
Effect of Biochar on Vermicompost Production: Chemical, Biochemical, and Biological Properties
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of Biochar
2.2. Experimental Setup
2.3. Treatments
2.4. Chemical Analysis
2.5. Preparation of Samples for Enzymatic Assays
2.6. Enzymatic Activity
2.7. Microbial Activity
2.8. Microbial Respiration
2.9. Enzymatic Index
2.10. Statistical Analysis
3. Results
4. Discussion
4.1. E. fetida Population and Yield and Chemical Properties of Vermicompost
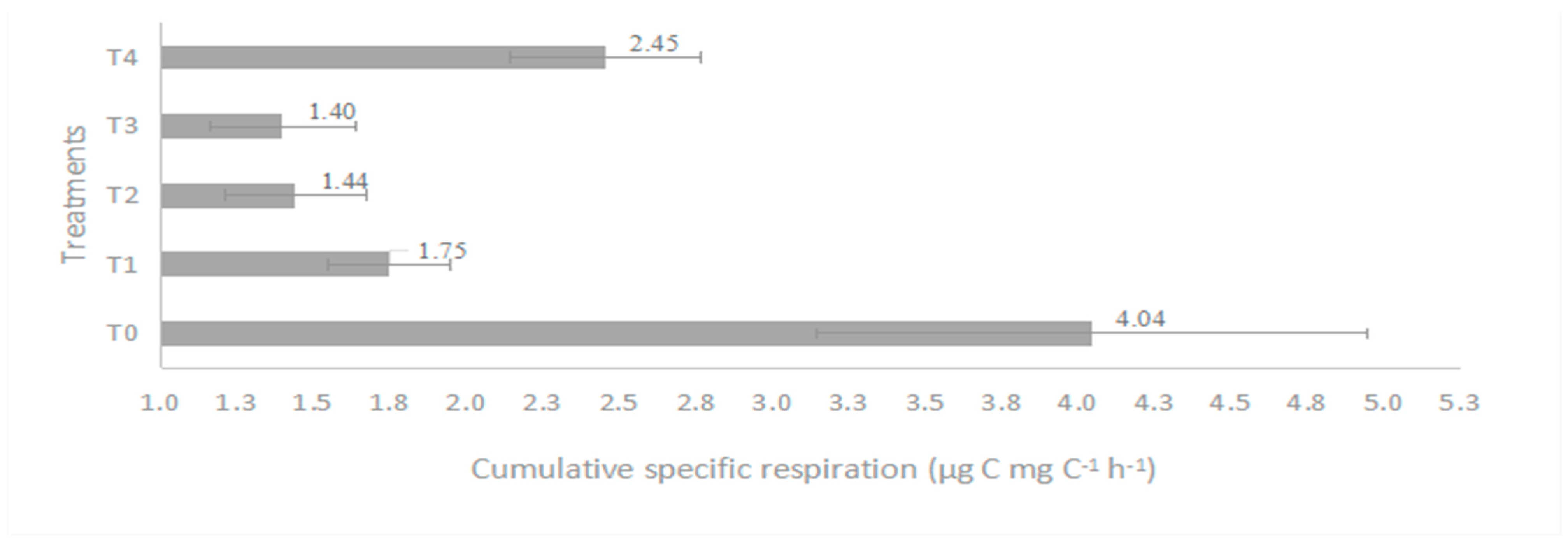
4.2. Microbial Respiration
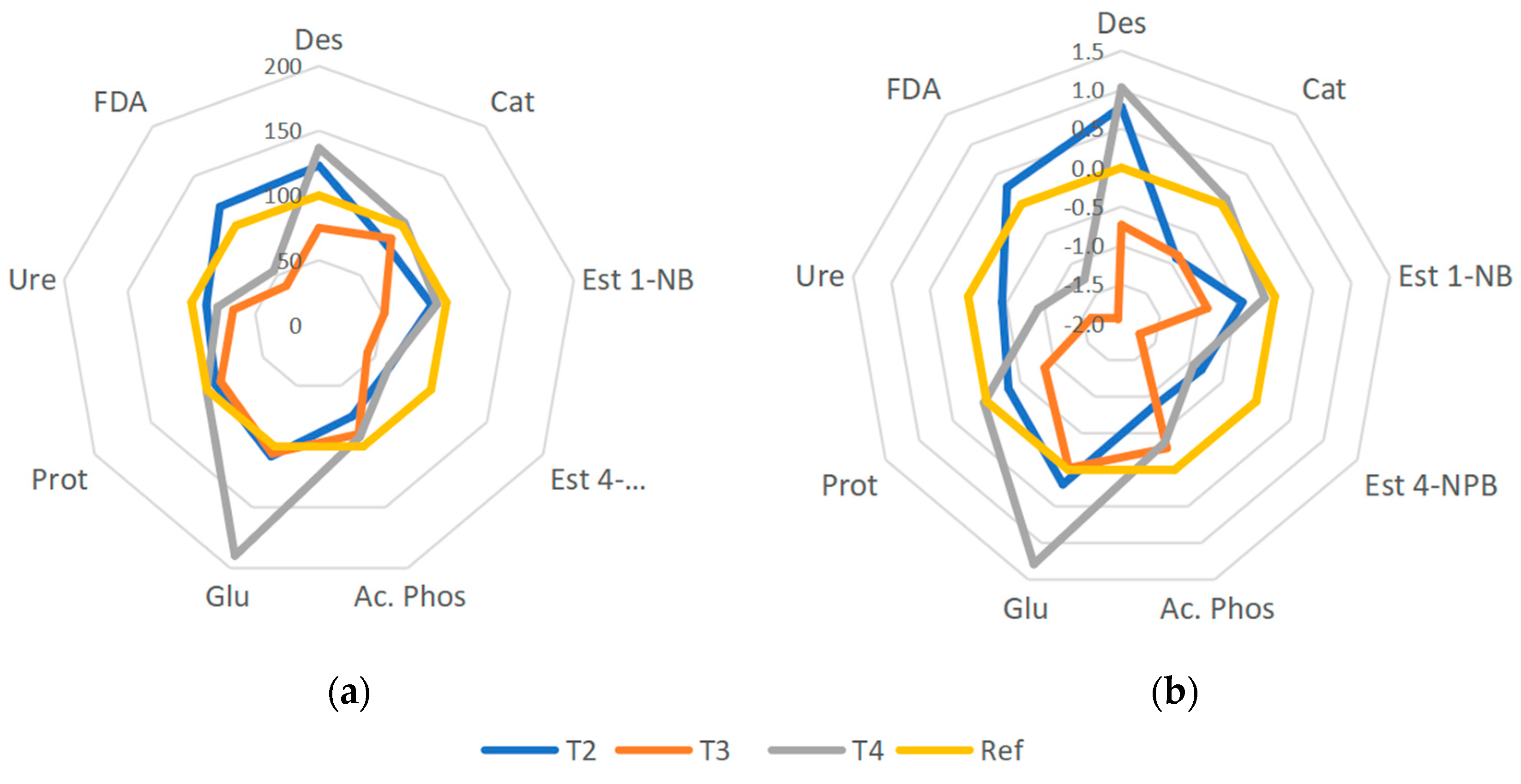
4.3. Enzymatic Activity and Global Index
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kolbe, A.; Aira, M.; Gómez-Brandón, M.; Pérez-Losada, M.; Domínguez, J. Bacterial succession and functional diversity during vermicomposting of the white grape marc Vitis vinifera v. albariño. Sci. Rep. 2019, 9, 7472. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Aira, M.; Kolbe, A.; Andrade, N.; Pérez-Losada, M.; Domínguez, J. Rapid bacterial community changes during vermicomposting of grape marc derived from red winemaking. Microorganisms 2019, 7, 473. [Google Scholar] [CrossRef]
- Domínguez, J.; Sánchez-Hernández, J.C.; Lores, M. Vermicomposting of wine-making products. In Handbook of Grape Processing By-Products: Sustainable Solutions; Galanakis, C.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 55–78. [Google Scholar]
- Nogales, R.; Fernández-Gómez, M.; Delgado-Moreno, L.; Castillo-Díaz, J.; Romero, E. Eco-friendly vermitechnological winery waste management: A pilot-scale study. SN Appl. Sci. 2020, 2, 4. [Google Scholar] [CrossRef]
- Rosado, D.; Ramos-Tapia, I.; Crandall, K.; Pérez-Losada, M.; Domínguez, J. Grapevine treatment with bagasse vermicompost changes the microbiome of albariño must and wine and improves wine quality. OENO One 2022, 56, 209–220. [Google Scholar] [CrossRef]
- Przemieniecki, S.; Zapałowska, A.; Skwiercz, A.; Damszel, M.; Telesiński, A.; Sierota, Z.; Gorczyca, A. An evaluation of selected chemical, biochemical, and biological parameters of soil enriched with vermicompost. Environ. Sci. Pollut. Res. 2020, 28, 8117–8127. [Google Scholar] [CrossRef]
- Antunes, M.; Esteves, V.; Guégan, R.; Crespo, J.; Fernandes, A.; Giovanela, M. Removal of diclofenac sodium from aqueous solution by isabel grape bagasse. J. Chem. Eng. 2012, 192, 114–121. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Lores, M.; Domínguez, J. Recycling and valorization of distilled grape marc through vermicomposting: A pilot-scale study. J. Mater. Cycles Waste Manag. 2023, 25, 1509–1518. [Google Scholar] [CrossRef]
- Jeffery, S.; Bezemer, T.M.; Cornelissen, G.; Kuyper, T.W.; Lehmann, J.; Mommer, L.; Sohi, S.P.; van de Voorde, T.F.J.; Wardle, D.A.; van Groenigen, J.W. The way forward in biochar research: Targeting trade-offs between the potential wins. GCB Bioenergy 2015, 7, 1–13. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management Science, Technology and Implementation, 2nd ed.; Routledge: London, UK, 2015. [Google Scholar]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.P.; Smith, P. Climate-smart soils. Nature 2016, 532, 49–57. [Google Scholar] [CrossRef]
- Calvelo Pereira, R.; Kaal, J.; Camps-Arbestain, M.; Pardo Lorenzo, R.; Aitkenhead, W.; Hedley, M.; Macías, F.; Hindmarsh, J.; Maciá-Agulló, J.A. Contribution to characterisation of biochar to estimate the labile fraction of carbon. Org. Geochem. 2011, 42, 1331–1342. [Google Scholar] [CrossRef]
- Garbuz, S.; Camps-Arbestain, M.; Mackay, A.; DeVantier, B.; Minor, M. The interactions between biochar and earthworms, and their influence on soil properties and clover growth: A 6-month mesocosm experiment. Appl. Soil Ecol. 2020, 147, 103402. [Google Scholar] [CrossRef]
- Gong, X.; Cai, L.; Li, S.; Chang, S.X.; Sun, X.; An, Z. Bamboo biochar amendment improves the growth and reproduction of Eisenia fetida and the quality of green waste vermicompost. Ecotoxicol. Environ. Saf. 2018, 156, 197–204. [Google Scholar] [CrossRef]
- Ameen, F.; Al-Homaidan, A.A. Improving the efficiency of vermicomposting of polluted organic food wastes by adding biochar and mangrove fungi. Chemosphere 2022, 286, 131945. [Google Scholar] [CrossRef]
- Honvault, N.; Houben, D.; Lebrun, M.; Vedere, C.; Nobile, C.; Guidet, J.; Kervroëdan, L.; Aubertin, M.L.; Rumpel, C.; Faucon, M.P.; et al. Positive or neutral effects of biochar-compost mixtures on earthworm communities in a temperate cropping system. Appl. Soil Ecol. 2023, 182, 104684. [Google Scholar] [CrossRef]
- Liesch, A.M.; Weyers, S.L.; Gaskin, J.W.; Das, K.C. Impact of two different biochars on earthworm growth and survival. Ann. Environ. Sci. 2010, 4, 1–9. Available online: https://openjournals.neu.edu/aes/journal/article/view/v4art1 (accessed on 1 January 2020).
- Bamminger, C.; Zaiser, N.; Zinsser, P.; Lamers, M.; Kammann, C.; Marhan, S. Effects of biochar, earthworms, and litter addition on soil microbial activity and abundance in a temperate agricultural soil. Biol. Fertil. Soils 2014, 50, 1189–1200. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Liang, C.; Fu, S.; Mendez, A.; Gasco, G. The effect of biochar and its interaction with the earthworm pontoscolex corethrurus on soil microbial Community structure in tropical soils. PLoS ONE 2015, 10, e0124891. [Google Scholar] [CrossRef]
- Domene, X. Chapter 11—A Critical Analysis of Meso- and Macrofauna Effects Following Biochar Supplementation. In Biochar Application Essential Soil Microbial Ecology; Elsevier: Amsterdam, The Netherlands, 2016; pp. 268–292. [Google Scholar] [CrossRef]
- Malińska, K.; Zabochnicka-Świątek, M.; Cáceres, R.; Marfà, O. The effect of precomposted sewage sludge mixture amended with biochar on the growth and reproduction of Eisenia fetida during laboratory vermicomposting. Ecol. Eng. 2016, 90, 35–41. [Google Scholar] [CrossRef]
- Paul, S.; Kauser, H.; Jain, M.; Khwairakpam, M.; Kalamdhad, A. Biogenic stabilization and heavy metal immobilization during vermicomposting of vegetable waste with biochar amendment. J. Hazard. Mater. 2020, 390, 121366. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, J.; Liu, X.; Chang, T.; Wang, Q.; Shaghaleh, H.; Hamoud, Y. Effects of biochar and vermicompost on microorganisms and enzymatic activities in greenhouse soil. Front. Environ. Sci. 2023, 10, 1060277. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.; Liu, S.; Li, Z.; Tan, X.; Huang, X.; Zheng, B. Biochar to improve soil fertility. A review. Agron. Sustain. Dev. 2016, 36, 2. [Google Scholar] [CrossRef]
- Ginebra, M.; Muñoz, C.; Calvelo Pereira, R.; Doussoulin, M.; Zagal, E. Biochar impacts on soil chemical properties, greenhouse gas emissions and forage productivity: A field experiment. Sci. Total Environ. 2022, 806, 150465. [Google Scholar] [CrossRef]
- Garg, V.K.; Gupta, R. Effect of Temperature variations on vermicomposting of household solid waste and fecundity of Eisenia fetida. Bioremediat J. 2011, 15, 165–172. [Google Scholar] [CrossRef]
- Sadzawka, A.; Carrasco, M.A.; Grez, R.; Mora, M.d.L.; Flores, H.; Neaman, A. Métodos de Análisis Recomendados para los Suelos de Chile: Revisión 2006; Serie Actas INIA No34; INIA: Santiago, Chile, 2006; 142p. [Google Scholar]
- Turner, B.L. Variation in pH optima of hydrolytic enzyme activities in tropical rain forest soils. Appl. Environ. Microbiol. 2010, 76, 6485–6493. [Google Scholar] [CrossRef]
- Von Mersi, W.; Schinner, F. An improved and accurate method for determining the dehydrogenase activity of soils with iodonitrotetrazolium chloride. Biol. Fert. Soils 1991, 11, 216–220. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Camiña, F.; Leirós, C.M.; Gil-Sotres, F. An improved method to measure catalase activity in soils. Soil Biol. Biochem. 1999, 31, 483–485. [Google Scholar] [CrossRef]
- Sánchez-Hernandez, J.C.; del Pino, J.N.; Domínguez, J. Earthworm-induced carboxylesterase activity in soil: Assessing the potential for detoxification and monitoring organophosphorus pesticides. Ecotoxicol. Environ. Saf. 2015, 122, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Popova, I.E.; Deng, S. A high-throughput microplate assay for simultaneous colorimetric quantification of multiple enzyme activities in soil. Appl. Soil Ecol. 2010, 45, 315–318. [Google Scholar] [CrossRef]
- Schinner, F.; Kandeler, E.; Ohlinger, R.; Margesin, R. Methods in Soil Biology; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Cheng, C.H.; Lehmann, J.; Thies, J.E.; Burton, S.D.; Engelhard, M.H. Oxidation of black carbon by biotic and abiotic processes. Org. Geochem. 2006, 37, 1477–1488. [Google Scholar] [CrossRef]
- Muñoz, C.; Ginebra, M.; Zagal, E. Variation of greenhouse gases fluxes and soil properties with addition of biochar from farm-wastes in volcanic and non-volcanic soils. Sustainability 2019, 11, 1831. [Google Scholar] [CrossRef]
- Hinojosa, M.B.; García-Ruíz, R.; Viñegla, B.; Carreira, J.A. Microbiological rates and enzyme activities as indicators of functionality in soils affected by the Aznalcóllar toxic spill. Soil Biol. Biochem. 2004, 36, 1637–1644. [Google Scholar] [CrossRef]
- Mijangos, I.; Albizu, I.; Epelde, L.; Amezaga, I.; Mendarte, S.; Garbisu, C. Effects of liming on soil properties and plant performance of temperate mountainous grasslands. J. Environ. Manag. 2010, 91, 2066–2074. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, W.; Burgeot, T.; Porcher, J.M. A novel “Integrated Biomarker Response” calculation based on reference deviation concept. Environ. Sci. Pollut. Res. 2012, 20, 2721–2725. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2020; Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina. Available online: http://www.infostat.com.ar (accessed on 1 January 2020).
- Alipour, M.; Fataei, E.; Nasehi, F.; Imani, A.A. Vermicompost quality and earthworm reproduction in different organic waste substrates. Int. J. Recycl. Org. Waste Agric. 2023, 12, 325–339. [Google Scholar] [CrossRef]
- Ferraz-Ramos, R.; Santana, N.; de Andrade, N.; Scheffer Romagna, I.; Tirloni, B.; de Oliveira Silveira, A.; Domínguez, J.; Josemar Seminoti Jacques, R. Vermicomposting of cow manure: Effect of time on earthworm biomass and chemical, physical, and biological properties of vermicompost. Bioresour. Technol. 2022, 345, 126572. [Google Scholar] [CrossRef] [PubMed]
- Boruah, T.; Barman, A.; Kalita, P.; Lahkar, J.; Deka, H. Vermicomposting of citronella bagasse and paper mill sludge mixture employing Eisenia fetida. Bioresour. Technol. 2019, 294, 122147. [Google Scholar] [CrossRef]
- Edwards, C.A. Breakdown of animal, vegetable and industrial organic wastes by earthworm. Agric. Ecosyst. Environ. 1988, 24, 21–31. [Google Scholar]
- Hirzel, J.; Salazar, F. Guía de Manejo y Buenas Prácticas de Aplicación de Enmiendas Orgánicas en Agricultura; Boletín INIA. Nº325; Instituto de Investigaciones Agropecuarias: Chillán, Chile, 2016; 56p. [Google Scholar]
- Sharma, K.; Garg, V.K. Comparative analysis of vermicompost quality produced from rice straw and paper waste employing earthworm Eisenia fetida (Sav.). Bioresour. Technol. 2018, 250, 708–715. [Google Scholar] [CrossRef]
- Santana, N.A.; Jacques, R.J.S.; Antoniolli, Z.I.; Martínez-Cordeiro, H.; Domínguez, J. Changes in the chemical and biological characteristics of grape marc vermicompost during a two-year production period. Appl. Soil Ecol. 2020, 154, 103587. [Google Scholar] [CrossRef]
- Wang, T.; Camps-Arbestain, M.; Hedley, M.; Bishop, P. Predicting phosphorus bioavailability from high-ash biochars. Plant Soil 2012, 357, 173–187. [Google Scholar] [CrossRef]
- Wang, T.; Camps Arbestain, M.; Hedley, M.; Bishop, P. Chemical and bioassay characterisation of nitrogen availability in biochar produced from dairy manure and biosolids. Org. Geochem. 2012, 51, 45–54. [Google Scholar] [CrossRef]
- Wang, T.; Camps-Arbestain, M.; Hedley, M. The fate of phosphorus of ash-rich biochars in a soil-plant system. Plant Soil 2014, 375, 61–74. [Google Scholar] [CrossRef]
- Muñoz, C.; Ginebra, M.; Zagal, E. Impact of Pyrolyzed and Unpyrolyzed Animal Manures on Soil Properties, Carbon Sequestration, and Clover Productivity in Andisol. Agronomy 2024, 14, 592. [Google Scholar] [CrossRef]
- Grutzmacher, P.; Puga, A.P.; Bibar, M.P.S.; Coscione, A.R.; Packer, A.P.; de Andrade, C.A. Carbon stability and mitigation of fertilizer induced N2O emissions in soil amended with biochar. Sci. Total Environ. 2018, 625, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Gascó, G.; Paz-Ferreiro, J.; Cely, P.; Plaza, C.; Méndez, A. Influence of pig manure and its biochar on soil CO2 emissions and soil enzymes. Ecol. Eng. 2016, 95, 19–24. [Google Scholar] [CrossRef]
- Ginebra, M.; Muñoz, C.; Zagal, E. Carbon stability and soil N2O emissions. Pyrolyzed or unpyrolyzed manure? J. Environ. Manag. 2022, 322, 116095. [Google Scholar] [CrossRef]
- Brassard, P.; Godbout, S.; Palacios, J.H.; Jeanne, T.; Hogue, R.; Dube, P.; Limousy, L.; Raghavan, V. Effect of six engineered biochars on GHG emissions from two agricultural soils: A short-term incubation study. Geoderma 2018, 327, 73–84. [Google Scholar] [CrossRef]
- Sheng, Y.; Zhu, L. Biochar alters microbial community and carbon sequestration potential across different soil pH. Sci. Total Environ. 2018, 622–623, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Hernandez, J.C.; Sandoval, M.; Pierart, A. Short-term response of soil enzyme activities in a chlorpyrifos-treated mesocosm: Use of enzyme-based indexes. Ecol. Indic. 2017, 73, 525–535. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameters | Unit | 1 BC1 | 1 BC2 | 1 CM | 1 B |
|---|---|---|---|---|---|
| pH | – | 5.73 | 8.2 | 10.24 | 6.78 |
| CE (1:5) | dS m−1 | – | – | – | 2.14 |
| MO | % | – | – | – | 95.45 |
| TN | g kg−1 | 2.8 | 33.9 | 8.4 | 18.4 |
| TC | g kg−1 | 841.5 | 418.8 | 116.1 | 553.7 |
| 2 Cno_ox | g kg−1 | 760.2 | 124.4 | 71.1 | - |
| C:N | – | 300.5 | 12.4 | 13.8 | 30.09 |
| N-NO3− | mg kg−1 | 380 | 7860 | 9400 | 25.5 |
| N-NH4+ | mg kg−1 | 0.55 | 9.58 | 5.79 | 79.8 |
| Av. K | g kg−1 | 0.32 | 16.15 | 14.78 | 11.29 |
| Av. P | g kg−2 | 1.58 | 13.17 | 22.19 | 2.71 |
| Av. Mg | g kg−3 | 0.06 | 12.85 | 12.32 | 1.09 |
| Av. Ca | g kg−4 | 0.71 | 30.57 | 223.8 | 4.93 |
| CaCO3-eq | % | 0.39 | 2.45 | 12.69 | – |
| S | mg kg−1 | <d.l | 260 | 230 | – |
| Fe | mg kg−1 | – | – | – | 295 |
| Mn | mg kg−1 | – | – | – | 17 |
| Zn | mg kg−1 | – | – | – | 7 |
| Cu | mg kg−1 | – | – | – | 16 |
| B | mg kg−1 | – | – | – | 20 |
| WB Moisture | % | – | – | – | 42.4 |
| Treatments | Adult Earthworms | Yield VC (g) |
|---|---|---|
| T0: B | - | 62.35 ± 2.61 c |
| T1: B + E | 97 ± 2.65 | 237.43 ± 30.80 b |
| T2: B + E + BC1 | 85 ± 14.97 | 266.75 ± 38.69 b |
| T3: B + E + BC2 | 90 ± 5.19 | 306.38 ± 16.81 a |
| T4: B + E + CM | 94 ± 4.92 | 311.38 ± 15.37 a |
| p-value | 0.30 | 0.0001 |
| CV % | 9.2 | 10.29 |
| Parameters | Units | T0: B | T1: B + E | T2: B + E + BC1 | T3: B + E + BC2 | T4: B + E + CM |
|---|---|---|---|---|---|---|
| pH (1:5) | -- | 5.51 | 5.03 | 4.92 | 5.76 | 6.61 |
| OM | % | 90.66 | 84.99 | 85.98 | 78.37 | 65.62 |
| Total N | % | 3.56 ± 0.15 ab | 3.74 ± 0.18 a | 3.33 ± 0.06 b | 3.66 ± 0.12 a | 2.94 ± 0.28 c |
| Total C | % | 46.60 ± 0.18 a | 43.65 ± 1.91 b | 48.40 ± 3.10 a | 42.73 ± 1.23 b | 35.0 ± 1.69 c |
| C/N Ratio | -- | 13.10 ± 0.53 b | 11.69 ± 0.18 c | 14.55 ± 1.12 a | 11.69 ± 0.11 c | 11.97 ± 0.58 c |
| N-NO3 | mg kg−1 | 291.90 | 1325.6 | 153.6 | 555.80 | 734.1 |
| N-NH4 | mg kg−1 | 62.70 | 85.10 | 72.70 | 60.20 | 50.30 |
| Av. N | mg kg−1 | 354.6 | 1410.7 | 226.3 | 616 | 784.4 |
| Av. P | g kg−1 | 3.36 | 3.62 | 2.83 | 11.42 | 8.33 |
| Av. K | g kg−1 | 26.64 | 22.08 | 18.43 | 18.68 | 18.01 |
| Av. Ca | g kg−1 | 7.29 | 7.72 | 6.51 | 13.44 | 74.07 |
| Av. Mg | g kg−1 | 2.41 | 2.29 | 1.93 | 4.70 | 3.38 |
| Mn | mg kg−1 | 42 | 105 | 108 | 273 | 22 |
| Cu | mg kg−1 | 52 | 48 | 39 | 88 | 53 |
| B | mg kg−1 | 30 | 41 | 32 | 48 | 37 |
| Enzymes | T1: B + E | T2: B + E + BC1 | T3: B + E + BC2 | T4: B + E + CM | p-Value |
|---|---|---|---|---|---|
| Des (µmol INFT h−1 g−1) | 1.96 ± 0.76 | 2.41 ± 0.18 | 1.47 ± 0.34 | 2.68 ± 0.70 | 0.11 |
| Cat (mmol H2O2 h−1 g−1) | 91.13 ± 7.61 | 73.84 ± 8.51 | 79.35 ± 3.7 | 93.80 ± 15.66 | 0.15 |
| Est 1-NB (µmol h−1 g−1) | 374.55 ± 245.6 | 331.19 ± 204.3 | 192.05 ± 69.9 | 347.52 ± 224.9 | 0.41 |
| Est 4-NPB (µmol h−1 g−1) | 2298.76 ± 900.8 | 1463.2 ± 436.8 | 992.97 ± 657.1 | 1442.99 ± 576.5 | 0.13 |
| Ac. Phos (µmol h−1 g−1) | 38.37 ± 12.92 | 28.87 ± 8.73 | 34.42 ± 10 | 35.34 ± 16.84 | 0.78 |
| Glu (µmol h−1 g−1) | 8.29 ± 3.15 | 8.96 ± 3.14 | 8.75 ± 4.26 | 15.73 ± 5.04 | 0.11 |
| Prot (mg Tyr eq h−1g−1) | 13.53 ± 2.22 | 12.49 ± 1.15 | 11.85 ± 5.62 | 13.57 ± 1.28 | 0.46 |
| Ure (µg NH4+ h−1 g−1) | 213.69 ± 39.27 | 188.71 ± 21.47 | 144.60 ± 45.96 | 169.91 ± 40.16 | 0.29 |
| FDA (µg F g−1) | 13.62 ± 1.84 a | 16.22 ± 5.59 a | 5.53 ± 0.43 b | 7.40 ± 0.62 ab | 0.005 ** |
| Treatments * | T-SQI | IBRv2 | GMean |
|---|---|---|---|
| T1: B + E | - | - | 48.58 ± 10.3 a |
| T2: B + E + BC1 | 64.58 ± 10.7 ab | 2.49 ± 1.03 | 43.33 ± 2.9 a |
| T3: B + E + BC2 | 45.95 ± 14.6 b | 8.93 ± 7.36 | 32.93 ± 10.2 b |
| T4: B + E + CM | 69.04 ± 9.6 a | 1.16 ± 2.1 | 45.44 ± 5.6 a |
| p-value | 0.05 * | 0.07 | 0.04 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antileo-Mellado, S.; Muñoz, C.; Sanchez-Hernandez, J.C.; Ginebra, M.; Sandoval, M. Effect of Biochar on Vermicompost Production: Chemical, Biochemical, and Biological Properties. Agronomy 2024, 14, 615. https://doi.org/10.3390/agronomy14030615
Antileo-Mellado S, Muñoz C, Sanchez-Hernandez JC, Ginebra M, Sandoval M. Effect of Biochar on Vermicompost Production: Chemical, Biochemical, and Biological Properties. Agronomy. 2024; 14(3):615. https://doi.org/10.3390/agronomy14030615
Chicago/Turabian StyleAntileo-Mellado, Silvia, Cristina Muñoz, Juan Carlos Sanchez-Hernandez, Milagros Ginebra, and Marco Sandoval. 2024. "Effect of Biochar on Vermicompost Production: Chemical, Biochemical, and Biological Properties" Agronomy 14, no. 3: 615. https://doi.org/10.3390/agronomy14030615