
Effects of Drought Stress and Postdrought Rewatering on Winter Wheat: A Meta-Analysis

Abstract
1. Introduction
2. Materials and Methods
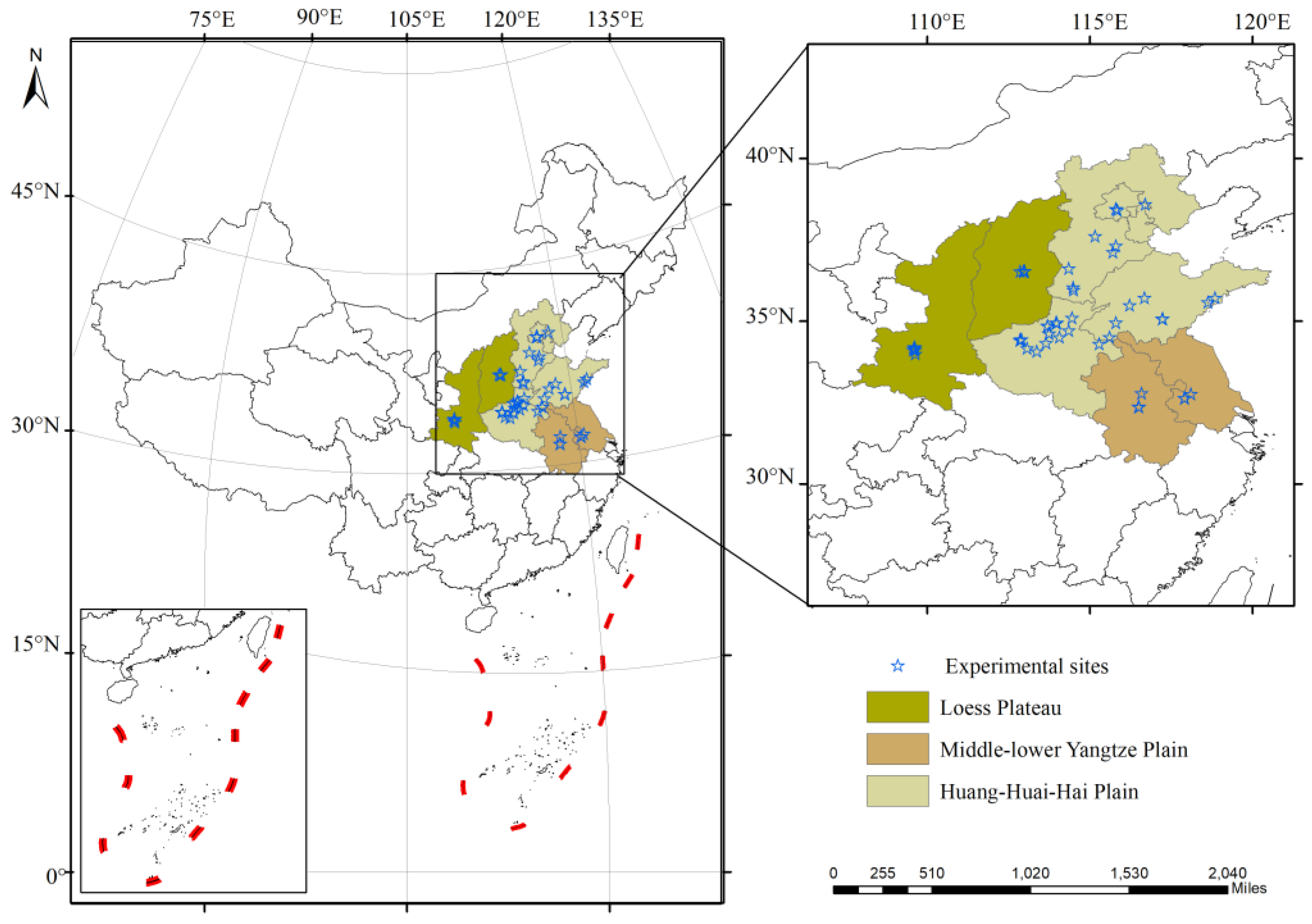
2.1. Data Search and Collection
2.2. Inclusion Criteria
- (1)
- The experimental crop was defined as winter wheat;
- (2)
- The planting methods must be field-grown or pot-grown;
- (3)
- (4)
- The experiment must include a well-watered control group and a drought stress (or postdrought rewatering) treatment group;
- (5)
- Must be a controlled experiment with rain protection to prevent natural precipitation;
- (6)
- The intensity of drought stress must be delineated by relative soil water content (RSWC) from 0–20 cm;
- (7)
- The data contain any of the plant growth parameters for winter wheat listed in Table 1;
- (8)
- Data from the same experiment appearing in different papers were included only once.
2.3. Data Classification and Subgroup Analysis
- (1)
- According to the 0–20 cm of RSWC, it was divided into three drought stress intensities: mild stress (55% < RSWC ≤ 65%), moderate stress (45% < RSWC ≤ 55%), and severe stress (RSWC ≤ 45%). The relative soil water content was calculated as follows:
- (2)
- The drought stress subdatabase was divided according to the duration of drought stress, and the rewatering subdatabase was divided according to the duration of rewatering. The number of days of duration was calculated from the beginning of the corresponding treatment to the end of the day of the measurement, which was divided into the following categories: 1–3 days, 4–7 days, 8–15 days, and >15 days;
- (3)
- The growth stages were divided as shown in Table 2, which includes single-stage drought and multi-stage successive drought;
- (4)
- The planting methods are classified as field and pot;
- (5)
- The experimental areas were divided into the Loess Plateau, the Huang-Huai-Hai Plain, and the Middle–Lower Yangtze Plain.
2.4. Statistical Analyses
3. Results
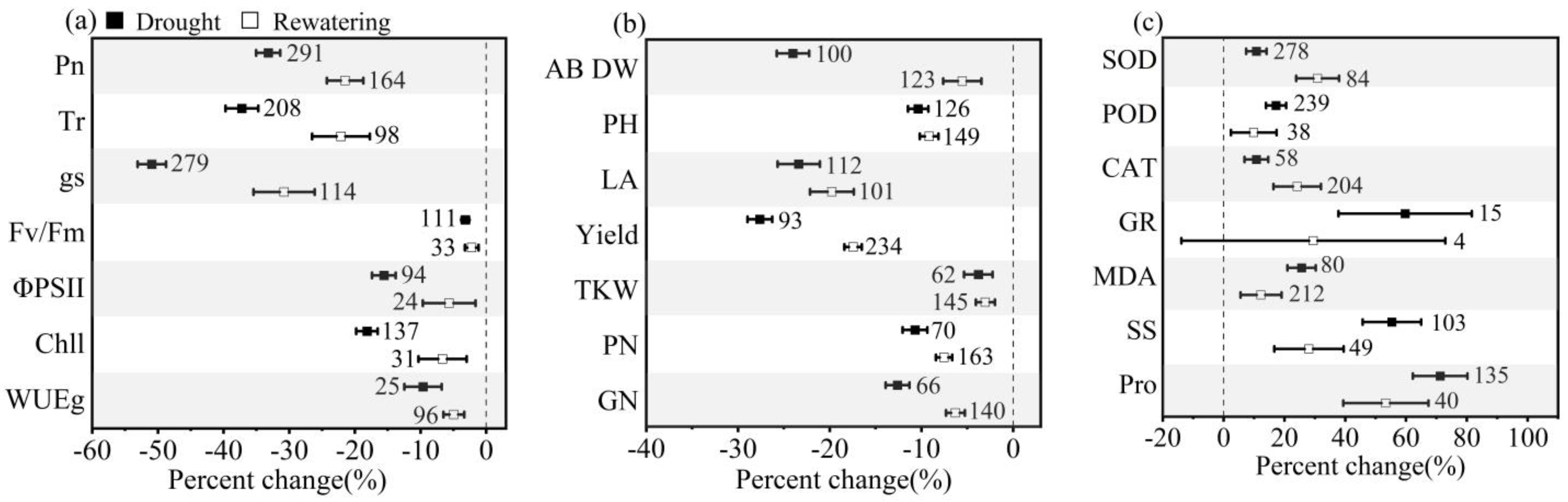
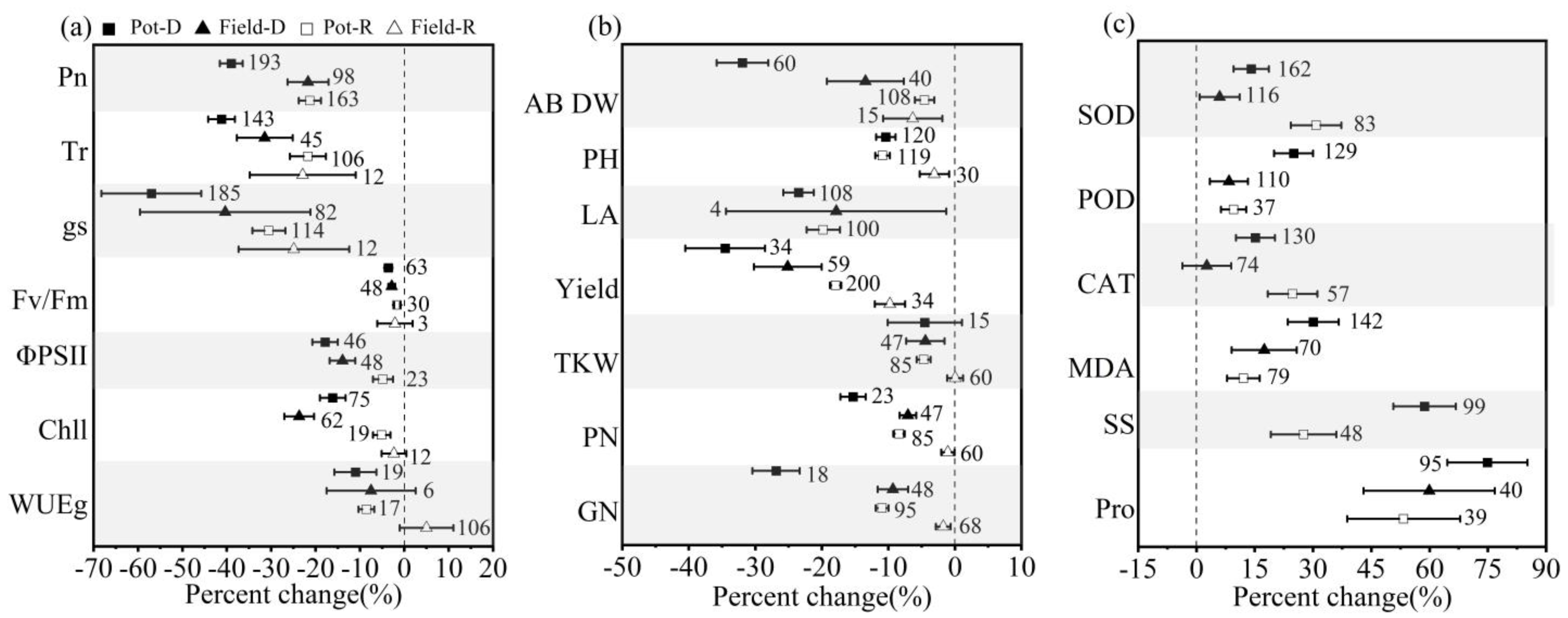
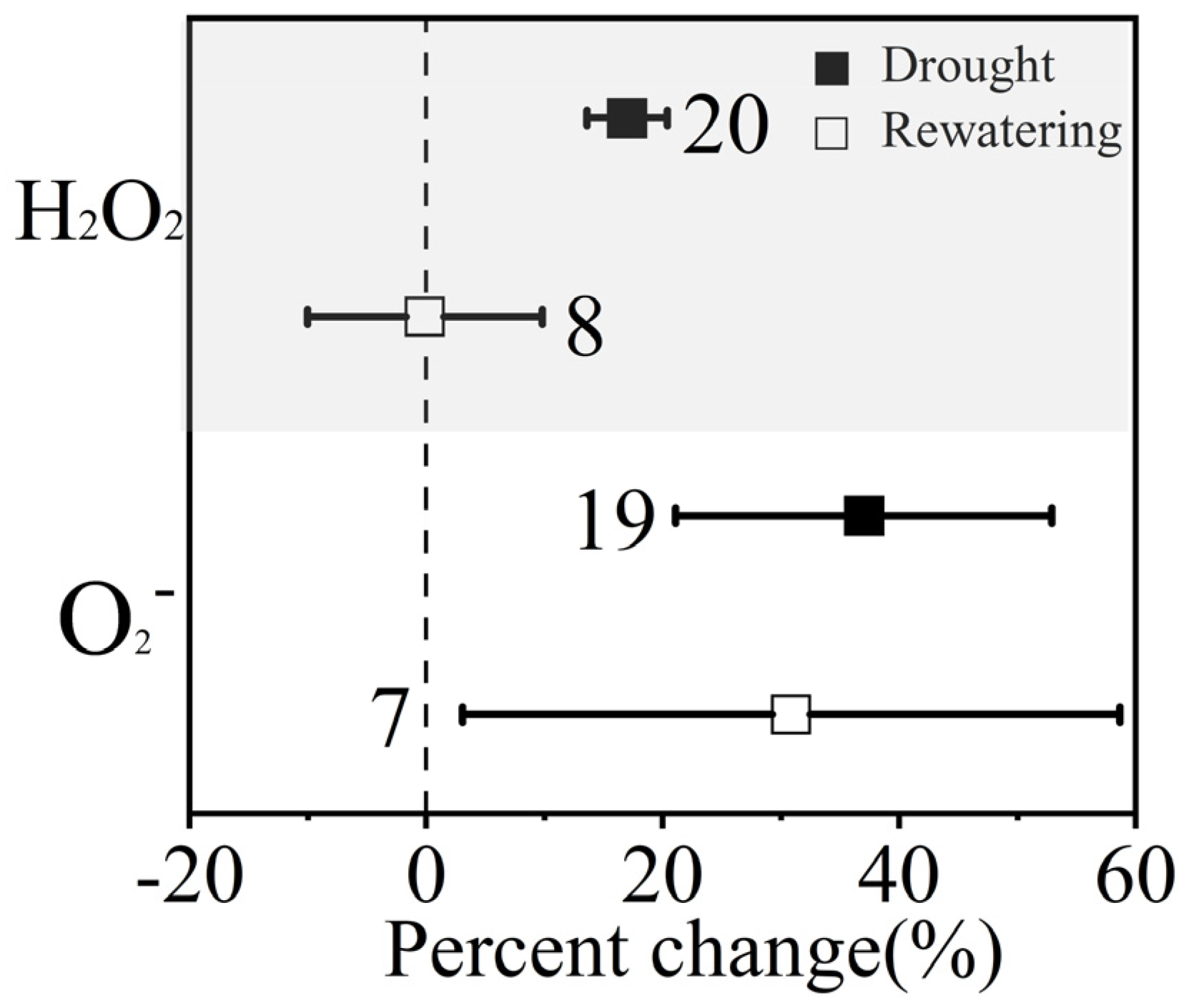
3.1. Overall Effects of Drought Stress and Postdrought Rewatering on Winter Wheat
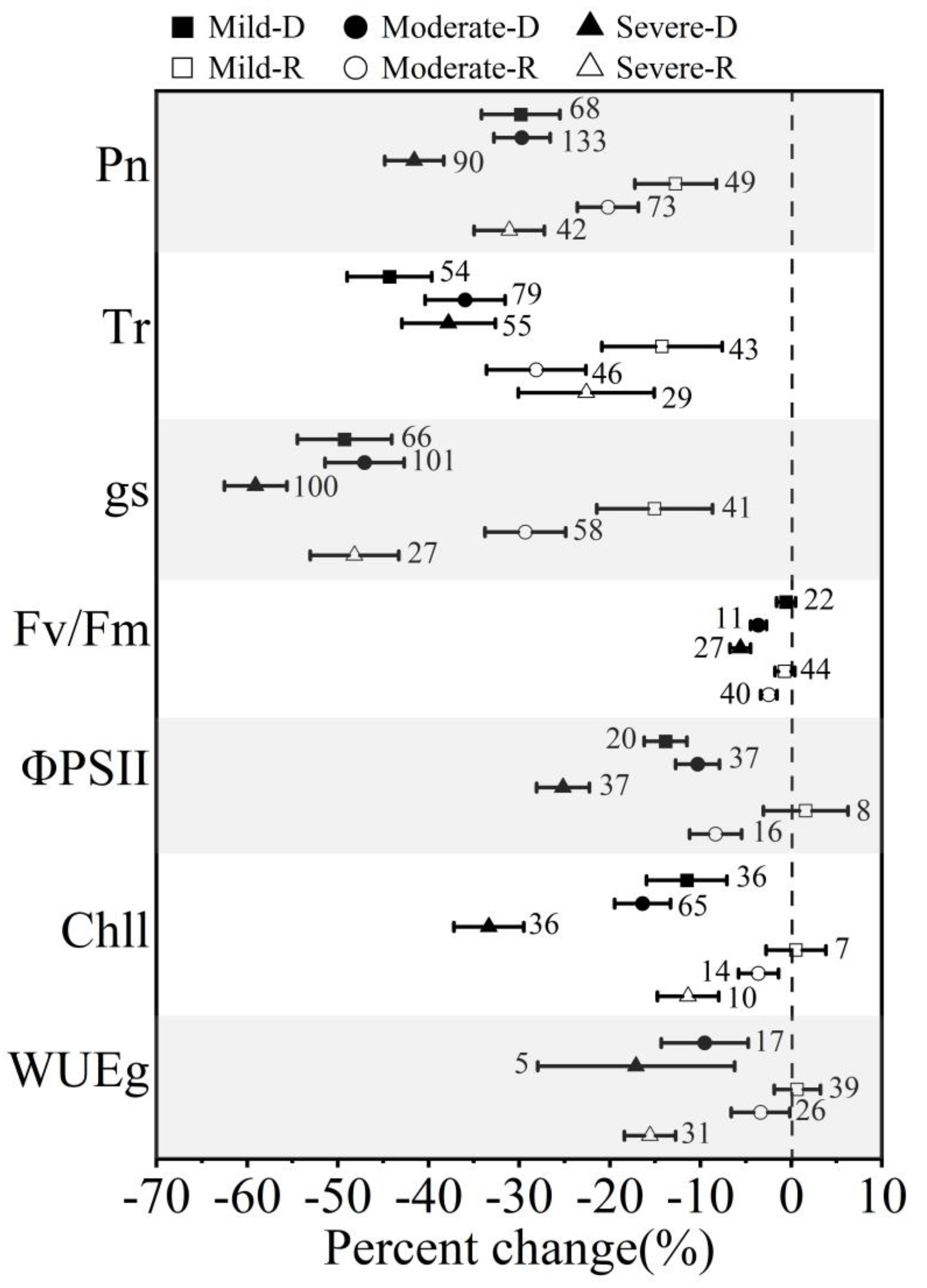
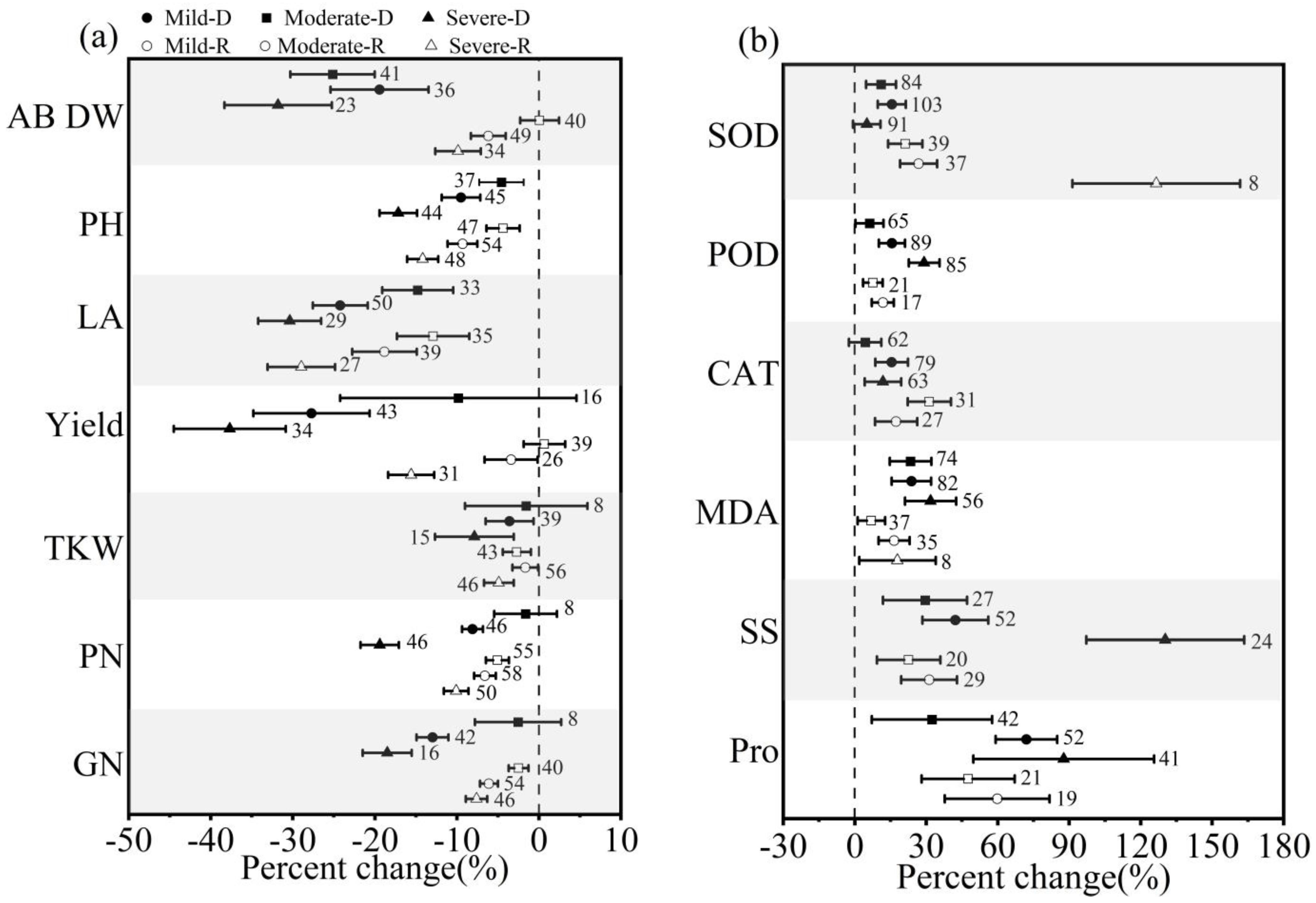
3.2. Effect of Stress Intensities on Winter Wheat in Response to Drought and Postdrought Rewatering
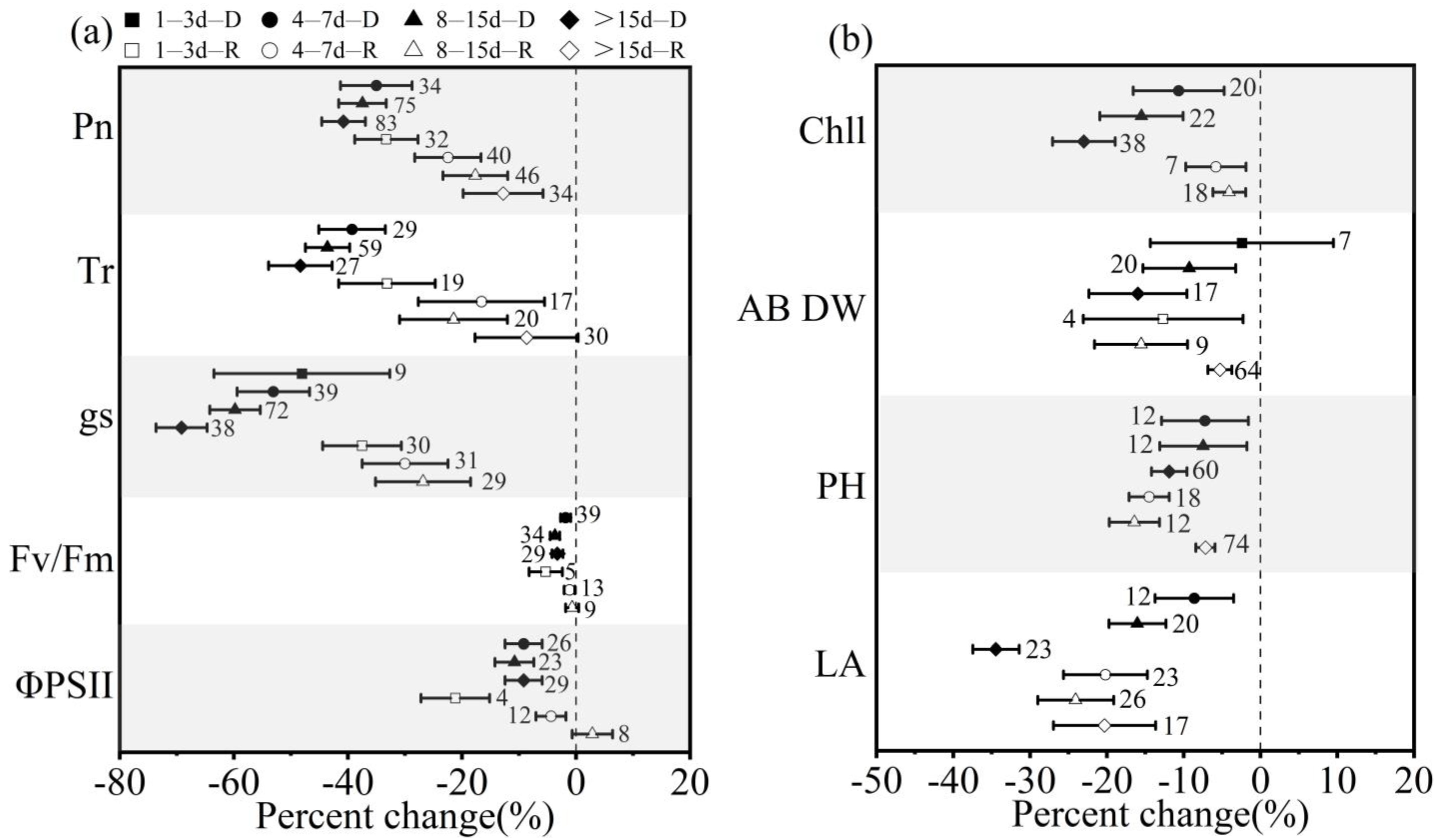
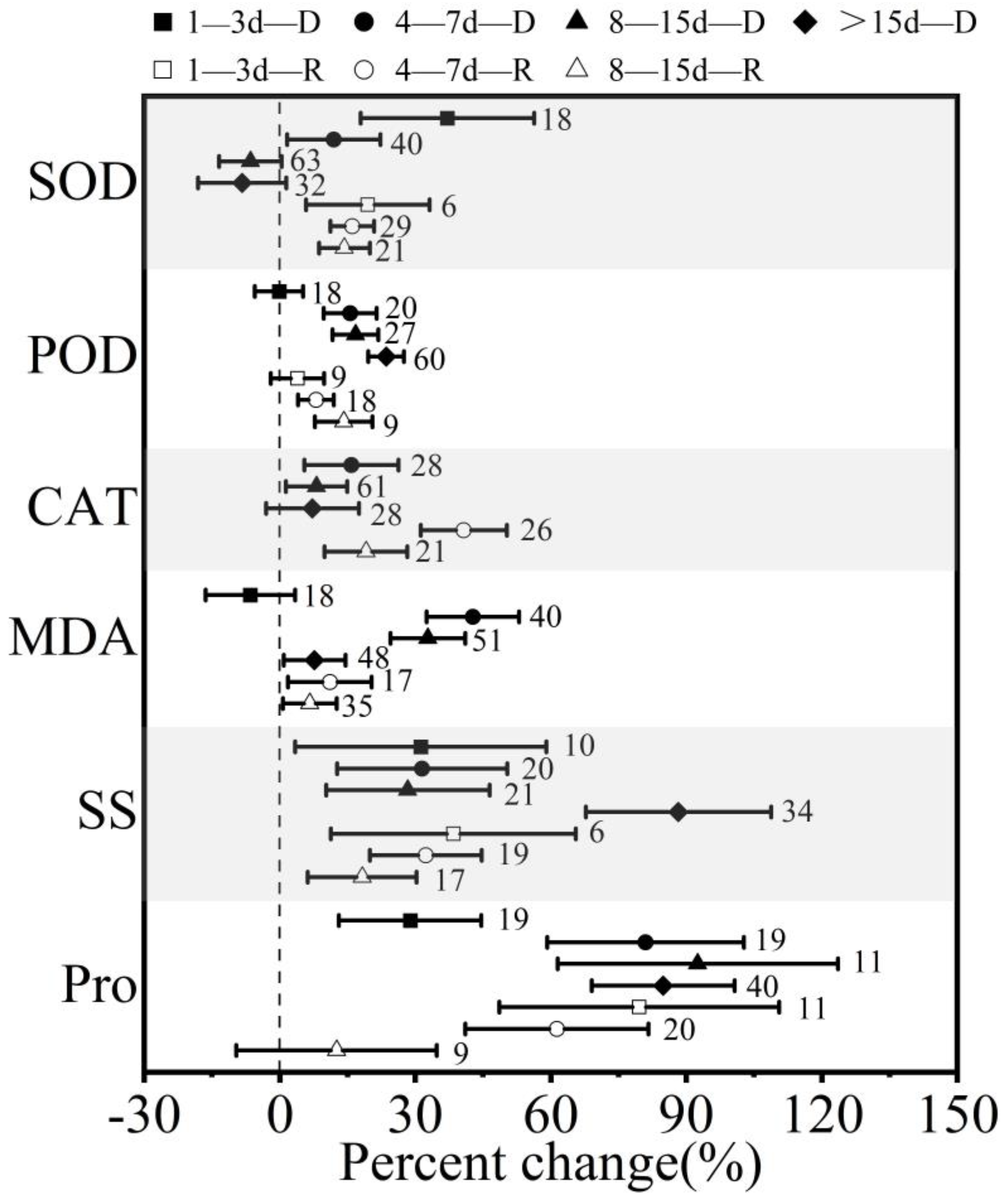
3.3. Effect of Drought (Rewatering) Treatment Durations on Winter Wheat in Response to Drought and Postdrought Rewatering
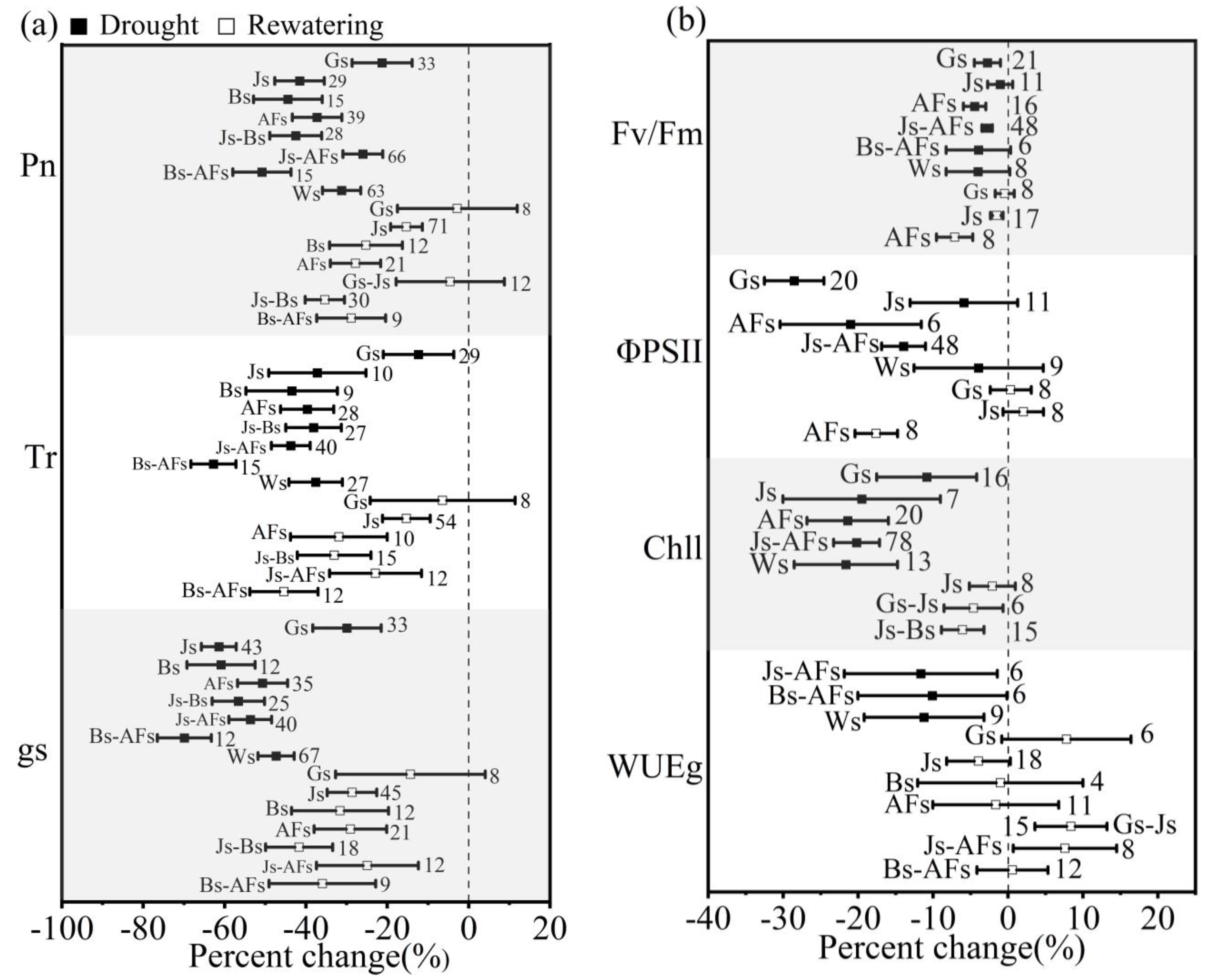
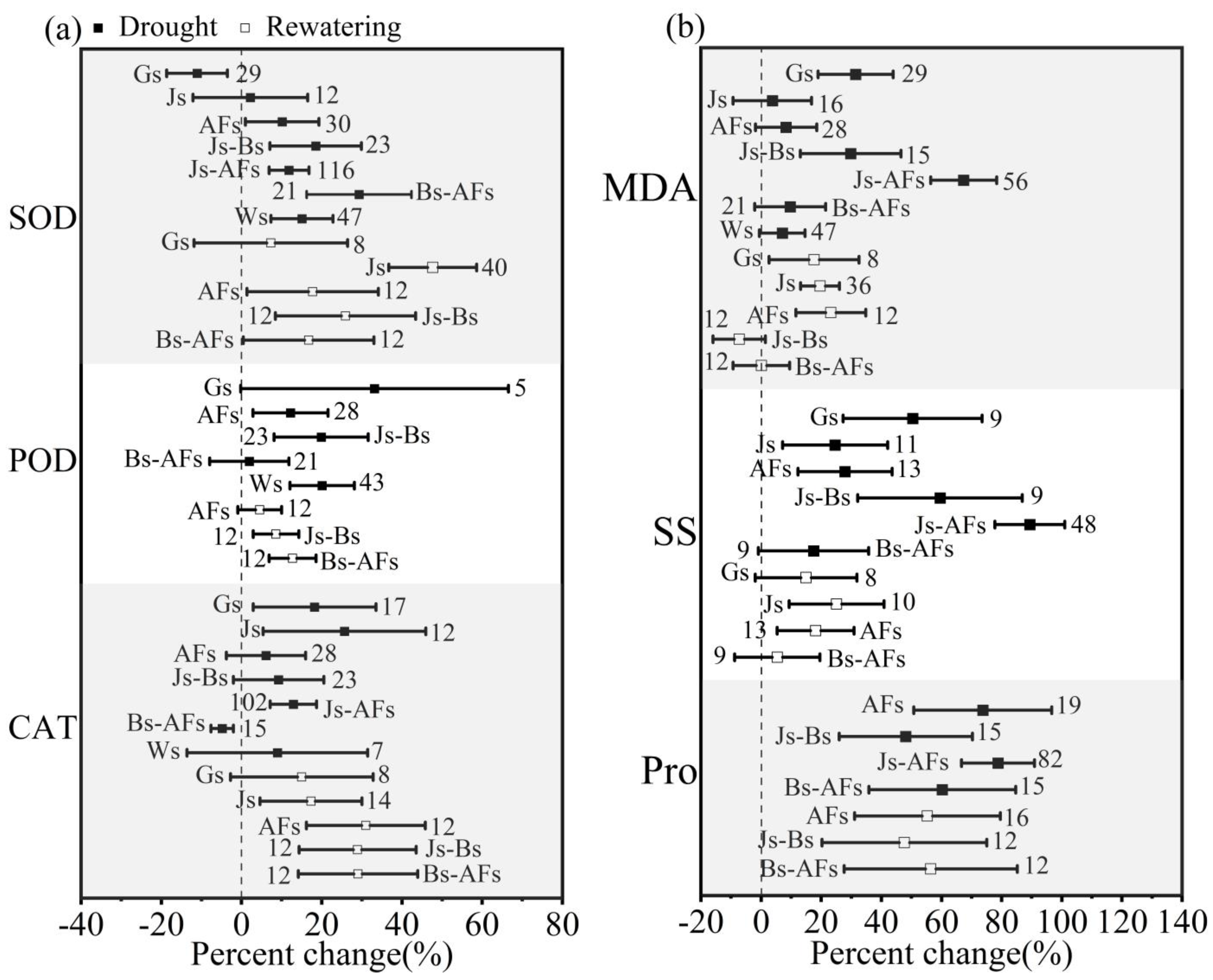
3.4. Effect of Growth Stages on Winter Wheat in Response to Drought and Postdrought Rewatering
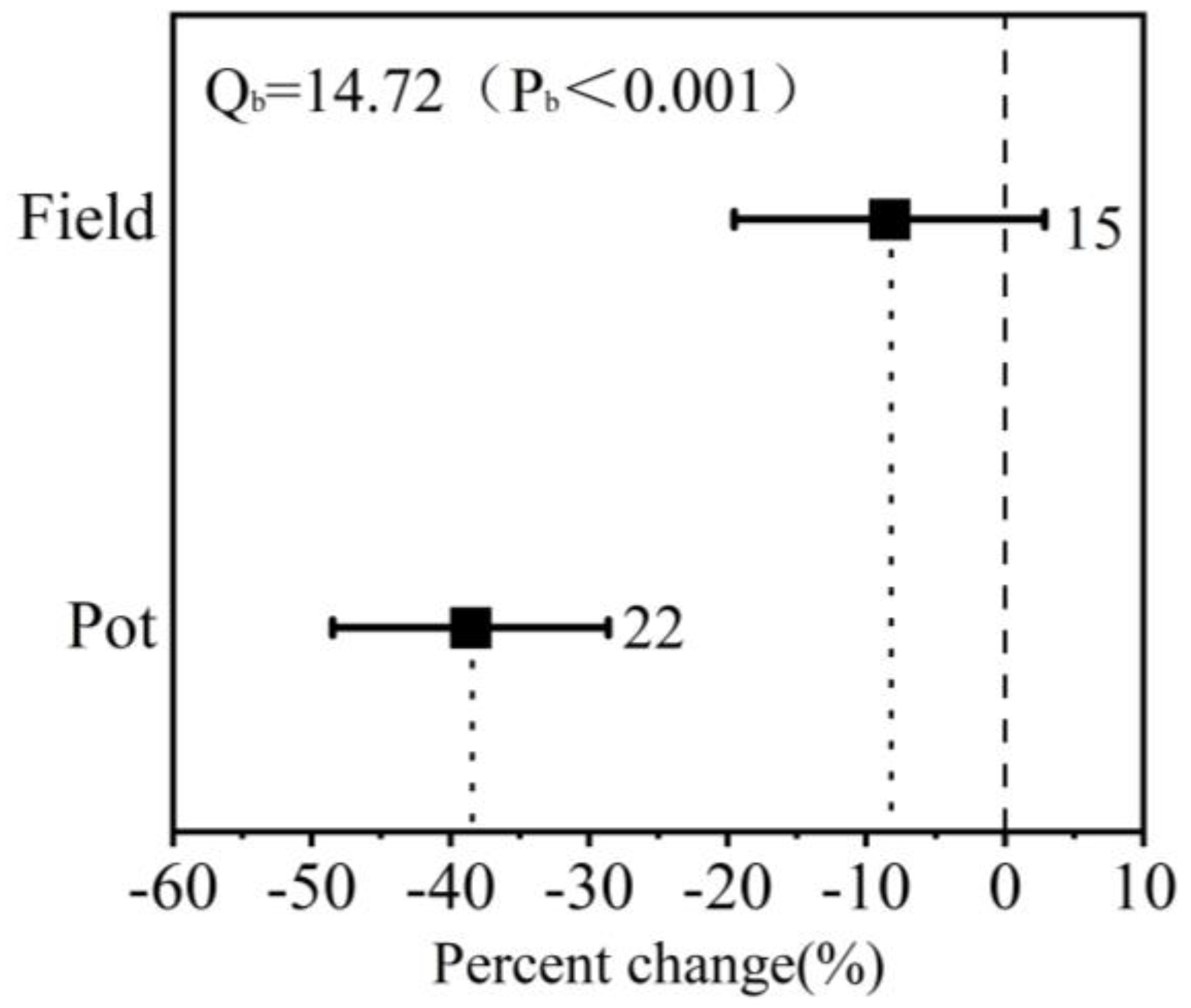
3.5. Effect of Planting Methods on Winter Wheat in Response to Drought and Postdrought Rewatering
3.6. Effect of Experimental Areas on Winter Wheat in Response to Drought and Postdrought Rewatering
4. Discussion
4.1. How Winter Wheat Respond to Drought Stress and Postdrought Rewatering
4.2. The Impact of Stress Intensities and Durations of Drought (Rewatering) on Winter Wheat in Response to Drought Stress and Postdrought Rewatering
4.3. The Impact of Growth Stages on Winter Wheat in Response to Drought Stress and Postdrought Rewatering
4.4. The Impact of Planting Methods on Winter Wheat in Response to Drought Stress and Postdrought Rewatering
4.5. The Impact of Experimental Areas on Winter Wheat in Response to Drought Stress and Postdrought Rewatering
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Han, J.; Zhang, Z.; Cao, J.; Luo, Y.; Zhang, L.; Li, Z.; Zhang, J. Prediction of winter wheat yield based on multi-source data and machine learning in China. Remote Sens. 2020, 12, 236. [Google Scholar] [CrossRef]
- Li, J.Y.; Yao, D.X.; Guo, X.L.; Wang, Z.M.; Zhang, Y.H. Effects of post-anthesis drought on photosynthesis, antioxidant properties, and C4 photosynthetic enzyme activity in winter wheat. Chin. J. Ecol. 2023, 42, 49–57. [Google Scholar]
- Lou, R.T.; Li, D.X.; Li, Y.B.; Bian, Z.P.; Zhu, Y.N. Effect of pre-anthesis drought hardening on post-anthesis physiological characteristics, yield and WUE in winter wheat. Phyton-Int. J. Exp. Bot. 2021, 90, 245–257. [Google Scholar] [CrossRef]
- Zlatev, Z.; Lidon, F.C. An overview on drought induced changes in plant growth, water relations and photosynthesis. Emir. J. Food Agric. 2012, 24, 57–72. [Google Scholar]
- Pei, D.; Sun, Z.S.; Chen, S.L.; Zhang, X.Y.; Zhang, Z.H. Effects of water stress on physiological and agronomic features of winter wheat. Trans. CSAE 2006, 22, 68–72. [Google Scholar]
- Shanmugam, S.; Kjaer, K.H.; Ottosen, C.O.; Rosenqvist, E.; Kumari Sharma, D. The alleviating effect of elevated CO2 on heat stress susceptibility of two wheat (Triticum aestivum L.) cultivars. J. Agron. Crop. Sci. 2013, 199, 340–350. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Van, B.F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 2006, 28, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.H.; Merope, T.M.; Zhang, Y.L.; Zhang, W.F. Combining gas exchange and chlorophyll a fluorescence measurements to analyze the photosynthetic activity of drip-irrigated cotton under different soil water deficits. J. Integr. Agric. 2016, 15, 1256–1266. [Google Scholar] [CrossRef]
- Tombesi, S.; Frioni, T.; Poni, S.; Palliotti, A. Effect of water stress ‘memory’ on plant behavior during subsequent drought stress. Environ. Exp. Bot. 2018, 150, 106–114. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Z.; Li, Y.; Feng, W.; Tian, W. Flag leaf chlorophyll fluorescence characteristics and its response to drought stress in different cultivars of winter wheat. J. Triticeae Crops 2015, 35, 699–706. [Google Scholar]
- Meng, L.L.; Song, J.F.; Wen, J.; Zhang, J.; Wei, J.H. Effects of drought stress on fluorescence characteristics of photosystem II in leaves of Plectranthus scutellarioides. Photosynthetica 2016, 54, 414–421. [Google Scholar] [CrossRef]
- Huang, M.; Wu, J.Z.; Li, Y.J.; Yao, Y.Q.; Zhang, C.J.; Cai, D.X.; Jin, K. Effects of different tillage management on production and yield of winter wheat in dryland. Trans. CSAE 2009, 25, 50–54. [Google Scholar]
- Zhang, Y.P.; Zhang, Y.H.; Wang, Z.M.; Wang, Z.J. Characteristics of canopy structure and contributions of non-leaf organs to yield in winter wheat under different irrigated conditions. Field Crops Res. 2011, 123, 187–195. [Google Scholar] [CrossRef]
- Wang, W.S.; Wang, C.; Pan, D.Y.; Zhang, Y.K.; Luo, B.; Ji, J. Effects of drought stress on photosynthesis and chlorophyll fluorescence images of soybean (Glycine max) seedlings. Int. J. Agric. Biol. Eng. 2018, 11, 196–201. [Google Scholar] [CrossRef]
- Dabrowski, P.; Baczewska-Dabrowska, A.H.; Kalaji, H.M.; Goltsev, V.; Paunov, M.; Rapacz, M. Exploration of chlorophyll a fluorescence and plant gas exchange parameters as indicators of drought tolerance in perennial ryegrass. Sensors 2019, 19, 2736. [Google Scholar] [CrossRef] [PubMed]
- Faseela, P.; Sinisha, A.K.; BrestiČ, M.; Puthur, J.T. Chlorophyll a fluorescence parameters as indicators of a particular abiotic stress in rice. Photosynthetica 2019, 57, 108–115. [Google Scholar]
- Alves, A.A.C.; Setter, T.L. Response of cassava leaf area expansion to water deficit: Cell proliferation, cell expansion and delayed development. Ann. Bot. 2004, 94, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; Zhang, S.Q.; Cheng, M.; Jiang, H.; Zhang, X.Y.; Peng, C.H.; Lu, X.H.; Zhang, M.X.; Jin, J.X. Effect of drought on agronomic traits of rice and wheat: A Meta-analysis. Int. J. Environ. Res. Public Health 2018, 15, 839. [Google Scholar] [CrossRef]
- Leilah, A.A.; Al-khateeb, S.A. Statistical analysis of wheat yield under drought conditions. J. Arid Environ. 2005, 61, 483–496. [Google Scholar] [CrossRef]
- Zhang, L.X.; Li, S.X.; Zhang, H.; Liang, Z.S. Nitrogen rates and water stress effects on production, lipid peroxidation and antioxidative enzyme activities in two maize (Zea mays L.) genotypes. J. Agron. Crop Sci. 2007, 193, 387–397. [Google Scholar] [CrossRef]
- Nio, S.A.; Cawthray, G.R.; Wade, L.J.; Colmer, T.D. Pattern of solutes accumulated during leaf osmotic adjustment as related to duration of water deficit for wheat at the reproductive stage. Plant Physiol. Biochem. 2011, 49, 1126–1137. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Shan, C.Z.; Tang, J.X.; Hao, W.F. Effects of water stress on leaf physiological characteristics of Lomax 9133 seedlings. J. Jiangsu Agric. Sci. 2006, 22, 229–232. [Google Scholar]
- Lou, L.L.; Li, X.R.; Chen, J.X.; Li, Y.; Tang, Y.; Lv, J.Y. Photosynthetic and ascorbate-glutathione metabolism in the flag leaves as compared to spikes under drought stress of winter wheat (Triticum aestivum L.). PLoS ONE 2018, 13, e0194625. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Dai, T.B. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, 0232974. [Google Scholar] [CrossRef] [PubMed]
- Simova-Stoilova, L.; Demirevska, K.; Petrova, T.; Tsenov, N.; Feller, U. Antioxidative protection in wheat varieties under severe recoverable drought at seedling stage. Plant Soil. Environ. 2008, 54, 529–536. [Google Scholar] [CrossRef]
- Manaa, A.; Goussi, R.; Derbali, W.; Cantamessa, S.; Barbato, R. Photosynthetic performance of quinoa (Chenopodium quinoa Willd.) after exposure to a gradual drought stress followed by a recovery period. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2021, 1862, 148383. [Google Scholar] [CrossRef] [PubMed]
- Vanková, R.; Dobrá, J.; Storchová, H. Recovery from drought stress in tobacco: An active process associated with the reversal of senescence in some plant parts and the sacrifice of others. Plant Signal. Behav. 2012, 7, 19–21. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Mu, Q.; Cai, H.J.; Sun, S.K.; Wen, S.S.; Xu, J.T.; Dong, M.Q.; Saddique, Q. The physiological response of winter wheat under short-term drought conditions and the sensitivity of different indices to soil water changes. Agric. Water Manag. 2021, 243, 106475. [Google Scholar] [CrossRef]
- Siopongco, J.D.L.C.; Yamauchi, A.; Salekdeh, H.; Bennett, L.; Wade, L.J. Growth and water use response of doubled-haploid rice lines to drought and rewatering during the vegetative stage. Plant Prod. Sci. 2006, 9, 141–151. [Google Scholar] [CrossRef]
- Yao, N.; Song, L.B.; Liu, J.; Feng, H.; Wu, S.F.; He, J.Q. Effects of water stress at different growth stages on the development and yields of winter wheat in arid region. Sci. Agric. Sin. 2015, 48, 2379–2389. [Google Scholar]
- Izanloo, A.; Condon, A.G.; Langridge, P.; Tester, M.; Schnurbusch, T. Different mechanisms of adaptation to cyclic water stress in two South Australian bread wheat cultivars. J. Exp. Bot. 2008, 59, 3327–3346. [Google Scholar] [CrossRef]
- Gurevitch, J.; Hedges, L.V. Statistical issues in ecological Meta-analyses. Ecology 1999, 80, 1142–1149. [Google Scholar] [CrossRef]
- Philibert, A.; Loyce, C.; Makowski, D. Assessment of the quality of Meta-analysis in agronomy. Agric. Ecosyst. Environ. 2012, 148, 72–82. [Google Scholar] [CrossRef]
- Zhang, L.Q.; Wang, Y.L.; Lou, Z.X.; Hsu, L.F.; Chen, D.; Piao, R.Z.; Zhao, H.Y.; Cui, Z.J. Meta-analysis of factors affecting C-N fractions and yield of paddy soils by total straw return and N fertilizer application. Agronomy 2022, 12, 3168. [Google Scholar] [CrossRef]
- Li, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Spring freeze effect on wheat yield is modulated by winter temperature fluctuations: Evidence from Meta-analysis and simulating experiment. J. Agron. Crop Sci. 2015, 201, 288–300. [Google Scholar] [CrossRef]
- Li, P.F.; Ma, B.L.; Palta, J.A.; Ding, T.T.; Cheng, Z.G.; Lv, G.C.; Xiong, Y.C. Wheat breeding highlights drought tolerance while ignores the advantages of drought avoidance: A Meta-analysis. Eur. J. Agron. 2021, 122, 126–196. [Google Scholar] [CrossRef]
- Villegas, D.; Aparicio, N.; Blanco, R.; Royo, C. Biomass accumulation and main stem elongation of durum wheat grown under Mediterranean conditions. Ann. Bot. 2001, 88, 617–627. [Google Scholar] [CrossRef]
- Ruehr, N.K.; Gast, A.; Weber, C.; Daub, B.; Arneth, A. Water availability as dominant control of heat stress responses in two contrasting tree species. Tree Physiol. 2016, 36, 164–178. [Google Scholar] [CrossRef]
- Liu, H.N.; Wu, H.; Yao, S. Depth extraction of continuous data in Meta-analysis. Chin. J. Evid.-Based Med. 2017, 17, 117–121. [Google Scholar]
- Pan, Y.X.; Yang, R.; Qiu, J.X.; Wang, J.Y.; Wu, J.P. Forty-year spatio-temporal dynamics of agricultural climate suitability in China reveal shifted major crop production areas. Catena 2023, 226, 107073. [Google Scholar] [CrossRef]
- Rosenberg, M.S.; Adams, D.C.; Gurevitch, J. Metawin: Statistical software for Meta-analysis version 2. Q. Rev. Biol. 1998, 73, 126–128. [Google Scholar]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The Meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Rosenthal, R. The file drawer problem and tolerance for null results. Psychol. Bull. 1979, 86, 638–641. [Google Scholar] [CrossRef]
- Luo, Y.Q.; Hui, D.F.; Zhang, D.Q. Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: A Meta-analysis. Ecology 2006, 87, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, H.R.; Sutton, A.J.; Borenstein, M. Publication Bias in Meta-Analysis, 1st ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006; pp. 1–7. [Google Scholar]
- Chandrasekaran, M.; Boughattas, S.; Hu, S.J.; Oh, S.H.; Sa, T. A Meta-analysis of arbuscular mycorrhizal effects on plants grown under salt stress. Mycorrhiza 2014, 24, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Li, X.R.; Tang, Y.; Zhou, C.J.; Zhang, L.X.; Lv, J.Y. A wheat WRKY transcription factor TaWRKY46 enhances tolerance to osmotic stress in transgenic Arabidopsis plants. Int. J. Mol. Sci. 2020, 21, 1321. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.H.; Bonnett, D.G.; Rebetzke, G.J. A 192bp allele at the Xgwm261 locus is not always associated with the Rht8 dwarfing gene in wheat (Triticum aestivum L.). Euphytica 2007, 157, 209–214. [Google Scholar] [CrossRef]
- Torrecillas, A.; Guillaume, C.; Alarcón, J.J.; Ruiz-Sánchez, M.C. Water relations of two tomato species under water-stress and recovery. Plant Sci. 1995, 105, 169–176. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, F. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J. Effects of water stress on photosystem II photochemistry and its thermostability in wheat plants. J. Exp. Bot. 1999, 50, 1199–1206. [Google Scholar] [CrossRef]
- Subrahmanyam, D.; Subash, N.; Haris, A.; Sikka, A.K. Influence of water stress on leaf photosynthetic characteristics in wheat cultivars differing in their susceptibility to drought. Photosynthetica 2006, 44, 125–129. [Google Scholar] [CrossRef]
- Abid, M.; Tian, Z.W.; Ata-UI-Karim, S.T.; Wang, F.; Liu, Y.; Zahoor, R.; Jiang, D.; Dai, T.B. Adaptation to and recovery from drought stress at vegetative stages in wheat (Triticum aestivum) cultivars. Funct. Plant Biol. 2016, 43, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.J.; Wang, R.R.; Jiang, G.Y.; Che, Z.Q.; Chen, T.P.; Hu, J.J.; Guo, R.G.; Peng, T.T.; Lei, Q.Q. Response of photosynthetic physiology and yield of spring wheat to drought-rewatering. J. Soil. Water Conserv. 2023, 37, 1–11. [Google Scholar]
- Ru, C.; Hu, X.T.; Chen, D.Y.; Wang, W.; Zhen, J.B. Photosynthetic, antioxidant activities, and osmoregulatory responses in winter wheat differ during the stress and recovery periods under heat, drought, and combined stress. Plant Sci. 2023, 327, 111557. [Google Scholar] [CrossRef]
- Selote, D.S.; Bharti, S.; Khanna-Chopra, R. Drought acclimation reduces O2− accumulation and lipid peroxidation in wheat seedlings. Biochem. Biophys. Res. Commun. 2004, 314, 724–729. [Google Scholar] [CrossRef]
- Asada, K. The water–water cycle in chloroplast: Scavenging of active oxygen and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Liu, X.J.; Wang, X.D.; Liu, P.; Bao, X.Y.; Hou, X.Y.; Yang, M.M.; Zhen, W.C. Rehydration compensation of winter wheat is mediated by hormone metabolism and de-peroxidative activities under field conditions. Front. Plant Sci. 2022, 13, 823846. [Google Scholar] [CrossRef]
- Aliyeva, D.R.; Aydinli, L.M.; Pashayeva, A.N.; Zulfugarov, I.S.; Huseynova, I.M. Photosynthetic machinery and antioxidant status of wheat genotypes under drought stress followed by rewatering. Photosynthetica 2020, 5895, 1217–1225. [Google Scholar] [CrossRef]
- He, J.N.; Duan, Y.; Hua, D.P.; Fan, G.J.; Wang, L.; Liu, Y. DEXH box RNA helicase-mediated mitochondrial reactive oxygen species production in Arabidopsis mediates crosstalk between abscisic acid and auxin signaling. Plant Cell 2012, 24, 1815–1833. [Google Scholar] [CrossRef]
- Qiu, J.R.; Xiang, X.Y.; Wang, J.T.; Xu, W.X.; Chen, J.; Xiao, Y. MfPIF1 of resurrection plant Myrothamnus flabellifolia plays a positive regulatory role in responding to drought and salinity stresses in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 3011. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.; Wang, K.; Wang, J.; Shi, L.; Du, L.; Ye, X. Expression of Arabidopsis Ornithine Aminotransferase (AtOAT) encoded gene enhances multiple abiotic stress tolerances in wheat. Plant Cell Rep. 2021, 40, 1155–1170. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Zhu, L.; Song, A.P.; Wang, H.B.; Chen, S.M.; Jiang, J.F.; Chen, F.D. Chrysanthemum (Chrysanthemum morifolium) CmICE2 conferred freezing tolerance in Arabidopsis. Plant Physiol. Biochem. 2020, 146, 31–41. [Google Scholar] [CrossRef]
- Varshney, R.K.; Barmukh, R.; Roorkiwal, M.; Qi, Y.; Kholova, Y.; Tuberosa, R.; Reynolds, M.P.; Tardieu, F.; Siddique, K.H.M. Breeding custom-designed crops for improved drought adaptation. Adv. Genet. 2021, 2, e202100017. [Google Scholar] [CrossRef]
- Raza, A.; Mubarik, M.S.; Sharif, R.; Habib, M.; Jabeen, W.; Zhang, C.; Chen, H.; Chen, Z.H.; Siddique, K.H.M.; Zhuang, W. Developing drought-smart, ready-to-grow future crops. Plant Genome 2022, 16, e20279. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.Y.; Xue, X.P.; Yang, Z.Q.; Zhao, H.; Dong, Z.Q.; Xu, Y.; Zou, J.L. Compensation effects of rewatering at flowering stage on leaf state and yield structure of winter wheat under drought. Chin. J. Agrometeorol. 2020, 41, 253–262. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Schoppach, R.; Sadok, W. Differential sensitivities of transpiration to evaporative demand and soil water deficit among wheat elite cultivars indicate different strategies for drought tolerance. Environ. Exp. Bot. 2012, 84, 1–10. [Google Scholar] [CrossRef]
- Zhang, F.; Guo, J.K.; Yang, Y.L.; He, W.L.; Zhang, L.X. Changes in the pattern of antioxidant enzymes in wheat exposed to water deficit and rewatering. Acta. Physiol. Plant. 2004, 26, 345–352. [Google Scholar] [CrossRef]
- Guo, Z.; Ou, W.; Lu, S.; Zhong, Q. Differential responses of antioxidative system to chilling and drought in four rice cultivars differing in sensitivity. Plant Physiol. Biochem. 2006, 44, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Szablińska-Piernik, J.; Lesław, B.L. Polar metabolites profiling of wheat shoots (Triticum aestivum L.) under repeated short-term soil drought and rewatering. Int. J. Mol. Sci. 2023, 24, 8429. [Google Scholar] [CrossRef]
- Gu, Y.F.; Ding, S.Y.; Gao, Z.Y.; Xing, Q. Effects of drought and rehydration on the distribution pattern of photosynthetic products in winter wheat. Strateg. Study CAE 2012, 14, 59–64. [Google Scholar]
- Feng, Z.Z.; Kazuhiko, K.; Elizabetha, A. Impact of elevated ozone concentration on growth, physiology, and yield of wheat (Triticum aestivum L.): A Meta-analysis. Glob. Change Biol. 2008, 14, 2696–2708. [Google Scholar] [CrossRef]
- Hamblin, A.P. Root characteristics of some temperate legume species and varieties on deep, free-draining entisols. Aust. J. Agric. Res. 1985, 36, 63–72. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Category | Parameter Abbreviation | Description |
|---|---|---|
| Gas exchange parameters, chlorophyll content, chlorophyll fluorescence parameters and water use efficiency | Pn | Leaf net photosynthetic rate |
| gs | Stomatal conductance | |
| Tr | Transpiration | |
| Fv/Fm | Quantum efficiency of PSII photochemistry in dark-adapted state | |
| ΦPSII | The quantum yield of electron transport | |
| Chll | Total chlorophyll concentration in leaves | |
| WUEg | Water use efficiency for grain | |
| Plant architecture | PH | Plant height |
| LA | Leaf area | |
| Yield, yield components, and biomass components | Yield | Yield |
| TKW | Thousand kernel weight | |
| GN | Grain number per panicle | |
| PN | Panicle number per pot or m2 | |
| AB DW | Aboveground dry weight | |
| Biochemical components | SOD | Superoxide dismutase activities in leaves |
| POD | Peroxidase activities in leaves | |
| CAT | Catalase activities in leaves | |
| GR | glutathione reductase activities in leaves | |
| MDA | Malondialdehyde concentration in leaves | |
| SS | Soluble sugar concentration in leaves | |
| Pro | Proline concentration in leaves | |
| H2O2 | hydrogen peroxide concentration in leaves | |
| superoxide anion concentration in leaves |
| Abbreviation | Division Description |
|---|---|
| Gs drought | Drought occurring at the regreening stage |
| Js drought | Drought occurring at the jointing stage |
| Bs drought | Drought occurring at the booting stage |
| AFs drought | Drought occurring at the anthesis stage, the filling stage or from the anthesis stage to maturity |
| Ws drought | Drought during the complete growth cycle |
| Gs-Js drought | Drought occurring from the regreening stage to the jointing stage |
| Gs-Bs drought | Drought occurring from the regreening stage to the booting stage |
| Gs-AFs drought | Drought occurring from the regreening stage to the anthesis stage (or the filling stage, maturity) |
| Js-Bs drought | Drought occurring from the jointing stage to the booting stage |
| Js-AFs drought | Drought occurring from the jointing stage to the anthesis stage (or the filling stage, maturity) |
| Bs-AFs drought | Drought occurring from the booting stage to the anthesis stage (or the filling stage, maturity) |
| Categorical Variable | Treatment Condition | Treatment (between D and R) | Stress Intensity | Treatment Duration | Growth Stage | Planting Method | Experimental Area |
|---|---|---|---|---|---|---|---|
| Pn | D | 48.82 *** | 28.74 *** | 2.70 | 59.78 *** | 44.37 *** | 49.76 *** |
| R | 37.28 *** | 22.97 *** | 61.88 *** | - | 26.43 *** | ||
| Tr | D | 36.43 *** | 6.41 * | 4.77 | 100.02 *** | 7.86 ** | 4.75 |
| R | 9.90 ** | 14.88 ** | 45.07 *** | 0.04 | 38.51 *** | ||
| gs | D | 67.21 *** | 19.01 *** | 19.19 *** | 77.10 *** | 32.37 *** | 116.89 *** |
| R | 64.01 *** | 3.97 | 12.07 | 0.80 | 5.46 | ||
| Fv/Fm | D | 2.77 | 45.38 *** | 10.35 ** | 12.93 * | 1.67 | 7.62 * |
| R | 7.24 ** | 15.28 *** | 31.47 *** | 0.24 | 37.54 *** | ||
| ΦPSII | D | 22.11 *** | 61.53 *** | 2.83 | 59.57 *** | 3.65 | 20.65 *** |
| R | 16.97 *** | 84.12 *** | 148.56 *** | - | 154.54 *** | ||
| Chll | D | 35.25 *** | 59.14 *** | 13.11 ** | 11.88 * | 10.85 *** | 0.23 |
| R | 33.05 *** | 2.62 | 5.25 | 3.04 | 1.64 | ||
| WUEg | D | 8.30 ** | 4.79 | - | 0.8 | 21.46 *** | 0.82 |
| R | 72.44 *** | - | 276.16 *** | 0.56 | 18.92 *** | ||
| AB DW | D | 176.72 *** | 7.33 * | 5.72 | 111.96 *** | 28.46 *** | 0.07 |
| R | 31.84 *** | 15.20 *** | 21.99 ** | 0.62 | 29.00 *** | ||
| PH | D | 2.43 | 52.45 *** | 4.30 | 40.61 *** | - | 0.09 |
| R | 49.50 *** | 61.96 *** | 107.44 *** | 42.33 *** | 13.61 ** | ||
| LA | D | 4.53 * | 29.44 *** | 105.78 *** | 70.97 *** | 0.97 | 2.50 |
| R | 28.16 *** | 1.76 | 22.88 *** | - | 1.37 | ||
| Yield | D | 17.99 *** | 13.91 *** | - | 24.73 *** | 5.21 * | 10.71 ** |
| R | 625.02 *** | - | 1102.15 *** | 48.50 *** | 53.01 *** | ||
| TKW | D | 0.65 | 3.29 | - | 20.63 *** | 0.01 | 4.22 * |
| R | 7.31 * | - | 277.84 *** | 34.77 *** | 5.51 | ||
| PN | D | 14.97 *** | 99.71 *** | - | 35.56 *** | 51.87 *** | 28.73 *** |
| R | 24.32 *** | - | 1143.39 *** | 154.20 *** | 0.63 | ||
| GN | D | 55.75 *** | 37.13 *** | - | 32.93 *** | 64.40 *** | 50.56 *** |
| R | 35.97 *** | - | 857.74 *** | 121.99 *** | 47.73 *** | ||
| SOD | D | 27.38 *** | 6.12 * | 29.72 *** | 39.39 *** | 5.17 * | 82.10 *** |
| R | 66.64 *** | 1.79 | 21.33 *** | - | 130.52 *** | ||
| POD | D | 2.89 | 26.64 *** | 48.41 *** | 58.77 *** | 21.15 *** | 80.60 *** |
| R | 2.12 | 7.09 * | 14.84 ** | - | - | ||
| CAT | D | 9.51 ** | 5.03 | 1.84 | 10.46 | 8.77 ** | 10.23 ** |
| R | 4.69 * | 10.63 ** | 4.19 | - | 3.60 | ||
| GR | D | 2.07 | - | - | - | - | - |
| R | - | - | - | - | - | ||
| MDA | D | 9.40 ** | 1.72 | 64.98 *** | 117.40 *** | 5.07 * | 43.00 *** |
| R | 5.50 | 0.73 | 34.98 *** | - | 4.17 | ||
| SS | D | 11.67 *** | 37.09 *** | 25.33 *** | 98.63 *** | - | 22.17 *** |
| R | 0.91 | 3.77 | 61.32 *** | - | 7.48 ** | ||
| Pro | D | 4.06 * | 31.85 *** | 27.04 *** | 5.97 | 2.02 | - |
| R | 0.65 | 14.64 *** | 0.24 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Yang, Z. Effects of Drought Stress and Postdrought Rewatering on Winter Wheat: A Meta-Analysis. Agronomy 2024, 14, 298. https://doi.org/10.3390/agronomy14020298
Wu H, Yang Z. Effects of Drought Stress and Postdrought Rewatering on Winter Wheat: A Meta-Analysis. Agronomy. 2024; 14(2):298. https://doi.org/10.3390/agronomy14020298
Chicago/Turabian StyleWu, Huizhen, and Zaiqiang Yang. 2024. "Effects of Drought Stress and Postdrought Rewatering on Winter Wheat: A Meta-Analysis" Agronomy 14, no. 2: 298. https://doi.org/10.3390/agronomy14020298
APA StyleWu, H., & Yang, Z. (2024). Effects of Drought Stress and Postdrought Rewatering on Winter Wheat: A Meta-Analysis. Agronomy, 14(2), 298. https://doi.org/10.3390/agronomy14020298