Abstract
Iodine is a critical trace element necessary for human and animal health owing to its role in thyroid hormone synthesis. Despite its importance, iodine deficiency remains a global health concern. Traditional methods to address this issue, such as salt iodization, face challenges like iodine loss during storage and cooking. The biofortification of plants, particularly carrots, offers a promising alternative. This study investigates iodine accumulation and distribution in carrots biofortified with potassium iodide (KI) and potassium iodate (KIO3) using single extraction at elevated temperatures with tetramethylammonium hydroxide followed by iodine determination by ICP-MS. Results show that iodine biofortification significantly increases the iodine content of various parts of the carrot, especially the leaves and the root peels. Carrots treated with iodate accumulate 2.7 times more iodine than those treated with iodide. The leaves exhibit the highest iodine content, particularly in iodate-treated plants, where levels can be over 24 times higher than those of control carrots. Root peel and roots also accumulate substantial amounts of iodine, with the iodate treatment resulting in 5.42 mg·kg−1 and 3.75 mg·kg−1 dry weight, respectively. The potential application of biofortified carrots can help meet human dietary iodine requirements; additionally, iodine-rich carrot leaves as livestock feed offer a sustainable approach to increasing the iodine intake in animals.
1. Introduction
Iodine is a vital trace element essential for human and animal health. It plays a crucial role in the synthesis of thyroid hormones, specifically T4 (tetraiodo-L-thyronine) and T3 (triiodo-L-thyronine) [1]. These hormones regulate the metabolism, growth, and development [2], as well as controlling the basal metabolic rate, protein synthesis, long bone growth, and neuronal maturation [3].
Apart from its role in hormone synthesis, iodine deficiency is associated with various health risks, including thyroid cancer [4]. Higher urinary iodine concentrations have been observed in breast cancer patients compared to controls, suggesting that urinary iodine concentrations might serve as a marker for breast cancer screening [5]. Additionally, improved iodine supply is linked to a decreased incidence of stomach cancer, indicating that iodine prophylaxis in deficient areas could protect against this disease [6]. These findings underscore the importance of addressing iodine deficiency in the environment.
Salt iodization is a common strategy to combat iodine deficiency, but several challenges must be addressed, such as iodine loss during storage and cooking. About 20% of iodine is lost during storage, and an additional 20–37% is lost during cooking or boiling [7]. Excessive salt consumption is associated with an increased risk of cardiovascular diseases [8], leading the World Health Organization to recommend limiting salt intake while ensuring that the salt used is iodized [9]. This highlights the need for alternative methods of iodine fortification.
Biofortification of plants is a promising and economically viable method to increase iodine intake. Unlike traditional fortification, which is based on the enrichment of macro- and micronutrients of the most frequently consumed food products with these elements in their mineral forms, biofortification enables the synthesis of organic iodine compounds within plant tissues, which are better absorbed and accumulated [10]. This method is also easily accepted by vegan diets [11,12]. However, caution is needed to avoid excessive iodine application to the soil, as it can harm crop yields [13].
Although iodine was not traditionally considered to be essential to plants and can be toxic if used excessively [13,14,15], new studies underline its important role in photosynthetic processes and defense mechanisms from abiotic and biotic stress through its involvement in various peroxidases [16]. The effects of iodine vary depending on plant species, application method, and soil properties [17,18]. Iodine accumulation in vegetables largely depends on xylem transportation [19], and foliar applications show promising results, especially in leafy vegetables. Soil-based biofortification strategies can be beneficial owing to the soil’s buffering capacity, which alleviates iodine’s toxic effects [18,20]. The true benefits, including human health and socio-economic advantages, lie within the health improvements brought by biofortification with iodine. Koronowicz et al. [11] indicate that the consumption of iodine-biofortified plants might hold potential for cancer treatment. Their study evaluates the effects of iodine-biofortified lettuce on the transcriptomic profile of the Caco-2 cancer cell line. They found that iodine, expected to act as a free ion or be partially incorporated into lettuce macromolecules, differently regulated the pathways of numerous transcription factors, leading to varied cellular effects. Specifically, they observed the inhibited proliferation of Caco-2 cells and the induced mitochondrial apoptosis and/or cell differentiation after treatment with biofortified lettuce, but not with potassium iodide (KI) alone. In light of Tonacchera et al. [21] demonstrating that the consumption of raw carrots biofortified with iodine through foliar fertilization during the growing season significantly increases urinary iodine concentrations in humans, potentially improving their iodine nutritional status, carrots are arguably a good target plant for using this agrotechnical method to gain potential health benefits.
The carrot (Daucus carota L.) is a globally important vegetable crop providing a source of important nutritional compounds through its carotenoid content, while adding flavor and texture to many dishes across the world [22]. Carrots are rich in nutritional compounds such as phenolics, carotenoids, polyacetylenes, sugars, and ascorbic acids, all of which provide antioxidant and anti-inflammatory properties. These compounds may reduce the risk of cancer and cardiovascular diseases [23]. Carrots also show a higher tolerance for iodine’s detrimental effects compared to other vegetables, making them suitable for iodine biofortification strategies [24].
Iodate is not as readily bioavailable as iodide, and its reduction would probably be needed prior to absorption. This is supported by the results of Kato et al. [25], who suggest that, during the cultivation of some plants such as rice, soybean, and also barley, the plants in question are able to reduce iodate to bioavailable iodide, with rhizosphere iodide being more bioavailable.
These studies underscore the potential of carrots as a dietary intervention to address iodine deficiency and explore new avenues in medical research, including cancer treatment. However, further groundwork is still needed before addressing the specifics of iodine’s health benefits, especially since research focused on the deposition of iodine in plant tissues and its transport within carrot plant organs is scarce. However, there is no reference value for organ-specific iodine allocation in carrots. Hong et al. [26], based on their study on pakchoi, celery, pepper, and radish, stated that iodine concentrations vary across different parts of a plant, typically following the sequence of roots > leaves > stems > fruits, with fruits generally containing the least iodine. This variance underscores the importance of effective iodine uptake and its potential implications for iodine content in dietary sources.
In soils, the application of iodate is preferred over iodide fortification. Several studies have demonstrated that the uptake of iodate from soils is generally more effective, likely due to its slower absorption by plants [18]. Because iodate is not as readily bioavailable as iodide, it must be reduced to iodide before uptake can occur, a process likely mediated biologically by plant roots or through microbial activity in the rhizosphere [25]. Additionally, interactions with humic substances may facilitate the reduction of iodate, as these compounds contain free redox-active groups in their structure [27].
As iodide is readily bioavailable, its use has been shown to have various adverse effects on lettuce and kohlrabi, leading them to exhibit common toxicity symptoms such as chlorosis and necrotic spots on the leaves. Lawson et al. [28], in their study, report a 20% decrease in the crop yield of plants fortified with iodide.
Some studies indicate that iodate is better absorbed by plants [18,29,30]; therefore, in this work, we choose both iodine species to determine which species would be more suitable for carrot biofortification. The aim of this study is to investigate the accumulation and distribution of both iodine species in various parts of carrots biofortified with iodide and iodate applied pre-sowing to the soil.
2. Materials and Methods
2.1. Carrot Cultivation
To study the accumulation and distribution of iodine in the different parts of a carrot, stock solutions of potassium iodide (KI) and potassium iodate (KIO3) with a concentration of 1000 mg∙L−1 were prepared by dissolving KI and KIO3 p.a. (Centralchem, Bratislava, Slovakia) in redistilled water. The soil used was identified as fluvisol (Akp horizon 0–30 cm) collected from an agricultural area in Žitný ostrov, Slovakia. The soil was loosened and air-dried. A total of 1 kg of soil was placed in plastic pots and thoroughly mixed with the iodine solutions to achieve a concentration of 10 mg∙kg−1 for each iodine species. The substrate was stabilized in the dark for 48 h. After this period, the soil was loosened again, and 20 carrot seeds (Daucus carota L., var. Korina, medium ripening, MoravoSeed, Mikulov, Czech Republic) were sown in each pot. Following germination, the number of seedlings was reduced to five per pot. The pots were placed outdoors in partial shade for 90 days from 2 May to 30 August 2023 and were watered as necessary to maintain optimal soil moisture for carrot growth.
2.2. Sample Processing
After three months, three carrot specimens were selected from each batch. The leaves and stems were separated, and the roots were peeled using a vegetable peeler. Each part was then cut into small pieces with a scalpel and dried at 25 °C for two weeks. After drying, the samples were crushed using a mortar and pestle. Representative soil samples were also collected from the pots. These soil samples were air-dried at room temperature and sieved through a 0.5 mm sieve to prepare them for iodine content determination.
2.3. Iodine Determination
Dried and ground biomass, weighing 0.25 g, was transferred to 15 mL centrifuge tubes with screw caps, to which 2 mL of 25% tetramethylammonium hydroxide solution (TMAH; Alfa Aesar, Karlsruhe, Germany) was added. The tubes were sealed and placed in a dry bath incubator (MK-20, Hangzhou Allsheng Instruments, Hangzhou, China) at 70 °C for 4 h and were subjected to manual shaking every 20 min. After extraction, the tubes were allowed to cool, and their contents were rinsed into 50 mL centrifuge tubes to a constant volume and centrifuged for 10 min at 10,000 rpm (Eppendorf 5804 R, Hamburg, Germany). The supernatant was then filtered through a nitrocellulose mixed ester membrane filter with a pore size of 0.45 µm (Advantec, Tokyo, Japan).
The iodine content in the soil was determined as follows: 100 mg of finely ground soil and 5 mL of 5% TMAH was added to 15 mL disposable polypropylene centrifuge tubes with screw caps. The tubes were placed in a dry bath and incubated for 4 h at 90 °C, with manual shaking every 20 min. After incubation, the samples were centrifuged (CM-6MT, ELMI, Riga, Latvia) at 2300 rpm for 30 min. The supernatant was then diluted with redistilled water to a final volume of 50 mL, resulting in a 1% TMAH concentration in the solution.
The iodine content of the solutions was measured using an inductively coupled plasma mass spectrometer (ICP-MS) (Thermo Scientific iCAP Q, Waltham, MA, USA), with 126Te used as an internal standard.
Calibration and standard solutions for ICP-MS were prepared from a standard KI solution (0.1 mol·L−1, HI 4011-01, Hanna Instruments, Villafranca Padovana, Italy) which included a 0.5% TMAH solution. The standard ICP-MS operating mode was used, with a calibration range of 0–127 µg·L−1. The 0.01 µg·L−1 LOD was calculated as 3× the standard deviation of the blank determination, while the 0.03 µg·L−1 LOQ was calculated as 10× the standard deviation of the blank determination.
The operating parameters for the ICP-MS are shown in Table 1. To evaluate the extraction efficiency of this method, certified reference materials (CRM) were used which are listed in Table 2.

Table 1.
ICP-MS operating conditions for 127I determination (standard mode).
Table 2.
Extraction efficiency of the used certified reference materials.
The uptake of iodine by carrots from the soil was described by the substrate to plant transfer factor (TF), which, in this case, was calculated as the iodine concentration in the plant parts of a carrot (mg kg−1) dry weight divided by the remaining iodine concentration in the substrate (dry weight), as follows:
3. Results and Discussion
The background iodine concentration in the untreated soil was determined through triplicate sampling to ensure the accuracy and reliability of the data. The results indicate that the soil used in this study is naturally iodine deficient, with a background iodine concentration of 0.63 mg·kg−1. This confirms that the soil provides a minimal iodine baseline, knowledge which is important to evaluate the effects of iodine fertigation treatments. The iodine concentration in the fertigated soil was measured both before sowing and after the 90-day cultivation period, following harvest. This approach allowed for the monitoring of any potential changes in iodine content over the course of plant growth and nutrient uptake.
The iodine concentration in the soil treated with KI was found to be 10.4 mg·kg−1, which decreased by 17% by the end of the cultivation period, probably due to volatilization and washout by the irrigation water. The iodine concentration in the soil treated with KIO3 was 9.9 mg·kg−1, with only a 9% decrease observed during the cultivation period. This finding highlights the potential retention by the soil of iodine supplied as KIO3, a phenomenon which could have implications for the sustained effectiveness of iodine fertigation practices in future crop cycles. The iodine concentrations of the used soil are presented in Table 3. The fresh and dry weight of the carrot parts are presented in Table 4. No statistically significant difference between control and iodide- or iodate-treated samples was observed based on fresh or dry weight.
Table 3.
Iodine content of the soil used for cultivation.
Table 4.
Fresh and dry biomass weight.
In control carrots, the distribution of iodine was relatively uniform across the various parts of the plant, with an average concentration of around 0.85 mg·kg−1 dry weight. This consistent distribution suggests that, in the absence of iodine fortification, the uptake and allocation of iodine by the plant are limited and evenly spread throughout the carrot. However, in biofortified carrots, the distribution of iodine varied significantly, following a clear descending order: leaf > stem > root peel > root. This indicates that soil fertigation leads to much higher iodine accumulation in the aerial parts of the plant, particularly in the leaves which develop during the early growth stages, with decreasing amounts in the stem and roots. The root peel, the outer layer of carrot roots, also exhibited notable iodine accumulation compared to the inner root tissue.
The differences in iodine concentration across these plant parts highlight the plant’s natural mechanisms of iodine transport and storage, with the aerial parts acting as primary sinks. These findings are further illustrated in Figure 1, where the distribution and content of iodine in the different parts of the carrot are depicted.
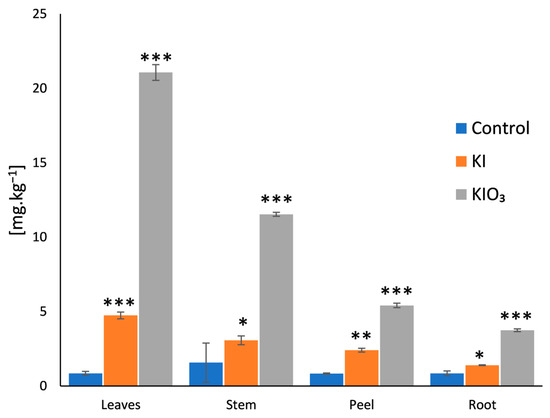
Figure 1.
Iodine accumulation in different plant parts. The columns represent the arithmetic means with the standard deviations (n = 5). Differences were assessed at * p = 0.05, ** p = 0.01, *** p = 0.001 using the student’s t-test for comparison with the control.
Biofortified carrots treated with iodate accumulated 2.7 times more iodine in their edible parts compared to those treated with iodide, indicating the significantly higher efficiency of iodate as a biofortification agent. The leaves contained the highest iodine concentrations in both treatments. Specifically, iodine levels in the leaves of iodide-treated samples were more than five times higher than those found in the control vegetables, while, in iodate-treated samples, iodine levels were over 24 times higher than those found in the control carrots. This dramatic increase suggests that iodine uptake from iodate is more effective in promoting iodine accumulation in the aerial parts of the plant.
The stems (aerial parts without the leaves) also showed substantial iodine accumulation, with more than seven times the iodine concentration found in untreated carrots when treated with iodate. Given that iodine content in the control carrots was relatively uniform across the different plant parts, these results suggest that naturally occurring iodine may undergo “aging”, becoming strongly bound to soil particles and, thus, less bioavailable. In contrast, newly introduced iodine through fertigation with KI and KIO3 solutions appears to be readily bioavailable, particularly during the early stages of plant development, and is efficiently transported via the xylem to the leaves and other aerial parts.
The ability of the leaves and stems to act as iodine sinks is particularly noteworthy, as it opens up additional opportunities for utilizing iodine-enriched plant material, for example as livestock feed or as nutrient-rich compost for further biofortification strategies.
These findings align with those reported by Hong et al. [24], who found that iodine levels in the carrot rhizome were about 50% of those found in the shoots, supporting the conclusion that iodine accumulation is significantly higher in the aboveground parts of the plant. A comparison of the accumulation rates observed in this study with data from other researchers is presented in Table 5, illustrating how different iodine species and treatments influence the overall iodine content in the various parts of a carrot.
Table 5.
Iodine accumulation in different carrot parts.
The root peel also accumulated a significant amount of iodine, containing 2.41 mg·kg−1 iodine following the iodide treatments and 5.42 mg·kg−1 dry weight iodine following the iodate treatments. Results obtained by Kapusta-Duch et al. [33] similarly indicate substantial iodine accumulation in the removed parts of carrot roots, suggesting that iodine accumulation in the periderm of storage roots may be the result of a plant defense mechanism against excessive intake of this trace element. Cooking unpeeled biofortified carrots has also been shown to result in higher iodine contents compared to peeled carrots [34]. The significance of the outer layer in alleviating the rate of iodine transfer has also been demonstrated in other root plants. Cerretani et al. [35] suggested that the potato skin might act as a physical barrier, limiting the leaching of iodine into the boiling water. However, it remains unclear whether this is due to the barrier effect or, simply, to the higher iodine accumulation in the root peel. Soil-to-plant transfer factors with the total iodine concentration in each plant part are presented in Table 6. The percentual distribution of the total accumulated iodine in different plant parts is depicted in Figure 2.
Table 6.
Soil-to-plant transfer factor (TF) values and total iodine concentrations in different plant parts.
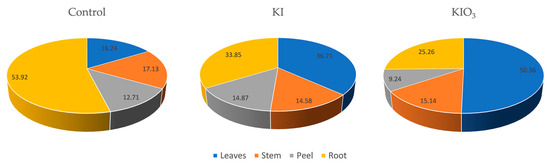
Figure 2.
Percentual distribution of the total accumulated iodine across different plant parts.
Although the available studies do not directly present the iodine content of the peel, it can be inferred that the higher iodine accumulation rate in the peel of biofortified carrots through soil fertigation plays a significant role. The first barrier in nutrient uptake from the soil is the outer tissues, comprising the rhizodermis or periderm. Bhandari et al. [36] reveal that the outer tissues of carrot roots contain higher levels of bioactive compounds such as free sugars, carotenoids, ascorbic acid, total phenols, and total flavonoids compared to the inner tissues. Thus, it is probable that the outer tissues of carrot roots, which also include part of the phloem, act as a sink for iodine deposited from the aerial parts of the plant via the phloem stream. Assuming the lower iodine content in the xylem and the higher iodine content in the aerial parts, this suggests a highly efficient transport of iodine from the roots through the xylem via the transpiration stream. However, despite the efficient transport, peeled roots of iodate-fortified carrots showed significantly higher iodine accumulation, reaching 3.75 mg·kg−1 dry weight, compared to the untreated samples. This is more than 4.3 times the amount found in untreated controls and 2.68 times higher than in carrots biofortified with iodide.
The high iodine content in iodate-fortified leaves, at 21 mg·kg−1 dry weight, makes dry carrot leaves a valuable addition to hay feed for livestock. This is also evident from Figure 2: iodine-treated carrots showed higher accumulated iodine levels in the leaves compared to the controls. Additionally, the iodate treatments resulted in a higher accumulation rate in the leaves compared to those engendered by the iodide treatments, whereas the iodide distribution throughout the plant was more uniform. The substantial iodine concentrations in these leaves not only allow for an efficient way to utilize all parts of the carrot plant, but also offers a potential solution for increasing iodine intake in livestock, which can be beneficial for their overall health and growth. Biofortified carrots grown in an open field can significantly contribute to meeting the recommended daily allowance (RDA) of iodine. Consuming 100 g of fresh carrots biofortified with a 50 mg·L−1 iodine solution can satisfy the RDA, while 200 g of carrots treated with a 500 mg·L−1 solution can meet the requirement [37]. Additionally, consuming 100 g of fresh carrots biofortified with 0.5 mg·L−1 iodide-containing water would cover approximately 38% of an adult’s daily iodine intake requirement [32]. In our case, considering the iodine concentration and moisture content of the carrot roots, consuming 100 g of fresh iodate-fortified carrots could cover 37% of the recommended daily allowance for an adult, a finding which is in good agreement with previous reports.
However, from a consumer’s perspective, the stability of enriched nutrients during processing and cooking is crucial, especially with respect to iodine due to its volatile nature. Biofortification can affect the quality during storage. Smoleń et al. [38] report that iodine (both KI and KIO3) supplied with relatively high doses of nitrogen decreased the quality of fresh carrots. However, after storage, the opposite relations were noted, suggesting a need for developing specific agronomic rules for iodine-fortified carrots intended for immediate consumption versus long-term storage. Skoczylas et al. [39] report an iodine loss index in the range of 36–39% during the production process (washing, peeling, blanching, juicing, sterilization, and storage) of carrot juice from non-fortified carrots. Most of the iodine in biofortified vegetables is present mostly in mineral forms, which can easily dissolve in water during cooking or blanching [40].
Piątkowska et al. [12] suggest that cooking not only causes iodine, loss but also affects its bioavailability from biofortified carrots. Cooking processes impact the quantity of iodine due to its volatile nature, with mean losses during different procedures in descending order: boiling (37–40%), microwave cooking (27%), shallow frying (27%), pressure cooking (22%), steaming (20%), deep frying (10–20%), and roasting (6–10%) [41,42]. Lower iodine concentrations in cooked and steamed biofortified carrots are reported by Kapusta-Duch et al. [33], noting that increased cooking time significantly decreases the iodine content. Biofortified peeled carrots cooked for 30 min or 40 min had lower iodine concentrations than raw control carrots. Comandini et al. [43] suggest that the low starch content in carrots could contribute to a lower iodine retention during cooking compared to potatoes, where iodine can be sequestered within the helical V-amylose component of the starch [44]. To minimize iodine losses during cooking, it is advisable to sprinkle salt on food after cooking rather than during [42]. Nonetheless, vegetables are excellent candidates for biofortification with trace elements, as they can often be consumed raw without thermal processing.
Statistically significantly higher iodine levels have been observed in the urine, feces, and selected tissues of rats fed a diet containing biofortified raw carrots compared to those on a diet without iodine and those consuming control cooked carrots. The intake of biofortified raw carrots significantly increased triiodothyronine concentrations compared to animals in other experimental groups. The highest thyroid-stimulating hormone levels were found in rats fed control cooked carrots. Additionally, the mRNA expression of selected genes in the rats’ heart was affected by different dietary treatments. These findings suggest that both biofortified raw and cooked carrots could be potential sources of iodine in daily diets to prevent iodine deficiency across various populations [12].
While we did not evaluate the qualitative parameters of carrots or observe any inhibitory effects on carrot development in our experiment, other studies have reported changes in the quality of biofortified carrots. However, iodine fertilization did not reduce yield and had no adverse effect on other nutritional characteristics of carrot roots according to Smoleń et al. [31]; from a consumer’s perspective, such biofortified carrots maintain their characteristics and can serve as additional sources of iodine. On the other hand, Hung et al. [45] suggest that iodate application might weaken nitrate reduction, leading to excessive nitrate accumulation, although no excessive accumulation of nitrates in carrot tissues was observed by Smoleń et al. [46] after KIO3 application.
Iodine fortification might result in a lighter coloring and an increased pH value of carrot juices depending on carrot cultivars. Additionally, it has been found to contribute to a slight decrease in dry matter, total soluble solids, protein, sugars, and β-carotene in juices compared to non-biofortified orange cultivars [39]. However, Smoleń et al. [31,47], based on a three-year study, suggest that these changes in the chemical composition of carrot roots might be more dependent on the cultivation period than on the application of iodine.
Kapusta-Duch et al. [33] observed a significant increase in total carotenoids in peeled, cooked, and steamed carrots biofortified with iodine compared to raw biofortified carrots. Carotenoids are converted to vitamin A in the body, a process which is essential for cell division, proliferation, and growth. Research indicates that vitamin A levels modulate thyroid gland homeostasis, alone or in interaction with other micronutrients, particularly iodine [48,49]. It is suggested that, after conversion to vitamin A, carotenoids influence thyroid hormone production, leading to higher body gain and feed efficiency ratios in rats fed cooked carrots biofortified with iodine [12].
Biofortified carrot juice from orange cultivars has been found to contain higher levels of caffeic, ferulic, and salicylic acids than analogous juices, except for those from the cultivar Kazan F1. However, biofortification has been found to decrease (+) catechin contents regardless of the carrot cultivar used for juice production [39].
Antioxidant activity has been found to be significantly higher in thermally processed, both unpeeled and peeled, biofortified carrots compared to raw biofortified carrots [33], although a slight decrease in antioxidant activity against DPPH has been documented in juices made from carrots fortified with iodine and Se [39]. Conversely, Bieżanowska-Kopeć et al. [34] observed an increase in antioxidant contents, along with iron and polyphenols, in carrots fortified with iodate.
4. Conclusions
This study demonstrates the effectiveness of iodine biofortification in carrots, with potassium iodate (KIO3) proving more efficient than potassium iodide (KI) in increasing iodine content across different parts of the carrot plant. These findings underscore the potential of iodate fortification as a superior method for biofortifying carrots with iodine. The enhanced iodine content in both the roots and leaves of iodate-fortified carrots highlights the importance of selecting appropriate fortification compounds to achieve the desired nutritional outcomes. As we continue to explore and refine biofortification techniques, it is essential to consider the broader implications for food security, public health, and agricultural sustainability. In addition to their nutritional benefits, the use of iodate-fortified carrot leaves as an iodine-rich fertilizer can contribute to sustainable agricultural practices. By maximizing the utilization of the entire plant, farmers can reduce waste and improve the economic value of their crops. Moreover, incorporating iodine-rich feed into livestock diets can enhance the nutritional quality of animal products such as milk and meat, thereby indirectly benefiting human health.
Author Contributions
Conceptualization, E.D. and M.U.; Methodology, E.D. and M.B.; Validation, E.D. and M.B.; Formal analysis, P.D.; Investigation, E.D. and M.U.; Writing—original draft, E.D., P.M. and M.U.; Writing—review & editing, E.D., P.M., P.D. and M.U. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Scientific Grant Agency of the Ministry of Education of the Slovak Republic and the Slovak Academy of Sciences under the contracts VEGA No. 1/0139/22 and by the Specific research project from Brno University of Technology, No. FCH-S-24-8597.
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author/s.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sorrenti, S.; Baldini, E.; Pironi, D.; Lauro, A.; D’Orazi, V.; Tartaglia, F.; Tripodi, D.; Lori, E.; Gagliardi, F.; Praticò, M.; et al. Iodine: Its Role in Thyroid Hormone Biosynthesis and Beyond. Nutrients 2021, 13, 4469. [Google Scholar] [CrossRef]
- Andersson, M.; Braegger, C.P. The Role of Iodine for Thyroid Function in Lactating Women and Infants. Endocr. Rev. 2021, 43, 469–506. [Google Scholar] [CrossRef] [PubMed]
- Mégier, C.; Dumery, G.; Luton, D. Iodine and Thyroid Maternal and Fetal Metabolism during Pregnancy. Metabolites 2023, 13, 633. [Google Scholar] [CrossRef]
- Fan, L.; Meng, F.; Gao, Y.; Liu, P. Insufficient iodine nutrition may affect the thyroid cancer incidence in China. Br. J. Nutr. 2021, 126, 1852–1860. [Google Scholar] [CrossRef] [PubMed]
- Malya, F.U.; Kadioglu, H.; Hasbahceci, M.; Dolay, K.; Guzel, M.; Ersoy, Y.E. The correlation between breast cancer and urinary iodine excretion levels. J. Int. Med. Res. 2017, 46, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Gołkowski, F.; Szybiński, Z.; Rachtan, J.; Sokołowski, A.; Buziak-Bereza, M.; Trofimiuk, M.; Hubalewska-Dydejczyk, A.; Przybylik-Mazurek, E.; Huszno, B. Iodine prophylaxis—The protective factor against stomach cancer in iodine deficient areas. Eur. J. Nutr. 2007, 46, 251–256. [Google Scholar] [CrossRef]
- WHO. Recommended Iodine Levels in Salt and Guidelines for Monitoring Their Adequacy and Effectiveness; World Health Organization: Geneva, Switzerland, 1996. [Google Scholar]
- He, F.J.; Burnier, M.; MacGregor, G.A. Nutrition in cardiovascular disease: Salt in hypertension and heart failure. Eur. Heart J. 2011, 32, 3073–3080. [Google Scholar] [CrossRef]
- WHO. Global Strategy on Diet, Physical Activity and Health; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Smoleń, S.; Kowalska, I.; Halka, M.; Ledwożyw-Smoleń, I.; Grzanka, M.; Skoczylas, Ł.; Czernicka, M.; Pitala, J. Selected Aspects of Iodate and Iodosalicylate Metabolism in Lettuce Including the Activity of Vanadium Dependent Haloperoxidases as Affected by Exogenous Vanadium. Agronomy 2020, 10, 1. [Google Scholar] [CrossRef]
- Koronowicz, A.A.; Kopeć, A.; Master, A.; Smoleń, S.; Piątkowska, E.; Bieżanowska-Kopeć, R.; Ledwożyw-Smoleń, I.; Skoczylas, Ł.; Rakoczy, R.; Leszczyńska, T.; et al. Transcriptome Profiling of Caco-2 Cancer Cell Line following Treatment with Extracts from Iodine-Biofortified Lettuce (Lactuca sativa L.). PLoS ONE 2016, 11, e0147336. [Google Scholar] [CrossRef]
- Piątkowska, E.; Kopeć, A.; Bieżanowska-Kopeć, R.; Pysz, M.; Kapusta-Duch, J.; Koronowicz, A.A.; Smoleń, S.; Skoczylas, Ł.; Ledwożyw-Smoleń, I.; Rakoczy, R.; et al. The Impact of Carrot Enriched in Iodine through Soil Fertilization on Iodine Concentration and Selected Biochemical Parameters in Wistar Rats. PLoS ONE 2016, 11, e0152680. [Google Scholar] [CrossRef]
- Dai, J.L.; Zhu, Y.G.; Zhang, M.; Huang, Y.Z. Selecting iodine-enriched vegetables and the residual effect of iodate application to soil. Biol. Trace Elem. Res. 2004, 101, 265–276. [Google Scholar] [CrossRef]
- Dai, J.L.; Zhu, Y.G.; Huang, Y.Z.; Zhang, M.; Song, J.L. Availability of iodide and iodate to spinach (Spinacia oleracea L.) in relation to total iodine in soil solution. Plant Soil 2006, 289, 301–308. [Google Scholar] [CrossRef]
- Duborská, E.; Urík, M.; Kubová, J. Interaction with soil enhances the toxic effect of iodide and iodate on barley (Hordeum vulgare L.) compared to artificial culture media during initial growth stage. Arch. Agron. Soil Sci. 2018, 64, 46–57. [Google Scholar] [CrossRef]
- Kiferle, C.; Martinelli, M.; Salzano, A.M.; Gonzali, S.; Beltrami, S.; Salvadori, P.A.; Hora, K.; Holwerda, H.T.; Scaloni, A.; Perata, P. Evidences for a Nutritional Role of Iodine in Plants. Front. Plant Sci. 2021, 12, 616868. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Prom-u-thai, C.; Guilherme, L.R.G.; Rashid, A.; Hora, K.H.; Yazici, A.; Savasli, E.; Kalayci, M.; Tutus, Y.; Phuphong, P.; et al. Iodine biofortification of wheat, rice and maize through fertilizer strategy. Plant Soil 2017, 418, 319–335. [Google Scholar] [CrossRef]
- Duborská, E.; Urík, M.; Bujdoš, M.; Kubová, J. Aging and Substrate Type Effects on Iodide and Iodate Accumulation by Barley (Hordeum vulgare L.). Water Air Soil Pollut. 2016, 227, 407. [Google Scholar] [CrossRef]
- Herrett, R.A.; Hatfield, H.H., Jr.; Crosby, D.G.; Vlitos, A.J. Leaf abscission induced by the iodide ion. Plant Physiol. 1962, 37, 358. [Google Scholar] [CrossRef]
- Mackowiak, C.L.; Grossl, P.R.; Cook, K.L. Iodine toxicity in a plant-solution system with and without humic acid. Plant Soil 2005, 269, 141–150. [Google Scholar] [CrossRef]
- Tonacchera, M.; Dimida, A.; De Servi, M.; Frigeri, M.; Ferrarini, E.; De Marco, G.; Grasso, L.; Agretti, P.; Piaggi, P.; Aghini-Lombardi, F.; et al. Iodine Fortification of Vegetables Improves Human Iodine Nutrition: In Vivo Evidence for a New Model of Iodine Prophylaxis. J. Clin. Endocrinol. Metab. 2013, 98, E694–E697. [Google Scholar] [CrossRef]
- Augšpole, I.; Kince, T.; Dukalska, L. Content of sugars, dietary fibre and vitamin C in hybrids of “Nante” carrots cultivated in Latvia. Res. Rural Dev. 2012, 1, 137–142. [Google Scholar]
- Ahmad, T.; Cawood, M.; Iqbal, Q.; Ariño, A.; Batool, A.; Tariq, R.M.S.; Azam, M.; Akhtar, S. Phytochemicals in Daucus carota and Their Health Benefits-Review Article. Foods 2019, 8, 424. [Google Scholar] [CrossRef]
- Hong, C.L.; Weng, H.X.; Qin, Y.C.; Yan, A.L.; Xie, L.L. Transfer of iodine from soil to vegetables by applying exogenous iodine. Agron. Sustain. Dev. 2008, 28, 575–583. [Google Scholar] [CrossRef]
- Hong, C.-L.; Weng, H.-X.; Yan, A.L.; Xie, L.-L. Characteristics of iodine uptake and accumulation by vegetables. Yingyong Shengtai Xuebao 2007, 18, 2313–2318. [Google Scholar]
- Kato, S.; Wachi, T.; Yoshihira, K.; Nakagawa, T.; Ishikawa, A.; Takagi, D.; Tezuka, A.; Yoshida, H.; Yoshida, S.; Sekimoto, H.; et al. Rice (Oryza sativa L.) roots have iodate reduction activity in response to iodine. Front. Plant Sci. 2013, 4, 47204. [Google Scholar] [CrossRef]
- Reiller, P.; Mercier-Bion, F.; Gimenez, N.; Barré, N.; Miserque, F. Iodination of humic acid samples from different origins. Radiochim. Acta 2006, 94, 739. [Google Scholar] [CrossRef]
- Lawson, P.G.; Daum, D.; Czauderna, R.; Meuser, H.; Hartling, J.W. Soil versus foliar iodine fertilization as a biofortification strategy for field-grown vegetables. Front. Plant Sci. 2015, 6, 450. [Google Scholar] [CrossRef]
- Smolen, S.; Sady, W. Influence of iodine form and application method on the effectiveness of iodine biofortification, nitrogen metabolism as well as the content of mineral nutrients and heavy metals in spinach plants (Spinacia oleracea L.). Sci. Hortic. 2012, 143, 176–183. [Google Scholar] [CrossRef]
- Gonzali, S.; Kiferle, C.; Perata, P. Iodine biofortification of crops: Agronomic biofortification, metabolic engineering and iodine bioavailability. Curr. Opin. Biotechnol. 2017, 44, 16–26. [Google Scholar] [CrossRef]
- Smoleń, S.; Baranski, R.; Ledwożyw-Smoleń, I.; Skoczylas, Ł.; Sady, W. Combined biofortification of carrot with iodine and selenium. Food Chem. 2019, 300, 125202. [Google Scholar] [CrossRef] [PubMed]
- Dobosy, P.; Endrédi, A.; Sandil, S.; Vetési, V.; Rékási, M.; Takács, T.; Záray, G. Biofortification of Potato and Carrot With Iodine by Applying Different Soils and Irrigation With Iodine-Containing Water. Front. Plant Sci. 2020, 11, 593047. [Google Scholar] [CrossRef]
- Kapusta-Duch, J.; Bieżanowska-Kopeć, R.; Smoleń, S.; Pysz, M.; Kopeć, A.; Piątkowska, E.; Rakoczy, R.; Koronowicz, A.; Skoczylas, Ł.; Leszczyńska, T. The effect of preliminary processing and different methods of cooking on the iodine content and selected antioxidative properties of carrot (L.) biofortified with (potassium) iodine. Folia Hortic. 2017, 29, 11–24. [Google Scholar] [CrossRef]
- Bieżanowska-Kopeć, R.; Pysz, M.; Kapusta-Duch, J.; Kopeć, A.; Smoleń, S.; Koronowicz, A.; Piątkowska, E.; Rakoczy, R.; Skoczylas, Ł.; Leszczyńska, T. The effects of peeling and cooking on the mineral content and antioxidant properties in carrots enriched with potassium iodate and/or selenite (SeIV) and selenite (SeVI). Int. J. Food Sci. Nutr. 2016, 67, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Cerretani, L.; Comandini, P.; Fumanelli, D.; Scazzina, F.; Chiavaro, E. Evaluation of iodine content and stability in recipes prepared with biofortified potatoes. Int. J. Food Sci. Nutr. 2014, 65, 797–802. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Choi, C.S.; Rhee, J.; Shin, Y.K.; Song, J.W.; Kim, S.H.; Kang, S.; Lee, J.G. Influence of Root Color and Tissue on Phytochemical Contents and Antioxidant Activities in Carrot Genotypes. Foods 2022, 12, 120. [Google Scholar] [CrossRef] [PubMed]
- Signore, A.; Renna, M.; D’Imperio, M.; Serio, F.; Santamaria, P. Preliminary Evidences of Biofortification with Iodine of “Carota di Polignano”, An Italian Carrot Landrace. Front. Plant Sci. 2018, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- Smoleń, S.; Sady, W.; Ledwożyw-Smoleń, I.; Strzetelski, P.; Liszka-Skoczylas, M.; Rożek, S. Quality of fresh and stored carrots depending on iodine and nitrogen fertilization. Food Chem. 2014, 159, 316–322. [Google Scholar] [CrossRef]
- Skoczylas, Ł.; Tabaszewska, M.; Smoleń, S.; Słupski, J.; Liszka-Skoczylas, M.; Barański, R. Carrots (Daucus carota L.) Biofortified with Iodine and Selenium as a Raw Material for the Production of Juice with Additional Nutritional Functions. Agronomy 2020, 10, 1360. [Google Scholar] [CrossRef]
- Smoleń, S.; Kowalska, I.; Kováčik, P.; Halka, M.; Sady, W. Biofortification of Six Varieties of Lettuce (Lactuca sativa L.) with Iodine and Selenium in Combination with the Application of Salicylic Acid. Front. Plant Sci. 2019, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Goindi, G.; Karmarkar, M.G.; Kapil, U.; Jagannathan, J. Estimation of losses of iodine during different cooking procedures. Asia Pac. J. Clin. Nutr. 1995, 4, 225–227. [Google Scholar]
- Rana, R.; Raghuvanshi, R.S. Effect of different cooking methods on iodine losses. J. Food Sci. Technol. 2013, 50, 1212–1216. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Comandini, P.; Cerretani, L.; Rinaldi, M.; Cichelli, A.; Chiavaro, E. Stability of iodine during cooking: Investigation on biofortified and not fortified vegetables. Int. J. Food Sci. Nutr. 2013, 64, 857–861. [Google Scholar] [CrossRef]
- Yu, X.; Houtman, C.; Atalla, R.H. The complex of amylose and iodine. Carbohydr. Res. 1996, 292, 129–141. [Google Scholar] [CrossRef]
- Hung, C.-C.; Wong, G.; Dunstan, W. Iodate Reduction Activity in Nitrate Reductase Extracts from Marine Phytoplankton. Bull. Mar. Sci. 2005, 76, 61–72. [Google Scholar]
- Smoleń, S.; Skoczylas, Ł.; Ledwożyw-Smoleń, I.; Rakoczy, R.; Liszka-Skoczylas, M.; Kopeć, A.; Piątkowska, E.; Bieżanowska-Kopeć, R.; Koronowicz, A.; Kapusta-Duch, J.; et al. The quality of carrot (L.) cultivated in the field depending on iodine and selenium fertilization. Folia Hortic. 2016, 28, 151–164. [Google Scholar] [CrossRef]
- Smoleń, S.; Skoczylas, Ł.; Ledwożyw-Smoleń, I.; Rakoczy, R.; Kopeć, A.; Piątkowska, E.; Bieżanowska-Kopeć, R.; Koronowicz, A.; Kapusta-Duch, J. Biofortification of Carrot (Daucus carota L.) with Iodine and Selenium in a Field Experiment. Front. Plant Sci. 2016, 7, 730. [Google Scholar] [CrossRef]
- Capriello, S.; Stramazzo, I.; Bagaglini, M.F.; Brusca, N.; Virili, C.; Centanni, M. The relationship between thyroid disorders and vitamin A: A narrative minireview. Front. Endocrinol. 2022, 13, 968215. [Google Scholar] [CrossRef]
- Carazo, A.; Macáková, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients 2021, 13, 1703. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).