Abstract
Conyza bonariensis L. (hairy fleabane) is a significant weed in production systems, especially due to its evolving resistance to various herbicides. In Uruguay, control failures of C. bonariensis have been reported following the use of glyphosate and ALS inhibitors. The objective of this study was to investigate the occurrence of the multiple and cross-resistance of C. bonariensis to glyphosate, chlorimuron, and diclosulam, and to assess the efficacy of alternative herbicides against these resistant biotypes. Seeds were collected from sites where plants had survived herbicide applications during the 2019/2020 and 2020/2021 seasons. Following initial screenings, biotypes were selected to establish independent dose–response curves for glyphosate, diclosulam, and chlorimuron. For each herbicide, four biotypes of C. bonariensis were tested: one susceptible (S), two putatively moderately resistant (MR) biotypes, and one putatively highly resistant (R) biotype. In each assay, eight herbicide doses were used (1/32; 1/16; 1/8; 1/2; 1; 2; and 4X for S and MR biotypes, and 1/8; 1/2; 1; 2; 4; 8; and 16X for R biotypes) based on the recommended dose (1x) for each herbicide, with four repetitions per treatment. Each assay was completely replicated twice. Resistance was confirmed through testing in two plant generations (G1 and G2). The findings reveal high levels of multiple and cross-resistance in C. bonariensis to glyphosate, diclosulam, and chlorimuron. In general, herbicides with alternative action mechanisms effectively controlled C. bonariensis exhibiting multiple and cross-resistance. This study confirms the first case of C. bonariensis cross-resistance to diclosulam and chlorimuron, and the first occurrence of multiple and cross-resistance to glyphosate, diclosulam, and chlorimuron in this species.
1. Introduction
The evolution of herbicide-resistant weed biotypes represents a significant challenge in agricultural systems, leading to substantial reductions in crop productivity and increased environmental impacts associated with their management. Within the Conyza genus, over 100 cases of herbicide resistance have been reported, predominantly in Conyza bonariensis L. Cronquist (hairy fleabane), Conyza canadensis L. Cronquist (horseweed), and Conyza sumatrensis Retz. E. Walker (Sumatran fleabane). These species exhibit resistance to herbicides, including inhibitors of 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) and acetolactate synthase (ALS), synthetic auxins, and photosystem I, among others [1,2].
In Uruguay, C. bonariensis (synonym: Erigeron bonariensis L.), a member of the Asteraceae family native to South America, is particularly problematic in agricultural production systems. It infests both winter and summer crops as well as pastures and is widely distributed across regions in Argentina, Brazil, Paraguay, and Uruguay [3]. The prevalence of C. bonariensis has escalated with the adoption of no-tillage systems and the introduction of glyphosate-resistant soybean. This trend, observed since the 1990s in Uruguay and its agricultural neighboring regions, poses significant challenges [4].
The detrimental impact of C. bonariensis on crop yields primarily arises from its competition for essential resources such as water, nutrients, and light. At densities reaching 2.7 plants m−2, Conyza spp. are capable of reducing soybean yields by as much as 50% [5]. C. bonariensis is a herbaceous type of weed, characterized by an annual life cycle, which can thrive in a variety of agricultural settings. Remarkably, a single plant can produce over 800,000 seeds [6,7], which can be dispersed over long distances by wind, contributing to its widespread distribution.
The high seed output and adaptability of C. bonariensis to production systems, coupled with the repeated and frequent use of herbicides with the same mode of actions, have led to the selection of resistant biotypes. To date, 21 cases of herbicide-resistant C. bonariensis have been reported globally, primarily involving resistance to glyphosate and paraquat [2]. In Uruguay, glyphosate was the principal herbicide for controlling C. bonariensis for many years. However, since 2013, with the emergence of glyphosate-resistant C. bonariensis biotypes, ALS-inhibiting herbicides have gained popularity for pre- and post-emergence management of this species in soybeans [4]. Yet, recent reports indicate failures in controlling C. bonariensis with ALS inhibitors, particularly diclosulam and chlorimuron.
In light of these events, the objective of this study was to investigate the presence of multiple and cross-resistance to glyphosate, chlorimuron, and diclosulam in Uruguayan C. bonariensis biotypes. Additionally, the study aimed to evaluate the effectiveness of alternative herbicides in controlling these resistant biotypes.
2. Materials and Methods
2.1. Seed Collection and Preliminary Studies
Seeds from 46 Conyza spp. biotypes were collected in agricultural fields and pastures across various departments in Uruguay during the summer seasons of 2019/2020 and 2020/2021. These biotypes were gathered from fields where control failures were reported by farmers and agronomists after applying recommended doses of glyphosate and/or diclosulam and/or chlorimuron. Sampling in each field was conducted based on the distribution of Conyza plants. To ensure representative samples, approximately 1 hectare was surveyed in an inverted “V” pattern, starting 10 m inside the paddock from the crop edge. In each field, seeds from at least 10 Conyza plants were collected, stored in paper bags, air-dried at room temperature, and then stored under refrigerated conditions (4 °C).
A screening study was carried out using the collected seeds, with glyphosate, diclosulam, and chlorimuron each being applied at their recommended doses individually to all Conyza biotypes. From this initial study and the morphological characterization of biotypes as described by Ulzurrun et al. [8], a total of five C. bonariensis biotypes were selected for subsequent experiments (Table 1). However, it was not possible to select susceptible biotypes of C. bonariensis from this collection. Thus, a known susceptible biotype (biotype CB-I) from Rio Grande do Sul, Brazil, previously identified as susceptible (S) by Kaspary et al. [9], was used as a control. Surviving plants of the initial screening from each biotype, generation 0 (G0), were allowed to self-pollinate in order to increase homozygosity of generation 1 (G1). These seeds were stored in paper bags at ca. 4 °C for a minimum of three months to break dormancy before use in subsequent experiments. Dose–response experiments were conducted with G1 of the four biotypes for each herbicide. An untreated plant from each biotype of G1 was allowed to self-pollinate to produce generation 2 (G2), for which dose–response experiments were repeated.

Table 1.
Origin of Conyza bonariensis biotypes and characterization after the screening with glyphosate (GLY), diclosulam (DIC), and chlorimuron (CHL). INIA-LE, Colonia-Uruguay, 2023.
2.2. Dose–Response Experiments
Approximately 1.5 g of mature seeds from each biotype was immersed in water for 24 h to stimulate germination and then sown into plastic trays filled with a commercial substrate (BioTerra Sustrato®, Canelones, Uruguay). Following the emergence of the first true leaves, seedlings were transplanted into 0.3 L plastic pots containing a 3:1:1 mixture of soil, substrate, and vermiculite (v/v), with one plant per pot. Three separate herbicide dose–response assays were conducted for glyphosate, chlorimuron, and diclosulam. All experiments followed a randomized block design with four replications and were repeated under greenhouse conditions (25 ± 5 °C, 14/10 h light/dark cycle, and daily irrigation) in 2021 (G1-repetitions 1 and 2) and 2022 (G2-repetitions 1 and 2). For each herbicide, four C. bonariensis biotypes were tested: one susceptible (S), two putatively moderately resistant (MR) biotypes, and one putatively highly resistant (R) biotype. Herbicide treatments involved 8 doses, considering the recommended rate (1X) of 720 g ae ha−1 for glyphosate, 25.2 g ai ha−1 for diclosulam, and 50 g ai ha−1 for chlorimuron. Doses of 1/32; 1/16; 1/8; 1/2; 1; 2; and 4X were applied to S and MR biotypes, and 1/8; 1/2; 1; 2; 4; 8; and 16X to R biotypes. Applications were made on plants at the 4–6 leaf stage using a cabinet sprayer fitted with XR8010 nozzles, set to deliver 150 L ha−1 at 250 kPa.
Plant response (%) was assessed 28 days after herbicide application (DAA); visual scores were assigned to each experimental unit (a pot with one plant), where 0% indicated no damage, and 100% indicated complete plant death. Shoot dry mass (SDM) measurements were also conducted 28 DAA. Above ground biomass of each plant was harvested, placed in paper bags, oven-dried at 70 °C until constant weight was achieved, and then weighed. Relative SDM was calculated by comparing the weight of treated plants with their corresponding untreated controls (considered as 100%).
2.3. Alternative Herbicides for Managing Resistant C. bonariensis Biotypes
To explore management alternatives for resistant biotypes, an experiment was conducted using a completely randomized factorial design with a 6 × 8 arrangement. Plants from each of the six C. bonariensis biotypes (CB-I, CB-II, CB-III, CB-IV, CB-V, and CB-VI) were subjected to either a control treatment (unsprayed) or to treatments with various herbicides: imazethapyr, ammonium-glufosinate, paraquat, diquat, 2,4-D, dicamba, and saflufenacil (Table 2). Four replicates were also used for this study, which was repeated twice in a greenhouse from October to December 2022. Second generation (G2) seeds from the six C. bonariensis biotypes were grown, treated, and evaluated as previously described in Section 2.2.
Table 2.
Herbicide alternatives tested for managing glyphosate-, diclosulam-, and chlorimuron-resistant C. bonariensis biotypes. INIA-LE, Colonia-Uruguay, 2023.
2.4. Statistical Analysis
The data obtained from the dose–response experiments for each herbicide and generation were analyzed using analysis of variance (ANOVA) via the F-test (p < 0.05) with the Statistical Analysis System (SAS® OnDemand for Academics, 2022). Results from experiments conducted in different years showed no significant year effect, allowing for the pooling of data. Subsequently, the data were fitted to a three-parameter model using the SigmaPlot statistical program, version 10.0 (Systat Software, Inc., San Jose, CA, USA), according to the following equation:
where Y represents the dependent variable (either control % or dry mass); X denotes the independent variable (herbicide dose in g ae or ai ha−1); “a” is the upper limit; X050 is the herbicide dose (in g ae or ai ha−1) that reduces the dependent variable to 50% of the upper limit; and “b” describes the slope around X050. Using this equation, the following coefficients were calculated: the X50 value, representing the herbicide dose required for 50% control (C50) or a 50% reduction in SDM (GR50) for each biotype. Additionally, the resistance factor (RF) was calculated by dividing the C50 and GR50 values of resistant and susceptible biotypes, respectively (C50R/C50S and GR50/GR50S). The resistance factor quantifies how many times greater the dose needed to achieve 50% control or growth reduction in resistant biotypes is compared to that required for susceptible biotypes [10].
Y = a/(1 + exp (−(X − X050)/b))
The results regarding alternative herbicides for managing resistant C. bonariensis biotypes were analyzed by ANOVA using the Statistical Analysis System (SAS® OnDemand for Academics, 2022). The data showed no significant interaction between years, and thus were pooled for further analysis. Significant differences between treatments were declared after Tukey’s test at the 5% significance level.
3. Results
3.1. Confirmation of C. bonariensis Resistance to Glyphosate
The evaluation of resistance levels in generation 1 (G1) of C. bonariensis to glyphosate indicated that biotype CB-IV is highly resistant, while biotypes CB-II and CB-III exhibit moderate to mild resistance, and biotype CB-I is susceptible, as expected (Figure 1). Control and shoot dry mass (SDM) responses to varying doses of glyphosate confirmed significant differences among the tested biotypes (Figure 1A,B). Biotype CB-I was effectively controlled at doses around 100 g ha−1, whereas biotype CB-IV was not completely controlled even at the highest dose of 11,520 g ha−1. Biotypes CB-II and CB-III showed intermediate resistance, achieving 100% control with a glyphosate dose of 2880 g ha−1. Shoot dry mass reduction followed a similar pattern, with biotype CB-I being susceptible, CB-II and CB-III moderately resistant, and CB-IV highly resistant. The resistance factor (RF), based on SDM reduction, varied from 12.47 for biotype CB-III to 110.22 for biotype CB-IV (Figure 1B; Table 3).
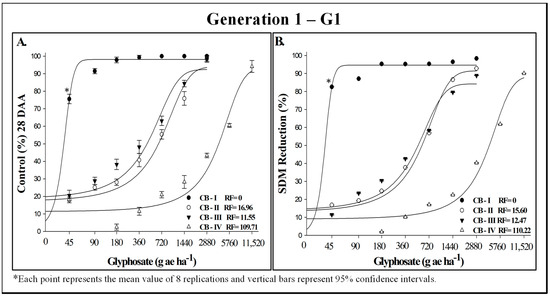
Figure 1.
Control (%) (A) and shoot dry mass (SDM) reduction (B) in generation 1 of putatively glyphosate-susceptible (CB-I) and -resistant (CB-II, CB-III, and CB-IV) Conyza bonariensis biotypes, evaluated 28 days after herbicide application. INIA-LE, Colonia-Uruguay, 2023.
Table 3.
Resistance factor (RF) and parameters of the log-logistic equation used to calculate glyphosate doses required for 50% control (C50) and 50% growth reduction (GR50) in generations 1 and 2 of putatively glyphosate-susceptible (CB-I) and -resistant (CB-II, CB-III, and CB-IV) C. bonariensis biotypes, evaluated 28 days after herbicide application (DAA). INIA-LE, Colonia, Uruguay, 2023.
Glyphosate dose–response experiments on generation 2 (G2) yielded similar results (Figure 2A,B). The RF values, derived from SDM reduction, were slightly higher for G2 compared to G1. Nonetheless, the resistance patterns remained consistent, with intermediate resistance in biotypes CB-II and CB-III, and high resistance in biotype CB-IV, exhibiting RF values of 19.00, 13.60, and 120.87, respectively (Figure 2A).
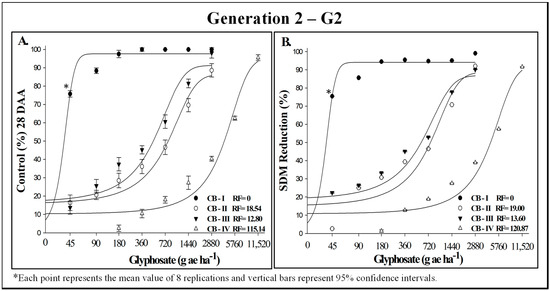
Figure 2.
Control (%) (A) and shoot dry mass (SDM) reduction (B) in generation 2 of putatively glyphosate-susceptible (CB-I) and -resistant (CB-II, CB-III, and CB-IV) Conyza bonariensis biotypes, evaluated 28 days after herbicide application. INIA-LE, Colonia-Uruguay, 2023.
The values for 50% control (C50) and 50% growth reduction (GR50) demonstrated marked differences among biotypes in both generations tested (Table 3). Biotype IV exhibited C50 and GR50 values exceeding 3700 g ha−1 in G1 and over 4000 g ha−1 in G2 for both variables. In contrast, C50 values for biotypes II and III ranged from 645.86 to 407.90, while GR50 values varied between 423.51 and 656.99 (Table 3). The application of logistic-type mathematical models provided an excellent fit to the data, with determination coefficients exceeding 92% (Table 3).
3.2. Confirmation of C. bonariensis Resistance to Diclosulam
The evaluation of resistance levels in G1 of C. bonariensis to the herbicide diclosulam indicated that biotype CB-IV is highly resistant, while biotypes CB-II and CB-V exhibit moderate resistance, and biotype CB-I is susceptible (Figure 3). The RFs calculated using C50 and GR50 values were similar. Based on SDM, CB-II, CB-V, and CB-IV were 2.89, 9.26, and 57.82 times more resistant to diclosulam than CB-I, respectively (Figure 3 and Table 4).
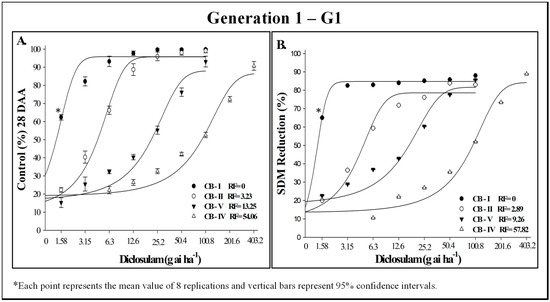
Figure 3.
Control (%) (A) and shoot dry mass (SDM) reduction (B) in generation 1 of putatively diclosulam-susceptible (CB-I) and -resistant (CB-II, CB-IV, and CB-V) C. bonariensis biotypes, evaluated 28 days after herbicide application. INIA-LE, Colonia, Uruguay, 2023.
Table 4.
Resistance factor (RF) and parameters of the log-logistic equation used to calculate diclosulam doses required for 50% control (C50) and 50% growth reduction (GR50) in generations 1 and 2 of putatively diclosulam-susceptible (CB-I) and -resistant (CB-II, CB-IV and CB-V) C. bonariensis biotypes, evaluated 28 days after herbicide application (DAA). INIA-LE, Colonia, Uruguay, 2023.
The biotype’s response to diclosulam in G2 was similar to that observed in G1 (Figure 4). Biotype CB-IV remained the most resistant, not being completely controlled even at a dose of 403.2 g ai ha−1. The data from the diclosulam dose–response experiments fitted well to the log-logistic model, yielding determination coefficients greater than 0.91 for both plant generations and across both variables of control and SDM reduction (Table 3). Regarding C50, GR50, and RFs values, results were consistent across generations for all biotypes (Table 4).
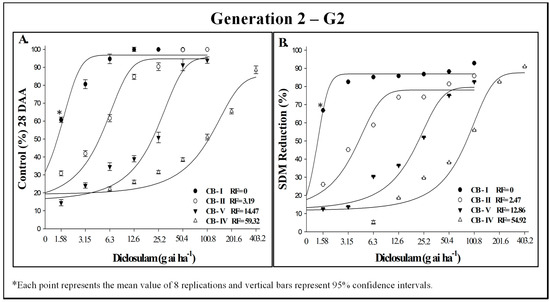
Figure 4.
Control (%) (A) and shoot dry mass (SDM) reduction (B) in generation 2 of putatively diclosulam-susceptible (CB-I) and -resistant (CB-II, CB-IV, and CB-V) C. bonariensis biotypes, evaluated 28 days after herbicide application. INIA-LE, Colonia, Uruguay, 2023.
3.3. Confirmation of C. bonariensis Resistance to Chlorimuron
The resistance of G1 C. bonariensis biotypes CB-III, CB-IV, and CB-VI to chlorimuron was evaluated, and compared with the susceptible control biotype CB-I (Figure 5). The three biotypes putatively resistant to chlorimuron, according to the initial screening, were not controlled with the recommended field rate of 50 g ai ha−1. Specifically, biotypes CB-III and CB-IV showed only 30% control at this rate. Based on SDM reduction data, CB-VI was classified as moderately resistant (RF 7.54), while biotypes CB-III and CB-IV were classified as highly resistant to chlorimuron (RFs 20.71 and 79.73, respectively) (Figure 5B).

Figure 5.
Control (%) (A) and shoot dry mass (SDM) reduction (B) in generation 1 of putatively chlorimuron-susceptible (CB-I) and -resistant (CB-III, CB-IV, and CB-VI) C. bonariensis biotypes, evaluated 28 days after herbicide application. INIA-LE, Colonia, Uruguay, 2023.
Resistance to chlorimuron was also confirmed in G2 C. bonariensis biotypes (Figure 6). The observed control levels and RFs were similar to those observed for G1. For biotype CB-IV, even the highest tested dose of chlorimuron, 800 g ai ha−1, did not result in complete plant death, achieving only approximately 80% control (Figure 6A). In terms of SDM reduction, biotypes CB-III and CB-IV again exhibited the highest resistance levels, with RFs of 21.52 and 89.07, respectively (Figure 6B).

Figure 6.
Control (%) (A) and shoot dry mass (SDM) reduction (B) in generation 2 of putatively chlorimuron-susceptible (CB-I) and -resistant (CB-III, CB-IV, and CB-VI) C. bonariensis biotypes, evaluated 28 days after herbicide application. INIA-LE, Colonia, Uruguay, 2023.
Data from both generations (G1 and G2) adequately fitted the log-logistic equation proposed, with R2 coefficients equal to or greater than 0.93 for the variables of control and SDM reduction (Table 5). The two most resistant biotypes (CB-III and CB-IV) consistently demonstrated high tolerance to chlorimuron, as evidenced by both control and shoot dry mass (SDM) reduction variables in both plant generations. For these biotypes, the C50 and GR50 values were higher than the recommended chlorimuron field application rate of 50 g ai ha−1, exceeding 200 g ai ha−1 for biotype CB-IV.
Table 5.
Resistance factor (RF) and parameters of the log-logistic equation used to calculate chlorimuron doses required for 50% control (C50) and 50% growth reduction (GR50) in generations 1 and 2 of putatively chlorimuron-susceptible (CB-I) and -resistant (CB-III, CB-IV, and CB-VI) C. bonariensis biotypes, evaluated 28 days after herbicide application (DAA). INIA-LE, Colonia, Uruguay, 2023.
3.4. Alternative Herbicides for Managing Resistant C. bonariensis Biotypes
Conyza bonariensis biotypes resistant to the tested herbicides (glyphosate, diclosulam, and chlorimuron) demonstrated significant differences in their response to treatment with alternative herbicides (Table 6 and Table 7). Among the resistant biotypes, the lowest control effectiveness was observed with the herbicide imazethapyr (an ALS inhibitor), particularly in biotype CB-IV, which exhibited only 22.75% control. A higher level of control, approximately 80%, was achieved for biotypes CB-III, CB-V, and CB-VI, while in biotypes CB-I and CB-II, plant death was nearly complete, exceeding 96%. (Table 6).
Table 6.
Evaluation of control (%) in Conyza bonariensis biotypes susceptible and resistant to glyphosate, diclosulam, and chlorimuron, treated with alternative herbicides. Assessment conducted 28 days after application. INIA-LE, Colonia, Uruguay, 2023.
Table 7.
Shoot dry mass (SDM) accumulation of Conyza bonariensis biotypes susceptible and resistant to glyphosate, diclosulam, and chlorimuron, treated with alternative herbicides. Assessment conducted 28 days after application. INIA-LE, Colonia, Uruguay, 2023.
Herbicides with mechanisms of action different from those to which the studied biotypes evolved resistance, such as ammonium-glufosinate, paraquat, diquat, and saflufenacil, demonstrated high control efficacy across all tested C. bonariensis biotypes, with control values exceeding 96%. However, the herbicide 2,4-D showed more limited control against biotypes CB-IV and CB-VI, with control rates of 70.25% and 90.50%, respectively. In contrast, dicamba, a Group 4 herbicide similar to 2,4-D, achieved control rates exceeding 97% for all tested biotypes (Table 6).
The SDM values (g plant−1) for C. bonariensis biotypes treated with various herbicides mirrored the control results (Table 7). After being sprayed with imazethapyr, biotype CB-IV’s dry matter accumulation was 1.985 g plant−1, which was not different from the untreated control. This biotype also showed intermediate dry matter accumulation after treatment with 2,4-D (1.356 g plant−1). The remaining herbicides tested demonstrated high levels of SDM reduction in plants from all C. bonariensis biotypes compared to the untreated control.
4. Discussion
The reliance on chemical control for managing C. bonariensis in Uruguayan production systems has led to the selection of biotypes that are now difficult to control with herbicides that were commonly effective in the past, such as glyphosate, diclosulam, and chlorimuron. A resistance factor (RF) above 100 in biotype CB-IV clearly demonstrates the high level of glyphosate resistance observed in this study. This outcome is consistent with the intense selection pressure resulting from the widespread and almost exclusive use of this herbicide for managing C. bonariensis within agricultural crops. This trend became particularly pronounced with the introduction of glyphosate-resistant soybean and corn cultivars, coupled with the widespread adoption of direct drilling practices reliant on this herbicide [4]. Glyphosate-resistant C. bonariensis biotypes have previously been reported in other countries of the region, such as Brazil [11,12,13] and Argentina [14], and more recently, in other parts of the world such as Colombia and South Africa [15,16]. Unlike the findings of this study, these reports generally indicate relatively low levels of glyphosate resistance, with RFs typically below 10. However, studies involving other Brazilian glyphosate-resistant C. bonariensis biotypes have reported RFs above 50 [9]. In the current study, high levels of glyphosate resistance were observed in both plant generations, confirming the heritability of this trait. This aspect is crucial for validating the selection of herbicide-resistant weed populations.
The mechanisms underlying herbicide resistance in these C. bonariensis biotypes might be associated with modifications at the glyphosate target site, specifically a mutation at position 106 of the EPSPS (P106S), as described in previous studies [17,18]. However, these studies link such DNA changes to cases of low-level resistance. Other research additionally suggests an increase in EPSPS enzyme activity and correlates these changes with biotypes that exhibit high-level glyphosate resistance [9,19]. Nevertheless, it is possible that non-target site mechanisms may also contribute to glyphosate resistance, including reduced absorption and translocation, vacuolar sequestration, and altered metabolism [16,20,21,22,23].
Dose–response experiments with acetolactate synthase (ALS)-inhibiting herbicides from different chemical families, diclosulam and chlorimuron, confirmed the occurrence of cross-resistance to these two herbicides in both plant generations (G1 and G2). Resistance to these herbicides emerged as ALS inhibitors have been extensively used to manage glyphosate resistance, leading to the evolution of populations with multiple and cross-resistance [24]. For C. bonariensis, no international reports of resistance to chlorimuron or diclosulam exist, nor are there known cases of cross-resistance to two ALS inhibitors or multiple resistance including glyphosate [2]. However, in C. sumatrensis (Sumatran fleabane), a species within the same genus as C. bonariensis, cases of multiple resistance to glyphosate and chlorimuron have already been documented in Brazil and Paraguay [1,25].
In a study involving more than 1100 populations of these two Conyza species from various soybean-producing regions in Brazil, no C. bonariensis accessions resistant to chlorimuron were identified. However, out of this total, 39.6% of the Sumatran fleabane populations were resistant to both glyphosate and chlorimuron [26]. Furthermore, a population of C. sumatrensis has already been confirmed to exhibit multiple resistance to five herbicide action sites: PSII inhibitors (paraquat), PS I Electron Diversion (diuron), protoporphyrinogen oxidase inhibition (saflufenacil), enolpyruvyl shikimate phosphate synthase inhibition (glyphosate), and auxin mimics (2,4-D) [27]. Additionally, there is a reported case of cross-resistance in C. sumatrensis to the ALS inhibitors chlorimuron-ethyl and cloransulam-methyl [28]. However, no cases of resistance to diclosulam have been reported in Conyza spp. [2].
Diclosulam and chlorimuron are both ALS inhibitors, but they belong to different chemical families: triazolopyrimidine-type 1 and sulfonylurea, respectively. The difference in chemical structure and intensity of use might contribute to the higher incidence of resistance reported for chlorimuron, which currently has 63 documented cases in various countries compared to only five for diclosulam [2]. However, in this study, a high level of resistance was observed in C. bonariensis, with an RF exceeding 50. The selection of diclosulam-resistant biotypes in Uruguay is likely attributed to the use of this herbicide as an alternative for managing Conyza spp. populations that are poorly controlled by glyphosate in pre- and post-emergent soybean cultivation [4]. The intensive use of flumetsulam, a herbicide belonging to the same family as diclosulam, may have also contributed to the selection pressure. Flumetsulam is by far the most frequently used herbicide for weed control in legume-containing sown pastures. Crop and pasture fields coexist across a significant portion of Uruguay’s agricultural area and Conyza spp. is a common weed of both type of fields. Interestingly, the evolution of multiple and cross-herbicide resistance to glyphosate, diclosulam, and chlorimuron in these C. bonariensis biotypes represents the first reported case for the species.
In agricultural systems, resistance at such a complex level poses significant challenges in managing this important weed species. Since C. bonariensis is a self-pollinating species, the dispersion and accumulation of resistance genes, potentially leading to complex resistance scenarios, would be expected to be slower than in outcrossing species [29]. However, its high seed production capacity (ca. 800,000 seeds per plant) [7], and its ability to disperse seeds effectively through wind could significantly contribute to the spread of biotypes with multiple herbicide resistances to previously unaffected areas. Despite this, the current study suggests that using herbicides with different mechanisms of action remains a viable control strategy for these biotypes. Similar effectiveness was observed in other studies where herbicides successfully controlled resistant Conyza spp. with up to six true leaves [30]. However, the herbicide imazethapyr proved ineffective in controlling biotype IV, which displays cross-resistance to diclosulam and chlorimuron. This pattern suggests a potential cross-resistance to imidazolinones as well, another chemical family within the ALS inhibitors. Moreover, difficulties in controlling some C. bonariensis biotypes with 2,4-D were noted in this study. Comparable results were observed in field experiments, where control efficacy at 28 days after application was around 80% [31]. Control levels improved when this herbicide was combined with saflufenacil or ammonium-glufosinate, offering management options for existing resistance cases [32].
In this context, implementing measures to mitigate and decelerate the selection process is critical [31]. These include the use of herbicides with different mechanisms of action, either in rotation or as tank mixes; sequential application in the pre- and post-emergence of crops and/or weeds; the integration of cover crops and other cultural strategies into herbicidal management plans; and the adoption of additional tools beyond chemical control. These practices are essential in preventing the selection of resistant weed biotypes and are fundamental to the sustainability of production systems.
5. Conclusions
Based on the results of this study, we can formally confirm the first case of cross-resistance in C. bonariensis to the herbicides diclosulam and chlorimuron (ALS inhibitors). This study also establishes the first case of both multiple and cross-resistance in C. bonariensis to glyphosate (an EPSPS inhibitor), diclosulam, and chlorimuron.
Biotypes of C. bonariensis exhibiting multiple and cross-resistance can, as of now, still be effectively controlled using herbicides with alternative mechanisms of action. However, integrating the use of these herbicides, in conjunction with other non-chemical management strategies, is critical for controlling these resistant biotypes and maintaining the sustainability of production systems.
Author Contributions
Conceptualization and investigation, T.E.K., M.E.W.B., S.M.H. and M.A.G.; formal analysis, T.E.K., M.E.W.B. and M.A.G.; data collection, T.E.K., M.E.W.B. and S.M.H.; writing—review and editing, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fondo Vaz Ferreira (Ministerio de Educación y Cultura-Uruguay)—Project identification: FVF/2021/013 and the National Agricultural Research Institute of Uruguay—INIA.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Albrecht, A.J.P.; Thomazini, G.; Albrecht, L.P.; Pires, A.; Lorenzetti, J.B.; Danilussi, M.T.Y.; Silva, A.F.M.; Adegas, F.S. Conyza sumatrensis resistant to paraquat, glyphosate and chlorimuron: Confirmation and monitoring the first case of multiple resistance in Paraguay. Agriculture 2020, 10, 582. [Google Scholar] [CrossRef]
- Heap, I. International Survey of Herbicide-Resistant Weeds. 2023. Available online: http://www.weedscience.org (accessed on 2 November 2023).
- Kissmann, K.G.; Groth, D. Plantas Infestantes e Nocivas, 2nd ed.; BASF: São Paulo, Brazil, 1999. [Google Scholar]
- Garcia, M.A.; Meneses, L.V.; Kaspary, T.E. Weed problems in uruguayan agriculture: Evolution and current situation. Outlooks Pest Manag. 2021, 32, 203–207. [Google Scholar] [CrossRef]
- Trezzi, M.M.; Vidal, R.A.; Patel, F.; Miotto, E.; Debastiani, F.; Balbinot, A.A.; Mosquen, R. Impact of Conyza bonariensis density and establishment period on soyabean grain yield, yield components and economic threshold. Weed Res. 2015, 55, 34–41. [Google Scholar] [CrossRef]
- Lorenzi, H. Manual de Identificação e Controle de Plantas Daninhas: Plantio Direto e Convencional, 7th ed.; Instituto Plantarum: Nova Odessa, Brazil, 2014. [Google Scholar]
- Kaspary, T.E.; Lamego, F.P.; Cutti, L.; Aguiar, A.C.d.M.; Rigon, C.A.G.; Basso, C.J. Growth, phenology, and seed viability between glyphosate-resistant and glyphosate-susceptible hairy fleabane. Bragantia 2017, 76, 92–101. [Google Scholar] [CrossRef]
- De Ulzurrun, P.D.; Acedo, M.B.; Garavano, M.E.; Gianelli, V.; Ispizúa, V.N. Morphological Characterization of Conyza blakei, Conyza bonariensis, Conyza sumatrensis and Conyza lorentzii in the Southeast of Buenos Aires (Argentina). Bol. Soc. Argent. Bot. 2018, 53, 359–373. [Google Scholar] [CrossRef]
- Kaspary, T.E.; Lamego, F.P.; Langaro, A.C.; Ruchel, Q.; Agostinetto, D. Investigation of the mechanism of resistance to glyphosate herbicide in hairy fleabane. Planta Daninha 2016, 34, 555–564. [Google Scholar] [CrossRef][Green Version]
- Burgos, N.R.; Tranel, P.J.; Streibig, J.C.; Davis, V.M.; Shaner, D.; Norsworthy, J.K.; Ritz, C. Review: Confirmation of resistance to herbicides and evaluation of resistance levels. Weed Sci. 2013, 61, 4–20. [Google Scholar] [CrossRef]
- Moreira, M.S.; Nicolai, M.; Carvalho, S.J.P.; Christoffoleti, P.J. Resistência de Conyza canadensis e C. bonariensis ao herbicida glyphosate. Planta Daninha 2007, 25, 157–164. [Google Scholar] [CrossRef]
- Vargas, L.; Bianchi, M.A.; Rizzardi, M.A.; Agostinetto, D.; Dal Magro, T. Buva (Conyza bonariensis) Resistente ao glyphosate na região sul do Brasil. Planta Daninha 2007, 25, 573–578. [Google Scholar] [CrossRef]
- Lamego, F.P.; Vidal, R.A. Resistance to glyphosate in Conyza bonariensis and Conyza canadensis biotypes in Rio Grande Do Sul, Brazil. Planta Daninha 2008, 26, 467–471. [Google Scholar] [CrossRef]
- Puricelli, E.; Faccini, D.; Metzler, M.; Torres, P. Differential susceptibility of Conyza bonariensis biotypes to glyphosate and als-inhibiting herbicides in Argentina. Agric. Sci. 2015, 6, 22–30. [Google Scholar] [CrossRef]
- Granados, E.; Zelaya, I.; Plaza, G. Widespread occurrence of glyphosate-resistant hairy fleabane (Erigeron bonariensis L.) in Colombia and weed control alternatives. Agronomy 2023, 13, 683. [Google Scholar] [CrossRef]
- Okumu, M.N.; Robbertse, P.J.; Vorster, B.J.; Reinhardt, C.F. The molecular, morphological and genetic characterization of glyphosate resistance in Conyza bonariensis from South Africa. Plants 2022, 11, 2830. [Google Scholar] [CrossRef] [PubMed]
- Powles, S.B.; Preston, C. Evolved glyphosate resistance in plants: Biochemical and genetic basis of resistance. Weed Technol. 2006, 20, 282–289. [Google Scholar] [CrossRef]
- Alarcón-Reverte, R.; García, A.; Urzúa, J.; Fischer, A.J. Resistance to glyphosate in junglerice (Echinochloa colona ) from California. Weed Sci. 2013, 61, 48–54. [Google Scholar] [CrossRef]
- Salas, R.A.; Dayan, F.E.; Pan, Z.; Watson, S.B.; Dickson, J.W.; Scott, R.C.; Burgos, N.R. EPSPS gene amplification in glyphosate-resistant italian ryegrass (Lolium perenne ssp. multiflorum) from Arkansas. Pest Manag. Sci. 2012, 68, 1223–1230. [Google Scholar] [CrossRef]
- Ferreira, E.A.; Galon, L.; Aspiazú, I.; Silva, A.A.; Concenço, G.; Silva, A.F.; Oliveira, J.A.; Vargas, L. Glyphosate translocation in hairy fleabane (Conyza bonariensis) biotypes. Planta Daninha 2008, 26, 637–643. [Google Scholar] [CrossRef]
- Ge, X.; D’Avignon, D.A.; Ackerman, J.J.H.; Collavo, A.; Sattin, M.; Ostrander, E.L.; Hall, E.L.; Sammons, R.D.; Preston, C. Vacuolar glyphosate-sequestration correlates with glyphosate resistance in ryegrass (Lolium spp.) from Australia, South America, and Europe: A 31P NMR Investigation. J. Agric. Food Chem. 2012, 60, 1243–1250. [Google Scholar] [CrossRef]
- Moretti, M.L.; Hanson, B.D. Reduced Translocation is involved in resistance to glyphosate and paraquat in Conyza bonariensis and Conyza canadensis from California. Weed Res. 2017, 57, 25–34. [Google Scholar] [CrossRef]
- Pan, L.; Yu, Q.; Han, H.; Mao, L.; Nyporko, A.; Fan, L.J.; Bai, L.; Powles, S. Aldo-keto reductase metabolizes glyphosate and confers glyphosate resistance in Echinochloa colona. Plant Physiol. 2019, 181, 1519–1534. [Google Scholar] [CrossRef]
- Peterson, M.A.; Collavo, A.; Ovejero, R.; Shivrain, V.; Walsh, M.J. The Challenge of herbicide resistance around the world: A current summary. Pest Manag. Sci. 2018, 74, 2246–2259. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.; Oliveira, R.S.; Constantin, J.; Francischini, A.C.; Osipe, J.B. Multiple resistance of Conyza sumatrensis to chlorimuron-ethyl and to glyphosate. Planta Daninha 2014, 32, 409–416. [Google Scholar] [CrossRef]
- Mendes, R.R.; Takano, H.K.; Netto, A.G.; Junior, G.J.P.; Cavenaghi, A.L.; Silva, V.F.V.; Nicolai, M.; Christoffoleti, P.J.; Junior, R.S.D.O.; De Melo, M.S.C.; et al. Monitoring glyphosate-and chlorimuron-resistant Conyza spp. populations in Brazil. An. Acad. Bras. Cienc. 2021, 93, e20190425. [Google Scholar] [CrossRef] [PubMed]
- Pinho, C.F.; Lourenço Leal, J.F.; dos Santos Souza, A.; de Oliveira, G.F.P.B.; de Oliveira, C.; Langaro, A.C.; Lopes Machado, A.F.; Christoffoleti, P.J.; Saes Zobiole, L.H. First evidence of multiple resistance of sumatran fleabane (Conyza sumatrensis (Retz.) E.Walker) to five- mode-of-action herbicides. Aust. J. Crop Sci. 2019, 13, 1688–1697. [Google Scholar] [CrossRef]
- Vital Silva, V.; Mendes, R.; Suzukawa, A.; Adegas, F.; Marcelino-Guimaraes, F.; Oliveira, R. A Target-site mutation confers cross-resistance to als-inhibiting herbicides in Erigeron sumatrensis from Brazil. Plants 2022, 11, 467. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.M.; Good-Avila, S.V. Macrophylogenetic analyses of the gain and loss of self-incompatibility in the Asteraceae. New Phytol. 2007, 173, 401–414. [Google Scholar] [CrossRef]
- Budd, C.M.; Soltani, N.; Robinson, D.E.; Hooker, D.C.; Miller, R.T.; Sikkema, P.H. Control of glyphosate resistant canada fleabane with saflufenacil plus tankmix partners in soybean. Can. J. Plant Sci. 2016, 96, 989–994. [Google Scholar] [CrossRef]
- da Silva, P.V.; de Barros, D.M.; Domingos, E.R.; Monquero, P.A.; Dias, R.d.C.; Vendruscolo, M.A. Control of hairy fleabane in sequential and pre-emergence applications in soybean crops. Rev. Caatinga 2023, 36, 748–756. [Google Scholar] [CrossRef]
- Cantu, R.M.; Albrecht, L.P.; Albrecht, A.J.P.; Silva, A.F.M.; Danilussi, M.T.Y.; Lorenzetti, J.B. Herbicide alternative for Conyza sumatrensis control in pre-planting in no-till soybeans. Adv. Weed Sci. 2021, 39, e2021000025. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).