1. Introduction
The pink bollworm,
Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae), is a major pest of Malvaceae plants, particularly cotton [
1,
2]. This species has a global distribution and has been found to cause very serious infestation levels in many cotton-producing geographical zones [
2,
3]. In contrast with other moth species that infest cotton, the larval development of
P. gossypiella is totally completed within the cotton boll, and the infestation is internal and cryptic, being visible from the exit hole, when it is too late to take control measures [
4,
5]. Moreover, it can rapidly build high population densities that completely destroy even 100% of the cotton production in a very short period of time. There are several reports that show that this species is expanding [
6], despite the fact that it was recently successfully eradicated from the US [
7].
Internal feeding necessitates the adoption of measures that control this species before entering the cotton boll; otherwise, any insecticidal application is likely to be ineffective. It is well established that
P. gossypiella has developed a considerable level of resistance to several insecticides with different modes of action in many geographical zones [
8]. At the same time, it has been found to be resistant to Cry1Ac- and Cry2Ab-expressing cotton [
9,
10,
11]. In this context, control of this species should be based on a careful selection of measures that are to be taken, which, apart from the insecticides, should include other measures such as the application of plant growth regulators to accelerate cotton maturation [
12], or the mechanical destruction of cotton residues in the field, to reduce the numbers of overwintering larvae [
13].
Apart from the above, monitoring is considered as the keystone in any Integrated Pest Management (IPM)-based strategy against
P. gossypiella [
14,
15]. This is usually carried out through samplings in the fruiting bodies of the plant, and, especially, the use of traps baited with the male attractant of the species, which is known commercially as gossyplure, i.e., Z,Z- and Z,E-hexadecadienyl acetate [
16,
17,
18,
19,
20,
21]. Athanassiou et al. [
22] have shown that the species can be successfully monitored by both Funnel and sticky traps, but these trapping devices provided dissimilar results, in terms of the succession of the generations. In that study, the authors have underlined that Funnel traps, with the use of an insecticide as a killing agent, were more effective than sticky traps, but only when population densities were high, while trapping location and the amount of pheromone were less important [
22]. Pheromone traps are capable of providing early detection estimates, while earlier studies have shown that trap captures can be used to predict infestation patterns in the cotton field [
23,
24], and, as a result, the concomitant insecticidal applications [
25].
Although the trapping of moths in field studies is based on just trap deployment, this is only a theoretical approach, as improper trap placement may over- or underestimate population densities of a given species, leading to false estimations and predictions [
26,
27,
28,
29]. For another major pest of cotton, the cotton bollworm,
Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), it was found that monitoring through pheromone traps was highly influenced by trap height, while this effect varied throughout the growing season [
29]. Also, trap color is a critical parameter, a phenomenon that has not been clarified in detail in the case of moth species that are active during the dark hours of the day, such as
P. gossypiella [
30,
31]. In this context, the availability of an extremely wide variety of commercially available traps and pheromones constitutes the selection of the devices that are to be used for a rather complicated procedure that requires additional attention.
Based on the above and considering that there is still inadequate information on the factors that affect the trapping of P. gossypiella, we performed a series of field tests to illustrate the influence of certain factors, such as trap type and trap height. For the experimentation, we selected the area of Thessaly, which is by far the most important cotton-producing geographical zone of the European Union (EU).
2. Materials and Methods
2.1. Experimental Sites
This field study took place in the regional unit of Larissa (Region of Thessaly, Central Greece), in three experimental fields, in Nikaia, Nees Karyes, and Koilada, throughout the 2019 growing season. The experimental site in Nikaia (altitude: 90 m; 22°81′ E and 39°34′ N) was a 6 Ha field surrounded by cotton and wheat fields, whereas the 2 Ha experimental field in Nees Karyes (altitude: 146 m; 22°28′ E and 39°30′ N) was bordered only by cotton fields. Finally, the experimental field in Koilada (altitude: 140 m; 22°18′ E and 39°34′ N) was 4 Ha and in between maize, sunflower, and wheat fields. These three locations were selected due to their large cotton-producing areas. In all sites, cotton planting occurred in mid- or end-April (Nikaia and Nees Karyes: 20 April 2019, variety Celia; Koilada: 28 April 2019, variety Elpida), followed by standard cultivation practices (e.g., drip irrigation, standard fertilization scheme, etc.). Trapping activities commenced in late June and concluded in late September 2019. Throughout this period, the recorded temperatures in the region of Larissa ranged from 15.2 to 31.0 °C in June, 17.7 to 33.1 °C in July and 17.3 to 32.6 °C in August. Rainfall amounts for June, July, and August were 25.6, 19.0, and 16.4 mm, respectively. The meteorological conditions prevailing during the trials, in terms of temperature and precipitation, were typical for summer in the region of Larissa (high temperatures and low precipitation).
2.2. Trap Types and Pheromones
Three trap types were used, i.e., white Delta traps (Trécé, Inc., Adair, IA, USA), red Delta traps (Russell IPM, Deeside, UK), and green Funnel traps (Hellafarm S.A., Stylida, Greece). The bottom of the white and red Delta traps was covered with a sticky cardboard sheet for the capture of male moths that was replaced after each evaluation, whereas transfluthrin-impregnated papers (0.4% w/w, VAPONA, Sarantis SA, Athens, Greece) were introduced inside each Funnel trap as killing agents. The pheromone lures (PecGos) used contained the gossyplure sex-attractant pheromone [(Z,Z) and (Z,E)-7,11-hexadecadienyl acetate] and were provided by Novagrica Hellas S.A. (Athina, Greece).
2.3. Experimental Design
In a first field trial, one trap from each of the three different trap types, i.e., a white Delta trap, a red Delta trap, and a green Funnel trap, was installed in each of the three experimental sites with their lowest part at 60 cm from the ground level, taking care to place them at a distance of 50 m from each other. Traps were inspected at regular intervals (4–5 days) starting from 28 June to 12 September. Traps were rotated after each inspection to mitigate the individual location effect, whereas pheromone lures were replaced every three weeks. In a second trial, three white Delta traps were installed in each of the three experimental sites at three different heights, namely, with their lowest part at 30, 60, and 90 cm above the ground, and traps were inspected as previously described.
2.4. Statistical Analysis
Data were first checked for normality and homogeneity of variances using Shapiro–Wilk and Levene’s tests, respectively. Afterwards, data were subjected to One-way Analysis of Variance (ANOVA) with adult captures as the response variable and trap type and trap height as the main effects. For the comparison of the means, the Tukey–Kramer HSD test was used at the 0.05 significance level [
32]. Additionally, to assess the synchronization between pairs of captures across the different trap types on the same evaluation date, correlation coefficient values were computed.
4. Discussion
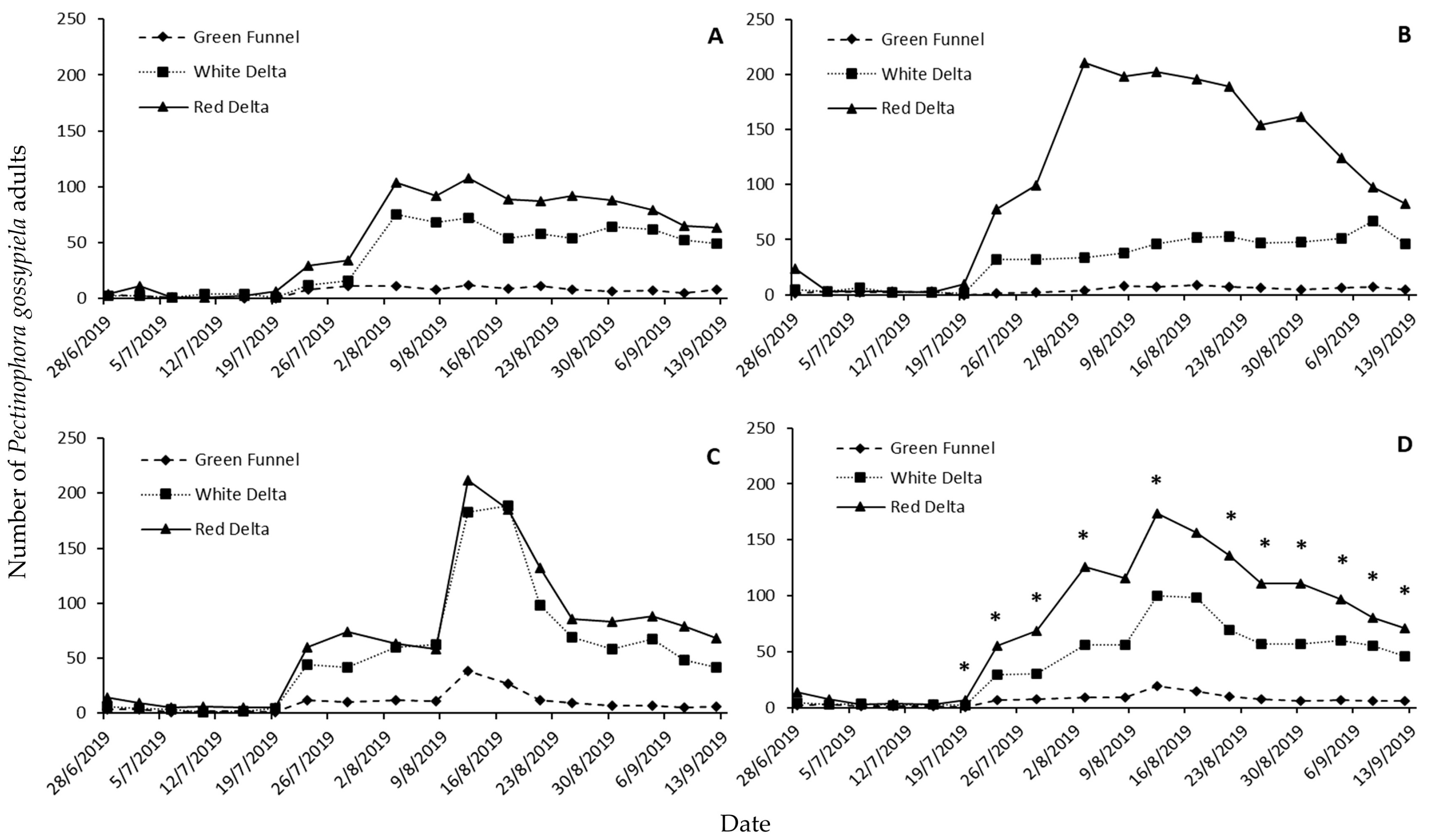
Our results show that Funnel traps had poor performance on the captures of
P. gossypiella adults, as compared with the respective figures for the other two trap types. This finding contradicts with data that had been previously reported from the same area (Thessaly) [
22], where Funnel traps were more effective than the Delta traps. However, the Funnel traps (black stripe Funnel moth trap; Agrisense BCS, Pontypridd, UK) that had been used in those tests were different, and thus, these results may not be directly comparable with the findings of the current study. However, in sticky traps, the adults are captured on a sticky surface, and in many cases, Funnel traps require the addition of a killing agent, such as a tablet that contains insecticide, which may have a repulsive action to the adults that are attracted by the pheromonic source. Earlier studies have shown that the presence of an insecticide may moderate the capture capacity of trapping devices. For instance, the addition of plugs containing dichlorvos in McPhail traps reduced the captures of adults of the Mediterranean fruit fly,
Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) [
33].
Our overall trap performance demonstrated that Delta traps provided similar results, but at certain periods of our monitoring season, red traps performed better than the white ones. Several studies have shown that trap color is a critical parameter that determines trap performance for many moth species [
30,
31,
34]. For the pine processionary moth,
Thaumetopoea pityocampa (Denis & Schiffermüller) (Lepidoptera: Thaumetopoeidae), it has been found that white- and yellow-colored traps were more effective for the capture of adults when these traps had been placed in pine trees [
35]. Similarly, Athanassiou et al. [
36] found that Funnel traps of brighter colors were superior than those of darker colors for the capture of adults of the jasmine moth,
Palpita unionalis (Hübner) (Lepidoptera: Pyralidae). However, traps performed in a dissimilar way only in some of the trap-check dates, which, in majority, were the dates when
P. gossypiella captures were high. Apart from the effect of color as a visual stimulus to the insects that approach the trap, the color might have indirectly affected captures through its effect on non-target taxa. Indicatively, it has been reported that red- or green-colored pheromone sticky traps had less effect on non-target species, especially beneficial insects, as they caught fewer bees compared to the white, yellow, or blue traps, allowing more free space in the sticky surface for the target species [
37]. Similar results have been reported by Bian et al. [
38] who suggested that fewer natural enemies were captured on color-optimized sticky card traps compared to yellow commercial ones. Although we did not collect quantitative data for the captured non-target taxa individuals, we observed that red traps had lower non-target taxa numbers as compared with the white ones, which may be one of the main reasons for their highest efficacy on
P. gossypiella. This effect may be important when
P. gossypiella adult numbers were high, as this can be depicted in surfaces that are not saturated with large-bodied “visitor” species. Along these lines, it should be highlighted that since the use of traps for insect pest monitoring may exert a negative impact on beneficial insects (natural enemies, e.g., parasitoids and predators, pollinators, etc.), special care should be taken when designing monitoring strategies to mitigate their environmental impact.
Interestingly, the traps tested here exhibited a dissimilar performance in the different areas tested. Hence, both Delta traps performed in the same way in Nikaia and Koilada, but not in Nees Karyes. While we are unaware of the reasons that caused these dissimilarities, we hypothesize that this could be attributed to the non-target taxa numbers that might be higher in the case of the white traps. Also, these differences may have been influenced by the individual trapping locations, since some locations provided higher numbers of
P. gossypiella adults than others. In fact, location is a critical parameter and should be taken into account when a trap-based monitoring protocol is planned. For instance, Karakantza et al. [
39] reported high variation in the total captures of
H. armigera adults among twenty cotton fields. The differences found among the traps that had been placed at different heights, although less pronounced as compared with those of the different trap types, were considerable during the second half of the monitoring period, when
P. gossypiella adult numbers were high. We found that traps that had been placed at 90 cm captured less adults as compared with the other two trap categories. Given that most of the fruiting bodies of the cotton plants were, for most of the growing period, at low heights, it is likely that adults were mostly active in this zone. Also, trap performance in traps that had been placed at 90 cm might have been affected by factors that have less influence at lower heights (e.g., wind, etc.). Similarly, for
H. armigera, it has been found that traps positioned at the canopy level in pigeon pea cultivation collected a greater quantity of male adults compared to traps positioned one and two feet above and below the canopy level [
40]. However, this is not always the case, given that in another study, it was found that the installation of pheromone traps at different heights in cotton fields did not significantly affect adult
H. armigera captures [
29].
Apart from the differences noted here, all traps had a similar “synchronization” of captures, in terms of the population fluctuation during the entire experimental period. This means that all treatments tested provided similar information on the succession of generations of
P. gossypiella throughout the monitoring period, and thus, all traps can be used with success to estimate the time of increased populations of this species. Nevertheless, trap performance was different in terms of early detection and trap sensitivity, i.e., the ability of certain traps to detect adults earlier than others. The detection sensitivity as categorized here underlines noticeable differences that also partially explain differential trap performance. For instance, a high number of the Funnel traps captured no
P. gossypiella adults, which is indicative of their poorer performance for this species. Moreover, the fact that the red traps were the only ones that captured >100 adults underlines the ability of these traps to capture higher moth numbers when
P. gossypiella population densities are high. High capture capacity and detection sensitivity may not be always combined in the same trap design, but there are previous studies that show that the detection sensitivity is a desirable characteristic, as they can time more accurately the decision for the application of control measures [
22,
29].
In summary, our tests underline the need for the adoption of a standardized trapping protocol, especially in area-wide management strategies against P. gossypiella in cotton. We demonstrated that specific traps are more effective than others, while trap height is an important parameter at certain periods of the growing season. Hence, unless these parameters are taken into account, the results of trapping may be inaccurate and lead to over- or underestimation of the population densities of this species. Given that larvae of this species are “internal feeders”, the early detection of adult activity is essential for the application of insecticides before the increase in the infestation level that is likely to constitute any measures considered ineffective.







{kind=link}
{kind=link}