Simple Summary
As a promising substitute for chemical insecticides, β-asarone has a broad application prospect in the field of crop protection. In order to improve the insecticidal activity of β-asarone, a series of derivatives were prepared by active substructure splicing strategy, and their insecticidal activities were evaluated. Compound 10, containing the 2-chloropyridine functional group, had the strongest insecticidal activity against the small brown planthopper (SBPH), Laodelphax striatellus (Fallén) (Hemiptera: Delphacidae), which was 8.31 times higher than that of β-asarone. The ABC transporter gene MDR49 might be a candidate insecticidal target of compound 10. Furthermore, compound 10 showed excellent efficacy against Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) and Tetranychus cinnabarinus (Boisduval) (Acarina: Tetranychidae).
Abstract
The small brown planthopper (SBPH), Laodelphax striatellus (Fallén) (Hemiptera: Delphacidae), is an increasing threat to Gramineae crops, posing significant risks to both the environment and food safety. β-asarone, as a promising green alternative to chemical insecticides, possesses wide application prospects in the crop protection field. To enhance the insecticidal activity of β-asarone, a series of derivatives were prepared through an active substructure splicing strategy, and their insecticidal activities against SBPH were evaluated. Among the 7 commercial compounds with chemical structures similar to β-asarone and 12 structural derivatives of β-asarone, compound 10, which incorporates the 2-chloropyridine functional group from flupyrimin, exhibited the most potent insecticidal activity against SBPH, with an 8.31-fold increase in insecticidal activity compared to β-asarone. Furthermore, transcriptome analysis showed that among the selected genes that may play important roles in insecticidal activity, an ABC transporter gene, MDR49, was most significantly down-regulated. MDR49 was highly expressed in the 4th-instar nymphs, with the highest expression level in the fat body, midgut, and abdomen. RNA interference (RNAi) against MDR49 significantly reduced susceptibility to compound 10 in SBPH, which revealed that MDR49 may be the candidate insecticidal target of compound 10. Additionally, the insecticidal spectrum revealed that compound 10 showed excellent efficacy against Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) and Tetranychus cinnabarinus (Boisduval) (Acarina: Tetranychidae). This study indicates that compound 10 could be further developed as a novel eco-friendly pesticide.
1. Introduction
The small brown planthopper (SBPH), Laodelphax striatellus (Fallén) (Hemiptera: Delphacidae), is one of the most destructive pests of Gramineae crops, such as rice, maize, and wheat in Asia, North Africa, and the warm temperate regions of Europe [1,2]. SBPH can cause a devastating disaster to agricultural production by sucking the crop phloem and spreading various plant viruses, such as rice stripe virus (RSV) and rice black-streaked dwarf virus (RBSDV) [3,4,5]. Although commercial chemical insecticides have been widely used to manage SBPH, several major issues have emerged. Firstly, the long-term usage of current commercial chemical insecticides leads to cross-resistance and poor control effectiveness. Additionally, environmental pollution and chemical residues contradict the principle of protecting the environment and ensuring food safety [6,7]. Hence, it is urgent to develop novel, effective, and environmentally friendly insecticides for sustainable pest management. Natural product-based agrochemicals offer significant advantages, including species-specific targeting, unique modes of action, and low mammalian toxicity. Furthermore, these compounds tend to decompose rapidly, minimizing their environmental impact. Plant-derived insecticidal substances thus provide a rich resource for discovering and developing promising safe insecticides.
Acorus calamus is a medicinal plant with pesticidal, antifungal, and antibacterial properties in Asia [8,9]. β-asarone, which is isolated and purified from A. calamus, has toxicity to several pests, such as Plutella xylostella (Linnaeus) (Lepidoptera: Plutellidae), Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae), Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae), Nilaparvata lugens (Stal) (Hemiptera: Delphacidae), and Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) [10,11,12,13,14]. Although β-asarone demonstrates broad-spectrum insecticidal activity, its efficacy is lower than that of commercial chemical insecticides. In the N. lugens susceptible strain, the median lethal dose (LD50) of β-asarone was 0.187 μg/nymph [15]. However, the LD50 of triflumezopyrim, which is a novel meso-ionic or zwitterionic chemical insecticide, was 2.129 × 10−4 μg/nymph in N. lugens [16]. Likewise, the median lethal concentration (LC50) of β-asarone and cyantraniliprole against B. tabaci was 15.51 and 1.57 mg/L, respectively [14]. Therefore, enhancing the insecticidal activity of botanical compounds is of great practical significance for their broader application in agricultural production.
In this study, β-asarone and seven commercial compounds with chemical structures similar were assembling active functional groups from highly effective chemical insecticides (cyantraniliprole, chlorantraniliprole, flubendiamide, flupyrimin, sulfoxaflor, and triflumezopyrim) through an active substructure splicing strategy to obtain 12 derivatives of β-asarone. Through toxicity assays, we evaluated the insecticidal activity of 19 β-asarone analogs and derivatives against the SBPH. The compound with the highest insecticidal efficacy (Compound 10) was further analyzed for its mechanism of action and insecticidal spectrum.
2. Materials and Methods
2.1. Insects and Reagents
A total of 6 species of insects were used in the study. The SBPH was the main insect tested in this study, and the other 5 insects, including Semiaphis heraclei (Takahashi) (Hemiptera: Aphididae), Aphis craccivora Koch (Hemiptera: Aphididae), B. tabaci, Rhopalosiphum maidis (Fitch) (Hemiptera: Aphididae), and Tetranychus cinnabarinus (Boisduval) (Acarina: Tetranychidae), were mainly used to verify the insecticidal activity of compound 10. The SBPH strain was originally collected from Haian (32.57° N, 120.45° E; Jiangsu Province, China) and maintained in the laboratory with rice seedlings. The S. heraclei was collected from Pingyi (35.60° N, 117.75° E; Shandong Province, China) and fed with celery. The A. craccivora was collected from Linqing (36.68° N, 115.72° E; Shandong Province, China) and maintained in the laboratory with broad beans. The B. tabaci was picked from cotton in our greenhouses. The R. maidis and the T. cinnabarinus were presented by the Institute of Plant Protection, Shandong Academy of Agricultural Sciences. All the insects were reared in the laboratory under the conditions of 25 ± 1 °C, 70–80% humidity, 16 h light/8 h dark photoperiod.
β-asarone (compound 1), α-asaron (compound 2), asarylaldehyde (compound 3), cis-Anethol (compound 4), eugenol (compound 5), isoeugenol (compound 6), asaronic acid (compound 7), and pterostilbene (compound 8) were all purchased from Bide Pharmatech Ltd. (Shanghai, China) with a purity above 99%.
2.2. Modification and Synthesis of Active Compounds
Based on the chemical structure of β-asarone, its commercially available structural analogs (compounds 2–8) were selected. Additionally, using the active substructure splicing strategy, active functional groups from highly effective chemical insecticides (cyantraniliprole, chlorantraniliprole, flubendiamide, flupyrimin, sulfoxaflor, and triflumezopyrim) were assembled onto the framework of similar chemical structure compounds to β-asarone, leading to the synthesis of a series of β-asarone derivatives (compounds 9–20). The synthetic procedures are shown in the Supporting Information. Column chromatography was generally performed on a silica gel (200–300 mesh) gradient elution with petroleum ether and ethyl acetate, and TLC inspections on silica gel GF-254 plates with petroleum ether: ethyl acetate. Both 1H and 13C NMR spectra were recorded on a Bruker DP-X500 MHz spectrometer (Bruker Corporation, Beijing, China). Chemical shifts were reported in ppm relative to internal tetramethylsilane for 1H NMR (7.260 ppm) and CDCl3 (77.16 ppm) for 13C NMR. High-resolution mass spectra (HRMS) were collected on Waters LCT Premier™ with an ESI mass spectrometer (Waters Corporation, Beijing, China). The melting points were measured on a STUART-SMP3 Melt-Temp apparatus (Stuart Equipment, Beijing, China) without correction.
2.3. Toxicity Bioassay
The LD50 of each compound was tested with the 3rd-instar nymphs of SBPH. The bioassay of β-asarone and 19 other compounds was performed according to the topical application method [17], and each compound was diluted with acetone to generate five concentrations. The nymphs were anesthetized with ice, and then a 0.05 μL liquid drop solution was applied topically to the prothorax notum of the nymphs by a MICRO2T SMARTouchTM device (SMARTouchTM Controller for UMP3/Nanoliter 2010, World Precision Instruments, lnc., Sarasota, FL, USA). The nymphs treated with acetone were employed as the control. Each treatment contained thirty insects and was repeated three times. The mortality was checked 72 h after treatment.
The insecticidal activities of compound 10 against S. heraclei, A. craccivora, B. tabaci, R. maidis, and T. cinnabarinus were detected. The compound 10 was diluted with acetone to generate five concentrations. The wingless adults of S. heraclei, A. craccivora, and R. maidis of the same size were selected as the test insects. After treatment, they were fed on leaves of celery, broad beans, and corn, respectively. Adult B. tabaci and T. cinnabarinus with the same development stage and similar size were selected as experimental insects, and fed with cotton and kidney beans after compound 10 treatment. All treatments consisted of three replicates, each containing 30 test insects, and mortality was measured 72 h after treatment.
2.4. Sample Collection
2.4.1. Sample Collection for Transcriptome Sequencing and Quantitative Validation
The 3rd-instar nymphs were treated with LD50 dose (0.051 μg/nymph) of compound 10, and the survival insects were collected at 72 h. The sample was immediately frozen in liquid nitrogen and then stored at −80 °C for transcriptome sequencing and quantitative validation. Each treatment contained three biological replications, and each replication contained thirty insects.
2.4.2. Sample Collection for Instar Expression Profile and Tissue Expression Profile
The 1st- to 5th-instar nymphs, the adults of female and male of SBPH were collected for instar expression profile analysis. For the tissue expression profile, the tissues of the head, thorax, abdomen, midgut, and fat body of SBPH were collected from the 5th-instar nymphs. The tissue of the ovary was collected from the four-day-old female adults. Each sample included three independent biological replicates, the instar samples each contained thirty insects, and the tissue samples each were dissected from fifty insects.
2.5. RNA Isolation, Library Construction, and Sequencing
The total RNA of SBPH samples was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s procedure. Then, RNA quality was determined by 5300 Bioanalyser (Agilent, Santa Clara, CA, USA) and quantified using the ND-2000 (NanoDrop Technology, Wilmington, DE, USA). The samples meet the criteria with OD (260/280) = 1.8~2.2, OD (260/230) ≥ 2.0, and RQN ≥ 6.5, 28 S:18 S ≥ 1.0, >1 μg could be used to construct a sequencing library. Then, the library construction and sequencing were performed at Shanghai Majorbio Bio-pharm Biotechnology Co., Ltd., Shanghai, China, following the manufacturer’s instructions. The SBPH RNA-seq transcriptome library was prepared following Illumina® Stranded mRNA Prep, Ligation (San Diego, CA, USA) using 1 μg of total RNA. Firstly, messenger RNA was isolated using oligomeric (dT) beads by the polyA selection method and then fragmented using fragment buffer. Secondly, double-stranded cDNA was synthesized using the SuperScript double-stranded cDNA synthesis kit (Invitrogen) with random hexamer primers. Then, according to the library construction plan, the synthesized cDNA is subjected to end repair, phosphorylation, and adapter addition. The sequencing library was performed using the NovaSeq kit on the NovaSeqX Plus platform (PE150).
The raw paired-end reads were trimmed and quality controlled by fastp with default parameters [18]. Then, the clean data extracted from the sample are used for de novo assembly with Trinity [19]. To improve assembly quality, all assembly sequences filtering through CD-HIT and TransRate were evaluated using BUSCO (Benchmarking Universal Single Copy Orthologies) [20,21,22]. The assembled transcripts were searched against the NCBI protein non-redundancy (NR), Clusters of Orthologous Groups of proteins (COG), and Kyoto Encyclopedia of Genes and Genomes (KEGG) databases [23]. The annotations for uniquely assembled transcripts, used to describe biological processes, molecular functions, and cellular components were obtained through the BLAST2GO program [24].
2.6. Differential Expression Analysis and Function Enrichment
The expression levels of each transcript were calculated using the TPM (the scripts per million reads) method to identify differentially expressed genes (DEGs) between two different groups. The abundance of genes was quantified by RSEM [25]. DESeq2 was used to perform the differential expression analysis with Log2FC ≥ 1 and FDR < 0.05 [26]. Additionally, functional enrichment analysis including GO and KEGG was conducted to determine DEGs that were significantly enriched in GO terminology and metabolic pathways, at Bonferroni-corrected p-values < 0.05 compared with the whole-transcriptome background. GO functional enrichment and KEGG pathway analysis were performed using Goatools 1.4.4 and Python 1.0.0 scipy software, respectively.
2.7. Quantitative Real-Time PCR
The StarScript III All-in-one RT Mix with gDNA Remover (Genstar, Beijing, China) was utilized to synthesize the first-strand cDNA of mRNA from total RNA. The relative mRNA expression was analyzed by RT-qPCR, which was carried out by a LineGene 9600 Plus real-time PCR detection system (BIOER, Hangzhou, China) with the 2×RealStar Fast SYBR qPCR Mix (Low ROX) (Genstar, Beijing, China). Beta-tubulin was employed as the reference gene and the 2−∆∆Ct method was used to calculate the relative expression. The RT-qPCR reaction included three independent biological and technical replications. The primers are listed in Table S1.
2.8. Gene Identification and Phylogenetic Analysis
The ABC transporter gene, which attracts attention in this study, has been annotated to multidrug resistance protein 49-like protein (MDR49) and uploaded to the NCBI by other researchers (GenBank accession number: KF828784.1). The accuracy of nucleoside sequence has been demonstrated by PCR. Other ABC transporter genes from SBPH and N. lugens were obtained from our previous study [27]. A phylogenetic tree was generated by the ClustalW alignment of the amino acid sequences using the neighbor-joining method with 1000 bootstrap replications in MEGA 5.05.
2.9. RNA Interference
A T7 high-yield transcription kit (Invitrogen) was used to synthesize the double-stranded RNA (dsRNA) of enhanced green fluorescent protein (EGFP) and MDR49 according to the manufacturer’s instructions. The primers for dsRNA synthesis are listed in Table S1. The 3rd-instar nymphs of SBPH were injected with 100 nL (4 µg/µL) dsMDR49 or dsEGFP, which served as the treatment and control. The knock-down efficiency was analyzed by RT-qPCR at 1.5, 3, and 5 days after injection with dsRNA, respectively. In a parallel experiment, the 3rd-instar nymphs of SBPH injected with dsRNA 72 h later were treated with an LD50 dose compound 10 and then transferred to rice seedlings. Mortality was detected at 72 h. Each treatment contained thirty insects and was repeated three times.
2.10. Statistical Analysis
SPSS 23.0 software (IBM Corporation, Chicago, IL, USA) was used for statistical analysis of the bioassay results. The insecticide treatment mortality was corrected with control mortality using Abbott’s formula. The LD50, slope, 95% confidence interval (CI), and χ2 were calculated using probit analysis and the chi-square test. The significance of differences between treatments was analyzed by the SPSS 23.0 software. The statistical significance of gene expression and mortality between the treatment and control groups was calculated using the Student’s t-test. The asterisk (*) and asterisks (**) indicate p < 0.05 and p < 0.01. The instar expression profile and tissue expression profile were compared by the analysis of variance (ANOVA) followed by Duncan’s multiple comparisons among different treatments. Different letters indicate significant differences (p < 0.05).
3. Results
3.1. The Synthesis of Insecticidal Active Compounds
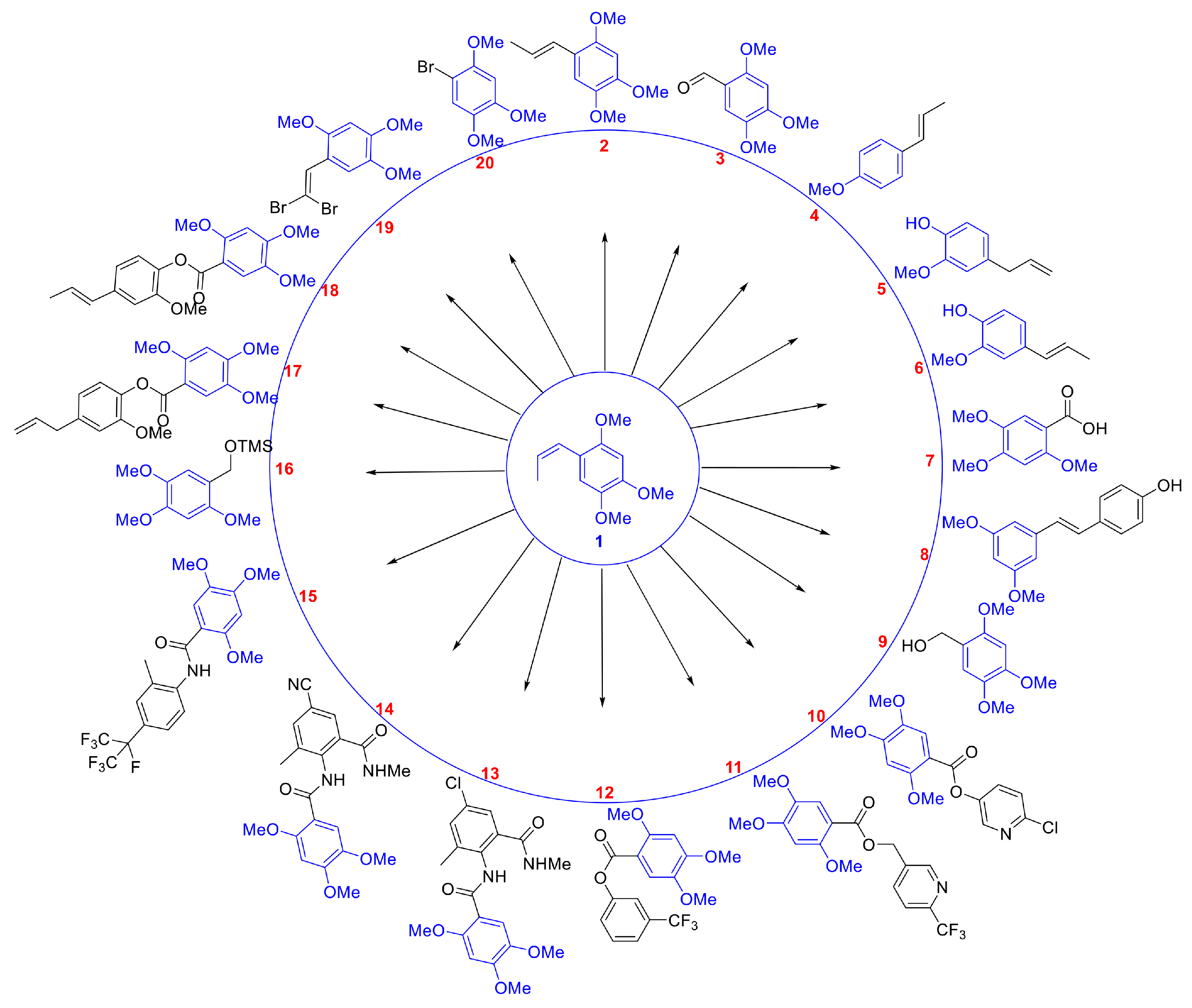
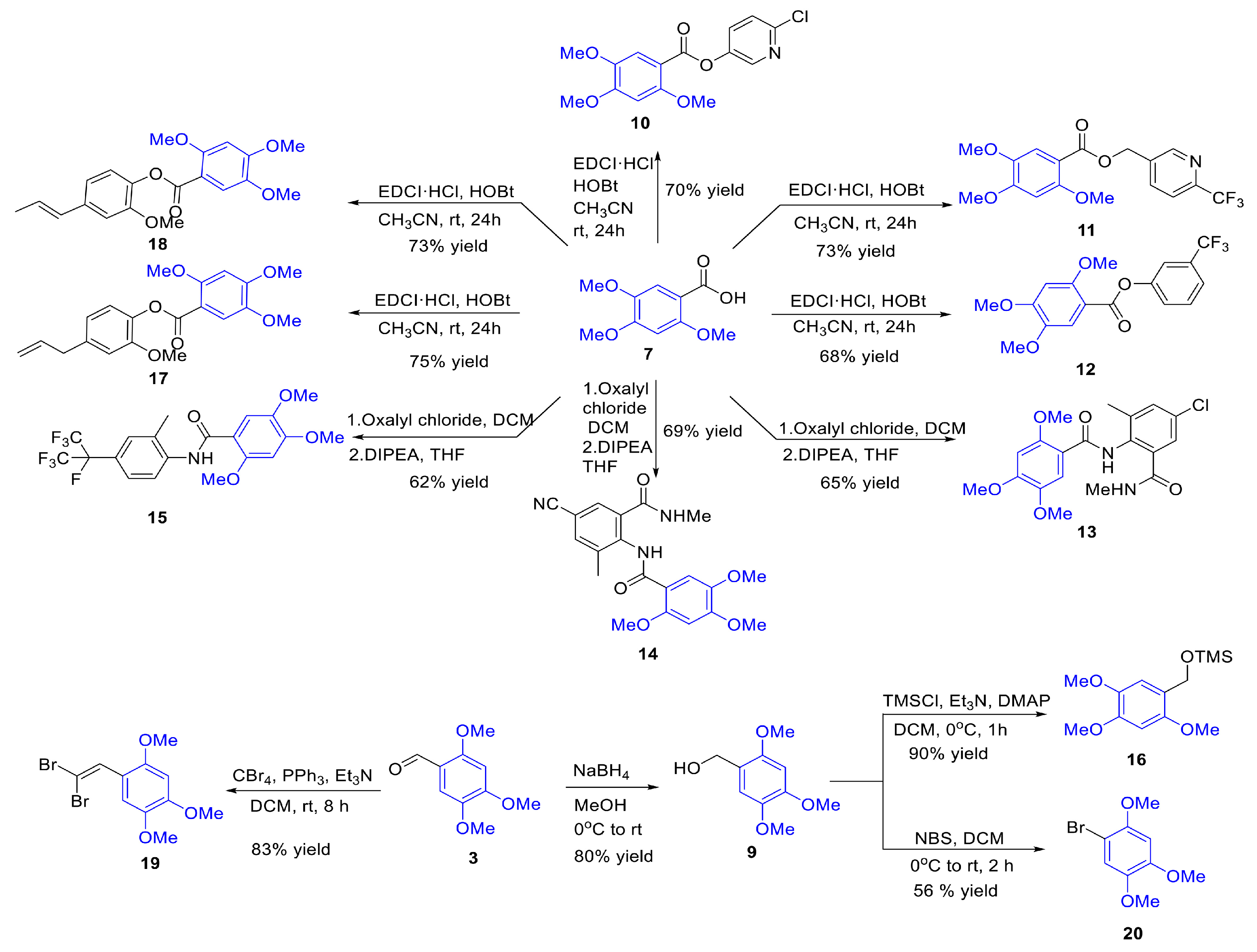
Compounds 2 to 8 are structurally similar to β-asarone, while compounds 9 to 20 are a series of derivatives based on β-asarone and its analogs. The structures of these twenty compounds are shown in Figure 1. Aryl (heteroaryl) ester or amide functional groups are key active moieties in various commercial insecticides, particularly halogens (chlorine, bromine, fluorine) and trifluoromethyl groups on aromatic or heteroaromatic rings, which significantly impact insecticidal activity. Based on the structure–activity relationship (SAR) studies and active substructure splicing strategy, compound 7 was esterified with active functional groups from highly effective chemical insecticides (cyantraniliprole, chlorantraniliprole, flubendiamide, flupyrimin, sulfoxaflor, and triflumezopyrim) to yield a series of esters and amides (compounds 10–15, 17–18). Additionally, compound 3 underwent a reduction reaction to form compound 9, which was subsequently derivatized to form compound 16 and compound 20. Compound 3 could also undergo a substitution reaction to yield compound 19 (Figure 2). Synthetic procedures and methods of compounds 9 to 20 were described in the Supporting Data. The physicochemical properties, i.e., 1H and 13C NMR of compounds 9–20 are presented in Table S2 and Supporting Data.
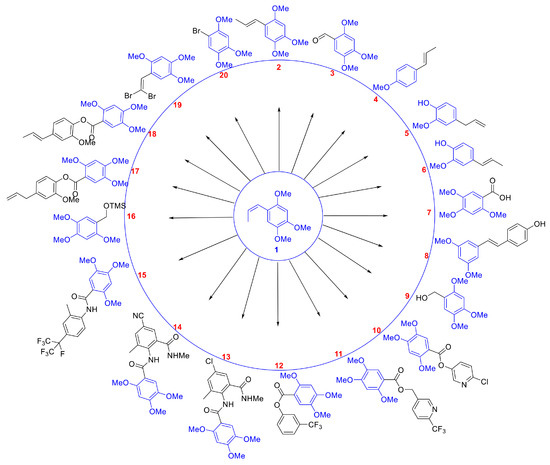
Figure 1.
Two-dimensional structures of 20 compounds derived from β-asarone.
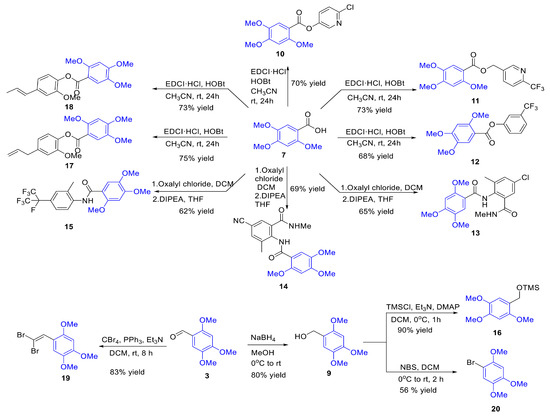
Figure 2.
The synthesis process of compounds 9–20. EDCI·HCl = N-(3-dimethylaminopropyl)-n’-ethylcarbodiimide hydrochloride; HOBt = 1-Hydroxybenzotriazole; DCM = Dichloromethane; DIPEA = N,N-Diisopropylethylamine; DMAP = 4-Dimethylaminopyridine; NBS = N-Bromosuccinimide.
3.2. Insecticide Activity of 20 Compounds
Except for compound 15, which was insoluble, the insecticidal activity of the other 19 compounds was determined by biological assay (Table 1). The insecticidal activity of all compounds was superior to that of β-asarone. Among them, compound 10, compound 8, and compound 7 exhibited the most potent insecticidal activity against SBPH, with LD50 values of 0.051, 0.073, and 0.085 μg/nymph, respectively (Table 1). Statistically, according to non-overlapping confidence intervals, there was no difference in efficacy between compound 10 and compounds 7 and 8.

Table 1.
Insecticidal activity of 20 compounds against SBPH.
3.3. Overall Analysis of Sequencing Data
Because of the best insecticidal activity, compound 10 was selected to run the follow-up research. Three control samples and three compound 10 treatment samples were sequenced, yielding an average of 44,572,031 raw reads and 44,007,361 clean reads. More than 74% of the clean reads were successfully assembled and aligned to the genome of SBPH. Data quality evaluation showed Q30 values above 89.50% and Q20 values above 96.27% in all samples, with the GC percentage counts ranging from 40.04% to 42.22% (Table S3). The clean reads were finally assembled into 59,004 unigenes with a mean length of 1094 bp and an N50 length of 2141 bp (Table S4).
3.4. Functional Annotation and Analysis of Differentially Expressed Genes (DEGs)
By Blastx homology search, 18,934, 16,003, 22,637, 25,684, 17,295, and 17,941 unigenes matched entries in the database of GO, KEGG, eggNOG, NR, Swiss-Prot, and Pfam, respectively (Figure S1). The biological function of these unigenes was further elucidated through GO and KEGG assignments (Figure S2).
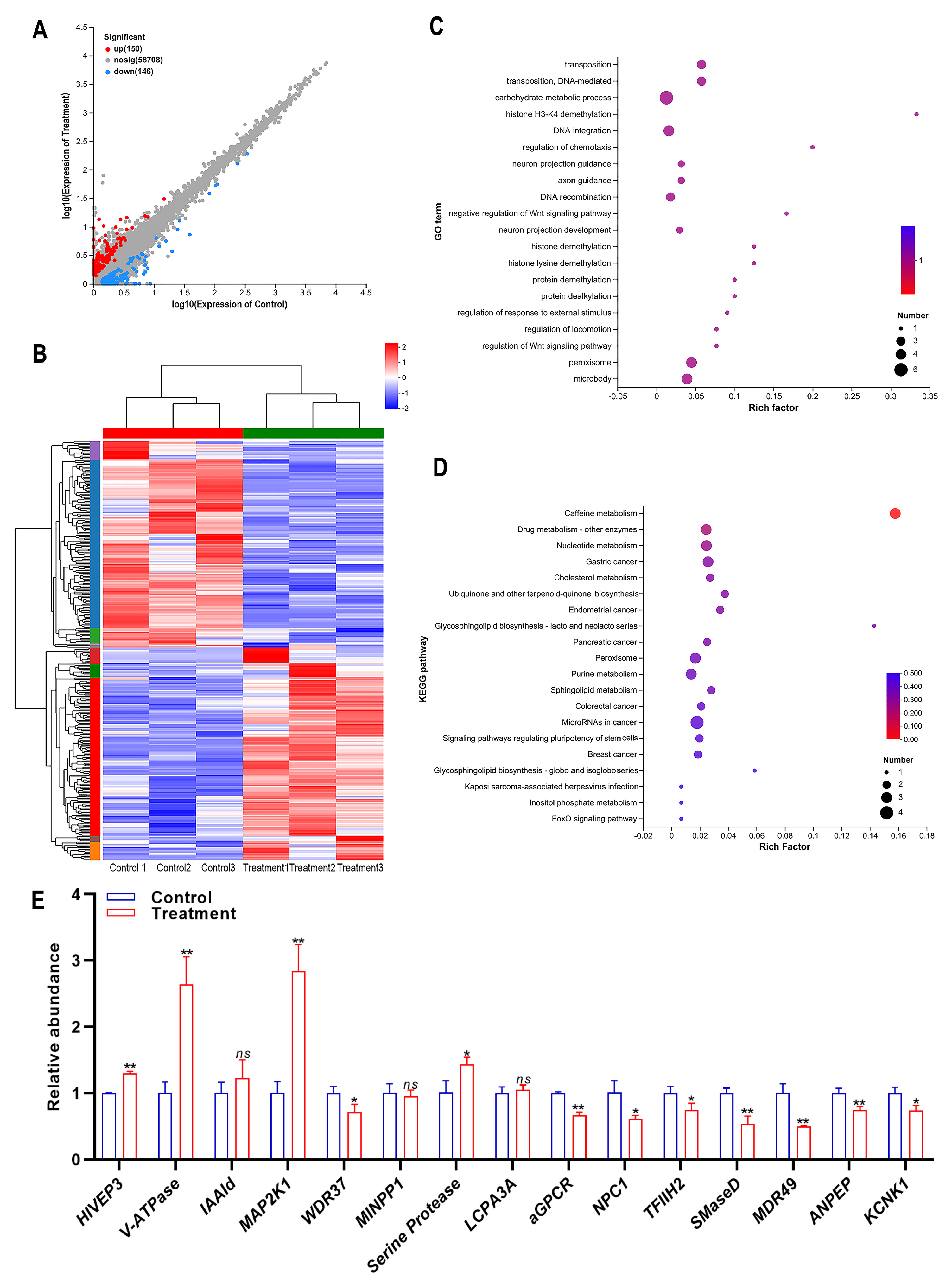
To identify the related genes that play crucial roles in the insecticidal mechanism of compound 10, the DEGs between the treatment of compound 10 and control samples were analyzed. A total of 296 genes (150 upregulated genes and 146 downregulated genes) were significantly differentially expressed in the treatment samples compared with the control (Figure 3A). A cluster heat map of DEGs between treatment and control is shown in Figure 3B. The function of DEGs was predicted through GO and KEGG enrichment analyses. The top 20 significantly enriched GO terms of the DEGs are presented in Figure 3C. The terms carbohydrate metabolic process, DNA integration, peroxisome, and microbody were the most significantly enriched terms (Figure 3C). The top 20 significantly enriched KEGG pathways of the DEGs are presented in Figure 3D. The Caffeine metabolism was the most significantly enriched pathway (Figure 3D).
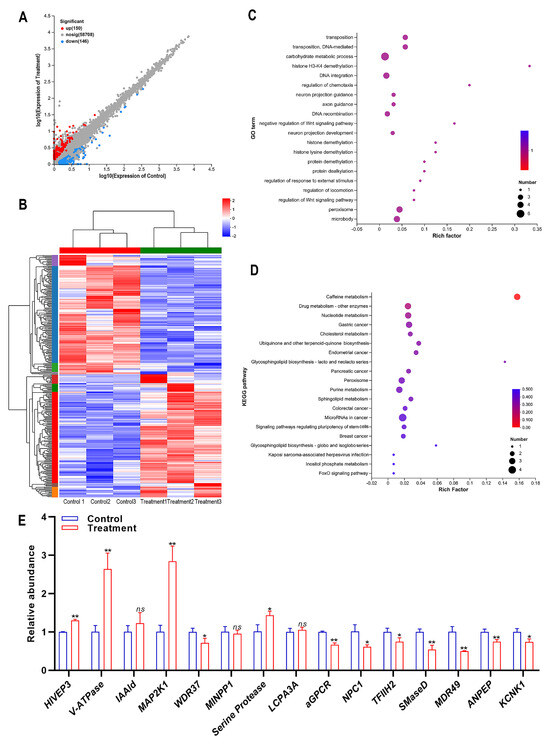
Figure 3.
Differential gene expression analysis of SBPH in response to compound 10 treatment. (A) Number of up- and down-regulated differentially expressed genes (DEGs). (B) Cluster heat map of DEGs. X-axis: sample name, Y-axis: DEGs. The colors indicated the expression levels of DEGs. (C) GO enrichment analysis of DEGs. The horizontal axis represents the enrichment degree, and the vertical axis represents the enrichment GO Term. The dot size indicates the number of differential genes enriched in a certain GO term. Dot colors indicate different p-values; the Rich factor represents the number of differential genes belonging to a GO Term/the total number of genes belonging to the GO Term. The greater the Rich factor, the higher the GO enrichment degree. (D) Enrichment analysis of the KEGG pathway for DEGs. The horizontal axis represents the Rich factor, and the vertical axis represents the enriched KEGG pathway. The dot size indicates the number of differential genes enriched in a KEGG pathway. Dot colors indicate different p-values; the Rich factor represents the number of differential genes belonging to a KEGG pathway/the total number of genes belonging to the KEGG pathway. The greater the Rich factor, the higher the enrichment degree of the KEGG pathway. (E) Quantitative verification of DEGs by RT-qPCR. The asterisk (*) indicates p < 0.05. The asterisks (**) indicate p < 0.01. The ns represents no significant difference. Results are shown as the average ± standard error (SE).
3.5. Screening of DEGs Associated with the Insecticidal Activity of Compound 10
The DEGs, which may play important roles in insecticidal activity, were selected to verify the accuracy of the transcriptome sequencing. These DEGs included transcription factor HIVEP3 (human immunodeficiency virus type I enhancer binding protein 3), V-ATPase (vesicle-fusing ATPase), IAAld (indole-3-acetaldehyde oxidase), MAP2K1 (mitogen-activated protein kinase kinase 1), WDR37 (WD repeat-containing protein 37), MINPP1 (multiple inositol polyphosphate phosphatase), serine protease, LCPA3A (larval cuticle protein A3A), aGPCR (Adhesion G protein-coupled receptor), NPC1 (Niemann-Pick C1 protein), TFIIH2 (general transcription factor IIH subunit 2-like), SMaseD (Sphingomyelinase D), MDR49, ANPEP (aminopeptidase), and KCNK1 (potassium channel subfamily K member 1). The result showed that, except IAAld, MINPP1, and LCPA3A, the quantitative validation results of DEGs were consistent with the transcriptome. Furthermore, MDR49 was the most significantly down-regulated DEG (Figure 3E).
3.6. MDR49 Affects the Insecticidal Activity of Compound 10 Against SBPH
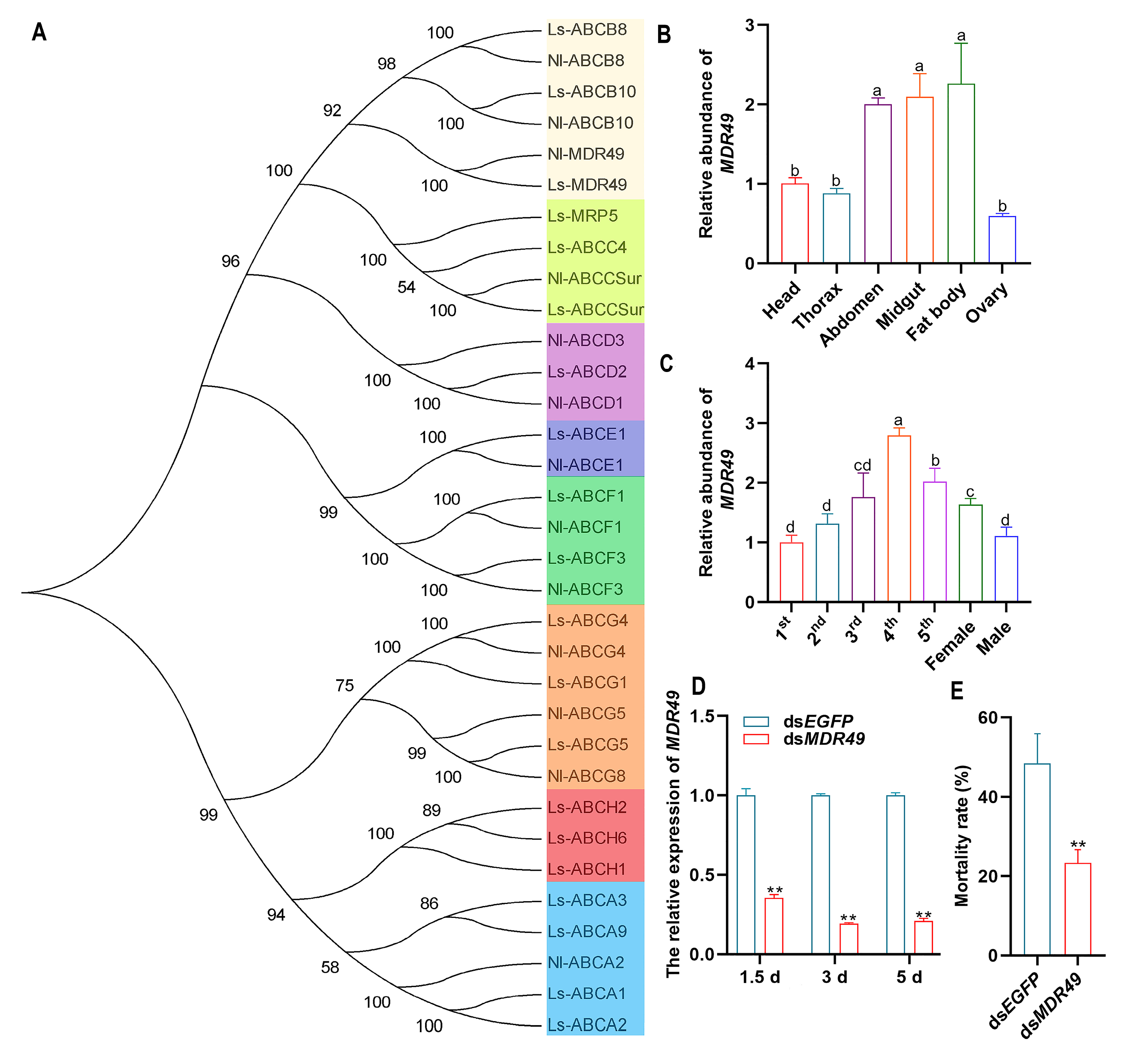
The ABC transporters are classified into eight subfamilies from ABCA to ABCH in insects. In this study, the most down-regulated gene in the transcriptome, MDR49, belongs to the ABCB subfamily (Figure 4A). To analyze the function of MDR49, the expression patterns of different developmental stages and tissues were analyzed in SBPH. The tissue expression profile revealed that MDR49 was expressed in all tissues of SBPH and was expressed at high levels in the fat body, midgut, and abdomen (Figure 4B). The instar expression profile showed that MDR49 was variously expressed at all developmental stages and was highly expressed in 4th-instar nymphs, followed by the nymphs of the 5th-instar (Figure 4C).
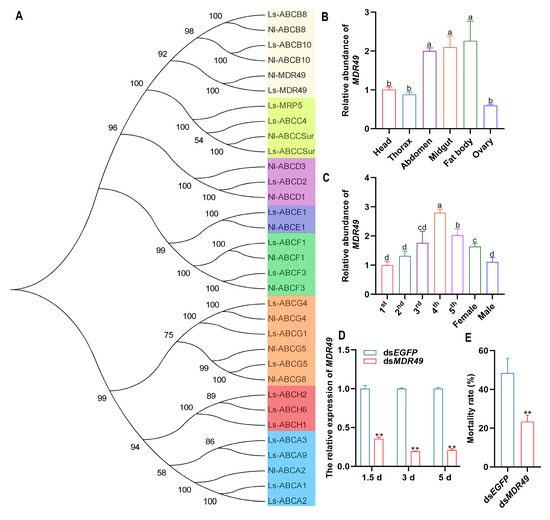
Figure 4.
Effect of MDR49 on compound 10 insecticidal activity in SBPH. (A) Phylogenetic analysis of ABC transporter genes. The numbers above the branches indicate support for the phylogenies, and only values above 50% are shown. Ls, Laodelphax striatellus; Nl, Nilaparvata lugens. (B) Expression profile of MDR49 in different tissues in SBPH. (C) Expression profile of MDR49 at different developmental stages in SBPH. (D) The relative expression level of MDR49 after silencing. (E) Mortality rate of SBPH in response to compound 10 after silencing MDR49. The letters a, b, c, and d present significant differences (p < 0.05) in the expression. The asterisks (**) indicate p < 0.01. Results are shown as the average ± standard error (SE).
To confirm whether MDR49 plays a role in the insecticidal effect of compound 10, RNAi was conducted to knock down MDR49 in SBPH. The relative expression of MDR49 was decreased significantly at 1.5, 3, and 5 days after silencing when compared to dsEGFP (Figure 4D). The bioassay results showed that down-regulating the expression of MDR49 could significantly reduce the SBPH mortality after LD50 treatment of compound 10 (Figure 4E).
3.7. The Insecticidal Spectrum of Compound 10
Compound 10 also demonstrated excellent insecticidal activity against the other five agricultural pests. Compared to SBPH, compound 10 had similar toxicity against A. craccivora, R. maidis, and S. heraclei, whereas the toxicities against B. tabaci and T. cinnabarinus were 8.5 and 5.1 times higher, respectively (Table 2).
Table 2.
Insecticidal activity of compound 10 against other insects.
4. Discussion
The SBPH is a serious pest in the grass family and is at risk of developing resistance to most kinds of insecticides [28,29]. Therefore, there is an urgent need for strategies to reduce or replace the use of chemical insecticides to address the severe problems caused by their widespread application. One such approach is the development of botanical insecticides.
Botanical pesticides are preparations made from all or part of the organic substances of plant organisms and secondary metabolites, including the active ingredients extracted from plants and derivatives synthesized according to the active structures [30,31,32]. Due to the characteristics of safety, nonpollution, and environment-friendly, more and more attention has been paid to botanical pesticides. As we all know some active substances, such as azadirachtin, pyrethrins, matrine, osthole, and eucalyptol, are derived from Azadirachta indica, Tanacetum cinerariifoliums, Sophora flavescens, Cnidium monnieri, and Cinnamomum longepaniculatum, have been developed into various commercial insecticides or fungicides [33,34,35,36,37]. However, the amount of active substances in plants is very low, so relying on the active substances in the plants during the processing will result in high costs [38,39]. Therefore, it is of great significance to isolate these compounds, clarify their chemical structures, and simulate compounds with similar compositions for the development of agricultural pesticides. Active substructure splicing strategy has been widely used in developing higher effective and lower toxic insecticides, bactericides, fungicides, and algicides [40,41,42,43]. Several osthole-derived N-benzoylthioureas introduced the active thiourea moieties of thiourea pesticides exhibited broad-spectrum insecticidal activities and less toxicity to non-target organisms [40]. Isoxazoline, which contains fragments of good biological, was widely used in the development of new pesticides [44,45]. Three osthole derivatives, which introduced the isoxazoline moiety, improve its insecticidal activity to Mythimna separata and P. xylostella [40]. Compounds with steroid skeletons, such as Periplocoside X (PSX) and periplocoside A (PSA) isolated from the root bark of Periploca sepium Bunge, displayed insecticidal activity against various pests [46,47]. Steroidal oxazole derivatives were synthesized by introducing 1,3,4-oxadiazole heterocycles, which have diverse pharmacological properties, on snake tail glycoside X and snake tail glycoside A, exhibiting the similar effects as commercial insecticides, such as chlorpyrifos and thiamethoxam to Eriosoma lanigerum [48].
In the previous study, β-asarone, which is derived from A. calamus, has been identified as an insecticide with activity against Coleoptera, Lepidoptera, and Hemiptera pests [12,13,49,50]. However, there is a significant gap in insecticidal efficacy between β-asarone and conventional chemical insecticides. Therefore, improving the insecticidal effectiveness of β-asarone is essential for its broader application. So far, the commercial chemical insecticides, cyantraniliprole, chlorantraniliprole, flubendiamide, flupyrimin, sulfoxaflor, and triflumezopyrim are highly active broad-spectrum insecticides [51,52,53,54,55,56]. Based on this, the functional groups from these high-efficiency chemical insecticides were added to β-asarone through a series of chemical reactions to obtain multiple compounds. Fortunately, the insecticidal activity of almost all the derivatives was higher than that of β-asarone, with compound 10 showing an 8.31-fold increase in efficacy. Flupyrimin is a pesticide that acts on nicotinic acetylcholine receptor (nAChR), discovered by Meiji Seika Pharma, aand has unique biological properties, including outstanding potency to imidacloprid (IMI)-resistant rice pests together with superior safety toward pollinators [57]. The 2-chloropyridine functional group present in flupyrimin is also found in other insecticides such as imidacloprid, acetamiprid, nitenpyram, thiacloprid, fluopyrifuranone, cycloxaprid, paichongding, and guadipyr. Therefore, we introduced the 2-chloropyridine group into compound 7, a β-asarone analog, to obtain compound 10, which demonstrated excellent insecticidal activity against the small brown planthopper (SBPH). Compound 10 also demonstrated good insecticidal activities against S. heraclei, A. craccivora, B. tabaci, R. maidis, and T. cinnabarinus. However, compound 10 is one of the derivatives derived from the active functional groups of highly effective insecticides, and its possible ecotoxicological effects are not provided with sufficient information in this study. Studies have shown that most of the target osthole-derived N-benzoylthioureas displayed a more potent and broad-spectrum insecticidal effect than the parent osthole after the introduction of N-benzoylthioureas on the C-3′ position of osthole. Among them, the potent compounds B19 and B23 possessed good selectivity and were less toxic to non-target organisms [40]. Additionally, approximately 70% of antimicrobials are derived from natural products and their derivatives, such as penicillin G, amoxicillin, oxytetracycline, and minocycline. A series of novel honokiol/magnolol amphiphiles were prepared by mimicking the chemical structures and antibacterial properties of cationic antimicrobial peptides, and some showed excellent antibacterial activity against Gram-positive bacteria and clinical MRSA isolates [58]. Therefore, compound 10 may not have only excellent insecticidal activity but may also have a better ecotoxicological profile, which needs further research.
Clarifying the action mode of insecticide against pests could provide a sustainable strategy for pest management. Targets served as potential action sites for insecticides, and their changes in transcription levels have been shown to mediate insecticide sensitivity. In this study, bioinformatics and molecular biology methods were used to identify the related genes, which play important roles in the action mode of compound 10 against SBPH. Fifteen genes, such as MAP2K1, aGPCR, MDR49, NPC1, and KCNK1, were selected from the transcriptome, all of which have been reported to play an important role in the mechanism of pesticide action [59,60,61,62,63]. The technology of RNAi has great potential for application in the detection of insecticide target genes [64,65]. The study in Aphis gossypii confirmed that the action mode of 6-methylcoumarin against aphids was by inhibiting AChE [66]. RNAi suppression of the ryanodine receptor gene results in decreased susceptibility to chlorantraniliprole in Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae) [67]. In this study, decreasing the expression of an ABC transporter gene, MDR49, through RNAi reduced the sensitivity to compound 10 in SBPH. It is generally known that the ABC transporters play significant parts in the detoxification of insecticides in pests. Over-expression of MDR49 confers resistance to imidacloprid in N. lugens [68]. In Drosophila melanogaster, sensitivity to nine different insecticides was increased by silencing MDR65 [69]. Moreover, many reports have shown that the reduced expression of ABC transporters could also mediate resistance. A study on P. xylostella highlights that ABCC2 and ABCC3, which reduce expression by the MAPK signaling pathway, play a crucial role in field-evolved resistance to Cry1Ac [70]. In Spodoptera frugiperda, a SfABCC2 knockout strain using the CRISPR/Cas9 system displayed high resistance to Cry1F compared with the parental strain [71].
5. Conclusions
In summary, several β-asarone derivatives that showed better insecticidal activity to SBPH were obtained through a series of modifications. MDR49 was found to be down-regulated after compound 10 treatment and the importance of MDR49 in insecticidal activity of compound 10 was verified by RNAi. Additionally, compound 10 exhibited exceptional insecticidal activity against B. tabaci and T. cinnabarinus. Our results provide a scientific basis for developing compound 10 as a plant-derived pesticide for effective pest control. Further research is needed on its environmental properties such as toxicity to mammals.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14102420/s1, Figure S1: The number of unigenes annotated in the public database searched; Figure S2: Functional annotation of unigenes. (A) Go Function classification of all annotated unigenes from the transcriptome. (B) KEGG pathway classification of all annotated unigenes from the transcriptome; Table S1: Primers used in this study; Table S2: Information on 20 compounds; Table S3: Summary of the transcriptome sequencing data from the control and compound 10 treated samples; Table S4: Evaluation of assembly sequences; Supporting Data: The synthetic procedure of compound.
Author Contributions
Conceptualization, J.Z., Y.Y. and Y.Z. (Yun Zhou); Methodology, A.W., Y.Z. (Yun Zhou), and Y.Z. (Yifei Zhang); Software, X.F., X.J. and Y.Z. (Yanwei Zhu); Validation, A.W., Y.Z. (Yun Zhou), X.F., X.J., Y.Z. (Yanwei Zhu), Y.Z. (Yun Zhang), and C.X.; Formal analysis, X.F., Y.C., X.J. and Y.Z. (Yanwei Zhu); Investigation, A.W., Y.Z. (Yun Zhou), X.F., Y.C., Y.Z. (Yun Zhang), and C.X.; Resources, X.W.; Data curation, A.W., Y.Z. (Yun Zhou), X.F., X.W., Y.C., Y.Z. (Yifei Zhang), Y.Z. (Yun Zhang), and C.X.; Writing—original draft preparation, A.W., Y.Z. (Yun Zhou), J.Z. and Y.Y.; Writing—review and editing, C.S., M.Z., J.Z. and Y.Y.; Visualization, C.S. and M.Z.; Supervision, C.S. and M.Z.; Project administration, C.S., M.Z., J.Z. and Y.Y.; Funding acquisition, Y.Z. (Yifei Zhang), C.S., M.Z., J.Z. and Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Technology Innovation Guidance Plan of Shandong Province, China (2022LYXZ026); Central Guidance of Local Science and Technology Development Special Foundation (YDZX2022077); National Natural Science Foundation of China (32102211, 32302355); GDAS’ Project of Science and Technology Development (2018GDASCX-0926); Natural Science Foundation of Shandong Province (ZR2023QB024); Agricultural Scientific and Technological Innovation Project of Shandong Academy of Agricultural Sciences (CXGC2024F03, CXGC2024D03); and Key Research and Development Plan of Shandong Province (2022TZXD0034).
Data Availability Statement
The transcriptome data were deposited in the Gene Expression Omnibus (GEO) at the National Center for Biotechnology Information (NCBI) under accession number GSE272249.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Elzaki, M.E.A.; Zhang, W.; Feng, A.; Qiou, X.; Zhao, W.; Han, Z. Constitutive overexpression of cytochrome P450 associated with imidacloprid resistance in Laodelphax striatellus (Fallen). Pest Manag. Sci. 2016, 72, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, A.; Xue, C.; Tian, H.; Zhang, Y.; Zhou, M.; Zhao, M.; Liu, Z.; Zhang, J. MicroRNA PC-5p-3991_515 mediates triflumezopyrim susceptibility in the small brown planthopper through regulating the post-transcriptional expression of P450 CYP417A2. Pest Manag. Sci. 2024, 80, 1761–1770. [Google Scholar] [CrossRef]
- Tanno, F.; Nakatsu, A.; Toriyama, S.; Kojima, M. Complete nucleotide sequence of Northern cereal mosaic virus and its genome organization. Arch. Virol. 2000, 145, 1373–1384. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, L.; Ren, Y.D.; Wang, X.F. Rice black-streaked dwarf virus: From multiparty interactions among plant–virus–vector to intermittent epidemics. Mol. Plant Pathol. 2020, 21, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Hajano, J.; Raza, A.; Zhang, L.; Liu, W.; Wang, X. Ribavirin targets sugar transporter 6 to suppress acquisition and transmission of rice stripe tenuivirus by its vector Laodelphax striatellus. Pest Manag. Sci. 2020, 76, 4086–4092. [Google Scholar] [CrossRef]
- Dively, G.P.; Kamel, A. Insecticide residues in pollen and nectar of a cucurbit crop and their potential exposure to pollinators. J. Agric. Food Chem. 2012, 60, 4449–4456. [Google Scholar] [CrossRef]
- Wu, J.; Ge, L.; Liu, F.; Song, Q.; Stanley, D. Pesticide-induced planthopper population resurgence in rice cropping systems. Annu. Rev. Entomol. 2020, 65, 409–429. [Google Scholar] [CrossRef]
- Imai, T.; Masuda, R. Insecticidal activities of methyleugenol and β-asarone, from the herbal medicines Saishin and Sekishōkon, and other alkoxy-propenyl-benzene derivatives against the cigarette beetle Lasioderma serricorne (Coleoptera: Anobiidae). Appl. Entomol. Zool. 2017, 52, 183–188. [Google Scholar] [CrossRef]
- Arasu, M.V.; Viayaraghavan, P.; Ilavenil, S.; Al-Dhabi, N.A.; Choi, K.C. Essential oil of four medicinal plants and protective properties in plum fruits against the spoilage bacteria and fungi. Ind. Crops Prod. 2019, 133, 54–62. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, W.; Yang, C.; Hua, H. Supercritical fluid CO2 extraction of Acorus calamus L. (Arales: Araceae) and its contact toxicity to Sitophilus zeamais Motschusky (Coleoptera: Curculionidae). Nat. Prod. Res. 2012, 26, 1498–1503. [Google Scholar] [CrossRef]
- Reddy, S.G.E.; Kirti Dolma, S.; Koundal, R.; Singh, B. Chemical composition and insecticidal activities of essential oils against diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae). Nat. Prod. Res. 2016, 30, 1834–1838. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Sharma, S.; Sharma, S.; Kumari, A.; Kumar, D.; Nadda, G.; Padwad, Y.; Ogra, R.K.; Kumar, N. Chemical composition, cytotoxicity and insecticidal activities of Acorus calamus accessions from the western Himalayas. Ind. Crops Prod. 2016, 94, 520–527. [Google Scholar] [CrossRef]
- Xu, X.; Li, X.; Liu, Z.; Wang, F.; Fan, L.; Wu, C.; Yao, Y. Knockdown of CYP301B1 and CYP6AX1v2 increases the susceptibility of the brown planthopper to beta-asarone, a potential plant-derived insecticide. Int. J. Biol. Macromol. 2021, 171, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Fang, Y.; Che, W.; Zhang, Q.; Wang, J.; Luo, C. The toxicity, sublethal effects, and biochemical mechanism of β-asarone, a potential plant-derived insecticide, against Bemisia tabaci. Int. J. Mol. Sci. 2022, 23, 10462. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, X.; Wang, F.; Han, K.; Liu, Z.; Fan, L.; Hua, H.; Cai, W.; Yao, Y. Candidate detoxification-related genes in brown planthopper, Nilaparvata lugens, in response to β-asarone based on transcriptomic analysis. Ecotoxicol. Environ. Saf. 2019, 185, 109735. [Google Scholar] [CrossRef]
- Wen, S.; Xue, Y.; Du, R.; Liu, C.; Wang, X.; Wang, Y.; Liu, C.; Wang, S.; Wang, J.; Xia, X. Toxicity and sublethal effects of triflumezopyrim on the development and detoxification enzymatic activities in the small brown planthopper (SBPH), Laodelphax striatellus (Fallen). Crop Prot. 2021, 150, 105813. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, A.; Zhang, Y.; Xue, C.; Zhao, M.; Zhang, J. Activating pathway of three metabolic detoxification phases via down-regulated endogenous microRNAs, modulates triflumezopyrim tolerance in the small brown planthopper, Laodelphax striatellus (Fallén). Int. J. Biol. Macromol. 2022, 222, 2439–2451. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Smith-Unna, R.; Boursnell, C.; Patro, R.; Hibberd, J.M.; Kelly, S. TransRate: Reference-free quality assessment of de novo transcriptome assemblies. Genome Res. 2016, 26, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, A.; Wang, M.; Zhang, Y.; Zhang, J.; Zhao, M. ATP-binding cassette transporters ABCF2 and ABCG9 regulate rice black-streaked dwarf virus infection in its insect vector, Laodelphax striatellus (Fallén). Bull. Entomol. Res. 2022, 112, 327–334. [Google Scholar] [CrossRef]
- Gao, B.; Wu, J.; Huang, S.; Mu, L.; Han, Z. Insecticide resistance in field populations of Laodelphax striatellus (Fallén) (Homoptera: Delphacidae) in China and its possible mechanisms. Int. J. Pest Manag. 2008, 54, 13–19. [Google Scholar] [CrossRef]
- Xue, Y.; Liu, C.; Liu, D.; Ding, W.; Li, Z.; Cao, J.; Xia, X. Sensitivity differences and biochemical characteristics of Laodelphax striatellus (Fallén) to seven insecticides in different areas of Shandong. China Insects 2022, 13, 780. [Google Scholar] [CrossRef]
- Cantrell, C.L.; Dayan, F.E.; Duke, S.O. Natural products as source for new pesticides. J. Nat. Prod. 2012, 75, 1231–1242. [Google Scholar] [CrossRef]
- Kiyota, H. Synthetic studies of biologically active natural products contributing to pesticide development. J. Pestic. Sci. 2020, 45, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; The International Natural Product Sciences Taskforce; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Lorsbach, B.A.; Sparks, T.C.; Cicchillo, R.M.; Garizi, N.V.; Hahn, D.R.; Meyer, K.G. Natural products: A strategic lead generation approach in crop protection discovery. Pest Manag. Sci. 2019, 75, 2301–2309. [Google Scholar] [CrossRef] [PubMed]
- Marrone, P.G. Pesticidal natural products-status and future potential. Pest Manag. Sci. 2019, 75, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Wessels, F.J.; Lorsbach, B.A.; Nugent, B.M.; Watson, G.B. The new age of insecticide discovery-the crop protection industry and the impact of natural products. Pestic. Biochem. Physiol. 2019, 161, 12–22. [Google Scholar] [CrossRef]
- Yang, Y.; Zhu, Q.; Zhang, K.; Zhao, S. Synthesis, antimosquito activities, photodegradation, and toxic assessment of novel pyrethroids containing 2-chlorobiphenyl and 2-chlorophenylpyridine. Pest Manag. Sci. 2021, 77, 2773–2784. [Google Scholar] [CrossRef]
- Singh, S.; Mukherjee, A.; Jaiswal, D.K.; de Araujo Pereira, A.P.; Prasad, R.; Sharma, M.; Kuhad, R.C.; Shukla, A.C.; Verma, J.P. Advances and future prospects of pyrethroids: Toxicity and microbial degradation. Sci. Total Environ. 2022, 829, 154561. [Google Scholar] [CrossRef]
- Wiratno, S.; Trisawa, I.M. Research progress, formulation, and utilization of botanical pesticide. J. Litbang Pert. 2013, 32, 150–155. [Google Scholar]
- Wang, Y.; Shi, Z. Strain pesticide research and development progress of quasi deinsectization. Chem. Reag. 2024, 46, 50–58. [Google Scholar]
- Yang, R.; Liu, Z.; Han, M.; Cui, L.; Guo, Y. Preparation and biological evaluation of novel osthole-derived N-benzoylthioureas as insecticide candidates. J. Agric. Food Chem. 2022, 70, 15737–15746. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Z.; Xu, H.; Liu, Y.; Yang, X.; Sun, T.; Lu, X.; Shi, F.; Yang, Q.; Chen, W.; et al. Synthesis, antifungal activity, and 3D-QASR of novel 1,2,3,4-tetrahydroquinoline derivatives containing a pyrimidine ether scaffold as chitin synthase inhibitors. J. Agric. Food Chem. 2022, 70, 9262–9275. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, S.; Li, T.; Ma, S.; Wang, P.; Wang, G.; Su, S.; Ding, Y.; Yang, L.; Zhou, X.; et al. Design, synthesis and bioactivity evaluation of novel 2-(pyrazol-4-yl)-1,3,4-oxadiazoles containing an imidazole fragment as antibacterial agents. Molecules 2023, 28, 2442. [Google Scholar] [CrossRef]
- Huang, S.; Zuo, L.; Zhang, L.; Guo, X.; Cheng, C.; He, Y.; Cheng, G.; Yu, J.; Liu, Y.; Chen, R.; et al. Design, synthesis, and mode of action of thioacetamide derivatives as the algicide candidate based on active substructure splicing strategy. J. Agric. Food Chem. 2024, 72, 7021–7032. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Han, M.; Yan, X.; Cheng, W.; Tang, Z.; Cui, L.; Yang, R.; Guo, Y. Design, synthesis, and biological evaluation of novel osthole-based isoxazoline derivatives as insecticide candidates. J. Agric. Food Chem. 2022, 70, 7921–7928. [Google Scholar] [CrossRef]
- Shan, X.; Lv, M.; Wang, J.; Qin, Y.; Xu, H. Acaricidal and insecticidal efficacy of new esters derivatives of a natural coumarin osthole. Ind. Crops Prod. 2022, 182, 114855. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Shi, B.J.; Hu, Z.N. The insecticidal activity of periplocoside NW. Chin. Bull. Entomol. 2008, 45, 950–952. [Google Scholar]
- Li, Y.; Zeng, X.N.; Wang, W.Z.; Luo, C.H.; Yan, Q.; Tian, M. Chemical constituents from the roots of Periploca sepium with insecticidal activity. J. Asian Nat. Prod. Res. 2012, 14, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Jiang, W.; Li, Q.; Li, T.; Wu, W.; Bai, H.; Shi, B. Design, synthesis, and study of the insecticidal activity of novel steroidal 1,3,4-oxadiazoles. J. Agric. Food Chem. 2021, 69, 11572–11581. [Google Scholar] [CrossRef]
- Yooboon, T.; Kuramitsu, K.; Bullangpoti, V.; Kainoh, Y.; Furukawa, S. Cytotoxic effects of β-asarone on Sf9 insect cells. Arch. Insect Biochem. Physiol. 2019, 102, e21596. [Google Scholar] [CrossRef]
- Chen, H.P.; Yang, K.; Zheng, L.S.; You, C.X.; Wang, C.F. Repellant and insecticidal activities of shyobunone and isoshyobunone derived from the essential oil of Acorus calamus rhizomes. Pharmacogn. Mag. 2015, 11, 675–681. [Google Scholar]
- Liao, X.; Mao, K.; Ali, E.; Jin, R.; Li, Z.; Li, W.; Li, J.; Wan, H. Inheritance and fitness costs of sulfoxaflor resistance in Nilaparvata lugens (Stal). Pest Manag. Sci. 2019, 75, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jin, M.; Wang, N.; Yu, Q.; Xue, C. Combined transcriptomic and proteomic analysis of flubendiamide resistance Plutella xylostella. Entomol. Res. 2020, 10, 50. [Google Scholar] [CrossRef]
- Umetsu, N.; Shirai, Y. Development of novel pesticides in the 21st century. J. Pestic. Sci. 2020, 45, 54–74. [Google Scholar] [CrossRef]
- Zhang, S.; Gu, F.; Du, Y.; Li, X.; Gong, C.; Pu, J.; Liu, X.; Wang, X. Risk assessment and resistance inheritance of triflumezopyrim resistance in Laodelphax striatellus. Pest Manag. Sci. 2022, 78, 2851–2859. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, X.; Guo, L.; Yin, T.; Liu, D.; Liu, S.; You, X.; Xia, X. Risk of resistance and the metabolic resistance mechanism of Laodelphax striatellus (Fallén) to cyantraniliprole. Pestic. Biochem. Physiol. 2023, 197, 105685. [Google Scholar] [CrossRef]
- Ren, H.; Zhang, H.; Tan, Y.; Ni, R.; Shan, Y.; Li, F.; Dai, G.; Li, L.; Li, Y.; Pang, B. Sublethal effects of chlorantraniliprole on biological characteristics, detoxifying enzyme activity and gene expression profile in the Allium mongolicum Regel leaf beetle Galeruca daurica (Coleoptera: Chrysomelidae). J. Appl. Entomol. 2024, 3, 148. [Google Scholar] [CrossRef]
- Onozaki, Y.; Horikoshi, R.; Ohno, I.; Kitsuda, S.; Durkin, K.A.; Suzuki, T.; Asahara, C.; Hiroki, N.; Komabashiri, R.; Shimizu, R.; et al. Flupyrimin: A novel insecticide acting at the nicotinic acetylcholine receptors. J. Agric. Food Chem. 2017, 65, 7865–7873. [Google Scholar] [CrossRef]
- Guo, Y.; Hou, E.; Wen, T.; Yan, X.; Han, M.; Bai, L.P.; Fu, X.; Liu, J.; Qin, S. Development of membrane-active honokiol/magnolol amphiphiles as potent antibacterial agents against methicillin-resistant Staphylococcus aureus (MRSA). J. Med. Chem. 2021, 64, 12903–12916. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Pu, J.; Chen, F.; Wang, J.; Han, Z. Multiple ATP-binding cassette transporters are involved in insecticide resistance in the small brown planthopper, Laodelphax striatellus. Insect Mol. Biol. 2017, 26, 343–355. [Google Scholar] [CrossRef]
- Li, Z.; Davis, J.A.; Swale, D.R. Chemical inhibition of Kir channels reduces salivary secretions and phloem feeding of the cotton aphid, Aphis gossypii (Glover). Pest Manag. Sci. 2019, 75, 2725–2734. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Y.Q.; Lai, T.; Liu, X.J.; Guo, F.Y.; Guo, T.; Ding, W. Acaricidal mechanism of scopoletin against Tetranychus cinnabarinus. Front. Physiol. 2019, 10, 164. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.C.; Yue, X.R.; Kuang, W.Q.; Li, S.L.; Tang, R.; Zhang, Z.F.; Kurban, A.; Saif-Ur-Rehman; Zhao, C.; Liu, T.X.; et al. NPC1b as a novel target in controlling the cotton bollworm, Helicoverpa armigera. Pest Manag. Sci. 2020, 76, 2233–2242. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Cheng, Z.; Qin, J.; Sun, D.; Wang, S.; Wu, Q.; Crickmore, N.; Zhou, X.; Bravo, A.; Soberón, M.; et al. MAPK-mediated transcription factor GATAd contributes to Cry1Ac resistance in diamondback moth by reducing PxmALP expression. PLoS Genet. 2022, 18, e1010037. [Google Scholar] [CrossRef]
- Ma, K.; Li, F.; Liang, P.; Chen, X.; Liu, Y.; Tang, Q.; Gao, X. RNA interference of Dicer-1 and Argonaute-1 increasing the sensitivity of Aphis gossypii Glover (Hemiptera: Aphididae) to plant allelochemical. Pestic. Biochem. Physiol. 2017, 138, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Guo, F.; Luo, J.; Zhang, Y.; Liu, J.; Zhang, Y.; Zheng, X.; Wan, F.; Ding, W. Functional analysis of an upregulated calmodulin gene related to the acaricidal activity of curcumin against Tetranychus cinnabarinus (Boisduval). Pest Manag. Sci. 2021, 77, 719–730. [Google Scholar] [CrossRef]
- Zhou, H.; Jian, Y.; Shao, Q.; Guo, F.; Zhang, M.; Wan, F.; Yang, L.; Liu, Y.; Yang, L.; Li, Y.; et al. Development of sustainable insecticide candidates for protecting pollinators: Insight into the bioactivities, selective mechanism of action and QSAR of natural coumarin derivatives against aphids. J. Agric. Food Chem. 2023, 71, 18359–18374. [Google Scholar] [CrossRef]
- Wan, P.J.; Guo, W.Y.; Yang, Y.; Lü, F.G.; Lu, W.P.; Li, G.Q. RNAi suppression of the ryanodine receptor gene results in decreased susceptibility to chlorantraniliprole in Colorado potato beetle Leptinotarsa decemlineata. J. Insect Physiol. 2014, 63, 48–55. [Google Scholar] [CrossRef]
- Wang, L.X.; Tao, S.; Zhang, Y.C.; Pei, X.G.; Gao, Y.; Song, X.Y.; Yu, Z.T.; Gao, C.F. Overexpression of ATP-binding cassette transporter Mdr49-like confers resistance to imidacloprid in the field populations of brown planthopper, Nilaparvata lugens. Pest Manag. Sci. 2022, 78, 579–590. [Google Scholar] [CrossRef]
- Sun, H.; Buchon, N.; Scott, J.G. Mdr65 decreases toxicity of multiple insecticides in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2017, 89, 11–16. [Google Scholar] [CrossRef]
- Guo, Z.; Kang, S.; Chen, D.; Wu, Q.; Wang, S.; Xie, W.; Zhu, X.; Baxter, S.W.; Zhou, X.; Jurat-Fuentes, J.L.; et al. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in diamondback moth. PLoS Genet. 2015, 11, e1005124. [Google Scholar] [CrossRef]
- Jin, M.H.; Tao, J.H.; Qi, L.I.; Cheng, Y.; Sun, X.X.; Wu, K.M.; Xiao, Y.T. Genome editing of the SfABCC2 gene confers resistance to Cry1F toxin from Bacillus thuringiensis in Spodoptera frugiperda. J. Integr. Agric. 2021, 20, 815–820. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).