol agents in strawberry powdery mildew control strategies in high tunnel growing systems. Crop Prot. 2008, 27, 622–631. [Google Scholar] [CrossRef]
- Gautam, A.K.; Avasthi, S. Ampelomyces quisqualis Ces.–A mycoparasite of Euphorbia hirta powdery mildew in Himachal Pradesh, India. J. Phytopathol. Pest Manag. 2016, 3, 64–70. [Google Scholar]
- Parratt, S.R.; Barrès, B.; Penczykowski, R.M.; Laine, A.L. Local adaptation at higher trophic levels: Contrasting hyperparasite-pathogen infection dynamics in the field and laboratory. Mol. Ecol. 2017, 26, 1964–1979. [Google Scholar] [CrossRef] [PubMed]
- Carbó, A.; Teixidó, N.; Usall, J.; Solsona, C.; Torres, R. Formulated Ampelomyces quisqualis CPA-9 applied on zucchini leaves: Influence of abiotic factors and powdery mildew mycoparasitization. Eur. J. Plant Pathol. 2021, 161, 37–48. [Google Scholar] [CrossRef]
- Philipp, W.-D.; Beuther, E.; Hermann, D.; Klinkert, F.; Oberwalder, C.; Schmidtke, M.; Straub, B. Zur Formulierung des Mehltauhyperparasiten Ampelomyces quisqualis Ces. Z. Pflanzenkrankh. Pflanzenschutz 1990, 97, 120–132. [Google Scholar]
- Gu, Y.H.; Ko, W.H. Water agarose medium for studying factors affecting germination of conidia of Ampelomyces quisqualis. Mycol. Res. 1997, 101, 422–424. [Google Scholar] [CrossRef]
- Suzuki, T.; Murakami, T.; Takizumi, Y.; Ishimaru, H.; Kudo, D.; Takikawa, Y.; Matsuda, Y.; Kakutani, K.; Bai, Y.; Nonomura, T. Trichomes: Interaction sites of tomato leaves with biotrophic powdery mildew pathogens. Eur. J. Plant Pathol. 2018, 150, 115–125. [Google Scholar] [CrossRef]
- Speer, E.O. Ampelomyces cesati (Fungi, Sphaeropsidales). Taxon 1978, 27, 549–562. [Google Scholar] [CrossRef]
- Kiss, L.; Pintye, A.; Zséli, G.; Jankovics, T.; Szentiványi, O.; Hafez, Y.M.; Cook, R.T.A. Microcyclic conidiogenesis in powdery mildews and its association with intracellular parasitism by Ampelomyces. Eur. J. Plant Pathol. 2010, 126, 445–451. [Google Scholar] [CrossRef]
- Zeng, L.F.; Jiang, Y.C. An investigation of parasitic fungi on Erysiphe spp. RoPP 1986, 65, 419. [Google Scholar]
- Machida, K.; Trifonov, L.S.; Ayer, W.A.; Lu, Z.-X.; Laroche, A.; Huang, H.C.; Cheng, K.J.; Zantige, J.L. 3(2H)-Benzofuranones and chromanes from liquid cultures of the mycoparasitic fungus Coniothyrium minitans. Phytochemistry 2001, 58, 173–177. [Google Scholar] [CrossRef] [PubMed]
- McQuilken, M.P.; Gemmell, J.; Whipps, J.M. Some nutritional factors affecting production of biomass and antifungal metabolites of Coniothyrium minitans. Biocont. Sci. Technol. 2002, 12, 443–454. [Google Scholar] [CrossRef]
- McQuilken, M.P.; Gemmell, J.; Hill, R.A.; Whipps, J.M. Production of macrosphelide A by the mycoparasite Coniothyrium minitans. FEMS Microbiol. Lett. 2003, 219, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Beuther, E.; Philipp, W.D.; Grossmann, F. Untersuchungen zum hyperparasitismus von Ampelomyces quisqualis auf gurkenmehltau (Sphaerotheca fuliginea). Phytopath. Z. 1981, 101, 265–270. [Google Scholar] [CrossRef]
- Aylor, D.E. The role of intermittent wind in the dispersal of fungal pathogens. Annu. Rev. Phytopathol. 1990, 28, 73–92. [Google Scholar] [CrossRef]
- Brown, J.K.M.; Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef]
- Nonomura, T.; Matsuda, Y.; Yamashita, S.; Akahoshi, H.; Takikawa, Y.; Kakutani, K.; Toyoda, H. Natural woody plant, Mallotus japonicus, as an ecological partner to transfer different pathotypic conidia of Oidium neolycopersici to greenhouse tomatoes. Plant Protect. Sci. 2013, 49, S33–S40. [Google Scholar] [CrossRef]
- Suzuki, T.; Iwasaki, S.; Hisazumi, H.; Miyamoto, A.; Ogami, H.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; Nonomura, T. Inhibitory effects of blue light-emitting diode irradiation on Podosphaera xanthii conidial release and infection of melon seedlings. Agriculture 2022, 12, 198. [Google Scholar] [CrossRef]
- Rouse, D.I.; MacKenzie, D.R.; Nelson, R.R. Density dependent sporulation of Erysiphe graminis f. sp. tritici. Phytopathology 1984, 74, 1176–1180. [Google Scholar] [CrossRef]
- Bushnell, W.R. The role of powdery mildew research in understanding host-parasite interaction: Past, present, and future. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; APS Press: St. Paul, MN, USA, 2002; pp. 1–12. [Google Scholar]
- Suzuki, T.; Nakamura, R.; Takagi, N.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; Matsui, K.; Nonomura, T. Quantitative analysis of the lifelong production of conidia released from single colonies of Podosphaera xanthii on melon leaves using electrostatic techniques. Austral. Plant Pathol. 2019, 48, 297–307. [Google Scholar] [CrossRef]
- Ayabe, S.; Kimura, Y.; Umei, N.; Takikawa, Y.; Kakutani, K.; Matsuda, Y.; Nonomura, T. Real-time collection of conidia released from living single colonies of Podosphaera aphanis on strawberry leaves under natural conditions with electrostatic techniques. Plants 2022, 11, 3453. [Google Scholar] [CrossRef]
- Kiss, L. The role of hyperparasites in host plant–parasitic fungi relationships. In Biotic Interactions in Plant–Pathogen Associations; Jeger, M.J., Spence, N.J., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 227–236. [Google Scholar]
- Daoust, R.A.; Hofstein, R. Ampelomyces quisqualis, a new biofungicide to control powdery mildew in grapes. In Brighton Crop Protection Conference, Pest and Diseases; British Crop Protection Council: Farnham, UK, 1996; Volume 1, pp. 33–40. [Google Scholar]
- Sundheim, L.; Tronsmo, A. Hyperparasites in biological control. In Biocontrol of Plant Diseases; Mukerji, K.G., Garg, K.L., Eds.; CRC Press: Boca Raton, FL, USA, 1988; Volume 1, pp. 53–69. [Google Scholar]
- Kiss, L. How dangerous is the use of fungal biocontrol agents to nontarget organisms? New Phytol. 2004, 163, 453–455. [Google Scholar] [CrossRef] [PubMed]
- Paulitz, T.C.; Bélanger, R.R. Biological control in greenhouse systems. Annu. Rev. Phytopathol. 2001, 39, 103–133. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, R.R.; Labbé, C. Control of powdery mildews without chemicals: Prophylactic and biological alternatives for horticultural crops. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; APS Press: St. Paul, MN, USA, 2002; pp. 256–267. [Google Scholar]
- Kaur, L.; Gupta, B.; Sharma, I.M.; Joshi, A.K. Eco-friendly management of powdery mildew of mango through biocontrol agents. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 392–396. [Google Scholar] [CrossRef]
- Philipp, W.-D.; Hellstern, A. Biologische Mehltaubekämpfung mit Ampelomyces quisqualis bei reduzierter Luftfeuchtigkeit. Z. Pflanzenkrankh. Pflanzenschutz 1986, 93, 384–391. [Google Scholar]
- Verhaar, M.A.; Kerssies, A.; Hijwegen, T. Effect of RH on mycoparasitism of rose powdery mildew with and without treatments with mycoparasites. Z. Pflanzenkrankh. Pflanzenschutz 1999, 106, 158–165. [Google Scholar]
- Epton, H.A.S.; Hamed El Nil, Y.F. Improvement in the hyperparasitic activity of Ampelomyces quisqualis in the biocontrol of powdery mildew of cucumber. IOBC/WPRS Bull. 1993, 16, 86–89. [Google Scholar]
- Hijwegen, T. Biological control of cucumber powdery mildew with Tilletiopsis minor under greenhouse conditions. Neth. J. Plant Pathol. 1992, 98, 221–225. [Google Scholar] [CrossRef]
- McGrath, M.T.; Shishkoff, N. Evaluation of biocompatible products for managing cucurbit powdery mildew. Crop Prot. 1999, 18, 471–478. [Google Scholar] [CrossRef]
- Sztejnberg, A.; Galper, S.; Lisker, N. Conditions for pycnidial production and spore formation by Ampelomyces quisqualis. Can. J. Microbiol. 1990, 36, 193–198. [Google Scholar] [CrossRef]
- Gilardi, G.; Manker, D.C.; Garibaldi, A.; Gullino, M.L. Efficacy of the biocontrol agents Bacillus subtilis and Ampelomyces quisqualis applied in combination with fungicides against powdery mildew of zucchini. J. Plant Dis. Prot. 2008, 115, 208–213. [Google Scholar] [CrossRef]
- Hijwegen, T. Effect of seventeen fungicolous fungi on sporulation of cucumber powdery mildew. Neth. J. Plant Pathol. 1988, 94, 185–190. [Google Scholar] [CrossRef]
- Carbó, A.; Torres, R.; Usall, J.; Ballesta, J.; Teixidó, N. Biocontrol potential of Ampelomyces quisqualis strain CPA-9 against powdery mildew: Conidia production in liquid medium and efficacy on zucchini leaves. Sci. Hortic. 2020, 267, 109337. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).