
Analysis of the Photosynthetic Parameters, Grain Yield, and Quality of Different Winter Wheat Varieties over a Two-Year Period

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Field Trial
2.2. Measurement of Agro-Morphological Traits
2.3. Measurement of Grain and Flour Quality Traits
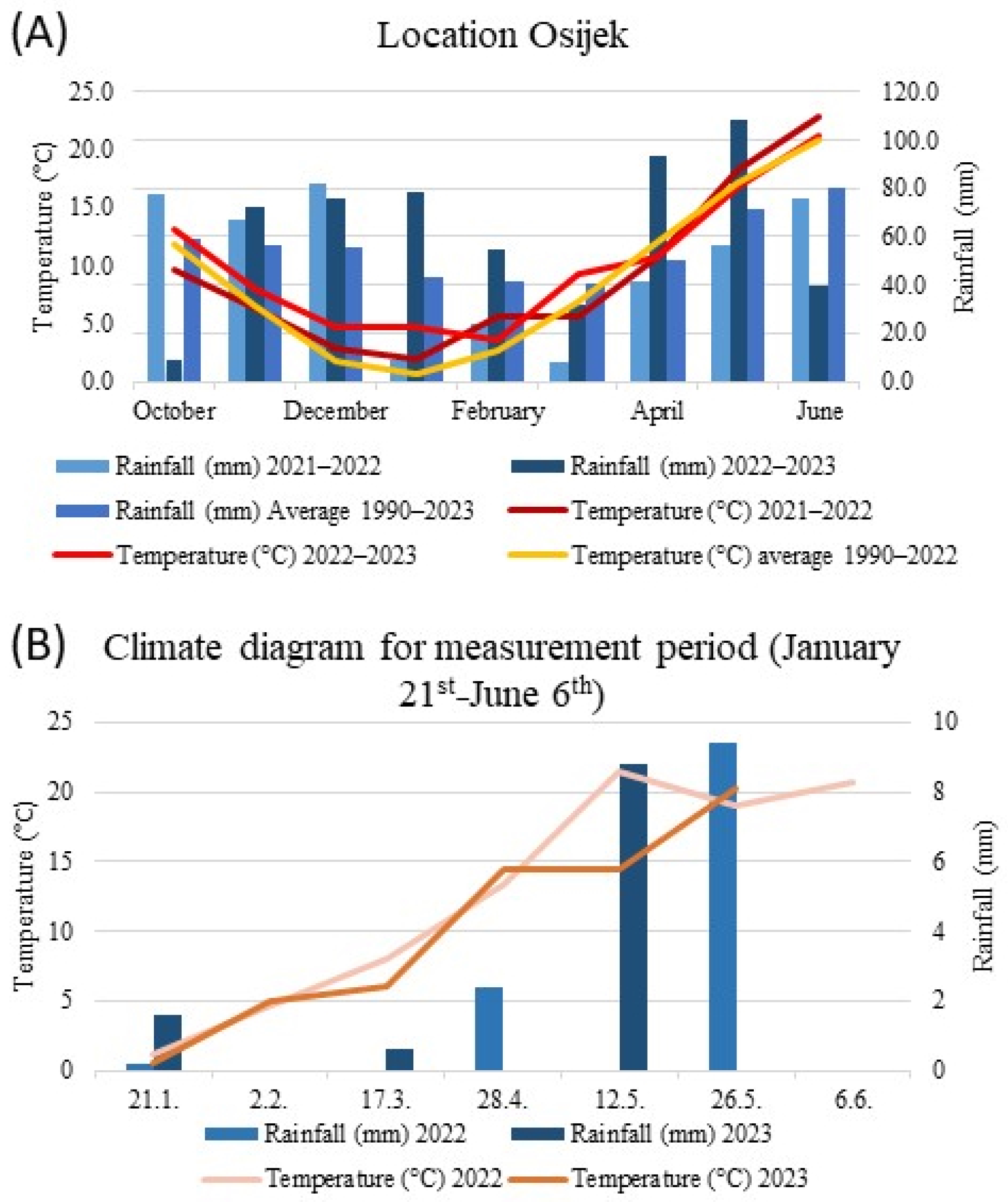
2.4. Chlorophyll a Fluorescence Measurement
2.5. Statistical Analysis
3. Results
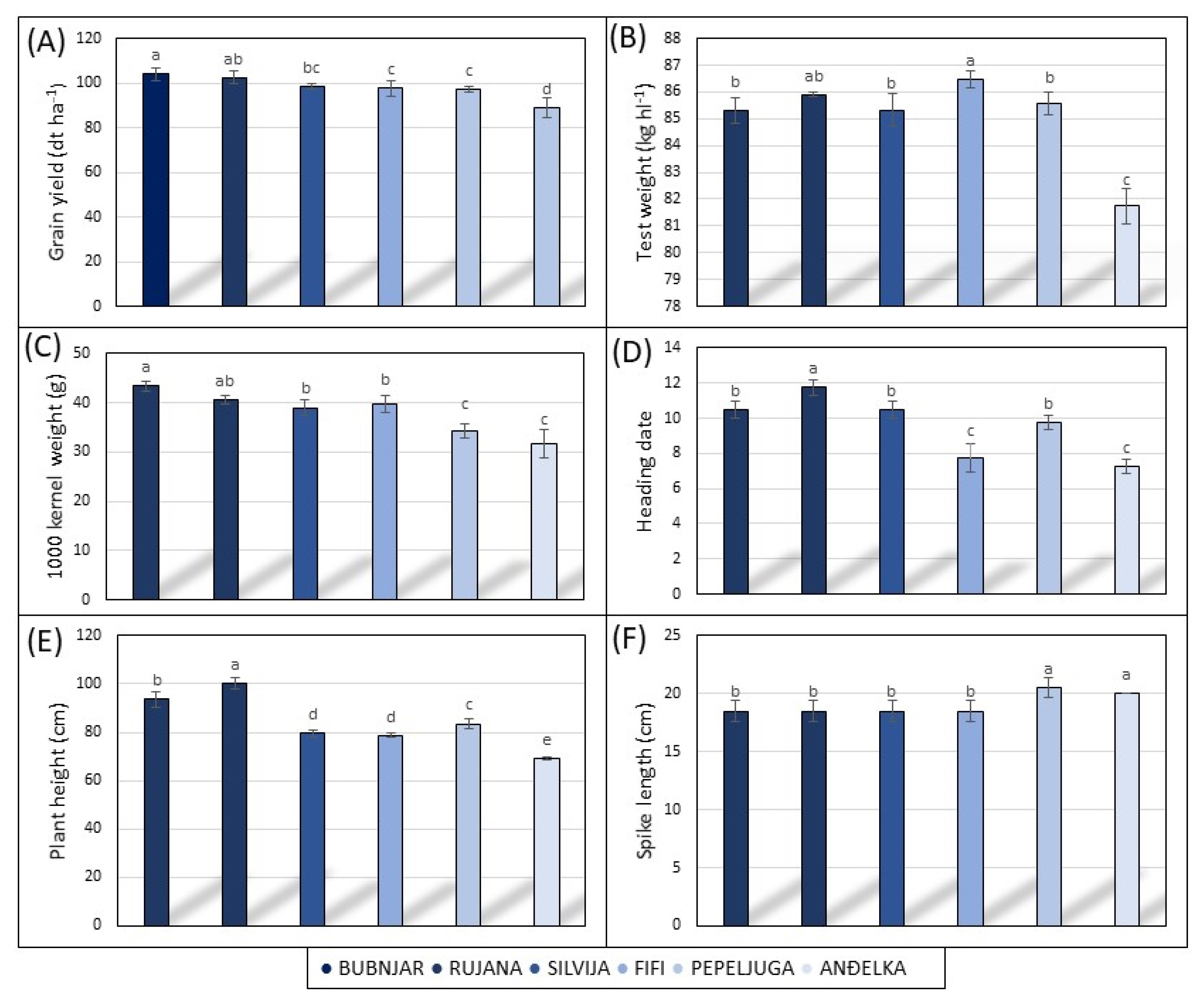
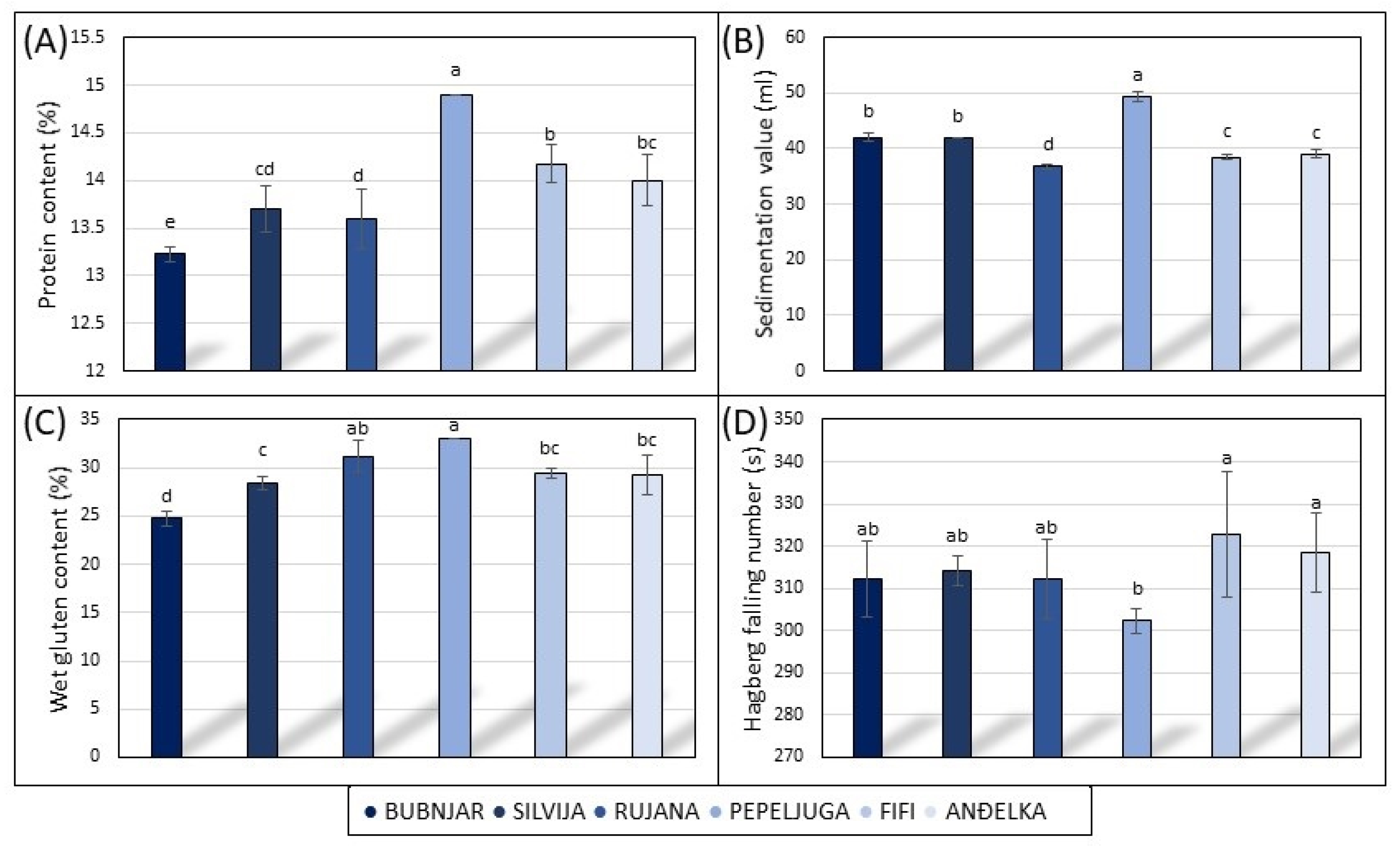
3.1. Agro-Morphological and Technologically Quality Traits in 2021/2022 Vegetative Season
3.2. Agro-Morphological and Technological Quality Traits in 2022/2023 Vegetative Season
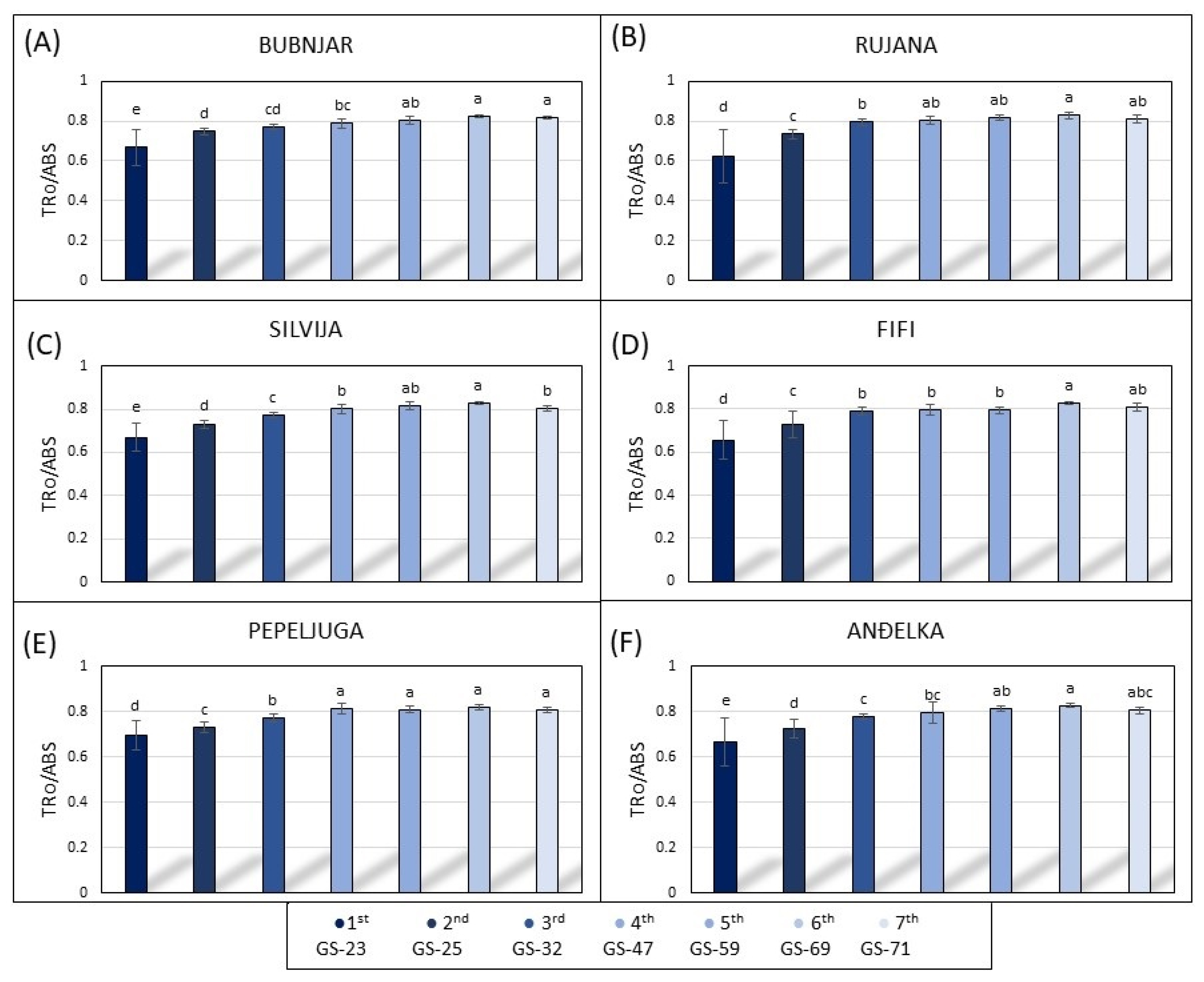
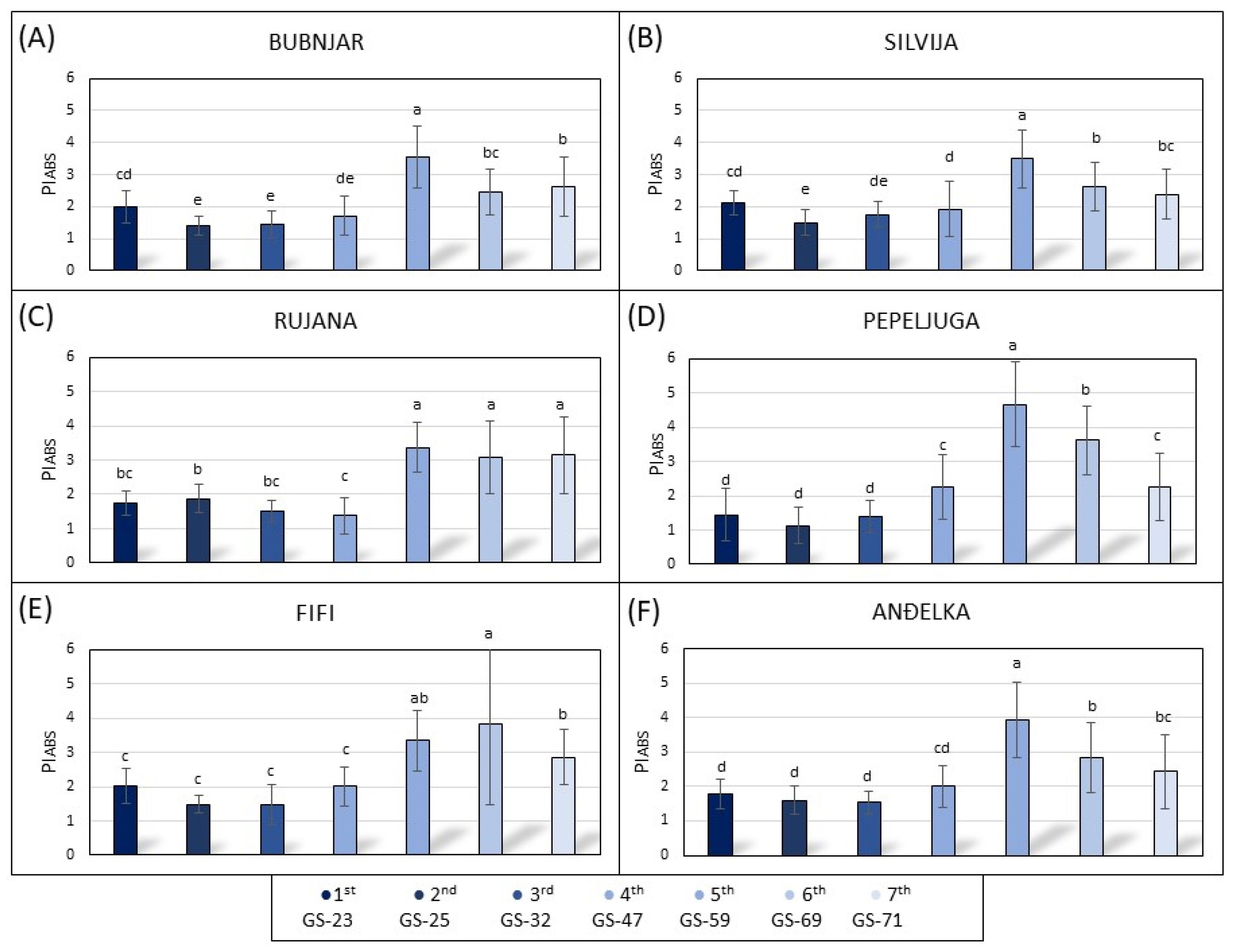
3.3. Photosynthetic Parameters of the Leaves during the 2021/2022 Growing Season
3.4. Photosynthetic Parameters of the Leaves during the 2022/2023 Growing Season
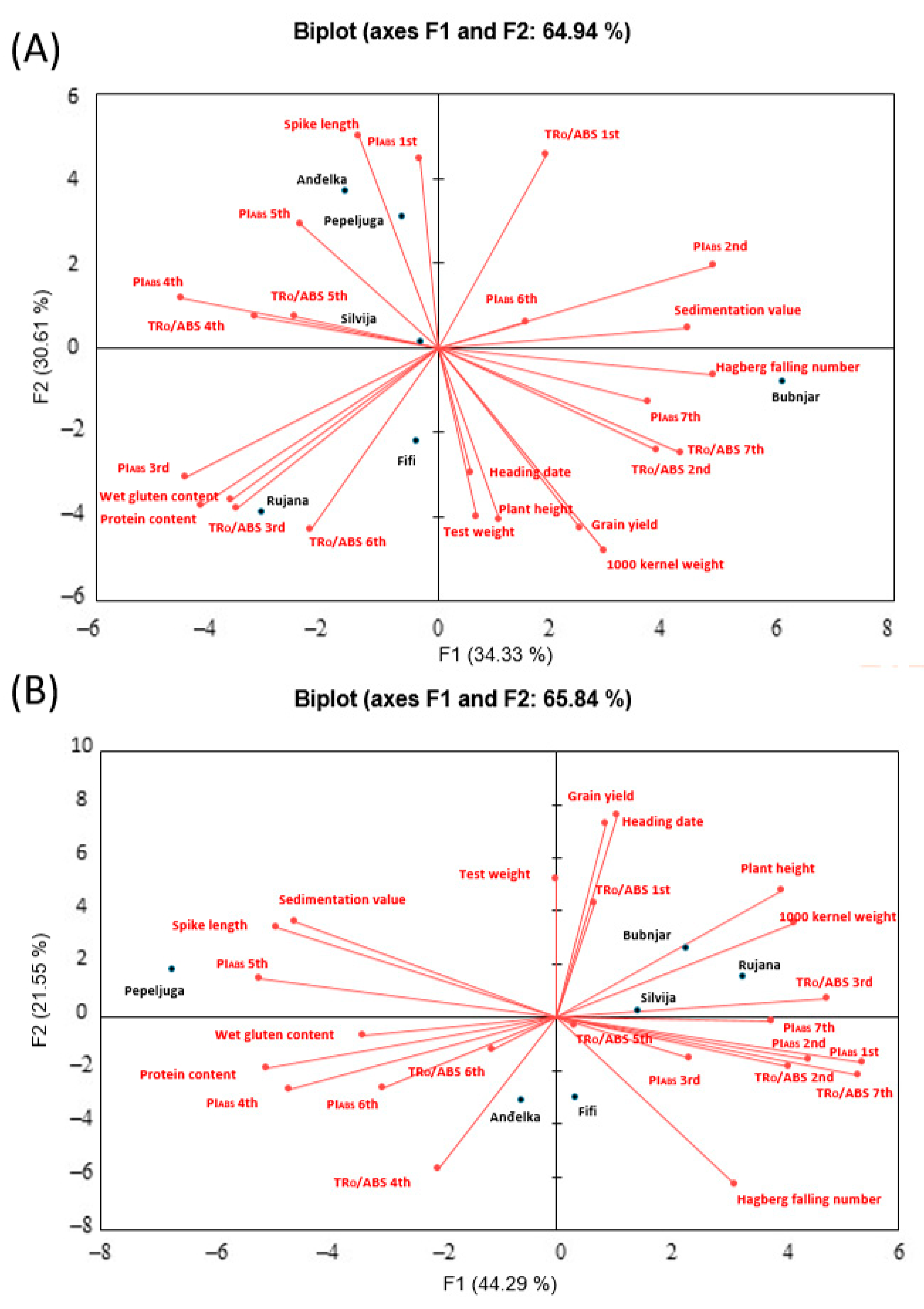
3.5. Correlation between Traits
4. Discussion
4.1. Vegetative Season 2021/2022
4.2. Vegetative Season 2022/2023
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Obembe, O.S.; Hendricks, N.P.; Tack, J. Decreased wheat production in the USA from climate change driven by yield losses rather than crop abandonment. PLoS ONE 2021, 16, e0252067. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Expanding wheat yields with expansin. New Phytol. 2021, 230, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.; Bayer, P.E.; Edwards, D. Climate change and the need for agricultural adaptation. Curr. Opin. Plant Biol. 2020, 56, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Mereu, V.; Gallo, A.; Trabucco, A.; Carboni, G.; Spano, D. Modeling high-resolution climate change impacts on wheat and maize in Italy. Clim. Risk Manag. 2021, 33, 100339. [Google Scholar] [CrossRef]
- Croatian Bureau of Statistics. Available online: https://podaci.dzs.hr/2022/hr/29384 (accessed on 23 May 2023).
- Onyemaobi, O.; Sangma, H.; Garg, G.; Wallace, X.; Kleven, S.; Suwanchaikasem, P.; Roessner, U.; Dolferus, R. Reproductive stage drought tolerance in wheat: Importance of stomatal conductance and plant growth regulators. Genes 2021, 12, 1742. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop Sci. 2011, 5, 764–777. [Google Scholar]
- Nouri-Ganbalani, A.; Nouri-Ganbalani, G.; Hassanpanah, D. Effects of drought stress condition on the yield and yield components of advanced wheat genotypes in Ardabil, Iran. J. Food Agric. Environ. 2009, 77, 228–234. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global Synthesis of Drought Effects on Maize and Wheat Production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef]
- Liu, J. Risk assessment of possible impacts of climate change and irrigation on wheat yield and quality with a modified CERES-Wheat model. J. Water Clim. Chang. 2021, 12, 2444–2459. [Google Scholar] [CrossRef]
- Tatar, O.; Bruck, H.; Asch, F. Photosynthesis and remobilization of dry matter in wheat as affected by progressive drought stress at stem elongation stage. J. Agron. Crop Sci. 2016, 202, 292–299. [Google Scholar] [CrossRef]
- Vukovic, R.; Stolfa Camagajevac, I.; Vukovic, A.; Sunic, K.; Begovic, L.; Mlinaric, S.; Sekulic, R.; Sabo, N.; Spanic, V. Physiological, biochemical and molecular response of different winter wheat varieties under drought stress at germination and seedling growth stage. Antioxidants 2022, 11, 693. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Ntuli, T.M. Drought and desiccation-tolerance and sensitivity in plants. In Botany; IntechOpen: London, UK, 2012; pp. 29–60. [Google Scholar]
- Brodersen, C.; Jansen, S.; Choat, B.; Rico, C.; Pittermann, J. Cavitation resistance in seedless vascular plants: The structure and function of interconduit pit membranes. Plant Physiol. 2014, 165, 895–904. [Google Scholar] [CrossRef]
- Senapati, N.; Stratonovitch, P.; Paul, M.J.; Semenov, M.A. Drought tolerance during reproductive development is important for increasing wheat yield potential under climate change in Europe. J. Exp. Bot. 2019, 70, 2549–2560. [Google Scholar] [CrossRef]
- Khadka, K.; Earl, H.J.; Raizada, M.N.; Navabi, A. A physio-morphological trait-based approach for breeding drought tolerant wheat. Front. Plant Sci. 2020, 11, 715. [Google Scholar] [CrossRef]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of climate change on agriculture and its mitigation strategies. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Launay, M.; Caubel, J.; Bourgeois, G.; Huard, F.; Cortazar-Atauri, I.G.; Bancal, M.O.; Brisson, N. Climatic indicators for crop infection risk: Application to climate change impacts on five major foliar diseases in Northern France. Agric. Ecosyst. Environ. 2014, 197, 147–158. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Hong, J.; Zhang, L.; Cooper, O.R.; Schultz, M.G.; Xu, X.; Wang, T.; Gao, M.; Zhao, Y.; Zhang, Y. Severe surface ozone pollution in China: A global perspective. Environ. Sci. Technol. 2018, 5, 487–494. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, H.; Wang, S.; Zheng, Y.; Mai, B.; Zhang, X. Assessment of photosynthesis and yield loss of winter wheat under ground-level ozone exposure. Environ. Technol. Innov. 2023, 29, 103013. [Google Scholar] [CrossRef]
- Parry, M.A.J.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef]
- Yan, Y.; Hou, P.; Duan, F.; Niu, L.; Dai, T.; Wang, K.; Zhao, M.; Li, S.; Zhou, W. Improving photosynthesis to increase grain yield potential: An analysis of maize hybrids released in different years in China. Photosynth. Res. 2021, 150, 295–311. [Google Scholar] [CrossRef]
- Long, S.P.; Marshall-Colon, A.; Zhu, X.G. Meeting the global food demand of the future by engineering crop photosynthesis and yield potential. Cell 2015, 161, 56–66. [Google Scholar] [CrossRef]
- Kubar, M.S.; Alshallash, K.S.; Asghar, M.A.; Feng, M.; Raza, A.; Wang, C.; Saleem, K.; Ullah, A.; Yang, W.; Kubar, K.A.; et al. Improving Winter Wheat Photosynthesis, Nitrogen Use Efficiency, and Yield by Optimizing Nitrogen Fertilization. Life 2022, 12, 1478. [Google Scholar] [CrossRef]
- Katanic, Z.; Mlinaric, S.; Katanic, N.; Cosic, J.; Spanic, V. Photosynthetic efficiency in flag leaves and ears of winter wheat during Fusarium head blight infection. Agronomy 2021, 11, 2415. [Google Scholar] [CrossRef]
- Spanic, V.; Sunic, K.; Duvnjak, J.; Yin-Gang, H.; Katanic, Z. Chlorophyll a fluorescence during flag leaf senescence of field-grown winter wheat plants under drought conditions. Ann. Appl. Biol. 2023, 183, 80–92. [Google Scholar] [CrossRef]
- Spanic, V.; Mlinaric, S.; Zdunic, Z.; Katanic, Z. Field study of the effects of two different environmental conditions on wheat productivity and Chlorophyl fluorescence induction (OJIP) parameters. Agriculture 2021, 11, 1154. [Google Scholar] [CrossRef]
- Spanic, V.; Sarcevic, H. Evaluation of effective system for tracing FHB resistance in wheat: An editorial commentary. Agronomy 2023, 13, 2116. [Google Scholar] [CrossRef]
- Duvnjak, J.; Loncaric, A.; Brkljacic, L.; Samec, D.; Sarcevic, H.; Salopek-Sondi, B.; Spanic, V. Morpho-physiological and hormonal response of winter wheat varieties to drought stress at stem elongation and anthesis stages. Plants 2023, 12, 418. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Spanic, V.; Cosic, J.; Zdunic, Z.; Drezner, G. Characterization of agronomical and quality traits of winter wheat (Triticum aestivum L.) for Fusarium head blight pressure in different environments. Agronomy 2021, 11, 213. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescent transient as a tool to characterise and screen photosynthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; CRC Press: New York, NY, USA, 2000; pp. 445–483. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2004; Volume 19. [Google Scholar]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.V.V.; Bheemanahalli, R.; Jagadish, S.V.K. Field crops and the fear of heat stress-opportunities, challenges and future directions. Field Crops Res. 2017, 200, 114–121. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Maarten van Ginkel, M.; Ribaut, J.M. Avenues for genetic modification of radiation use efficiency in wheat. J. Exp. Bot. 2000, 51, 459–473. [Google Scholar] [CrossRef]
- Slattery, R.A.; Ainsworth, E.A.; Ort, D.R. A meta-analysis of responses of canopy photosynthetic conversion efficiency to environmental factors reveals major causes of yield gap. J. Exp. Bot. 2013, 64, 3723–3733. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, W.Z.; Lin, W.; Han, X.Y.; Zhou, L.; Wang, Y.P. Grain Yield and WUE Responses to Different Soil Water Storage before sowing and water supplies during growing period for winter wheat in the loess tableland. Sci. Agric. Sin. 2017, 50, 3549–3560. [Google Scholar]
- Eitzinger, J.; Štastná, M.; Žalud, Z.; Dubrovskỳ, M.A. Simulation study of the effect of soil water balance and water stress on winter wheat production under different climate change scenarios. Agric. Water Manag. 2003, 61, 195–217. [Google Scholar] [CrossRef]
- Wu, J.D.; Li, J.C.; Wei, F.Z.; Wang, C.Y.; Zhang, Y.; Sun, G. Effects of nitrogen spraying on the post-anthesis stage of winter wheat under waterlogging stress. Acta Physiol. Plant. 2014, 36, 207–216. [Google Scholar] [CrossRef]
- Moore, F.C.; Lobell, D.B. The fingerprint of climate trends on European crop yields. Proc. Natl. Acad. Sci. USA 2015, 112, 2670–2675. [Google Scholar] [CrossRef]
- Sharma, R.C.; Tiwary, A.K.; Ortiz-Ferrara, G. Reduction in kernel weight as a potential indirect selection criterion for wheat grain yield under terminal heat stress. Plant Breed. 2008, 127, 241–248. [Google Scholar] [CrossRef]
- Mohammadi, M.; Sharifi, P.; Karimizadeh, R.; Shefazadeh, M.K. Relationships between grain yield and yield components in bread wheat under different water availability (dryland and supplemental irrigation conditions). Not. Bot. Horti Agrobot. Cluj Napoca 2012, 40, 195–200. [Google Scholar] [CrossRef]
- Nofouzi, F. Evaluation of seed yield of durum wheat (Triticum durum) under drought stress and determining correlation amongsome yield components using path coefficient analysis. Cuad. Investig. 2018, 10, 179–183. [Google Scholar]
- Makino, A. Photosynthesis, grain yield, and nitrogen utilization in rice and wheat. Plant Physiol. 2011, 155, 125–129. [Google Scholar] [CrossRef]
- Tilley, M.S.; Heiniger, R.W.; Crozier, C.R. Tiller initiation and its effects on yield and yield components in winter wheat. Agron. J. 2019, 111, 1323–1332. [Google Scholar] [CrossRef]
- Xie, Q.; Mayes, S.; Sparkes, D.L. Optimizing tiller production and survival for grain yield improvement in a bread wheat × spelt mapping population. Ann. Bot. 2016, 117, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Wang, Y.; Tang, H.; Sui, N.; Zhang, X.; Wang, F. Genetic, hormonal, and environmental control of tillering in wheat. Crop J. 2021, 9, 986–991. [Google Scholar] [CrossRef]
- Souza, T.; Magalhães, P.C.; Mauro de Castro, E.; Pereira de Albuquerque, P.R.; Marabesi, M.A. The influence of ABA on water relation, photosynthesis parameters, and chlorophyll fluorescence under drought conditions in two maize hybrids with contrasting drought resistance. Acta Physiol. Plant 2013, 35, 515–527. [Google Scholar] [CrossRef]
- Shunqing, A.; Gengshan, L.; Anhong, G. Consumption of available soil water stored at planting by winter wheat. Agric. Water Manag. 2003, 63, 99–107. [Google Scholar] [CrossRef]
- Izanloo, A.; Condon, A.G.; Langridge, P.; Tester, M.; Schnurbusch, T. Different mechanisms of adaptation to cyclic water stress in two South Australian bread wheat cultivars. J. Exp. Bot. 2008, 59, 3327–3346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zheng, B.; He, Y. Improving grain yield via promotion of kernel weight in high yielding winter wheat genotypes. Biology 2021, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Mahdy, R.E.; Ashehri, D.; Alatawi, H.A.; Al-Amrah, H.; Mahdy, E.E. Direct and indirect selection for grain yield and grain weight in late generations of bread wheat under drought stress and normal irrigation environments. Plants 2022, 11, 1604. [Google Scholar] [CrossRef]
- Spanic, V.; Drezner, G.; Dvojkovic, K.; Horvat, D. Traits of 25 Winter wheat varieties grown in Croatia in the last 100 years. Agron. Glas. 2016, 78, 3–16. [Google Scholar]
- Dubey, R.; Pathak, H.; Singh, S.; Chakrabarti, B.; Fagodiya, R.K. Impact of sowing dates on terminal heat tolerance of different wheat (Triticum aestivum L.) cultivars. Natl. Acad. Sci. Lett. 2019, 42, 445–449. [Google Scholar] [CrossRef]
- Zhou, H.; Riche, A.B.; Hawkesford, M.J.; Whalley, W.R.; Atkinson, B.S.; Sturrock, C.J.; Mooney, S.J. Determination of wheat spike and spikelet architecture and grain traits using X-ray Computed Tomography imaging. Plant Methods 2021, 17, 26. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, R.; Kumar, N.; Sihag, M.K.; Ray, A. Isolation, characterization of wheat gluten and its regeneration properties. J. Food Sci. Technol. 2015, 52, 5930–5937. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ghatak, A.; Bazargani, M.M.; Bajaj, P.; Varshney, R.K.; Chaturvedi, P.; Jiang, D.; Weckwerth, W. Spatial distribution of proteins and metabolites in developing wheat grain and their differential regulatory response during the grain filling process. Plant J. 2021, 107, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Turksoy, S.; Onar, D.A. Rapid detection of whole wheat gluten quality by a novel chemometric technique-glutopeak. Foods 2022, 11, 1927. [Google Scholar] [CrossRef]
- Laidig, F.; Piepho, H.P.; Rentel, D.; Drobek, T.; Meyer, U.; Huesken, A. Breeding progress, environmental variation and correlation of winter wheat yield and quality traits in German official variety trials and on-farm during 1983–2014. Theor. Appl. Genet. 2017, 130, 223–245. [Google Scholar] [CrossRef]
- Iizumi, T.; Ramankutty, N. How do weather and climate influence cropping area and intensity. Glob. Food Secur. 2015, 4, 46–50. [Google Scholar] [CrossRef]
- Nault, L.R. Arthropod transmission of plant viruses-a new synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Nancarrow, N.; Aftab, M.; Hollaway, G.; Rodoni, B.; Trębicki, P. Yield losses caused by barley yellow dwarf virus-PAV infection in wheat and barley: A three-year field study in South-Eastern Australia. Microorganisms 2021, 9, 645. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Fones, H.; Gurr, S. The impact of Septoria tritici blotch disease on wheat: An EU perspective. Fungal Genet. Biol. 2015, 79, 3–7. [Google Scholar] [CrossRef]
- Berraies, S.; Salah Gharbi, M.; Rezgui, S.; Yahyaoui, A. Estimating grain yield losses caused by septoria leaf blotch on durum wheat in Tunisia. Chil. J. Agric. Res. 2014, 74, 432–437. [Google Scholar] [CrossRef]
- Persic, V.; Bozinovic, I.; Varnica, I.; Babic, J.; Spanic, V. Impact of fusarium head blight on wheat flour quality: Examination of protease activity, technological quality and rheological properties. Agronomy 2023, 13, 662. [Google Scholar] [CrossRef]
- Spanic, V.; Zdunic, Z.; Drezner, G.; Viljevac Vuletic, M. Differences in physiological traits at the initial stage of Fusarium head blight infection in wheat. Biol. Plant. 2020, 64, 185–192. [Google Scholar] [CrossRef]
- Hlaváčová, M.; Klem, K.; Rapantová, B.; Novotná, K.; Urban, O.; Hlavinka, P.; Smutná, P.; Horáková, V.; Škarpa, P.; Pohanková, E.; et al. Interactive effects of high temperature and drought stress during stem elongation, anthesis and early grain filling on the yield formation and photosynthesis of winter wheat. Field Crops Res. 2018, 221, 182–195. [Google Scholar] [CrossRef]
- Gambin, B.L.; Borras, L.; Otegui, M.E. Source-sink relations and kernel weight differences in maize temperate hybrids. Field Crops Res. 2006, 95, 316–326. [Google Scholar] [CrossRef]
- Mu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Qin, J.; Cao, W. Long-term low radiation decreases leaf photosynthesis, photochemical efficiency and grain yield in winter wheat. J. Agron. Crop Sci. 2010, 196, 38–47. [Google Scholar] [CrossRef]
- Jablonskytė-Raščė, D.; Maikštėnienė, S.; Mankevičienė, A. Evaluation of productivity and quality of common wheat (Triticum aestivum L.) and spelt (Triticum spelta L.) in relation to nutrition conditions. Zemdirb. Agric. 2013, 100, 45–56. [Google Scholar] [CrossRef]
- Saint Pierre, C.; Peterson, J.; Ross, A.; Ohm, J.B.; Verhoeven, M.; Larson, M.; Hoefer, B. White wheat grain quality changes with genotype, nitrogen fertilization, and water stress. Agron. J. 2008, 100, 414–420. [Google Scholar] [CrossRef]
- Bogard, M.; Allard, V.; Martre, P.; Hezmenez, E.; Snape, J.W.; Orford, S.; Griffiths, S.; Gaju, O.; Foulkes, J.; Le Gouis, J. Identifying wheat genomic regions for improving grain protein concentration independently of grain yield using multiple inter-related populations. Mol. Breed. 2012, 31, 587–599. [Google Scholar] [CrossRef]
- Geyer, M.; Mohler, V.; Hartl, L. Genetics of the inverse relationship between grain yield and grain protein content in common wheat. Plants 2022, 11, 2146. [Google Scholar] [CrossRef] [PubMed]
- Munier-Jolain, N.; Salon, C. Are the carbon costs of seed production related to the quantitative and qualitative performance? An appraisal for legumes and other crops. Plant Cell Environ. 2005, 28, 1388–1395. [Google Scholar] [CrossRef]
- Acreche, M.M.; Slafer, G.A. Variation of grain nitrogen content in relation with grain yield in old and modern Spanish wheats grown under a wide range of agronomic conditions in a mediterranean region. J. Agric. Sci. 2009, 147, 657–667. [Google Scholar] [CrossRef]
- Tanaka, H.; Gorafi, Y.S.A.; Fujita, M.; Sasaki, H.; Tahir, I.S.A.; Tsujimoto, H. Expression of seed storage proteins responsible for maintaining kernel traits and wheat flour quality in common wheat under heat stress conditions. Breed. Sci. 2021, 71, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, I.F. Interaction between drought and chronic high temperature during kernel filling in wheat in a controlled environment. Ann. Bot. 2002, 90, 469–476. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Quantity |
|---|---|
| pHKCl | 6.25 |
| Humus | 2.00–2.20% |
| P2O5 | 39.70 mg 100 g−1 |
| H2O | 37.70mg 100 g−1 |
| Agro-Technical Measure | Agent and Concentrations of Active Compounds |
|---|---|
| 2021/2022 | |
| Pre-sowing seed treatment in 2021 | Maxim Extra 050 FS (fludioxonil 25 g L−1; difenoconazole 25 g L−1) |
| Weed control in November 2021 | Sharpen 330 EC (pendimethalin 330 g L−1) |
| Pest control in April 2022 | Vantex 60 CS (gamma-cyhalothrin 60 g L−1) |
| Weed control in May 2022 | Lodin (fluroxypyr-metil 295.5 g L−1) + Tribe 75 WG (tribenuron 750 g kg−1) |
| Pest control in May 2022 | Cythrin Max (cypermethrin 500 g L−1) |
| 2022/2023 | |
| Pre-sowing seed treatment in 2022 | Maxim Extra 050 FS (fludioxonil 25 g L−1; difenoconazole 25 g L−1) |
| Weed control in February 2023 | Alister New (diflufenican 120 g L−1; jodosulfuron 7.5 g L−1; mesosulfuron 9 g L−1) |
| Weed control in May 2023 | Sekator (amidosulfuron 100 g L−1; jodosulfuron metil 25 g L−1) + Tribe 75 WG (tribenuron 750 g kg−1) |
| Pest control in April 2023 | Cyclone (lambda-cyhalothrin 50 g L−1) |
| Pest control in May 2023 | Cyclone (lambda-cyhalothrin 50 g L−1) |
| Source of Variability | DF | Mean Square | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Grain Yield | Test Weight | 1000 Kenel Weight | Heading Date | Plant Height | Spike Length | Protein Content | Sedimentation Value | Wet Gluten Content | Hagberg Falling Number | ||
| Variety (V) | 5 | 712.8 *** | 64.0 *** | 206.0 *** | 40.6 *** | 1204.1 *** | 13.6 *** | 1.1 *** | 67.1 *** | 36.3 *** | 829.0 ** |
| Year (Y) | 1 | 24,797.5 *** | 314.7 *** | 278.4 *** | 80.1 *** | 475.7 *** | 1160.3 *** | 0.1 | 125.5 *** | 1.2 | 1210.0 ** |
| V × Y | 5 | 186.2 *** | 15.4 *** | 6.58 | 3.0 *** | 63.5 *** | 3.43 ** | 0.8 *** | 54.0 *** | 15.6 *** | 464.0 ** |
| Error | 36 | 9.1 | 0.6 | 2.76 | 0.4 | 4.7 | 1.9 | 0.1 | 2.5 | 1.4 | 183 |
| Source of Variability | DF | Mean Square | |
|---|---|---|---|
| TRO/ABS | PIABS | ||
| Variety (V) | 5 | 0.01 | 1.72 ** |
| Year (Y) | 1 | 0.03 | 32.93 *** |
| Measurement point (M) | 6 | 0.32 *** | 206.87 *** |
| V × Y | 5 | 0.01 | 0.82 |
| V × M | 30 | 0.01 | 2.16 *** |
| Y × M | 6 | 0.13 *** | 65.73 *** |
| V × Y × M | 30 | 0.08 | 3.29 *** |
| Error | 1596 | 0.01 | 0.05 |
| Variety | Barley Yellow Dwarf Virus (%) | Yellow Rust (%) |
|---|---|---|
| Bubnjar | 3.8 | 0 |
| Silvija | 4.8 | 3 |
| Rujana | 2.8 | 5 |
| Pepeljuga | 4.0 | 10 |
| Fifi | 3.2 | 20 |
| Anđelka | 2.3 | 90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duvnjak, J.; Katanic, Z.; Sarcevic, H.; Spanic, V. Analysis of the Photosynthetic Parameters, Grain Yield, and Quality of Different Winter Wheat Varieties over a Two-Year Period. Agronomy 2024, 14, 478. https://doi.org/10.3390/agronomy14030478
Duvnjak J, Katanic Z, Sarcevic H, Spanic V. Analysis of the Photosynthetic Parameters, Grain Yield, and Quality of Different Winter Wheat Varieties over a Two-Year Period. Agronomy. 2024; 14(3):478. https://doi.org/10.3390/agronomy14030478
Chicago/Turabian StyleDuvnjak, Jurica, Zorana Katanic, Hrvoje Sarcevic, and Valentina Spanic. 2024. "Analysis of the Photosynthetic Parameters, Grain Yield, and Quality of Different Winter Wheat Varieties over a Two-Year Period" Agronomy 14, no. 3: 478. https://doi.org/10.3390/agronomy14030478