Abstract
The oriental fruit moth, Grapholita molesta, is one of the most destructive orchard pests worldwide. Due to its behavior of boring into twigs and fruits, controlling the pest at the larval stage using insecticides is challenging. We studied the enemy species at the larval stage of G. molesta from seven fruit-growing areas by sampling peach shoots and fruits bored by its larvae. The occurrence and biological characteristics of the dominant enemy were verified using field investigation and laboratory experiments. The results showed that the dominant enemy species of larval G. molesta was identified as Therophilus festivus. The larval, cocoon, and pupal stages of T. festivus could not survive at 35 °C. The developmental durations of these three stages at 20 °C were significantly longer than those at 25 °C and 28 °C. Consecutive years of investigation in Xinxiang, China, indicated that T. festivus can parasitize G. molesta larvae collected at each generation. The parasitism rates in June (45.16%), July (63.50%), and August (48.37%) were significantly higher than those in April (15.27%). Most field research in China has focused on the release of parasitoids to control G. molesta eggs. Our study establishes a foundation for the control of G. molesta larvae using T. festivus.
1. Introduction
The oriental fruit moth, Grapholita molesta (Busck, 1916) (Lepidoptera: Tortricidae), a major orchard pest native to the Asian subcontinent, is found in many fruit-growing regions worldwide [1]. In China, it occurs in most fruit-growing regions except Tibet. It causes shoot dieback and fruit drop in apple, pear, peach, and other fruit trees of the family Rosaceae, thus leading to substantial economic losses [2]. At the adult stage, the main control methods are pheromone-mediated mating disruption and pesticides. At the egg stage, it can be controlled by species from the Trichogramma genus and insecticides. However, the larval stage is the main hazard stage, owing to the larvae being able to bore and feed on twigs and fruits. Neonate larvae typically bore into shoots or fruits within several hours of hatching and feed on the plant’s parts until the mature larvae emerge and pupate, leaving a short period for their effective control at the larval stage. Therefore, the control effect of insecticides on G. molesta larvae is limited. Thus, taking advantage of natural enemies is an effective strategy for the sustainable control of G. molesta larvae.
Using natural enemies to manage pests in agroecosystems is a safe and sustainable biological control approach. Parasitoid wasps are natural enemies of arthropod pests. However, the species of natural enemies of G. molesta vary worldwide. For example, in Japan, the parasitism rate of Lytopylus rufipes (Nees, 1814) (Hymenoptera: Braconidae) reached 91.5% [3]. This parasitic wasp can parasitize all instars of G. molesta and has been introduced to the United States to prevent and control G. molesta [4]. The United States has at least 130 species of native and introduced parasitic wasps that parasitize G. molesta. Among them, Macrocentrus ancylivorus (Rohwer, 1923) (Hymenoptera: Braconidae) is a dominant species for biocontrol applications. In Brazil, M. ancylivorus is one of the most abundant enemies in orchards and successfully colonizes hosts. However, its introduction was unsuccessful in Japan and Australia [5,6]. In China, parasitic natural enemies of G. molesta were found in Henan Province in the 1980s, including Eriborus sp. (Hymenoptera: Ichneumonidae), Trathala flavo-orbitalis (Cameron, 1907) (Hymenoptera: Ichneumonidae), and Phanerotoma planifrons (Nees, 1816) (Hymenoptera: Braconidae). In Jinzhong, Shanxi Province, the larval–pupal parasitoids Pimpla disparis (Viereck, 1911) (Hymenoptera: Ichneumonidae) and Elodia morio (Fallén, 1820) (Diptera: Tachinidae) were the dominant natural enemies of G. molesta [7]. In Baoding, Hebei Province, the natural enemy Therophilus festivus (Muesebeck, 1953) (Hymenoptera: Braconidae) was found, and the parasitism rate reaches 43% [8].
The aforementioned natural enemies in China were obtained from field sampling. However, further studies were not conducted. Most field research in China on the release of natural enemies to control G. molesta has focused on the genus Trichogramma, which has a wide range of hosts [9]. Species belonging to the Trichogramma genus are egg parasitoids of G. molesta. However, they are polyphagous and vulnerable to adverse weather conditions, pesticides, fungicides, and herbicides [10,11,12], which significantly impacts their control effect. Thus, identifying the strongly parasitic natural enemies of G. molesta at the larval stage will facilitate new strategies for preventing and controlling G. molesta via the introduction of parasitic wasps that exhibit different ecological traits.
Due to the difficulty in controlling G. molesta at the larval stage and the knowledge gaps regarding natural enemies at this stage, we examined the dominant natural enemy species at the larval stage from different fruit-growing regions and identified its biological characteristics and occurrence dynamics. This study aims to provide a biological control agent to target G. molesta larvae.
2. Materials and Methods
2.1. Investigation of the Total Parasitism Rates and the Dominant Natural Enemy Species at the Larval Stage of G. molesta
From 2018 to 2022, peach shoots bored by G. molesta larvae were sampled from peach orchards in seven cities or counties in five Provinces in China to investigate the total parasitism rates and the dominant natural enemies. Peach shoots were stored in plastic insect-rearing boxes (40 cm × 28 cm × 15 cm) at 28 °C and 75% relative humidity (RH) with a 15:9 (L:D) photoperiod. Two layers of gauze covered the boxes and served as a substrate for mature larvae that emerged from the peach shoots. Mature larvae on the gauze were collected, placed in Petri dishes (9 cm in diameter and 1.5 cm in height) containing two layers of gauze, and individually monitored daily until the parasitoid emerged. The total parasitism rate was calculated by dividing the number of larvae parasitized by all species of parasitoids by the total number of larvae collected [13].
2.2. Dominant Parasitoid Identification
The dominant parasitoid was identified using morphological features and molecular identification techniques. Morphological features of the adult parasitoids were captured using an ultra-depth-of-field three-dimensional microscope (VHX-6000, Keyence Corporation, Osaka, Japan) and identified at the species level per morphological diagnostic identification criteria [14] and the literature [8,15]. The molecular identification techniques used have been reported previously [16,17]. The total genomic DNA of the adult parasitoid was extracted using an insect DNA Kit (D0925-01, Omega Bio-Tek Inc, Norcross, GA, USA) per the manufacturer’s instructions. The mitochondrial cytochrome oxidase subunit 1 gene (mtDNA COI) was amplified using the primers LCO1490 (5′-GGTCAACAAATCATAAAGATATTGG-3′) and HCO2198 (5′-TAAACTTCAGGGTGACCAAAAAATCA-3′). PCR products were purified using a PCR purification kit (DR0101050, Zhejiang Easy-Do Biotechnology Co., Ltd., Hangzhou, China) and sequenced using Sanger sequencing. The COI gene sequence similarities of the dominant parasitoid were searched using the Basic Local Alignment Search Tool (BLAST, https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome, accessed on 6 May 2024).
2.3. Observation of Parasitic Mode and Life History of T. festivus
The parasitic mode of T. festivus was observed to determine whether the parasitic wasps exhibited larval–parasitic or egg–larval parasitism. The emerging male and female adults of T. festivus were placed in a plastic box (2000 mL) and fed 10% honey water. Two days later, eggs and young larvae of G. molesta that had bored into Fuji apples were exposed to T. festivus simultaneously. Subsequently, three days later, T. festivus was removed from the box and G. molesta eggs and young larvae were allowed to grow in the Fuji apples until mature larvae emerged. We monitored each G. molesta larva developed from eggs or young larvae daily until T. festivus emerged.
The generational life history of T. festivus was determined. Specifically, the newly hatched G. molesta larvae were placed on Fuji apples, which were then housed in a 120-mesh cage (40 cm × 40 cm × 40 cm) in a climate chamber at 28 ± 0.5 °C at 75 ± 5% relative humidity (RH) with a 16:8 h (L:D) photoperiod. Female and male T. festivus were then introduced to the cage. A week later, the apples were collected and placed in plastic insect-rearing boxes until mature G. molesta larvae emerged. The emergence of T. festivus from the mature G. molesta larvae and their life history were observed. Images were captured using the ultra-depth-of-field three-dimensional microscope.
2.4. Effect of Temperature on T. festivus Development
Suitable temperatures for T. festivus development were also investigated. The parasitized mature G. molesta larvae that emerged from peach shoots on the first day were collected, placed into the Petri dishes containing two layers of gauze, and individually monitored daily until the T. festivus larvae emerged. The pre-emergence period of T. festivus larvae was calculated at 28 °C. This period refers to the time from the initial emergence of parasitized mature G. molesta larvae from peach shoots to the first emergence of T. festivus larvae from the mature G. molesta larvae. The T. festivus larvae, pupae, and cocoons were individually placed at temperatures of 20 °C, 25 °C, 28 °C, and 35 °C to observe the effect of temperature on their survival rates. The temperatures used in the experiment were based on the temperature range of the naturally occurring season. Each treatment was repeated three times, with each repetition containing a minimum of five individuals. The developmental period of T. festivus larvae, pupae, and cocoons was recorded at temperatures of 20, 25, and 28 °C. Each treatment was repeated a minimum of ten times, with each repetition involving a single individual. These experiments were conducted under conditions of 75 ± 5% RH and a 16:8 h (L:D) photoperiod.
2.5. Successive Yearly Dynamics of T. festivus in Xinxiang Orchard
The control ability of T. festivus against G. molesta larvae was investigated in a peach orchard with extensive management. From 2021 to 2023, peach shoots bored by G. molesta larvae at each generation in the same peach orchard (approximately 4.7 ha, with 2- to 7-year-old peach trees) in Xinxiang were continuously collected and transported to the laboratory for breeding. The occurrence and dynamics of T. festivus in the orchard were studied. In addition, peach fruits and nearby apples bored by G. molesta larvae were also collected. The parasitism rates were calculated by dividing the number of larvae parasitized by T. festivus by the total number of larvae collected. The orchard was managed as follows: sex-attractant isotropic diffusers were hung in March and supplemented with pyrethroid insecticide or chlorantraniliprole for controlling G. molesta. Insecticide was sprayed once or twice per month, and as the fruits gradually ripened, the control was largely pesticide-free.
2.6. Statistical Analyses
Data were checked for normality and homogeneity of variance before the parametric analysis. Developmental durations for the larval, pupal, and cocoon stages of T. festivus at different temperatures were analyzed using a nonparametric analysis (Kruskal–Wallis test) because of the non-normality of the data. Mortality data for the larval, pupal, and cocoon stages of T. festivus at different temperatures were subjected to arcsine square root transformation and subsequently analyzed using a one-way analysis of variance (ANOVA) followed by the least significant difference (LSD) test. The parasitism rates of T. festivus to each generation of G. molesta larvae in the Xinxiang orchard were analyzed using one-way ANOVA followed by the LSD test. The parasitism rates of T. festivus to G. molesta larvae from peach shoots and ripe apples in the Xinxiang orchard were analyzed using a t test. All tests were performed using SPSS version 26 (IBM Corp., Armonk, NY, USA).
3. Results
3.1. Investigation of the Total Parasitism Rate and the Dominant Natural Enemies
Natural enemies were found in G. molesta larvae collected from the seven orchards. The total parasitism rate of natural enemies against G. molesta larvae from Laohekou, Jianning, Guixi, Dangshan, and Zhengzhou in April was less than 9%, while the parasitism rate in Xinxiang in July was up to 56.58%. Natural enemy species included one species of parasitic fly and five species of parasitic wasps. The parasitic fly and some species of parasitic wasp had very small populations in specific areas. Therophilus festivus had the greatest number (87.1%) of individuals in all orchards (Table 1).

Table 1.
Investigation of the total parasitism rate of natural enemies of Grapholita molesta among the geographical populations.
3.2. Morphological and Molecular Identification of the Dominant Parasitoid
Adult morphological characteristics of the dominant parasitoid were as follows:
Head. Antennae with 29–32 segments, filiform, blackish brown, length of scape 1.4–1.7 times its width, length of first, second, and penultimate segments 2.8–4.6, 2.5–3.0, and 1.6–2.5 times their width, respectively; mandible brown; palpi yellow; face setose, largely smooth and finely punctulate; clypeus largely smooth and moderately convex; frons with medial ridge; vertex and temple shiny and smooth.
Mesosoma. Pronotum largely smooth with carinae anteriorly and some punctures dorso-posteriorly; mesoscutum spaced punctulate; scutellum shiny and smooth; mesopleuron spaced punctulate below precoxal sulcus and the remainder shiny and largely smooth; metapleuron densely setose and dorsally rather coarsely punctate.
Wings. Translucent and infuscated; veins translucent or light brown; one pterostigma on approximately two-thirds of the costal margin of the forewing; forewing 3.2–3.7 mm; second submarginal cell narrow and petiolate; vein SR1 straight; vein r very short.
Legs. Length of hind leg femur, tibia, and basitarsus 3.2–5.5, 6.1–9.9, and 6.5–10.5 times their width, respectively; length of outer and inner spur of middle leg tibia 0.3–0.6 and 0.6–0.7 times the middle leg basitarsus, respectively; length of outer and inner spur of hind leg tibia 0.3–0.6 and 0.3–0.8 times the hind leg basitarsus, respectively; fore- and mid-legs (except coxae) yellow; coxa, femur, and end of tibia of hind legs black, basal half of hind tibia entirely white.
Body. Color black; length 3.6–4.8 mm; ovipositor sheath 3.3–4.5 mm. The characteristics of male and female adult wasps are similar except for ovipositors (Figure 1). The morphological characteristics are consistent with those of T. festivus [8,14,15].

Figure 1.
Morphological characteristics of Therophilus festivus: (A) female adult; (B) male adult, dorsal view; (C) antenna; (D) head, ventral view; (E) head, dorsal view; (F) compound eye; (G) wing; (H) foreleg; (I) middle and hind legs.
The length of the COI sequence was 599 bp (GenBank accession number: PP998445) and showed 99.33% similarity to the sequence of T. festivus (KF385868.1). Therefore, the dominant parasitic wasp was identified as T. festivus (Hymenoptera: Braconidae: Agathidinae).
3.3. Parasitic Mode and Generational Life History of T. festivus
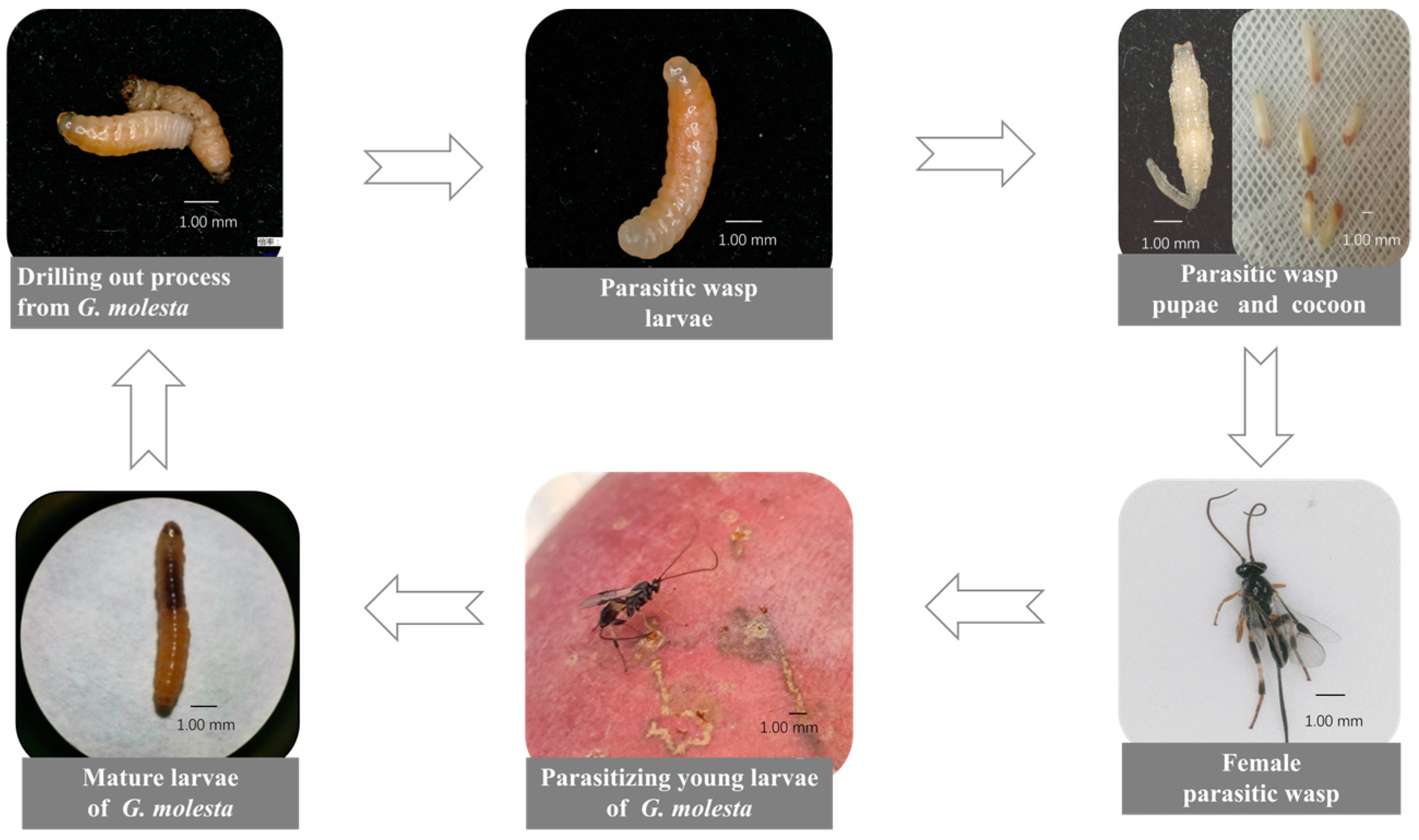
We showed that T. festivus can parasitize young G. molesta larvae without parasitizing the eggs, suggesting that T. festivus is a larval parasitoid rather than an egg–larval parasitoid. The parasitized young G. molesta larvae developed into mature larvae in the apples and then emerged from apples. Several days later, a larva of T. festivus emerged from the mature larva of G. molesta placed on the gauze and continued sucking until the mature larva died. Some T. festivus larvae formed cocoons on the gauze, while others pupated directly without cocoon formation owing to the artificial intervention: T. festivus larvae were moved to a smooth Petri dish immediately after they had emerged from the mature G. molesta larvae. Both the pupae and cocoons emerged normally at suitable conditions (Figure 2).
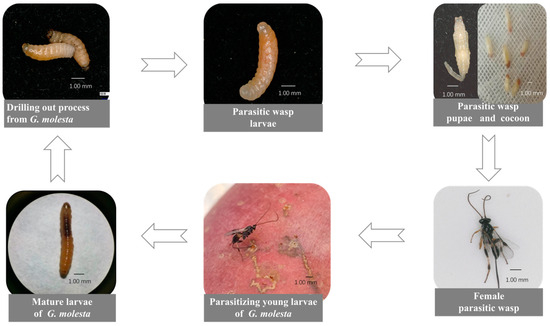
Figure 2.
Parasitism and acquisition process of Therophilus festivus.
3.4. Effect of Temperature on Developmental Duration of T. festivus
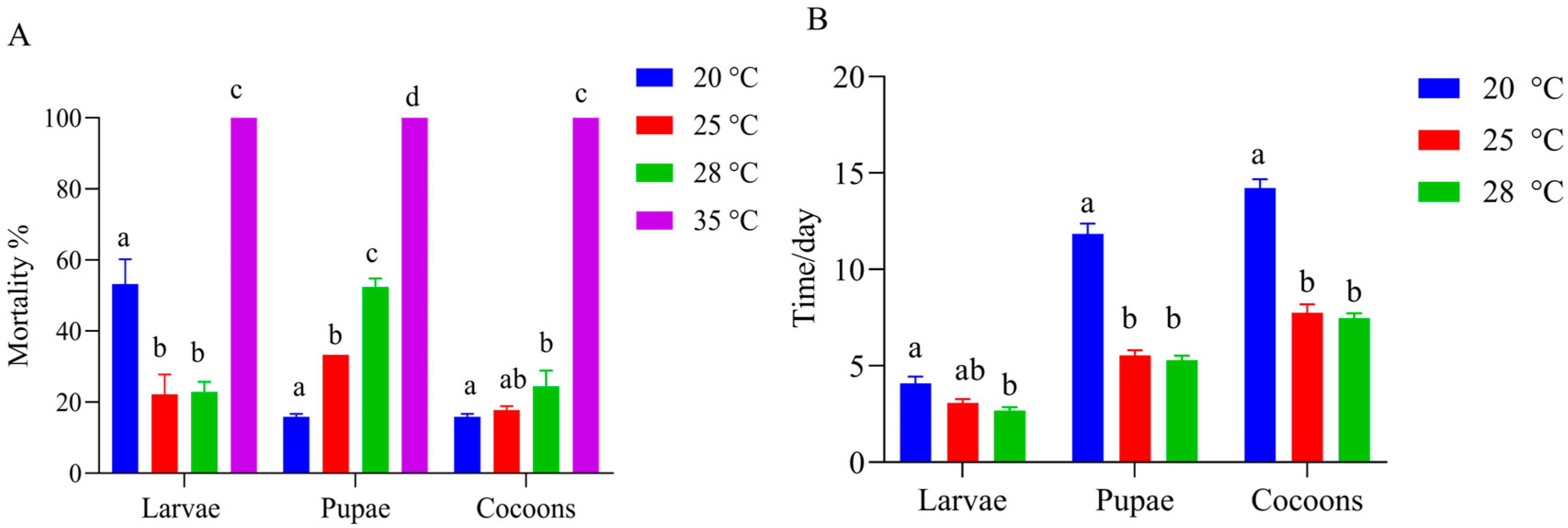
The results showed that T. festivus larvae emerged from the mature G. molesta larvae after an average of 4.76 d at 28 °C. At a temperature of 35 °C, there was 100% mortality at the larval, pupal, and cocoon stages. The mortality rate of larvae at 20 °C was significantly higher than that at 25 °C and 28 °C (20 °C vs. 25 °C, p = 0.006; 20 °C vs. 28 °C, p = 0.02), with no significant difference found between 25 °C and 28 °C. At the pupal stage, the mortality rate at 20 °C was markedly lower than that at 25 °C and 28 °C (20 °C vs. 25 °C, p = 0.005; 20 °C vs. 28 °C, p < 0.001). Furthermore, the mortality rate at 25 °C was also significantly lower than that at 28 °C (p = 0.002). There was no difference in cocoon mortality at 25 °C compared to that at both 20 °C and 28 °C. However, the mortality of cocoons at 20 °C was significantly lower than that at 28 °C (p = 0.025) (Figure 3A). The developmental durations of the three stages at 20 °C were significantly longer than those at 25 °C (larvae: p = 0.036; pupae: p < 0.0001; cocoons: p < 0.001) and 28 °C (larvae: p = 0.001; pupae: p < 0.001; cocoons: p < 0.001). No difference was observed between 25 °C and 28 °C (Figure 3B).
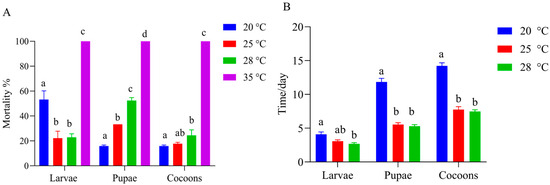
Figure 3.
Effect of temperature on Therophilus festivus development. (A) Effect of temperature on T. festivus mortality at the larval, pupal, and cocoon stages. At each stage, T. festivus individuals that failed to develop to the next stage at each temperature were defined as dead. Data represent the mean ± SE. Different lowercase letters at the same stage between different temperatures represent significant differences (LSD test, p < 0.05). (B) Effect of temperature on the developmental duration of T. festivus at the larval, pupal, and cocoon stages, respectively. Data represent the mean ± SE. Different lowercase letters at the same stage between different temperatures represent significant differences (Kruskal–Wallis test, p < 0.05).
3.5. Successive Years of Investigation of T. festivus in Xinxiang
The results of three consecutive years of investigation showed that larvae from the first generation (April) to the fourth generation (late August) were parasitized by T. festivus (Table 2). The parasitism rates in June, July, and August were significantly higher than those in April (June: p < 0.0001; July: p < 0.0001; August: p < 0.0001). In addition to peach shoots, T. festivus was found in the larvae sampled from young peach fruits (April) and mature apples (late August). The parasitism rate of T. festivus on G. molesta larvae from peach shoots was greater than that observed in ripe apples (p = 0.004) but less than that found in young peaches (p = 0.005).
Table 2.
Parasitism rate of Therophilus festivus to Grapholita molesta in Xinxiang orchard.
4. Discussion
We found several species of natural enemies of G. molesta larvae. Therophilus festivus was the dominant parasitic wasp, which had the largest population size and the highest parasitic rate. Therophilus festivus colonized hosts in all seven fruit-growing areas, while the other parasitoid species occurred in limited areas. Possible explanations for our findings are as follows: (i) Except for in Xinxiang, parasitoid species were only surveyed one or two times in the other locations without a seasonal survey, and the number of samples was insufficient; thus, species may have been missed in some areas. (ii) Some parasitoid species cannot colonize the hosts in some areas due to geographical locations, climate, and other conditions. Because insect phenologies mainly depend on temperature, with specific levels of accumulated degree days required for reaching each developmental stage, seasonal and geographical variations in temperature may largely affect the synchrony of the parasitoids with their hosts and, thus, their interaction level [18]. For instance, in contrast to Baoding, T. festivus was not detected in G. molesta from Jinzhong, where C. disparis and E. morio were the dominant parasitic wasps of G. molesta [7]. (iii) T. festivus may be a high-level specialist parasitoid of G. molesta, competing with other parasitoid species and contributing more to the mortality of G. molesta than generalist parasitoids [19]. (iv) Finally, management measures vary between orchards, including the type and schedule of insecticides that may harm certain parasitoid species [20].
Research on T. festivus has been limited to the specimen collection site, host species, and morphological description. The morphological features of our study’s dominant parasitoid of G. molesta are the same as those of T. festivus [8,15]. To further confirm the results, the dominant parasitoid was identified with molecular data from the COI gene. This gene has been previously applied to enhance species identification, such as the identification of Hymenoptera parasitoids in Tortricid fruit pests [16,17]. This is the first study to further confirm the species of T. festivus by molecular methods.
Therophilus festivus is understudied. We clarified its generational life history, revealed that it is a larval parasitoid, and identified the suitable temperature for development. However, the technology for breeding T. festivus in the laboratory remains lacking. For the further artificial release of T. festivus in orchards to control G. molesta, the large-scale rearing of T. festivus presents a pressing challenge. In the literature, the hosts of T. festivus include Grapholita prunivora (Walsh, 1868) (Lepidoptera: Tortricidae), G. molesta, Holocera pulverea (Meyrick, 1907) (Lepidoptera: Blastobasidae), Carposina niponensis (Walsingham, 1900) (Lepidoptera: Carposinadae), Haritalodes derogate (Fabricius, 1775) (Lepidoptera: Crambidae), Ostrinia furnacalis (Guenée, 1854) (Lepidoptera: Crambidae), Pectinophora gossypiella (Saunders, 1844) (Lepidoptera: Gelechiidae), Pseudohypatopa pulverea (Meyrick, 1907) (Lepidoptera: Blastobasidae), Helicoverpa armigera (Hübner, 1808) (Lepidoptera: Noctuidae), and Polyphagozerra coffeae (Nietner, 1861) (Lepidoptera: Cossidae) [15]. Further studies on the mature large-scale breeding technology using G. molesta and other hosts in the laboratory should be carried out to provide a theoretical basis for the application and release for controlling G. molesta in orchards.
Understanding the dynamics of natural enemies and host populations is a reference for developing biological control strategies. High parasitism capacity is a good criterion for selecting a parasitoid for a biological control program [21]. In peach shoots, except for the first-generation larvae in the orchard in Xinxiang with a specific parasitic rate of less than 16%, the parasitic rate of the remaining three larvae generations was more than 45%, which is similar to the results of Ran et al. [8]. Grapholita molesta undergoes five to six generations in the Xinxiang area, China, based on the law of effective accumulated temperature [22,23]. However, as the seasons change, the availability of new shoots in peach orchards decreases. Moreover, mature larvae of G. molesta begin their overwintering process in September, leading to a decline in the number of peach shoots damaged by larvae, making them increasingly difficult to collect. Consequently, we collected only four generations of larvae to investigate the parasitism rate. Our laboratory observations indicated that the adult stage of T. festivus is synchronized with the larval stage of G. molesta. Therefore, in practice, the timing of T. festivus occurrences can be predicted using predictive degree/day models for the development of G. molesta.
The low parasitism rate of first-generation larvae may be related to the size of the host populations, which limited the parasitism rate. Higher parasitism rates are reasonable as environmental suitability and the number of hosts increase. Host plant parts, manual interventions, and other factors may also affect the parasitism rate [18,24]. For example, the parasitism rate of T. festivus on G. molesta larvae sampled from peach shoots was lower than that observed in peach fruits but higher than that recorded in apple fruits. These variations may be attributed to the volatile compounds produced by peach shoots, peach fruits, and apple fruits during G. molesta feeding. Therefore, further investigation is required to verify the volatile compounds present in these three parts and their differential attractiveness to T. festivus. Manual intervention also affected the parasitism rate and the successful colonization of parasitic wasp populations, mainly because heavy use of pesticides in orchards adversely affects parasitic wasps. Pesticide treatment timing can greatly impact parasitoid populations and should be timed to avoid the most vulnerable stages of parasitoids, including T. festivus. In addition, the host may influence the parasitism rate. Notably, G. molesta is not the only host of T. festivus, as C. niponensis serves as another host for this wasp [15]. Carposina niponensis can cause damage in peach orchards, surrounding apple orchards, and other types of orchards. Therefore, the presence of alternative hosts may also impact the parasitism rate of T. festivus against G. molesta. However, field experiments are necessary to further confirm the parasitism of this wasp on other hosts.
Trichogramma genus species are promising natural enemies for G. molesta control [9,25,26], especially Trichogramma dendrolimi (Matsumura, 1926) (Hymenoptera: Trichogrammatidae). For each generation of G. molesta, releasing 900,000 T. dendrolimi per hectare in three batches, three days apart, with the distribution of release points spaced 10 m apart, can reduce fruit damage by half in an orchard [9]. Although T. dendrolimi application in orchards is viable for reducing G. molesta populations and fruit damage, a previous study showed that its combined application with Beauveria bassiana (Bals.-Criv.) Vuill. had complementary impacts on G. molesta [25]. Our study provided another compatible control tactic against G. molesta. Thus, the combined release of T. dendrolimi at the egg stage and T. festivus at the larval stage, along with the application of sex pheromones at the adult stage of G. molesta [27,28], may be effective in reducing G. molesta populations and fruit damage. To enhance the efficacy of biocontrol, the combined application of multiple biological control agents is often recommended. However, the simultaneous release of multiple biological control agents might influence their individual efficacy and pose potential risks related to competition. For example, entomopathogenic fungi can be infectious to beneficial insects such as parasitoids, adversely affecting their oviposition behavior or reducing the fitness of surviving wasp offspring [29]. In our study, Trichogramma wasps complete development within host eggs, preventing the eclosion of the damaging larval stage and reducing host population densities. Therophilus festivus develops within host larvae, inhibiting the emergence of the adult stage; they parasitize G. molesta at different developmental stages. Although releasing Trichogramma wasps may decrease G. molesta larval populations in fields, it does not affect the survival or population of T. festivus, as these wasps have alternative hosts available. Similarly, while releasing T. festivus may reduce the adult G. molesta population, which could result in decreased egg production, it does not affect the survival or population of Trichogramma wasps owing to their access to other hosts. Both Trichogramma wasps and T. festivus are effective biological control agents for G. molesta and have significant potential for joint application to improve overall efficacy.
Although our study did not investigate other management factors on T. festivus, the effectiveness of parasitoids is likely to increase if other management practices are incorporated into an integrated pest management program. For instance, the introduction of T. dendrolimi and T. festivus during the egg and larval stages of G. molesta can serve as an effective strategy for their control. Concurrently, it is essential to minimize insecticide usage in the field and avoid applications during peak activity periods of parasitic wasps. This approach not only enhances the efficacy of sustainable control measures for G. molesta but also aids in delaying the development of insecticide resistance within this pest population. Future research should select pesticides that exhibit low toxicity toward T. festivus while maintaining high toxicity against G. molesta for optimizing pest management strategies.
5. Conclusions
In summary, we identified T. festivus as the dominant natural enemy at the larval stage of G. molesta. We also revealed its biological characteristics and efficiency in controlling G. molesta in an orchard in Xinxiang. Therophilus festivus has been adapted to different geographical environments and has substantial potential for application. Therefore, further research is required before parasitic wasps can be successfully applied in the field (e.g., large-scale breeding technology and field-release technology of T. festivus). Our research provides a theoretical basis for controlling G. molesta larvae by the use of parasitic wasps.
Author Contributions
X.W.: conceptualization, writing—original draft, validation, project administration, investigation, data curation. Q.L.: investigation, methodology. J.W.: writing—review and editing, software. L.W.: writing—review and editing, validation. H.T.: writing—review and editing, supervision, funding acquisition. J.Z.: conceptualization, supervision, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research is co-supported by the Central Public-interest Scientific Institution Basal Research Fund (No. 1610192023301), the Scientific and Technological Project of Henan Province (No. 232102110070), the Central Public-interest Scientific Institution Basal Research Fund (No. 1610192023105), the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-2021-RIP-05), and the Agricultural Science and Technology Innovation Program, Chinese Academy of Agricultural Sciences (CAAS-ASTIP-2024-ZFRI-03).
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kirk, H.; Dorn, S.; Mazzi, D. Worldwide population genetic structure of the oriental fruit moth (Grapholita molesta), a globally invasive pest. BMC Ecol. 2013, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.T.; Hull, L.A.; Krawczyk, G. Comparative survival rates of oriental fruit moth (Lepidoptera: Tortricidae) larvae on shoots and fruit of apple and peach. J. Econ. Entomol. 2006, 99, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Hoshi, H.; Kawaguchi, E. Seasonal prevalence of occurrence of Lytopylus rufipes parasitic on larvae of oriental fruit moth, Grapholita molesta in peach orchards. Ann. Rept. Plant Prot. North Jpn. 2015, 66, 153–156. [Google Scholar]
- Liu, C.; Kainoh, Y. Laboratory rearing of Lytopylus rufipes (Hymenoptera: Braconidae: Agathidinae), a parasitoid wasp of the oriental fruit moth, Grapholita molesta (Lepidoptera: Tortricidae), using apple and a commercially available diet. Appl. Entomol. Zool. 2020, 55, 271–276. [Google Scholar] [CrossRef]
- Bailey, P. An attempt to control oriental fruit moth, Cydia molesta busck. by mass releases of Macrocentrus ancylivorus rohwer (Hymenoptera: Braconidae). Aust. J. Entomol. 1980, 18, 211–212. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Garcia, M.S.; Nava, D.E.; Botton, M.; Parra, J.R.; Guerrero, M. Selection of Trichogramma pretiosum lineages for control of Grapholita molesta in peach. Fla. Entomol. 2011, 94, 398–403. [Google Scholar] [CrossRef]
- Zhang, C.; You, X.Y.; Zhao, L.L.; Meng, H.; Ma, R.Y. Investigation on parasitic insect natural enemies of oriental fruit moth in Jinzhong. J. Shanxi Agric. Sci. 2012, 40, 389–392. (In Chinese) [Google Scholar]
- Ran, H.F.; Ma, A.H.; Liu, W.X.; Lu, Z.Y.; Li, J.C. Therophilus festivus (muesebeck, 1953)—A larval parasitoid wasp of oriental fruit moth: Morphology, biological characteristics and biological control potential. Chin. J. Biol. Control 2019, 35, 173–179. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, J.; Tang, R.; Fang, H.; Liu, X.; Michaud, J.P.; Zhou, Z.; Zhang, Q.; Li, Z. Laboratory and field studies supporting augmentation biological control of oriental fruit moth, Grapholita molest (Lepidoptera: Tortricidae), using Trichogramma dendrolimi (Hymenoptera: Trichogrammatidae). Pest Manag. Sci. 2021, 77, 2795–2803. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, X.; Huang, X.; Yu, X.; Zhang, W.; Zhang, X.; Mu, W. Comparative ecotoxicity of neonicotinoid insecticides to three species of Trichogramma parasitoid wasps (Hymenoptera: Trichogrammatidae). Ecotoxicol. Environ. Saf. 2019, 183, 109587. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, X.; Zhang, Z.; Liu, F.; Mu, W. Lethal and sublethal impact of sulfoxaflor on three species of Trichogramma parasitoid wasps (Hymenoptera: Trichogrammatidae). Biol. Control 2019, 134, 32–37. [Google Scholar] [CrossRef]
- Rakes, M.; Pasini, R.A.; Morais, M.C.; Araújo, M.B.; de Bastos Pazini, J.; Seidel, E.J.; Bernardi, D.; Grützmacher, A.D. Pesticide selectivity to the parasitoid Trichogramma pretiosum: A pattern 10-year database and its implications for integrated pest management. Ecotoxicol. Environ. Saf. 2021, 208, 111504. [Google Scholar] [CrossRef] [PubMed]
- Koutsogeorgiou, E.I.; Moysiadis, T.; Fifis, G.T.; Gogolashvili, N.E.; Chatzimpalasis, D.; Andreadis, S.S. Age- and density-dependent parasitism rate and development time of the generalist egg-parasitoid Ooencyrtus telenomicida (Hymenoptera: Encyrtidae) on eggs of the brown marmorated stink bug Halyomorpha halys. Insects 2024, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Van Achterberg, C.; Long, K.D. Revision of the Agathidinae (Hymenoptera, Braconidae) of Vietnam, with the description of forty-two new species and three new genera. ZooKeys 2010, 54, 1. [Google Scholar] [CrossRef]
- Gupta, A.; Sanjeev, U. First report of larval parasitism of Ethmia nigroapicella saalmüller (Lepidoptera: Gelechioidea: Ethmiidae) by Therophilus festivus (Muesebeck) (Braconidae: Agathidinae) in India. J. Biol. Control 2016, 30, 106–108. [Google Scholar] [CrossRef]
- Franck, P.; Maalouly-Matar, M.; Olivares, J. Molecular tools for the detection and the identification of Hymenoptera parasitoids in Tortricid fruit pests. Int. J. Mol. Sci. 2017, 18, 2031. [Google Scholar] [CrossRef]
- Wan, W.J.; Du, S.J.; Hansson, C.; Liu, W.X. A new species of Diglyphus Walker (Hymenoptera, Eulophidae) from China, with morphological characterizations and molecular analysis. ZooKeys 2023, 1148, 65. [Google Scholar] [CrossRef]
- Maalouly, M.; Franck, P.; Lavigne, C. Temporal dynamics of parasitoid assemblages parasitizing the codling moth. Biol. Control 2015, 82, 31–39. [Google Scholar] [CrossRef]
- Elzinga, J.A.; van Nouhuys, S.; van Leeuwen, D.J.; Biere, A. Distribution and colonisation ability of three parasitoids and their herbivorous host in a fragmented landscape. Basic Appl. Ecol. 2007, 8, 75–88. [Google Scholar] [CrossRef]
- Schindler, B.; Gavish-Regev, E.; Keasar, T. Parasitoid wasp community dynamics in vineyards following insecticide application. Front. Environ. Sci. 2022, 9, 785669. [Google Scholar] [CrossRef]
- Parra, J.; Coelho, A.J. Insect rearing techniques for biological control programs, a component of sustainable agriculture in Brazil. Insects 2022, 13, 105. [Google Scholar] [CrossRef] [PubMed]
- Li, W.L.; Li, D.X.; Dong, J.F.; Feng, Y.Q.; Li, Y. The research of threshold temperature and effective accumulated temperature of oriental fruit moth, Grapholita molesta Busck. J. Agric. Sci. 2010, 10, 80–82. (In Chinese) [Google Scholar]
- Zhou, H.R.; Liu, Q.; Tu, H.T. Synergistic trapping effect of mixed lure traps for the main Lepidopteran pests in peach orchards. J. Fruit Sci. 2024, 41, 1181–1187. (In Chinese) [Google Scholar]
- Tan, M.; Wu, H.; Li, Y.; Zhang, A.; Xu, J.; Chai, R.; Meng, Z.; Yan, S.; Jiang, D. Cadmium exposure through the food chain reduces the parasitic fitness of Chouioia cunea to Hyphantria cunea pupae: An ecotoxicological risk to pest control. Sci. Total Environ. 2023, 887, 164106. [Google Scholar] [CrossRef]
- Wu, Y.; Fang, H.; Liu, X.; Michaud, J.P.; Xu, H.; Zhao, Z.; Zhang, S.; Li, Z. Laboratory evaluation of the compatibility of Beauveria bassiana with the egg parasitoid Trichogramma dendrolimi (Hymenoptera: Trichogrammatidae) for joint application against the oriental fruit moth Grapholita molesta (Lepidoptera: Tortricidae). Pest Manag. Sci. 2022, 78, 3608–3619. [Google Scholar] [CrossRef]
- Zhao, J.; Liang, M.; Wang, Z.; Zhao, Y.; Cheng, J.; Du, Y. Evaluation and optimization of blends for attracting Trichogramma dendrolimi based on semiochemicals mediating tritrophic interactions in the orchard habitat. Biol. Control 2022, 173, 104998. [Google Scholar] [CrossRef]
- Tollerup, K.E.; Ann, R.; Shearer, P.W. Whole-farm mating disruption to manage Grapholita molesta (Lepidoptera: Tortricidae) in diversified new jersey orchards. J. Econ. Entomol. 2012, 105, 1712–1718. [Google Scholar] [CrossRef]
- Kong, W.; Wang, Y.; Guo, Y.; Chai, X.; Niu, G.; Li, Y.; Ma, R.; Li, J. Effects of disruption of Grapholita molesta (Lepidoptera: Tortricidae) using sex pheromone on moth pests and insect communities in orchards. Appl. Entomol. Zool. 2020, 55, 367–377. [Google Scholar] [CrossRef]
- Araujo, E.S.; Poltronieri, A.S.; Poitevin, C.G.; Mirás-Avalos, J.M.; Zawadneak, M.A.C.; Pimentel, I.C. Compatibility between entomopathogenic fungi and egg parasitoids (Trichogrammatidae): A laboratory study for their combined use to control Duponchelia fovealis. Insects 2020, 11, 630. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).